






本文内容速览:
蛋白质是生物体内各项生命活动的主要执行者,受到精密地调控。根据分子生物学的中心法则,遗传信息从DNA经由mRNA传递到蛋白质。在这个过程中包括四个主要的调控步骤:转录调控、转录后调控、翻译调控和翻译后调控,其中翻译调控占整个调控幅度的一半以上,超过了所有其他调控的总和(Schwanhäusser et al., 2011)。因此,翻译调控是生物体内非常重要的调控步骤。自20世纪70年代以来,许多研究聚焦于翻译调控过程,但由于技术难度和翻译调控机制的高度复杂性,直到近年来翻译调控才实现了全球范围内的研究。
翻译组学
从广义上来说,翻译组学(Translatomic)研究翻译调控过程中所涉及的所有成分,包括正在翻译的mRNA(也称为核糖体-新生肽链复合物mRNA,RNC-mRNA)、核糖体、转运RNA(tRNA)、调控性RNA(miRNA、lncRNA等)、新生多肽链和多种翻译调控因子(Zhao et al., 2019)。其中,研究RNC-mRNA是翻译组学的主要任务。RNC是指在翻译过程中由核糖体、新生肽链和mRNA组成的复合物,在这个复合物中,mRNA正在被核糖体读取并指导蛋白质的合成,RNC-mRNA的丰度和类型可以反映细胞中正在翻译的mRNA的情况。目前研究RNC-mRNA的技术有多聚核糖体图谱分析(Polysome profiling)、核糖体新生链复合物测序(RNC-seq)、翻译核糖体亲和纯化测序(TRAP-seq)和核糖体印迹测序(Ribosome profiling,又称为Ribo-seq),这些也是常见的翻译组学技术。
翻译组学技术
1、Polysome profiling
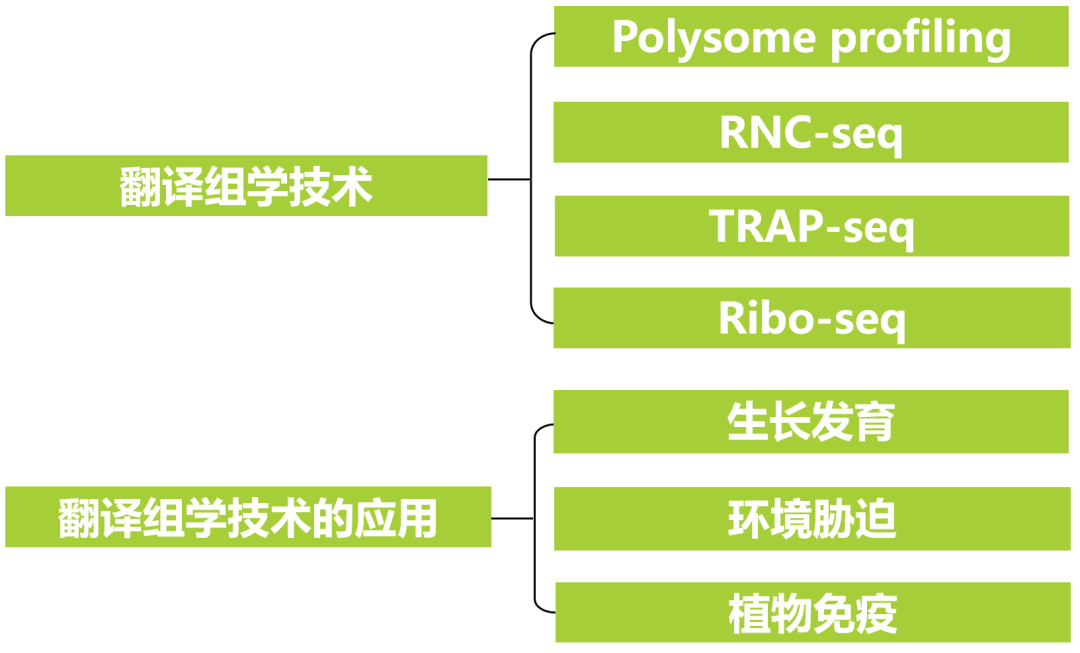
