







摘要:
植物高度是影响作物产量的关键农艺性状。芝麻植株的高度对产量表现、抗倒伏能力和植物结构都很重要。尽管芝麻品种之间的植株高度存在显著差异,但植株高度的遗传基础仍然大部分未知。在本研究中,为了探索芝麻植株高度发展的遗传见解,我们使用BGI MGIseq2000测序平台,对两种植株高度不同的芝麻品种(中芝13和ZZM2748)的茎尖在五个时间点进行了全面的转录组分析。在五个时间点中,共有16,952个基因在中芝13和ZZM2748之间表现出差异表达。KEGG和MapMan富集分析以及植物激素的定量分析表明,激素生物合成和信号传导途径与芝麻植株高度发展相关。鉴定了大量参与油菜素内酯(BR)、细胞分裂素(CK)和赤霉素(GA)生物合成和信号传导的候选基因,这些激素在两个品种之间存在主要差异,表明它们在植株高度调控中的关键作用。WGCNA分析揭示了一个与植株高度性状显著正相关的模块,并发现SiSCL9是网络中涉及植株高度发展的枢纽基因。进一步在转基因拟南芥中过表达验证了SiSCL9在增加植株高度26.86%的功能。总体而言,这些结果增进了我们对控制植株高度发展的调控网络的理解,并为改善芝麻植物结构提供了宝贵的遗传资源。
1. 引言
芝麻(Sesamum indicum L.,2n = 26)是一种传统的油料作物,在热带和亚热带地区有着悠久的栽培历史(Bedigian和Harlan,1986年)。目前,芝麻的主要生产国主要是依赖人工种植和收获的发展中国家(Andargie等人,2021年)。芝麻种子是人类健康的众多益处的优秀来源,包括油脂、蛋白质、脂肪酸、维生素和矿物质(Pathak等人,2014年)。同时,芝麻油因其含有的生育酚、生育三烯酚和木酚素而具有抗氧化活性,这些成分在人体内具有降低胆固醇和预防高血压的效果(Senila等人,2020年)。随着芝麻在食品、医药、化妆品和生物柴油等多种行业的广泛应用,芝麻的前景和重要性日益明显(Anilakumar等人,2010年)。
植物高度是决定作物植物结构的关键农艺性状(Li等人,2019年)。随着对耕地压力的增加,未来培育品种的理想植物结构将继续是主要的育种目标(Li等人,2019年)。通过在水稻和小麦中引入“绿色革命”基因,这些基因要么参与赤霉素(GA)的生物合成,要么参与信号传导,从而开发出了半矮秆水稻和小麦品种,显著提高了谷物生产力(Peng等人,1999年;Sasaki等人,2002年)。先前的研究表明,矮小或半矮小表型主要与内源植物激素的生物合成和信号转导有关,如赤霉素(GAs)、细胞分裂素(CK)、油菜素内酯(BR)和生长素(Wang等人,2017年;Shi等人,2020年;Nakazawa等人,2001年;Castorina和Consonni,2020年;Yu等人,2019年)。GA是影响植物高度的关键因素之一,因为小麦半矮秆基因Rh1和Rh2参与GA信号传导途径(Pearce等人,2011年),而水稻半矮秆基因Sd1涉及GA生物合成(Sasaki等人,2002年)。近几十年来,通过研究GA缺乏和过量突变体,揭示了植物激素GA在决定植物高度方面的作用。研究令人信服地证明,油菜素内酯(BR),一组植物特有的类固醇激素,是促进器官伸长和最终植物高度的主要内源激素(Singh和Savaldi-Goldstein,2015年)。最近,OsBSK1-1被鉴定为水稻中BR信号的正向调节因子。OsBSK1-1的敲除突变体对BR的敏感性较低,并表现出多效性表型,包括植株高度降低、分蘖数量减少和谷粒长度缩短(Tian等人,2023年)。生长素也参与调节植物高度,通过将更多的生长素从顶端运输到根尖,ZmPIN1a的过表达大幅降低了植株高度、节间长度和穗高(Li等人,2018年)。细胞分裂素是一类在多种生物过程中发挥重要作用的植物激素,包括细胞分裂和分化、地上部/地下部平衡、营养物质重新分配、种子数量以及对逆境的响应(Sakakibara,2006年)。CK在植物中以游离碱形式存在,包括N6-异戊烯腺嘌呤(IP)、反式玉米素(tZ)、顺式玉米素(cZ)和二氢玉米素(DHZ),或以结合形式存在,但其比例因植物种类而异。OsPUP1的过表达导致生长缺陷,包括叶片长度、植株高度、谷粒重量、穗长和谷粒数量减少,这可能是由于PUPs在CK运输中的潜在作用。CK在茎中的显著减少可能解释了矮小表型(Xiao等人,2020年)。
