







《WUSCHEL同源盒蛋白家族的全面鉴定和表达分析揭示了其在玉米(Zea mays L.)发育和非生物胁迫响应中的特殊作用》
摘要:
玉米是全球重要的粮食和经济作物。WUSCHEL (WUS)相关的同源盒(WOX)转录因子(TF)家族在植物发育过程和非生物胁迫响应中起着重要作用。然而,关于WOX基因在玉米中的功能研究却很少。本研究利用最新的玉米B73参考基因组,鉴定出22个潜在的ZmWOX基因家族成员。除了染色体5以外,这22个ZmWOX基因均匀分布在其他九条染色体上,并显示出三次串联重复和十次片段重复事件。基于系统发育特征,ZmWOX基因被分为三个类群(如WUS、中间组和古老组),同一类群中的大多数ZmWOX基因和蛋白质结构相似。跨物种共线性结果表明某些WOX基因可能在进化上是保守的。ZmWOX家族成员的启动子区域富含光、植物生长/激素和非生物胁迫响应元件。组织特异性表达评估显示,ZmWOX基因可能在玉米生殖器官的发生中起重要作用。转录组数据和RT-qPCR分析进一步显示,六个ZmWOX基因(如ZmWOX1、4、6、13、16和18)受温度、盐和渍水胁迫的正向或负向调节。此外,两个ZmWOX基因,ZmWOX1和ZmWOX18都在非生物胁迫下上调。ZmWOX18定位于细胞核并具有转激活活性,而ZmWOX1定位于细胞质和细胞核,且没有转激活活性。总体而言,本研究为ZmWOX基因的进化关系提供了新的视角,并可能为进一步探测ZmWOX基因的生物学功能提供了资源。
关键词:非生物胁迫;WOX基因家族;全基因组分析;玉米;WUS同源盒
1. 引言
顶端分生组织的发育和侧生器官的分化影响植物的形状和高度。同源盒(HB)基因家族是植物中的一个大型基因家族(包含14个亚家族)。这些基因类别的突变可导致胚胎期或其他发育时期的异常[1]。WUSCHEL相关同源盒(WOX)转录因子(TF)基因组是最重要的HB家族之一。其家族成员的蛋白质N端包含一个保守的DNA结合区域,由三个α螺旋组成(如同源域和HD,包括60-66个氨基酸)[2]。除了保守的HD基序,WOX蛋白还在C端包含一些功能域,如典型的WUS-box结构域(TLXLFP,七个氨基酸,X表示任何氨基酸)、EAR基序(ERF相关的两性抑制)和酸性域[3,4]。许多物种中的WOX蛋白可根据其结构相似性分为三组:古老组、中间组和现代/WUS组[5]。古老的WOX基因广泛分布于植物中,中间组成员存在于维管植物中,而WUS仅存在于种子植物中[6]。
根据生物信息学分析,WOX基因家族已在许多植物中被鉴定出来,如拟南芥(15个成员)[7-9]、甜瓜(Cucumis melo L.)(11个成员)[10]、莲花(Nelumbo nucifera)(15个成员)[11]、番茄(10个成员)[12]、陆地棉(40个成员)[13]、向日葵(Helianthus annuus L.)(18个成员)[14]、面包小麦(14个成员)[15]、水稻(13个成员)[16,17]和高粱(11个成员)[18]。在拟南芥中,WOX基因在调控茎尖分生组织(SAM)/根尖分生组织(RAM)/维管分生组织和器官(胚和花)发育的初级和次级生长过程中具有重要作用,这些功能已被系统报道[19,20]。AtWUS蛋白在SAM中维持干细胞稳态,以确保正常的植物发育[21-23]。类似于AtSUS,AtWOX5在RAM中对于干细胞稳态至关重要[21,24]。AtWOX3在侧生器官的起始和生长中起作用[25]。AtWOX4与AtWOX14功能丰富,并接受来自CLE41/44-PXY复合物的信号,负责维管组织的形成[26]。AtWOX14还通过增强赤霉素(GA)合成来促进维管细胞发育[27]。AtWOX13基因参与根和花的发育[28]。在水稻中,OsWUS基因在叶原基和RAM中表达[17]。与AtWOX3同源的OsWOX3可能在叶片的发育中发挥重要作用[29]。OsWOX4在早期叶片发育中起关键调控作用。敲低OsWOX4会导致叶片发育缺陷[30]。OsWOX11在新生的冠根中表达,随后在根分生组织的细胞分裂区表达[31]。