Polysome profiling基于蔗糖密度梯度离心法,利用核糖体沉降系数较大的特性,将游离的RNA、40S核糖体小亚基、60S核糖体大亚基、单个核糖体和多聚核糖体(结合有不同数目核糖体的mRNA)分散到不同的蔗糖密度梯度溶液中。通过离心,这些组分按照密度不同被分离开来(图1)。然后使用RNA-seq、微阵列、Northern blot、RT-qPCR分析各个组分中的mRNA,以反映转录本的翻译分布状况,还可使用质谱和Western blot分析组分中的蛋白质,以研究翻译过程中重要的翻译调控蛋白。这项技术通常用于检测翻译中发生巨大变化的RNA,如在高渗压力下,观察到单个核糖体组分的显著增加(Zhang et al., 2010);在氧化应激下,观察到与mRNA结合的核糖体数量显著增加(Zhong et al., 2015)。
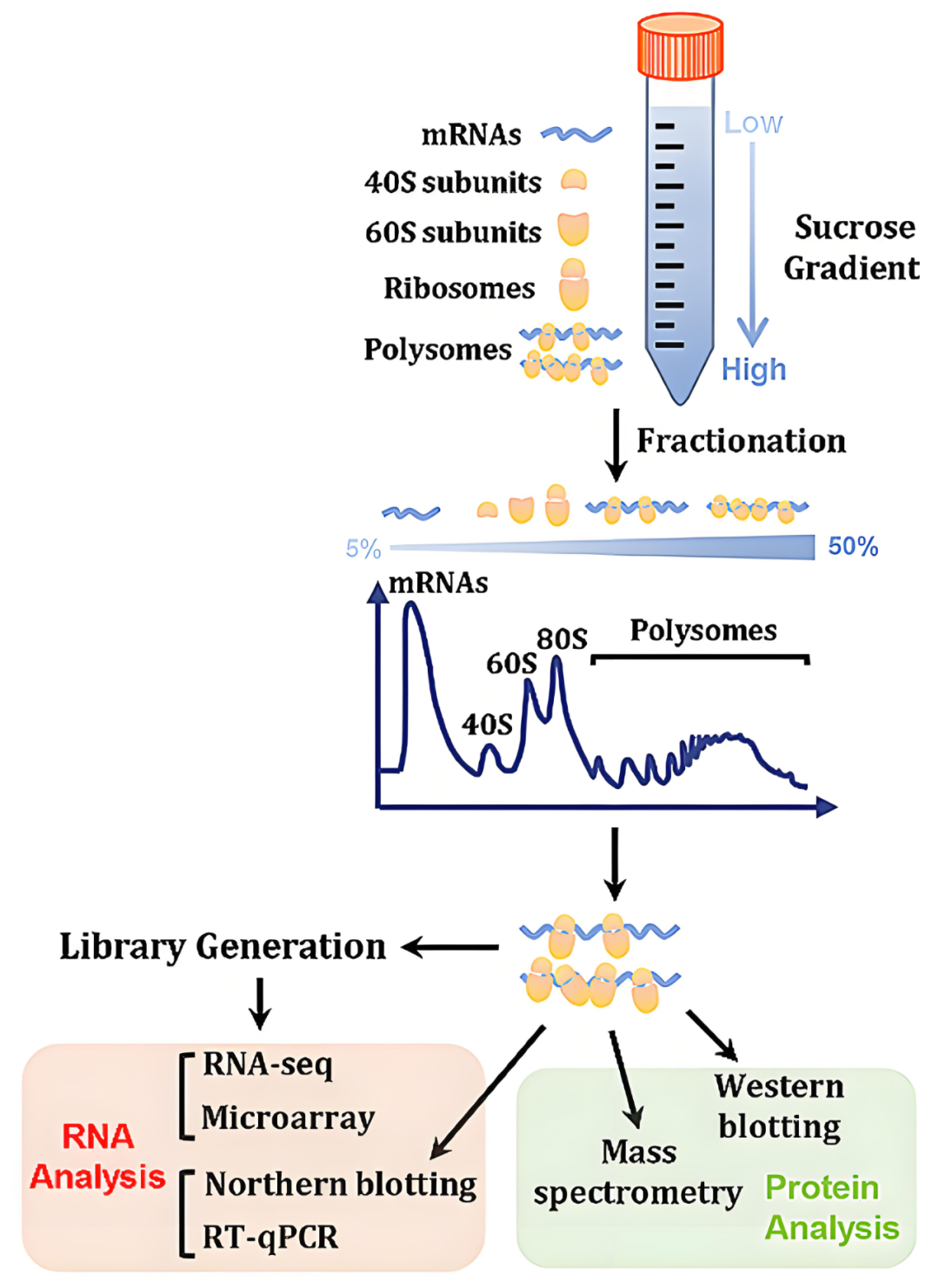
图1 Polysome profiling技术实验原理(Su et al., 2024)。
该技术可以检测核糖体密度分布,对胞质RNA做分级。该技术的主要缺点是难以对所有RNC-mRNA进行深入分析。这是因为在样品分级过程中,细胞质裂解物被蔗糖密度梯度溶液稀释,从多个蔗糖组分中回收RNC-mRNA的比例相当低。除非使用大量起始材料,否则难以满足全谱分析(如RNA-seq和微阵列)所需的样品量(King et al., 2016)。此外,超速离心可能导致假多聚体和脂质颗粒等高分子复合物与核糖体共沉淀。同时,该技术操作复杂、耗时长,不能提供核糖体的位置信息。
2、RNC-seq
RNC-seq又称为全长翻译mRNA测序(Wang et al., 2013),该技术在Polysome profiling的基础上进行了改良,使用30%的蔗糖溶液作为缓冲液,通过超速离心将核糖体组分与游离mRNA和其他细胞组分分离开来(图2)。