普遍认为,植株较高的芝麻品种更容易受到强风极端天气的影响,导致倒伏损害,这给机械化收获带来很大困难(Kafiriti和Mponda,2010年;Wang等人,2014年)。栽培芝麻的高质量参考基因组序列的完成为基因探索奠定了信息基础(Wang等人,2014年)。鉴于改善芝麻植株类型的重要性,已经检测到许多与芝麻植株高度相关的QTL位点、SNPs和候选基因。Wei等人(2015年)通过使用705份芝麻材料的GWAS揭示了14个与植株高度相关的显著位点,并鉴定了两个候选基因SiDFL1和SiILR1。应用高密度芝麻二倍体图谱发现了41个植株高度相关的QTLs,包括两个主要的QTLs,qPH-8.2和qPH-3.3(Wang等人,2016a)。然而,在芝麻突变体中对矮秆基因的鉴定研究有限。唯一报道的芝麻矮秆突变体DW607表现出节间长度和茎长的显著减少,SiDWF1编码的GID1B类蛋白参与赤霉素生物合成途径(Miao等人,2020年)。尽管如此,直到现在还没有关于阐明影响芝麻植株高度发展机制的报告。
为了全面概述芝麻植株高度的发育情况,我们进行了RNA测序,比较了两种植株高度差异显著的芝麻品种的基因表达模式。通过靶向代谢组分析直接定量了各种植物激素的水平。本研究旨在通过整合转录组、靶向代谢组和WGCNA分析,研究不同发育阶段的芝麻中转录组动态,并鉴定相关调控基因。这些结果增进了我们对控制植株高度发育的调控网络的理解,并为克服倒伏和机械化收获在芝麻中的需求提供了宝贵的遗传资源。
2. 材料与方法
2.1. 植物材料和组织准备
本研究选用了两种植株高度表型对比明显的芝麻品种:中芝13(高植株高度)和ZZM2748(低植株高度),并于2017年在中国武汉进行种植。芝麻茎尖在出现第六对真叶后,于五个时间点(0天、8天、16天、24天和34天)收集,采用三次生物重复。
2.2. RNA分离和RNA-seq分析
为了剖析中芝13和ZZM2748之间在转录组水平上植株高度差异的可能分子机制,我们对来自五个发育阶段的新鲜芝麻茎尖进行了转录组测序。RNA-seq分析使用了三次生物重复。使用Trizol试剂提取总RNA样本。在BGI MGIseq2000平台上准备了RNA-Seq文库并进行了测序。使用HTSeq 6.0软件(Anders和Huber,2010年)将基因表达水平标准化为每百万映射读数的千碱基转录本数**(RPKM)。通过DESeq2软件(http://bioconductor.org/packages/release/bioc/html/DESeq.html)获得了FDR < 0.01和|log2FC| > 1**的阈值的差异表达基因(DEGs)。通过Fisher检验分析了基因本体论(GO,http://www.geneontology.org)和京都基因与基因组百科全书(KEGG,http://www.genome.jp/kegg/)富集途径,调整后的p值< 0.05。
2.3. 植物激素含量的测定
通过液相色谱-串联质谱(LC-MS/MS)方法,测定了四种植物激素(生长素、细胞分裂素、赤霉素和油菜素内酯)在中芝13和ZZM2748两个品种的芝麻茎尖中的浓度,共五个时间点。激素提取和LC-MS/MS操作如下:首先用磷酸盐缓冲液(PBS,pH7)清洗茎尖,然后在液氮中冷冻并研磨成粉末;在加入同位素内标和800 μl甲醇:水(4:1,体积比)后,通过垂直振荡30秒,超声20分钟,20 ◦C下孵育1小时,4 ◦C下离心20分钟(1300 rpm);之后,取上清液进行固相萃取。化学分析采用超高效液相色谱法(UPLC,安捷伦1290;安捷伦科技,瓦尔德布隆,德国),在多反应监测(MRM)模式下由AB 5500Q-TRAP质谱仪(AB Sciex,福斯特城,加利福尼亚州,美国)收集数据。使用MultiQuant软件进行数据分析,基于标准曲线计算激素的倍数变化。
2.4. 通过qRT-PCR验证差异表达基因(DEGs)
实时定量PCR使用SYBR® Select Master Mix(2X)(Vazyme Biotec,南京,中国)在Light Cycler 480 II(Roche,巴塞尔,瑞士)上进行。为鉴定的候选基因设计了特异性引物,由Beacon Designer 8.0设计并在表S2中列出。芝麻中的组蛋白H3.3基因(SiH3.3)和AtUBQ10被用作内参基因,以标准化转录水平。每个基因的实时检测在相同条件下使用三个独立的生物重复进行。利用2 ΔΔCt方法(Livak和Schmittgen,2001)根据循环阈值计算基因表达。