除了在植物发育过程中起关键调控作用外,WOX转录因子(TF)还可以被各种非生物胁迫(例如干旱、低温和盐分)调节[32]。在水稻中,OsWOX11通过调节根毛的发育过程来提高水稻的抗旱能力[31]。包括OsWUS在内的八个OsWOX基因对干旱胁迫产生正响应。例如,OsWOX3和OsWOX5可以被盐诱导,而四个OsWOX家族成员对冷胁迫产生正响应[29]。棉花和黄瓜中的一些WOX基因家族成员也对几种环境胁迫产生正或负响应[18,33]。OsWOX13基因受干旱、寒冷和盐分调节。在水稻中,过表达OsWOX13显著增强了植物的抗旱胁迫能力[34]。过表达MdWOX13-1还通过激活活性氧清除系统增强了植物的抗旱胁迫能力[35]。
玉米是全球重要的作物。先前的研究基于古代基因组数据表明玉米中存在21个ZmWOX基因[16]。四个ZmWOX基因(如ZmNS1、ZmNS2、ZmWUS1和ZmWUS2)还参与了叶片发育并促进了玉米体细胞胚胎发生。ZmNS1和ZmNS2的双突变体导致叶片宽度变窄但叶片长度不变[36]。ZmWUS1和ZmWUS2均为拟南芥WUS的同源基因。同时,这两个基因的表达模式存在显著差异;ZmWUS1的转录在营养生长阶段发生动态变化,而ZmWUS2基因特异性地在幼叶原基中表达[37]。同时,ZmWUS2和BBM基因在玉米中的共同过表达显著增强了体细胞胚胎发生并缩短了基因转化的时间[38]。尽管已有上述发现,ZmWOX基因在玉米发育和非生物胁迫响应中的功能仍然大部分未知。
在本研究中,我们根据新组装的玉米B73参考基因组(Zm-B73-REFERENCE-NAM-5.0),鉴定了22个WOX基因并进行了全面分析。同时,探讨了ZmWOX基因在正常和非生物胁迫条件下的表达水平。此外,还分析了ZmWOX1和ZmWOX18的亚细胞定位和转录激活活性。本研究提供了关于ZmWOX基因进化关系的新信息,并可能有助于进一步了解ZmWOX基因的潜在功能。
2. 结果
2.1. ZmWOX基因的鉴定和特征
2010年的一项研究报告了包含21个WOX基因的玉米基因组[16]。在本研究中,基于新版玉米B73基因组(NAM-5.0),使用已知的AtWOX和OsWOX序列作为探针进行同源性搜索[15,16]。在去除重复序列并确认保守的HD结构域后,在玉米中鉴定出22个潜在的WOX基因(表1)。通过基因组搜索获得每个ZmWOX基因的详细位置信息(补充表S1)。这22个ZmWOX基因根据其在染色体区域中的位置被标记为ZmWOX1到ZmWOX22(补充表S1)。除了染色体5以外,其他九条染色体都含有ZmWOX基因。染色体8上有最多的ZmWOX基因(五个成员),其次是染色体3上有四个成员,而染色体1、7和9上分别只有一个ZmWOX基因(补充表S1)。ZmWOX基因的编码序列(CDS)从252 bp(ZmWOX8)到1551 bp(ZmWOX7)不等,22个ZmWOX蛋白包含83到516个氨基酸(aa)(平均:287 aa)(表1)。ZmWOX蛋白的等电点(PI)和分子量分别在5.56到10.52(平均:8.00)和10.01到53.26 kDa(平均:30.67)之间(表1)。根据网站的预测结果,大多数ZmWOX蛋白(17个成员)分布在细胞核中(表1),这与转录因子(TF)的定位一致。其他五个成员分别分布在叶绿体(三个成员)和线粒体(两个成员)(表1)。这可能负责质体中基因的转录调控。
2.2. ZmWOX蛋白的系统发育分析