由于所有的核糖体组分(RNC Pellet)都沉淀在管底,抽提沉淀中的RNA,即可得到RNC-mRNA,并能有效避免高浓度蔗糖的干扰,然后结合高通量测序即可获得所有RNC-mRNA的全长信息,包括丰度和类型。通过优化离心和蔗糖缓冲液条件,RNC的回收率可达90%,且在适当缓冲液条件下,RNC仍保持翻译活性(Zhang et al., 2009)。
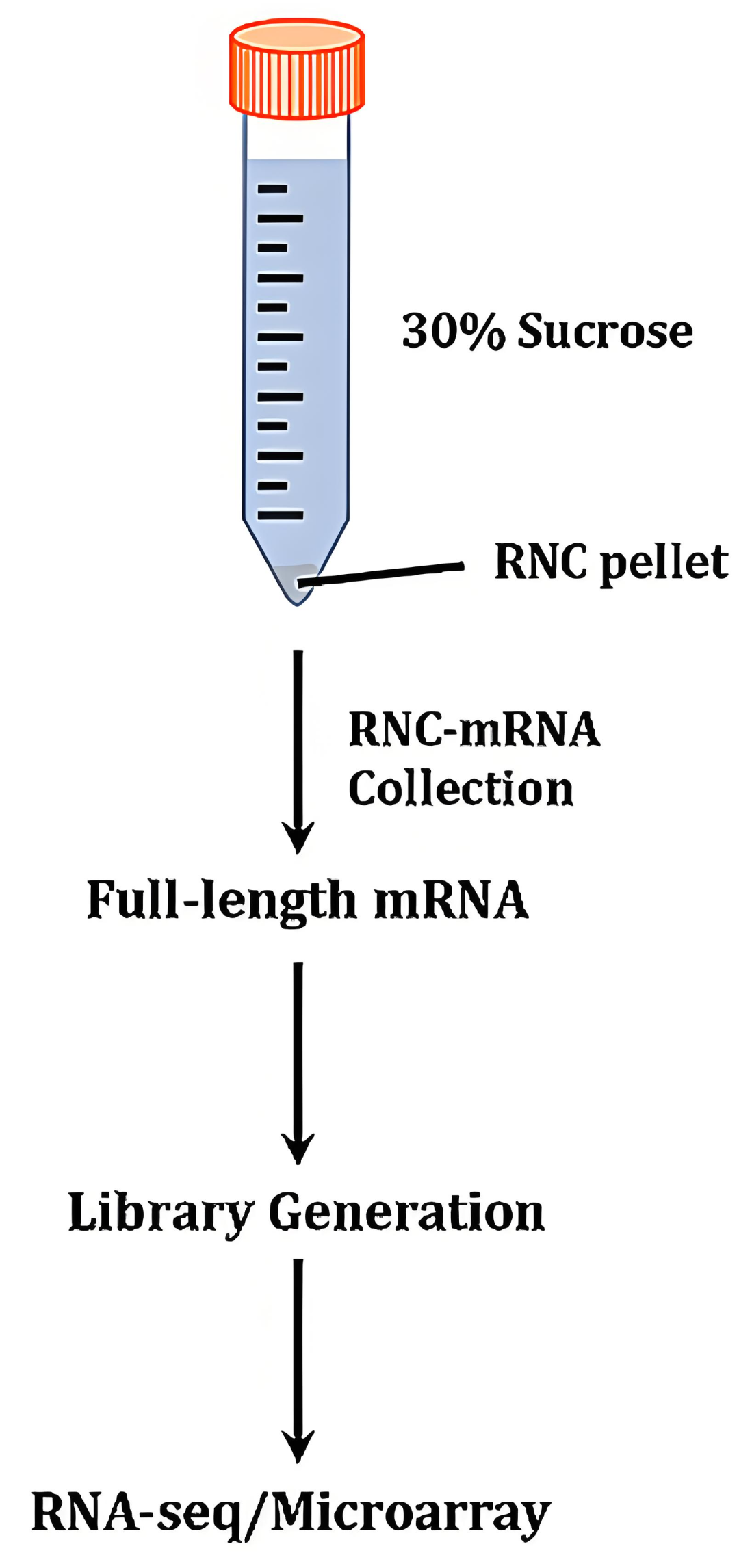
图2 RNC-seq技术原理(Su et al., 2024)。
由于核糖体与mRNA非共价结合,因此RNC非常脆弱,容易在分离过程中解离或导致mRNA降解。该技术的难点在于分离完整的RNC-mRNA,处理不当将得不到全长RNC-mRNA,并造成RNC-mRNA定量存在偏差。该技术也无法获得翻译过程中核糖体的位置信息。
3、TRAP-seq
TRAP-seq通过转基因技术给核糖体蛋白带上亲和纯化标签,并使用组织特异性启动子控制该标签的表达(Inada et al., 2002)。带标签的核糖体和与之结合的mRNA可以被特定的抗体捕获,去除rRNA后,进行RNA-seq或Microarray检测,从而获得特定组织类型中RNC-mRNA的信息(图3)。
图3 TRAP-seq技术原理(Su et al., 2024)。
TRAP-seq对分析复杂组织样品中的翻译组具有优势,可以分离出特定组织类型的RNC-mRNA,并且由于该技术不使用超速离心,不易受到假多聚体和脂质颗粒等非核糖体物质污染。然而,该技术需要通过转基因来产生标记的核糖体蛋白。这一过程耗时、昂贵,不适用于尚未建立稳定转化方法的物种(Wang et al., 2014)。此外,亲和标签可能会干扰核糖体的功能,改变自然的生理状态。该技术同样也不能提供核糖体的位置信息。
4、Ribo-seq