2.5. 加权相关网络分析
为了检测芝麻植株高度背后的枢纽基因,我们使用RNA-seq获得的表达基因(FPKM)值进行了加权相关网络分析(WGCNA)。WGCNA在R语言(版本4.0.2)中进行,使用R包WGCNA来生成加权相关网络(Langfelder和Horvath,2008)。默认的幂值为26。使用Cytoscape软件(Shannon等人,2003)绘制了黑色模块的调控网络。
3. 结果
3.1. 对比植株高度差异的两种芝麻品种的表型测定
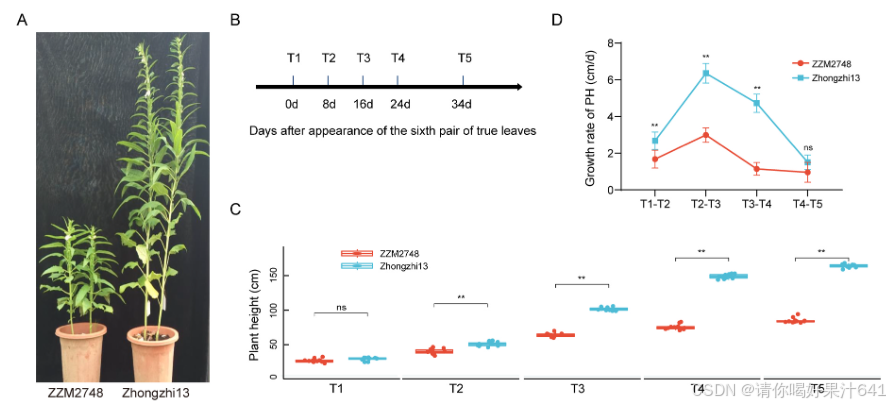
两种芝麻品种,中芝13和ZZM2748,被用于比较芝麻发育期间的植株高度(图1)。在五个时间点(T1、T2、T3、T4和T5)测量植株高度(PH),并从T1(中芝13为29.32厘米,ZZM2748为26.70厘米)持续增加到T5(中芝13和ZZM2748分别为163.96厘米和85.09厘米)(图1B和C)。在T1时,中芝13和ZZM2748的PH没有差异。在接下来的四个时间点,T2、T3、T4和T5,两种芝麻品种的PH之间观察到显著差异(P < 0.01)(图1C)。随着时间的推移,中芝13显示出比ZZM2748更高的生长速率,特别是在T1-T4期间(图1D)。中芝13和ZZM2748的生长速率在T2到T3阶段达到最大,表明这是植株高度发育的最重要阶段。
3.2. 植株高度发育的全局转录组分析
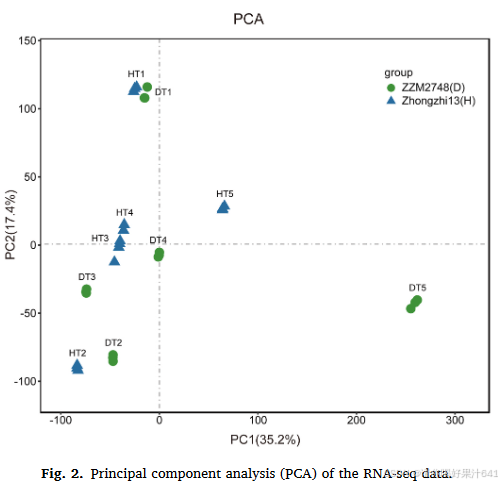
对植株高度发育的五个时间点的芝麻品种中芝13和ZZM2748进行了转录组动态分析。共使用了30个茎尖样本(2个品种×5个时间点×3个生物重复)进行转录组测序。在去除接头序列、模糊序列和低质量序列后,每个样本产生了55.6至76.8百万的清洁读段,这些读段用于进一步分析(表S1)。清洁读段中有91.16%至92.66%与中芝13参考基因组比对成功。所有样本的Q20值在97.52%至98.13%之间,Q30值在91.83%至93.91%之间,在不同的发育阶段。GC含量在45.01%至45.86%之间。在30个分析样本中共表达了28,615个基因。所有转录组数据反映了两个芝麻品种在植株高度发育期间转录组的足够覆盖度,并且数据可靠,适用于进一步分析。不同时间点的三个生物重复之间的Spearman相关系数范围在0.99至1.00之间(图S1)。为了研究中芝13和ZZM2748在植株高度发育期间转录组动态的全局差异,我们基于表达基因的FPKM值进行了主成分分析(PCA)(图2)。PCA结果显示,不同时间点之间通过PC2清晰分离。中芝13和ZZM2748的样本在T1和T2-T4期间集中分布,因此,这两个阶段的样本可能显示出类似的转录组水平。T5的样本通过PC1在不同基因型之间分离。总体而言,这些结果表明重复样本之间的相关性相对较高,且中芝13和ZZM2748在五个阶段显示出明显的时序依赖性转录组表达谱。
3.3. 两种芝麻品种之间的基因差异表达
为了进一步研究这两个芝麻品种在不同植株高度发育时间点的转录差异,我们鉴定了中芝13和ZZM2748之间的差异表达基因(DEGs)。在五个时间点共鉴定出16,952个DEGs(图3A)。具体来说,在中芝13和ZZM2748之间,五个时间点分别有1240、2518、2773、2755和7666个DEGs。此外,在T1、T3、T4和T5时间点,上调基因的数量高于下调基因的数量。我们鉴定了在五个时间点分别有792、943、1688、1706和4753个上调DEGs,以及448、1575、1085、1049和2913个下调DEGs。为了验证RNA-seq数据,我们通过实时定量逆转录聚合酶链反应(qRT-PCR)分析了八个随机选择的DEGs的基因表达变化。尽管所选基因在RNA-seq和qRT-PCR之间的倍数变化不同,但变化趋势是一致的(图S2)。
基于KEGG通路的差异表达基因(DEGs)分析揭示了几个在不同时间点独特或共同富集的生物过程(图3B)。具体来说,DEGs主要与以下16个主要类别相关:代谢途径、次生代谢物的生物合成、脂肪酸代谢、氨基酸的生物合成、碳代谢、氨基酸代谢产物、其他次生代谢物的生物合成、能量代谢、环境适应、糖苷生物合成与代谢、脂质代谢、辅酶和维生素代谢、其他氨基酸代谢、萜类和聚酮类代谢、复制和修复以及信号转导。有趣的是,KEGG通路富集分析表明,二萜生物合成途径(ko00904)、与赤霉素(GA)相关的萜类骨架生物合成(ko00900)以及与细胞分裂素(CK)生物合成相关的玉米素生物合成(ko00908)在不同时间点富集。植物激素信号转导途径(ko04075)在T2-T4时间点富集,此时两个品种的植株高度差异明显。这些结果暗示这三条途径可能有助于植株高度的发育。