拟南芥基因组包含15个WOX基因,这些基因的蛋白序列根据进化关系可以分为三组(例如现代/WUS、中间和古老组)[39]。我们使用高粱中已知的WOX蛋白序列(高粱中有12个WOX成员,补充表S1)、水稻(补充表S2)[17]、小麦(补充表S2)[15]、拟南芥(补充表S2)[8]和新鉴定的ZmWOX蛋白序列构建了一个系统发育树,以了解玉米中WOX基因的进化关系。如图1所示,来自五种植物的76个WOX蛋白也可以分为三类(例如WUS、中间和古老组),分别包含40、28和8个WOX基因(图1)。对于玉米WOX家族,十二个ZmWOX基因(例如ZmWOX2–5、9–11、14、16、17、21、22)属于WUS组,八个ZmWOX蛋白(例如WOX1、7、8、12、13、15、19、20)在中间组,只有两个ZmWOX基因(例如ZmWOX6和18)被划分到古老组(图1)。每组中的WOX蛋白都来自五种植物,表明这些物种中的WOX基因可能经历了相似的进化模式(图1)。同时,所有类别中的ZmWOX基因与SbWOX基因比与其他WOX更为接近(图1),表明玉米和高粱是进化上的直系同源关系。
2.3. ZmWOX基因的结构分析
使用MEME和TBtools分析了ZmWOX蛋白的基序和基因结构。根据MEME检测结果,22个ZmWOX中有10个保守基序。所有ZmWOX中都存在保守的HD结构域(例如,基序1和2,Helix1-Loop-Helix2-Turn-Helix3)(图S1)(图2A)。除了基序1和2外,一些基序仅存在于特定组中,反映了ZmWOX的多样性。中间组成员包含基序3、4、6、9和10。同时,除ZmWOX8(短序列)外,两个成员在C端均包含基序3-4(图2A)。ZmWOX6和ZmWOX18(古老成员)在N端还包含基序4(图2A)。在拟南芥中,WUS亚家族成员包含两个保守结构域:WUS盒和EAR样结构域。所有成员都包含WUS盒,而EAR基序仅在AtWUS中发现[39]。如图2A所示,除了ZmWOX9、11和14(短序列)外,所有玉米WUS成员中都鉴定出了基序5(WUS盒,TLXLFP,图S1)。我们还在ZmWOX2(ZmWUS1)和ZmWOX22(ZmWUS2)中发现了EAR结构域。
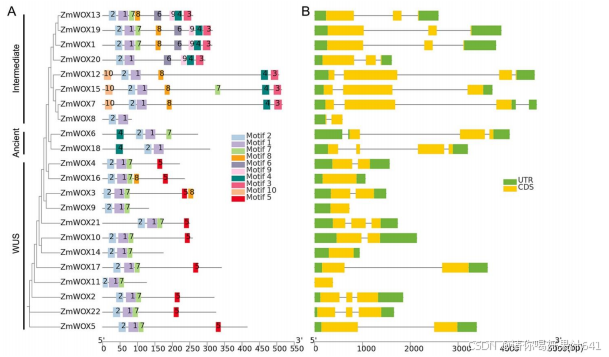
图2. ZmWOX基因的基因结构和保守蛋白基序。ZmWOX基因的聚类基于图1中显示的系统发育树。(A) ZmWOX蛋白中10个保守基序的分布(不同颜色的框)。基序1-10的常见序列显示在图S1中。(B) 22个ZmWOX基因的外显子-内含子结构。内含子表示为线条。
我们使用TBtools分析了ZmWOX基因的外显子-内含子分布。除了ZmWOX9和ZmWOX11仅有一个外显子且没有内含子外,所有ZmWOX都至少包含两个外显子和一个内含子(图2B)。同一组中的外显子和内含子长度相似,但在不同亚家族之间显示了较大的变异,主要是由于内含子序列的差异(图2B)。中间组成员的外显子序列比WUS和古老组的更长(图2B),导致了更长的蛋白序列(图2A)。总体而言,这些结果表明,同一组的ZmWOX基因在基因和蛋白质结构上显示出较小的变异,可能具有相同的功能。
2.4. ZmWOX基因的基因重复和共线性分析
除了染色体5外,22个ZmWOX基因广泛分布在其余九条染色体上(补充表S1和图3)。一些ZmWOX基因紧密排列,形成了三个串联重复事件(ZmWOX4/5/6、ZmWOX16/17/18和ZmWOX21/22)(图3)。根据基因重复事件分析,除了三个串联重复事件外(图3),在玉米基因组中还鉴定出了10对ZmWOX基因的片段重复(同源)(图3)。在这10对中,五对属于WUS家族(例如,ZmWOX2/22、ZmWOX3/9、ZmWOX4/16、ZmWOX5/17和ZmWOX10/14),四对属于中间组(例如,ZmWOX1/13、ZmWOX1/19、ZmWOX7/12和ZmWOX7/15),而古老组中只有ZmWOX6/18(图3)。这些结果表明,片段重复可能是推动ZmWOX家族扩展的主要力量。


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