Ribo-seq使用低浓度核糖核酸酶(RNase)处理细胞裂解物,降解掉没有核糖体覆盖的mRNA片段,可获得被核糖体保护的22-35nt mRNA小片段(称为Ribosome footprints,RFP),最后对这些RFP进行高通量测序(Ingolia et al., 2012)(图4)。使用此方法可获得所有RNC-mRNA上核糖体的分布和密度,推测起始密码子位置(包括非ATG起始)、密码子偏好性、上游开放阅读框(uORF)、翻译停滞等信息,从而能够更详细地分析翻译调控,这些方面是其他翻译组学技术无法做到的。2016年,超分辨率核糖体分析技术被开发出来,这是一种优化的Ribo-seq方法,能够在单个转录本中观察到每隔3nt的周期性分布,这种周期性反映了核糖体在mRNA上的移动和翻译过程中的结构特征(Hsu et al., 2016)。此外,该方法提高了Ribo-seq发现小开放阅读框(sORF)和未注释的新编码区域的能力。
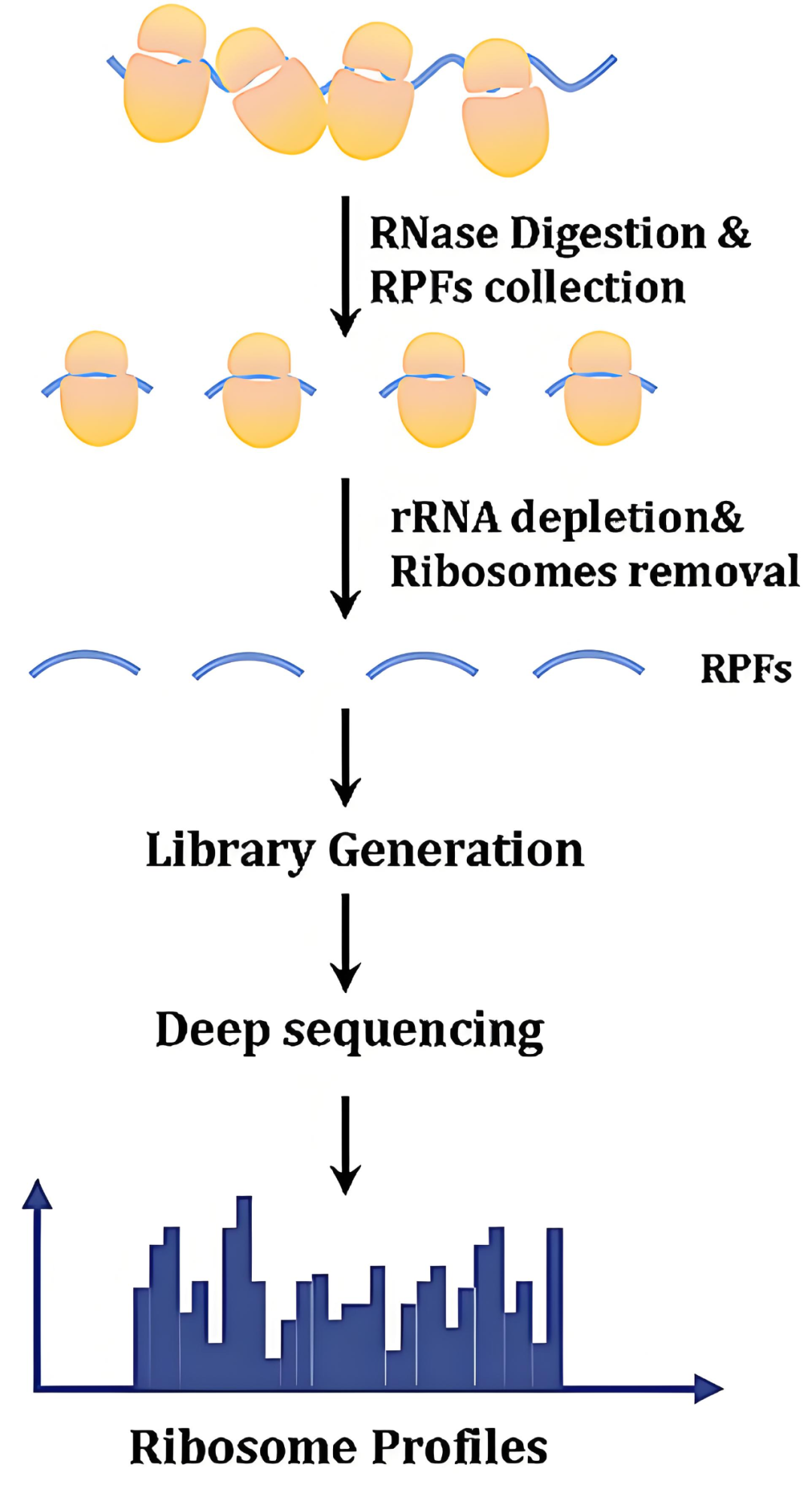
图4 Ribo-seq技术原理(Su et al., 2024)。
由于Ribo-seq主要分析核糖体与mRNA结合的位置,即编码区序列(CDS),因此无法高效分析与翻译调控高度相关的非编码区(UTR)(King et al., 2016)。此外,Ribo-seq产生的许多RFP会被比对到非编码RNA上,这表明该技术存在一定的假阳性率。另一个不足是RFP的长度较短,为了获得足够覆盖度,需要进行高深度测序,这意味着更高的测序和分析成本(Guttman et al., 2013)。
总之,前面列举的四种技术都有其优势和适用场景。选择哪种技术取决于具体的研究目标和实验条件。如果想要直观了解翻译动态变化图谱,Polysome Profiling是更好的选择;如果想要了解核糖体的组成和活性,RNC-seq更有优势;如果需要分析特定组织的翻译调控信息,TRAP-seq更适合;如果需要直接测定翻译效率、鉴定翻译起始位点、鉴定uORF以及新编码区域等,Ribo-seq是最佳选择。下面通过一些具体的文献案例来看看这些技术都是如何应用的。