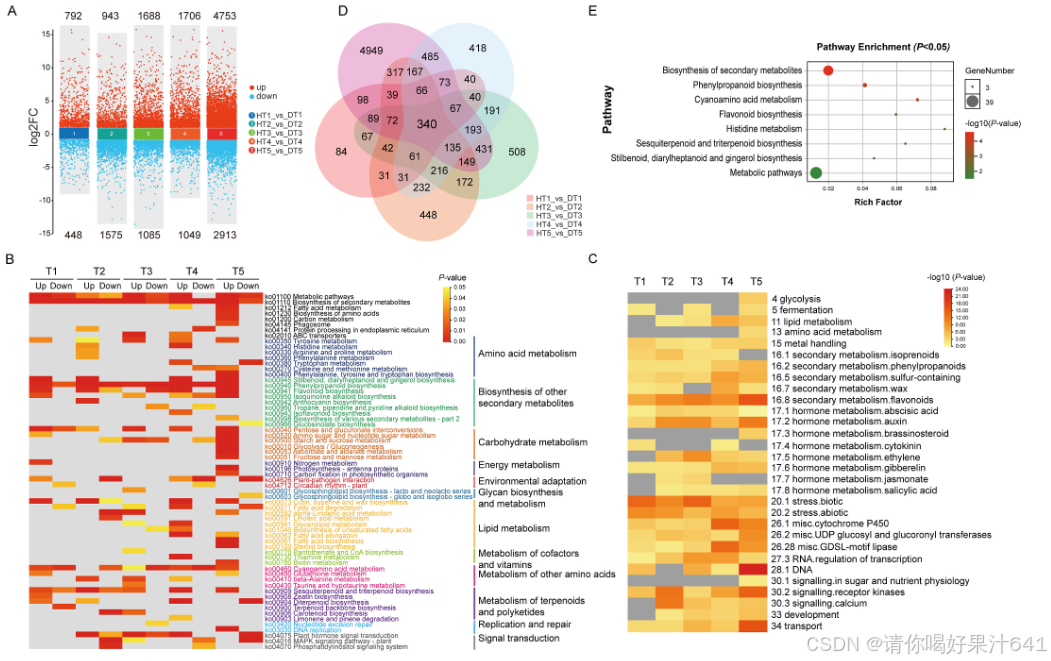
两个品种之间DEGs的富集MapMan功能类别在图3C中显示。这些DEGs富集到了细胞壁、多种次生代谢、脂质、氨基酸和激素代谢、蛋白质合成与降解、应激响应和信号传导等目录中。大多数富集的术语在多个时间点上都有过度表达。值得注意的是,DEGs的功能注释表明,在植株高度发育过程中,与多种激素代谢相关的基因,包括脱落酸、生长素、油菜素内酯、细胞分裂素、乙烯、赤霉素、茉莉酸和水杨酸等,在两个品种间都有过度表达。进一步比较了中芝13和ZZM2748在五个时间点之间的重叠DEGs,以鉴定涉及芝麻植株高度发育的核心基因。如图3D所示,在中芝13和ZZM2748之间共有340个基因重叠。对这些340个DEGs进行的进一步KEGG富集分析表明,影响次生代谢物(例如,酚丙烷、黄酮类和倍半萜及三萜)的基因有过度表达(图3E)。
3.3逻辑
-
差异表达基因(DEGs)鉴定:
- 目的:识别在不同植株高度发育阶段中芝13和ZZM2748两个芝麻品种之间的转录差异。
- 方法:通过RNA-seq技术在五个不同的时间点对两个品种的样本进行转录组测序,鉴定出差异表达的基因。
-
DEGs数量和变化趋势分析:
- 目的:量化每个时间点的DEGs数量,并比较上调和下调基因的数量。
- 方法:统计每个时间点的DEGs数量,并分析上调和下调基因的比例。
-
DEGs功能分类和富集分析:
- 目的:了解DEGs在生物学过程中的功能分类,并找出在特定生物学过程中富集的DEGs。
- 方法:利用KEGG和MapMan数据库对DEGs进行功能分类和富集分析,识别与植株高度发育相关的生物过程和代谢途径。
-
DEGs验证:
- 目的:验证RNA-seq结果的准确性。
- 方法:通过qRT-PCR技术对部分DEGs进行表达验证,比较RNA-seq和qRT-PCR结果的变化趋势是否一致。
-
核心基因鉴定:
- 目的:识别在植株高度发育中起关键作用的基因。
- 方法:比较两个品种在不同时间点的DEGs,找出共有的DEGs,进一步通过KEGG富集分析确定这些基因在次生代谢物合成中的作用。
-
激素代谢相关基因分析:
- 目的:探讨激素代谢在植株高度发育中的作用。
- 方法:分析DEGs的功能注释,特别关注与多种植物激素代谢相关的基因,这些基因在植株高度发育过程中可能起调控作用。
整体逻辑是通过转录组测序技术鉴定差异表达基因,然后通过功能分类和富集分析来了解这些基因在植株高度发育中的潜在作用,最后通过实验验证和核心基因鉴定来进一步确认这些基因的功能和重要性。这一过程有助于揭示控制芝麻植株高度发育的分子机制,并为后续的功能研究和育种提供理论基础和候选基因。
3.4. 通过LC-MS/MS定量分析植物激素含量以及参与植物激素生物合成和信号转导的基因表达谱
植物激素已被证实与植物高度密切相关。此外,DEGs的富集KEGG途径和MapMan注释与植物激素生物合成和信号转导相关。因此,我们对中芝13和ZZM2748在五个发育阶段的植物激素浓度变化进行了分析,这些阶段与选择的RNA-seq材料相同。通过同位素标记内标法的LC-MS/MS绝对定量分析了包括生长素、细胞分裂素、赤霉素和油菜素内酯在内的四种植物激素。共检测到12种代谢物,包括吲哚-3-乙酸(IAA)、甲基吲哚-3-乙酸(ME-IAA)、3-吲哚乙酸(IBA)、吲哚-3-甲醛(ICA)、N6-异戊烯腺嘌呤(IP)、反式玉米素(tP)、顺式玉米素(cZ)、二氢玉米素(DZ)、赤霉素A1(GA1)、赤霉素A3(GA3)、赤霉素A4(GA4)和油菜素内酯(BR)。
为了区分不同材料之间的差异,我们使用监督式偏最小二乘判别分析(PLS-DA)对数据进行了分析(图4A)。从得分图可以看出,根据品种,样本被有效地区分为两组。PLS-DA还提供了一个VIP分数,该分数描述了化合物对样本分类的贡献,更直观地显示了化合物在样本分类中的作用。共有三种植物激素的VIP分数≥1.5,分别是IP、BR和GA1(图4A)。这三种植物激素在所有阶段中芝13和ZZM2748之间显示出显著差异(P < 0.01)和一致的趋势(图4B和图S3)。