翻译组学技术的应用
翻译组学技术已经应用到植物生长发育、环境胁迫、植物免疫、物种进化等多个方面,揭示了各种生物过程中的翻译调控机制。
1、生长发育
水稻对全球粮食安全和人类健康至关重要,其产量和品质受到籽粒形状的显著影响,通常,长而窄的籽粒性状具有更好的外观品质和更高的商业价值(Zhao et al., 2022)。尽管已鉴定出许多影响水稻籽粒形状的基因或数量性状位点(QTL),但许多优良的基因未被充分应用于水稻育种,如正向调控GLW7(第7号染色体上的籽粒长度和重量关键基因)和GW7(第7号染色体上的籽粒宽度关键基因)的表达在育种中具有积极作用。近年来,结合Ribo-seq技术对uORF的鉴定和基因编辑技术,为正向调控基因表达提供了一种可行的思路。uORF是位于(5'-UTR)内的主要调控元件,可能影响主要ORF(mORF)的翻译,并位于18-57%的植物mORF的上游(Zhong et al., 2023)。最近的研究表明,编辑uORF能够改变植物表型,并且已在植物性状改良中得到应用(Wang et al., 2024)。然而,与水稻籽粒形状相关的uORF尚未被发表,这限制了具有育种价值的基因在改良水稻外观质量中的应用。
2024年10月,扬州大学张韬、刘巧泉课题组联合西南大学张勇课题组在Plant Physiology上发表了一篇题为“Improving rice grain shape through upstream open reading frame editing-mediated translation regulation”的研究论文,作者通过Ribo-seq数据鉴定出了OsGLW7和OsGW7的uORF,采用CRISPR-Cas9技术对两个uORF进行编辑,提高了两个基因的翻译水平,最终有效改良了水稻的籽粒形状、产量和品质(图5)。
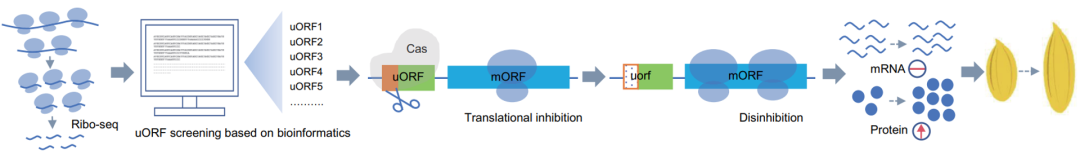
图5 Ribo-seq技术原理(Yang et al., 2024)。
首先,作者通过分析公共数据库中的Ribo-seq数据发现,OsGLW7和OsGW7 5'-UTR中分别存在1个和2个潜在的uORF。利用双荧光素酶报告基因实验进行初步评估,结果显示与自然条件下的uORF相比,GLW7-uorf1和GW7-uorf1序列中ATG突变为AAA提高了萤火虫荧光素酶(LUC)/海肾荧光素酶(REN)的活性比,而GW7-uorf2对活性比没有影响(图6A、C)。因此,作者通过CRISPR-Cas9技术对GLW7-uORF1和GW7-uORF1进行编辑。测序显示所有uorf 突变体在ATG位点存在插入或缺失,RT-qPCR结果显示GLW7和GW7的转录水平无显著变化(图6B、D),而Western结果表明GLW7蛋白水平显著提升(图6E)。这些结果表明,虽然转录水平没有变化,但是uORF序列发生突变提高了翻译水平。
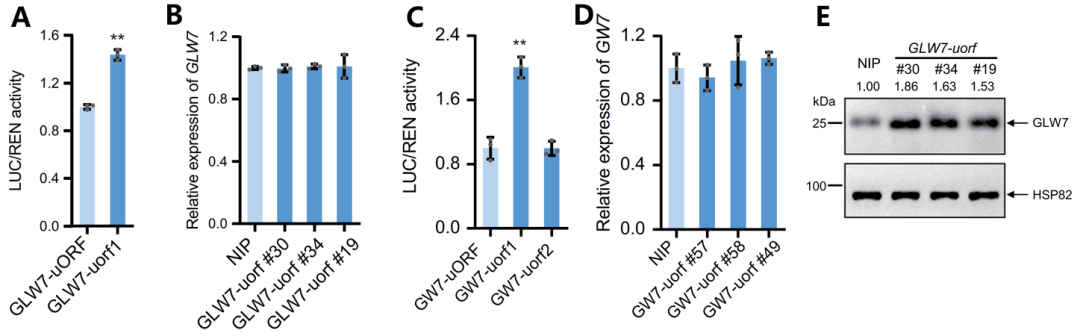
图6 GLW7和GW7的uORF序列突变提高了两个基因的翻译水平(Yang et al., 2024)。(A、C)双荧光素酶报告基因实验分析uORF序列的ATG突变为AAA对GLW7(A)和GW7(C) mORF翻译的影响;(B、D)野生型和uorf突变体幼穗中GLW7和GW7基因的转录水平,NIP,野生型日本晴;(E)Western blot检测GLW7的翻译水平,以水稻HSP82为内参。