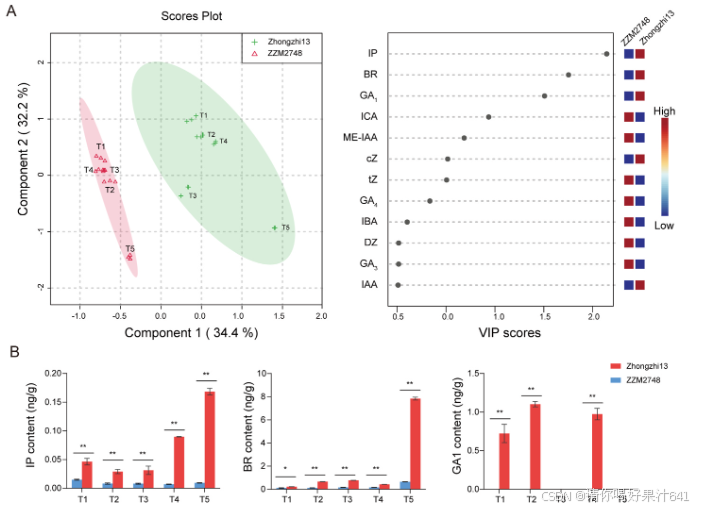
为了进一步研究植物激素相关基因在中芝13和ZZM2748植株高度发育中的参与情况,我们根据KEGG数据库和已发表的文献,概述了芝麻中假定的油菜素内酯(BR)、赤霉素(GA)和细胞分裂素(CK)生物合成和信号转导途径(图5)。具体来说,与BR、GA和CK的生物合成和信号转导相关的基因分别为23、38和37个(表S3)。这些基因在中芝13和ZZM2748之间的表达模式至少在一个时期有所变化。
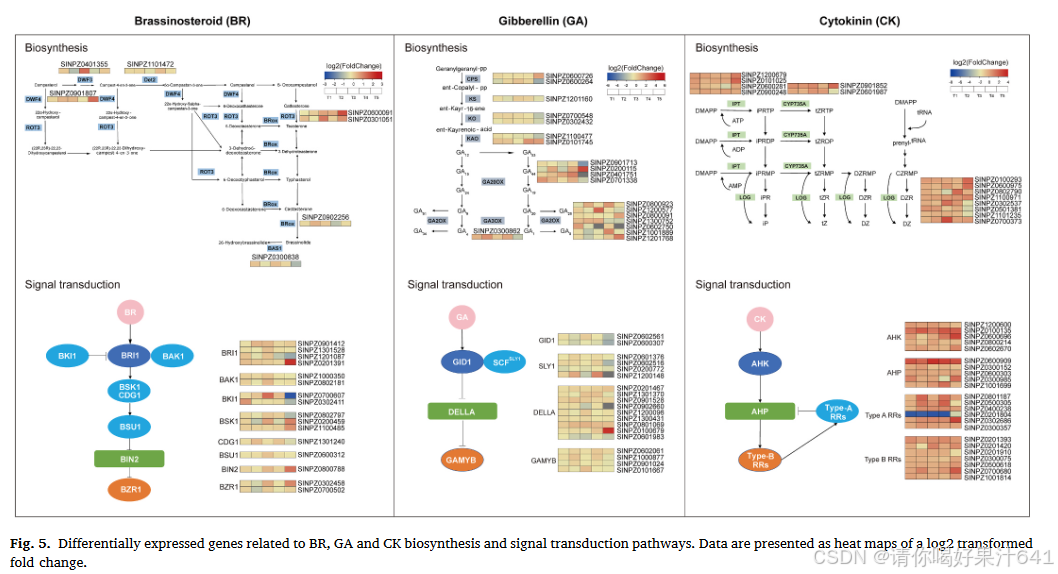
对于BR生物合成,四个基因(SINPZ0401355、SINPZ0901807、SINPZ0600091和SINPZ0301051)在T3、T4、T5时间点之一中,在中芝13中的表达量高于ZZM2748。对于BR信号转导,七个基因在T2-T5期间两个分析品种之间显著差异表达(|log2FC| > 1,FDR < 0.01)。共有四个基因(SINPZ0101745、SINPZ0200115、SINPZ0701338和SINPZ0300862)与GA生物合成相关,在植株高度发育的不同阶段在中芝13中高度表达。这些基因编码GA生物合成途径中的关键酶,包括CPS、KAO、GA20OX和GA3OX。还有两个基因(SINPZ1300752和SINPZ1200577),它们编码参与GA分解代谢的GA2OX家族成员,在T4和T5期间两个品种之间显著差异表达(|log2FC| > 1,FDR < 0.01),在ZZM2748中的表达量更高。此外,参与GA信号转导的七个基因在多个时间点在中芝13和ZZM2748之间显著差异表达(|log2FC| > 1,FDR < 0.01)。
对于CK生物合成,编码CK生物合成酶的十个基因,包括IPT、CYP735A1和LOG(SINPZ0101025、SINPZ1200679、SINPZ0900248、SINPZ0901852、SINPZ0601987、SINPZ1100971、SINPZ0302537、SINPZ0700373、SINPZ0100293和SINPZ0600975),在中芝13和ZZM2748之间显著差异表达(|log2FC| > 1,FDR < 0.01),并且在T3或T5期间这些基因在中芝13中的表达量高于ZZM2748。参与CK信号转导的十个基因的表达在中芝13和ZZM2748之间也显著不同(|log2FC| > 1,FDR < 0.01)。
因此,BR、GA和CK的生物合成和信号转导可能影响植株高度的发育。
3.4实验逻辑
-
植物激素与植株高度的关系:
- 确认植物激素与植株高度的密切关系,并以此为基础进行后续实验。
-
激素浓度分析:
- 对两个芝麻品种(中芝13和ZZM2748)在五个发育阶段的四种主要植物激素(生长素、细胞分裂素、赤霉素和油菜素内酯)浓度进行LC-MS/MS绝对定量分析,以检测激素水平的变化。
-
数据差异分析:
- 使用PLS-DA分析来区分不同品种间的激素水平差异,并识别出对样本分类贡献大的植物激素(IP、BR和GA1)。
-
基因表达与激素途径分析:
- 基于KEGG数据库和文献,概述芝麻中与BR、GA和CK生物合成和信号转导相关的基因,并分析这些基因在不同品种间的表达差异。
-
关键基因的表达模式分析:
- 分析特定基因(如BR生物合成和信号转导相关基因、GA生物合成和信号转导相关基因、CK生物合成和信号转导相关基因)在不同时间点的表达模式,以及它们在植株高度发育中的作用。
-
基因表达与激素水平的关联:
- 探讨基因表达变化与激素水平变化之间的关系,以及这些变化如何影响植株高度的发育。
-
结论:
- 得出BR、GA和CK的生物合成和信号转导可能影响植株高度发育的结论,并提出这些发现为未来研究提供了线索。
整个实验逻辑是通过对植物激素水平的定量分析和基因表达模式的比较分析,来揭示控制芝麻植株高度发育的潜在分子机制。这种方法有助于识别关键的激素和基因,为改良芝麻植株结构提供了科学依据。
3.5. 识别与植株高度高度相关的基因共表达网络模块
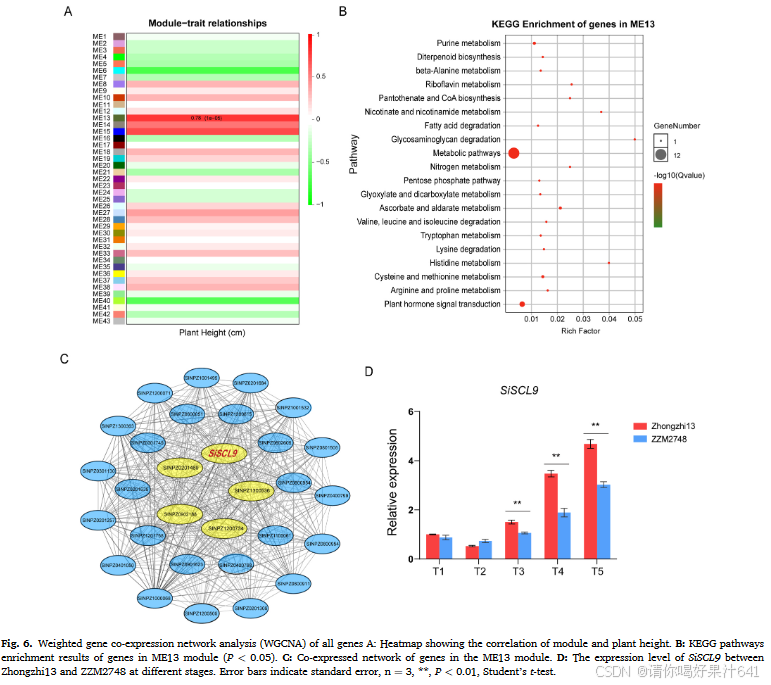
执行了加权基因共表达网络分析(WGCNA),以识别基于28,615个基因的表达谱与性状相关联的候选模块。WGCNA共鉴定出43个模块(包含23,239个基因)(图6A)。模块-性状相关性分析显示,ME13模块与植株高度性状显著正相关(r = 0.78)。图6B展示了ME13模块中1248个基因的KEGG分析。在富集度最高的前20个KEGG途径中,二萜生物合成、代谢和植物激素信号转导途径被富集,其中二萜生物合成途径与赤霉素(GA)生物合成相关。通过估算ME13模块内计算出的权重值和共表达关系,检测到与植株高度相关的五个枢纽基因:SINPZ0201469、SINPZ0902188、SINPZ1300536、SINPZ1200734和SINPZ0000728(图6C,表S4)。这些枢纽基因与其他基因高度相关(权重>0.15)。SINPZ0000728是WGCNA在ME13模块中发现的五个枢纽基因之一,编码一个属于GRAS基因家族的转录因子。由于与拟南芥中的同源基因AtSCL9(At2g37650),SINPZ0000728被命名为SiSCL9。SiSCL9的表达模式显示,在多个时间点中芝13中的表达显著更高(P < 0.01)(图6D)。考虑到在拟南芥中的同源基因注释以及在中芝13和ZZM2748中的基因表达水平,SiSCL9被选为进一步功能验证的对象。
3.5实验逻辑
这部分实验的逻辑可以概括为以下几个步骤:
-
共表达网络构建:
- 使用WGCNA方法,基于28,615个基因的表达数据,构建加权基因共表达网络,以识别与特定性状(植株高度)相关的基因模块。
-
模块鉴定与性状关联:
- 通过WGCNA分析,鉴定出43个共表达模块,这些模块包含23,239个基因,并找出与植株高度显著正相关的ME13模块(相关系数r = 0.78)。
-
功能富集分析:
- 对ME13模块中的1248个基因进行KEGG途径富集分析,以确定这些基因参与的主要生物学过程和代谢途径,特别是那些与植物激素信号转导和赤霉素生物合成相关的途径。
-
枢纽基因识别:
- 在ME13模块内,通过计算基因的权重值和共表达关系,识别出五个与植株高度高度相关的枢纽基因,这些基因与其他基因的共表达关系强(权重>0.15)。
-
候选基因选择与命名:
- 根据WGCNA分析结果,选择SINPZ0000728基因(命名为SiSCL9)作为候选基因,因为它编码一个属于GRAS家族的转录因子,并且与拟南芥中的已知基因AtSCL9同源。
-
表达模式分析:
- 分析SiSCL9在不同时间点在中芝13中的表达模式,发现其表达显著高于ZZM2748,这表明SiSCL9可能在植株高度发育中起重要作用。
-
功能验证:
- 基于SiSCL9与已知基因的同源性以及其在不同品种中的表达差异,选择SiSCL9进行进一步的功能验证,以确定其在植株高度调控中的具体作用。
整个实验逻辑是通过构建共表达网络,识别与植株高度相关的基因模块和枢纽基因,然后对这些基因进行功能富集分析和表达模式研究,最终选择候选基因进行功能验证,以揭示其在植株高度发育中的作用机制。这种方法有助于深入理解控制植株高度的分子网络,并为后续的基因功能研究和作物改良提供候选基因。
3.6. SiSCL9的过表达提高了转基因拟南芥的植株高度
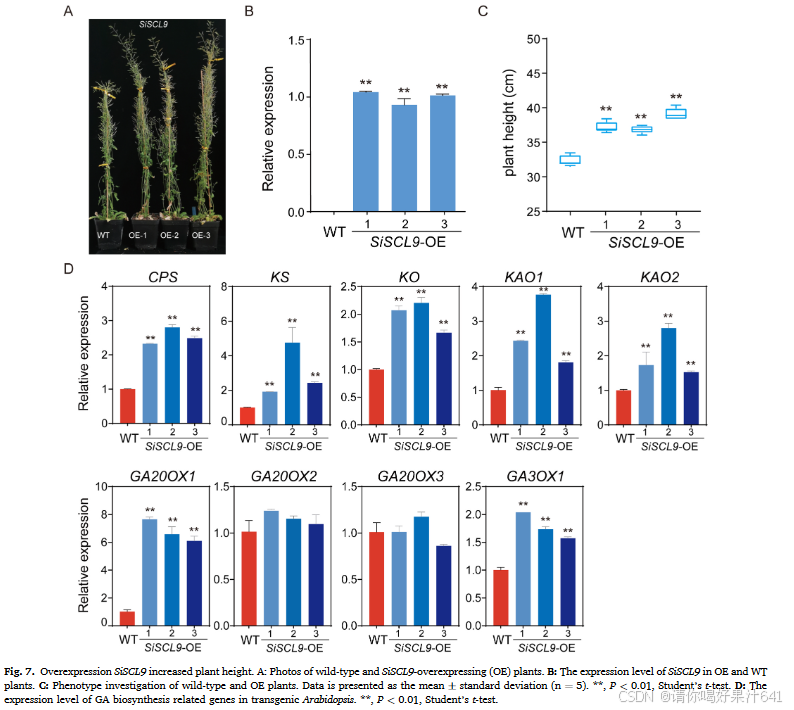
为了验证SiSCL9在植株高度中的功能,我们在35S启动子下过表达了SiSCL9转基因拟南芥。选择了三个转基因拟南芥过表达系(OE1、OE2和OE3)的种子进行进一步分析。过表达系中SiSCL9的表达水平显著高于野生型(图7B)。与野生型相比,SiSCL9的过表达显著增加了转基因拟南芥的植株高度26.86%(图7C)。为了进一步探究SiSCL9对植株高度的潜在调控机制,我们检测了转基因拟南芥中赤霉素生物合成相关基因的表达水平。包括CPS、KS、KO、KAO1、KAO2、GA20ox1和GA3ox1在内的赤霉素生物合成基因的表达在转基因拟南芥中显著上调(图7D)。
4. 讨论
芝麻,被誉为“油料作物之王”,是一种重要的特色油料作物。芝麻植株的高度对产量表现、抗倒伏能力和植物结构都很重要。芝麻的植株高度由顶部的连续生长构成的节间和节间长度组成。与其他作物相比,由于植株高度高、收获指数低以及对生物和非生物胁迫的敏感性,芝麻的产量能力较低(Uzun和Çağırgan,2006)。尽管已经鉴定了一些控制芝麻植株高度的QTL和候选基因,但决定植株高度的分子机制仍然不清楚(Wang等人,2016b;Miao等人,2020;Wei等人,2015)。因此,阐明遗传机制以探索新的矮秆或半矮秆芝麻遗传资源对于芝麻育种是必要的。