进一步功能分析显示,相较于野生型,所有GLW7-uorf 突变体显著增加了籽粒长度和长宽比,而不影响籽粒宽度,并且突变体水稻的直链淀粉含量较低,胶稠度更高,口感更柔软。这些结果表明,调控GLW7-uORF能有效改良籽粒形状、产量和品质。作者还分析了GW7-uorf 突变体的籽粒形状,结果显示其籽粒长度和长宽比增加,宽度减少,千粒重增加。此外,GW7-uorf 突变体的粉质度较低,但直链淀粉含量、胶稠度和黏度无明显变化。结果表明,编辑GW7的uORF可改善籽粒形状和重量,同时保持口感质量。
2、环境胁迫
高温胁迫是全球小麦生产的主要限制因素,在全球范围内造成了巨大的产量损失。了解与高温胁迫相关的分子机制将对提高小麦的耐热性和产量具有重要意义。2021年11月,中国农业大学彭惠茹和辛明明课题组在New Phytologist上发表了一篇题为“Stress granule-associated TaMBF1c confers thermotolerance through regulating specific mRNA translation in wheat (Triticum aestivum)”的研究论文,在高温胁迫下,小麦TaMBF1c基因上调表达,作者基于CRISPR-Cas9技术、Polysome profiling分析联合RNA-seq以及蛋白质互作组技术,揭示了TaMBF1c通过调节小麦中特定基因的翻译来调控高温胁迫反应。
作者首先在染色体7A、7B和7D上鉴定了TaMBF1c的同源基因(图7a)。RT-qPCR实验发现TaMBF1c同源基因的转录水平在高温胁迫下都急剧升高,并具有相似的表达模式(图7b)。作者分析发现TaMBF1c启动子中有三个热激元件(HSE),EMSA和转录激活实验证明HSFA6e可以与其结合并激活TaMBF1c的转录。表型分析发现,在高温胁迫条件下过表达株系相比于野生型表现出更强的耐热性,而沉默和基因编辑株系则表现出耐热性下降。这些结果表明TaMBF1c是小麦耐热所必需的。