在本研究中,选择了两种植株高度固有差异的芝麻品种,中芝13和ZZM2748,为分析植株高度差异的分子机制提供了有力的基础。全局转录组分析有助于发现新基因及其表达谱,并可能为研究植株高度提供宝贵的数据。动态转录组数据通过PCA清楚地突显了三个不同的时期(T1、T2-T4和T5)。这种聚类结果揭示了不同阶段之间的基因表达变化。在T1和T2-T4,中芝13和ZZM2748紧密聚集,表明这两个品种在这些阶段的转录组水平相似,并且从一个阶段到另一个阶段基因表达发生了显著变化。此外,差异表达分析揭示,在T2-T4(2518、2773和2755)比T1(1240)中芝13和ZZM2748之间有更多的差异表达基因(DEGs)。在当前研究中,KEGG富集分析表明,在T2-T4相比于T1,更多上调的DEGs富集在植物激素信号转导途径中。通过MapMan对DEGs的功能注释表明,与激素代谢相关的术语在大多数时间点显著富集(P < 0.05)。已经证明,植物激素信号转导与植株高度密切相关,阻断植物激素代谢和信号转导途径是植物矮化的机制之一(Wolters和Jürgens,2009;Ju等人,2018)。在芝麻中,从芝麻矮化突变体中克隆出的SiDWF1编码一个涉及赤霉素信号转导途径的GID1B类蛋白(Miao等人,2020)。
在本研究中,通过LC-MS/MS对四种植物激素(生长素、细胞分裂素、赤霉素和油菜素内酯)进行了分析,包括12种代谢物。根据PLS-DA分析结果,IP、BR和GA1被鉴定为中芝13和ZZM2748之间主要的差异激素。IP是植物中细胞分裂素的游离碱形式,它在调节植物生长和发育(如细胞分裂、根伸长和芽生长、叶衰老以及其他应激反应和病原体抗性)中发挥着重要作用(Kakimoto, 2003; Li et al., 2021)。在拟南芥中,细胞分裂素信号由三个传感器组氨酸激酶AHK2、AHK3和CRE1/AHK4感知,而磷酸转移蛋白(AHPs)是AHKs和响应调节因子(ARRs)之间的双组分磷酸中继介质(Kieber和Schaller, 2014)。A型ARRs在拟南芥中负向调节细胞分裂素信号(To et al., 2007)。在本研究中,编码A型响应调节因子(A型ARRs)的SINPZ0201804,在T1-T5期间在ZZM2748中的表达量显著高于中芝13(log2FC > 1, FDR <0.01)。AHPs作为细胞分裂素信号的冗余正向调节因子,影响植物发育的多个方面(Hutchison et al., 2006)。CK受体AHK和AHP基因家族成员SINPZ0201393、SINPZ0100135和SINPZ0600909,在T1-T5期间在中芝13中的表达量显著高于ZZM2748。BR是植物生长和发育所必需的类固醇激素,它们控制整个植物生命周期中各种细胞类型的分裂、伸长和分化(Planas-Riverola et al., 2019)。因此,由于编码BR合成和信号传导途径关键元素的基因发生突变,导致严重的矮化、器官生长和发育减少、植物生育力和产量降低(Sun et al., 2021)。蛋白BRASSINOSTEROID INSENSITIVE 1(BRI1)调节,响应油菜素内酯结合,涉及植物发育的信号级联反应,包括光和应激调控基因的表达、促进细胞伸长、正常的叶和叶绿体衰老以及开花(Singh和Savaldi-Goldstein, 2015)。编码BRI1的SINPZ0201391在两个分析品种之间在T1、T2和T5期间表达差异显著,中芝13中的表达量更高。赤霉素是植物高度的一个关键内源调节因子。在本研究中,内源生物活性赤霉素酸1(GA1)是两个品种之间的主要差异激素,与中芝13相比,ZZM2748中GA1的浓度显著降低。小麦中的‘绿色革命’基因Rht-B1/Rht-D1编码一个DELLA蛋白,作为GA信号传导途径中的负向调节因子。最近的研究揭示了一个Cla015407(GA3ox)基因控制西瓜的矮化结构(Sun et al., 2020)。在这里,编码GA20OX酶的SINPZ0200115和SINPZ0701338,在三个时间点上调。GA20氧化酶(GA20OX)负责形成C19-GA骨架时去除C-20,产物是几乎所有后续活性GAs的前体。CnGA20ox1的表达在高植株中大约是矮植株的3倍(Boonkaew et al., 2018)。在玉米中,GA20ox1基因也被报道控制植株高度(Voorend et al., 2016)。在拟南芥中,NAC转录因子JUB1被证明直接抑制GA3ox1和DWF4,导致GAs和BRs水平降低,从而表现出短下胚轴和矮化的表型(Shahnejat-Bushehri et al., 2016)。在水稻中,ckx4 ckx9双突变体中GA相关基因显著下调,这可能解释了ckx4 ckx9双突变体植株高度显著降低的原因(Rong et al., 2022)。总体而言,芝麻植株高度可能通过一个复杂和高度协调的植物激素相互作用网络来调节。
WGCNA被广泛用于推断共表达网络模块。SiSCL9是WGCNA分析中与植株高度高度相关的ME13模块中权重值最高的顶级基因。此外,SiSCL9在转基因拟南芥中的过表达导致植株高度增加了26.86%。在不同物种中,同源基因可能具有保守的功能,但嵌入在不同的转录调控网络中。GRAS家族蛋白已在近300种植物中报道,通常似乎通过转录调控影响植物生长、发育和对环境的响应(Bolle, 2004; Mayrose et al., 2006)。SiSCL9是AtSCL9的同源基因,属于GRAS家族的SCARECROW-LIKE(SCL)亚家族。DELLA蛋白属于植物特有的GRAS家族的一个亚家族,拟南芥包含五种DELLA蛋白(RGA、GAI、RGL1、RGL2和RGL3),它们在抑制GA响应中发挥作用(Lee et al., 2002)。SCL亚家族成员编码的GRAS蛋白没有GA响应的DELLA结构域。研究发现SCARECROW-LIKE 3(SCL3)参与GA途径并促进根内皮层(Heo et al., 2011)。其他研究揭示SCL3通过直接与DELLA相互作用自我调节其转录,并根据遗传研究,RGA和SCL3在GA信号和生物合成中可能具有相反的作用(Zhang et al., 2011)。在我们的研究中,与野生型相比,拟南芥转基因系中GA生物合成基因的表达水平上调。因此,SiSCL9的过表达可能增强GA生物合成活性,增加转基因拟南芥中GA水平,并最终促进植株高度,但分子机制尚不清楚,需要进一步研究。靶向SiSCL9可能有助于改善芝麻的架构。需要对这些候选基因进行功能研究,以了解其调控中涉及的分子机制,并发挥其全部潜力。
- 结论
为了应对耕地面积日益增加的压力并适应机械化收获,理想的芝麻植株结构将继续成为未来育种的主要目标。总之,本研究通过转录组、植物激素定量和WGCNA分析,为芝麻植株高度发育提供了全面的知识。研究发现了几个与植物激素生物合成和信号转导途径相关的候选基因,为未来的研究提供了线索。此外,SiSCL9被鉴定为影响植株高度的关键基因,并在转基因拟南芥中得到了进一步验证。我们的研究为育种者改善芝麻植株结构提供了重要的基础。
CReDiT作者贡献声明
陈晟:方法论、验证、撰写——原始草稿的准备,所有作者均阅读并批准了最终稿件。
宋胜男:方法论、验证、数据整理,所有作者均阅读并批准了最终稿件。
周旺怡:验证,所有作者均阅读并批准了最终稿件。
塞努瓦·塞格拉·科菲·多苏:撰写——审查与编辑,所有作者均阅读并批准了最终稿件。
周蓉:形式分析、调查,所有作者均阅读并批准了最终稿件。
张艳鑫:资源提供,所有作者均阅读并批准了最终稿件。
李东华:调查,所有作者均阅读并批准了最终稿件。
游军:概念化、撰写——审查与编辑、监督和项目管理,所有作者均阅读并批准了最终稿件。
王林海:概念化、资源提供、撰写——审查与编辑、监督和项目管理,所有作者均阅读并批准了最终稿件。
数据可用性:数据将根据请求提供。
致谢:这项研究得到了中国农业科学院农业科技创新项目(CAAS-ASTIP-2021-OCRI)、湖北省国际科技合作项目(2022EHB034)、湖北省科技创新项目(2021-620-000-001-035)、中央公益性科研院所基本科研业务费(Y2022XK11,1610172022010)、农业农村部油料作物生物学与遗传改良重点实验室开放项目(KF2022002)以及中国农业研究系统(CARS-14)的资助。
附录A. 补充数据
本文的补充数据可以在线查看,网址为:https://doi.org/10.1016/j.plaphy.2023.107695。


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