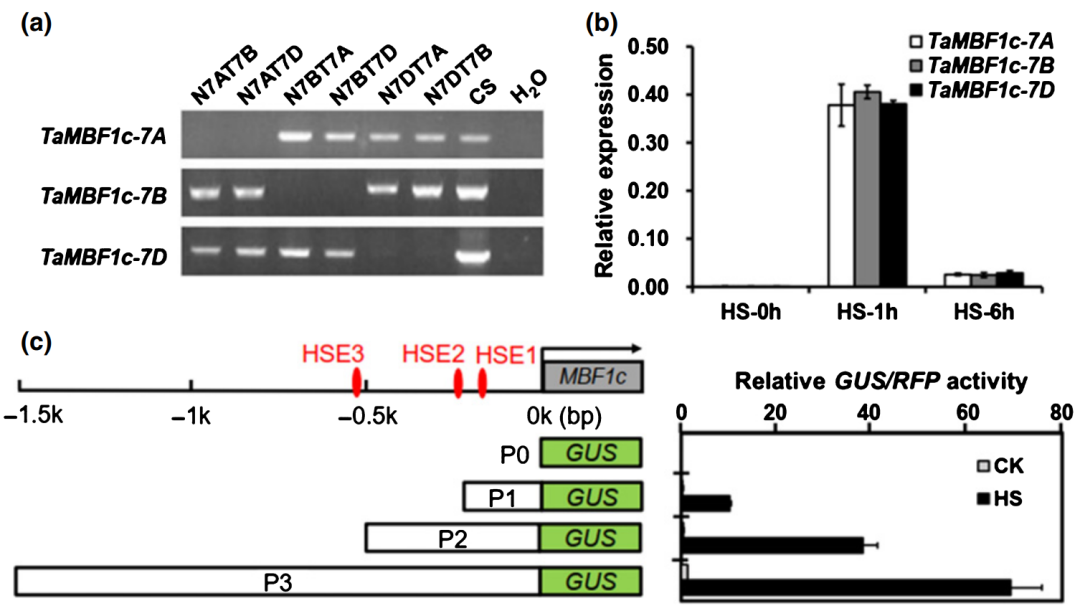
图7 TaMBF1c的染色体定位、表达模式和启动子活性分析(Tian et al., 2022)。(a)三个同源TaMBF1c基因的染色体位置。普通小麦是异源六倍体,其染色体组成为AABBCC,A、B、D 分别代表三个不同的染色体组。小麦品种Chinese Spring(CS)的单体-四体株系(Nullisomic-tetrasomic line)是特殊染色体变异材料(图中N7AT7B、N7AT7D、N7BT7A、N7BT7D、N7DT7A、N7DT7B),例如N7AT7B表示缺失了两条A染色体组的7号染色体,同时增加了两条B染色体组的7号染色体,可用于确定基因在小麦染色体上的具体位置;(b)RT-qPCR分析小麦TaMBF1c基因在高温胁迫下的表达模式;(c)TaMBF1c-7B的启动子在烟草高温胁迫条件下活性分析实验。红点表示位于TaMBF1c-7B启动子区域预测的HSE motif。TaMBF1c-7B起始密码子上游200bp、525bp或1500 bp分别截短为P1、P2或P3,并与GUS报告基因融合。通过RFP标准化GUS的活性水平。HS,高温胁迫处理;CK,对照。
作者对正常生长条件下(22℃;CK)、高温胁迫条件(42℃,3h;HS)和恢复条件(高温胁迫后22℃保温2h;RE)下WT和TaMBF1c敲除株系(KO-1)的Polysome profiling进行了比较,并对分离出多聚核糖体结合的RNA和总RNA进行测序。结果显示TaMBF1c影响了高温胁迫相关基因的翻译效率,这些基因在“序列特异性DNA结合”途径中显著富集(图8A)。此外,基因表达网络分析表明TaMBF1c与热休克蛋白HSP的翻译密切相关(图8B)。
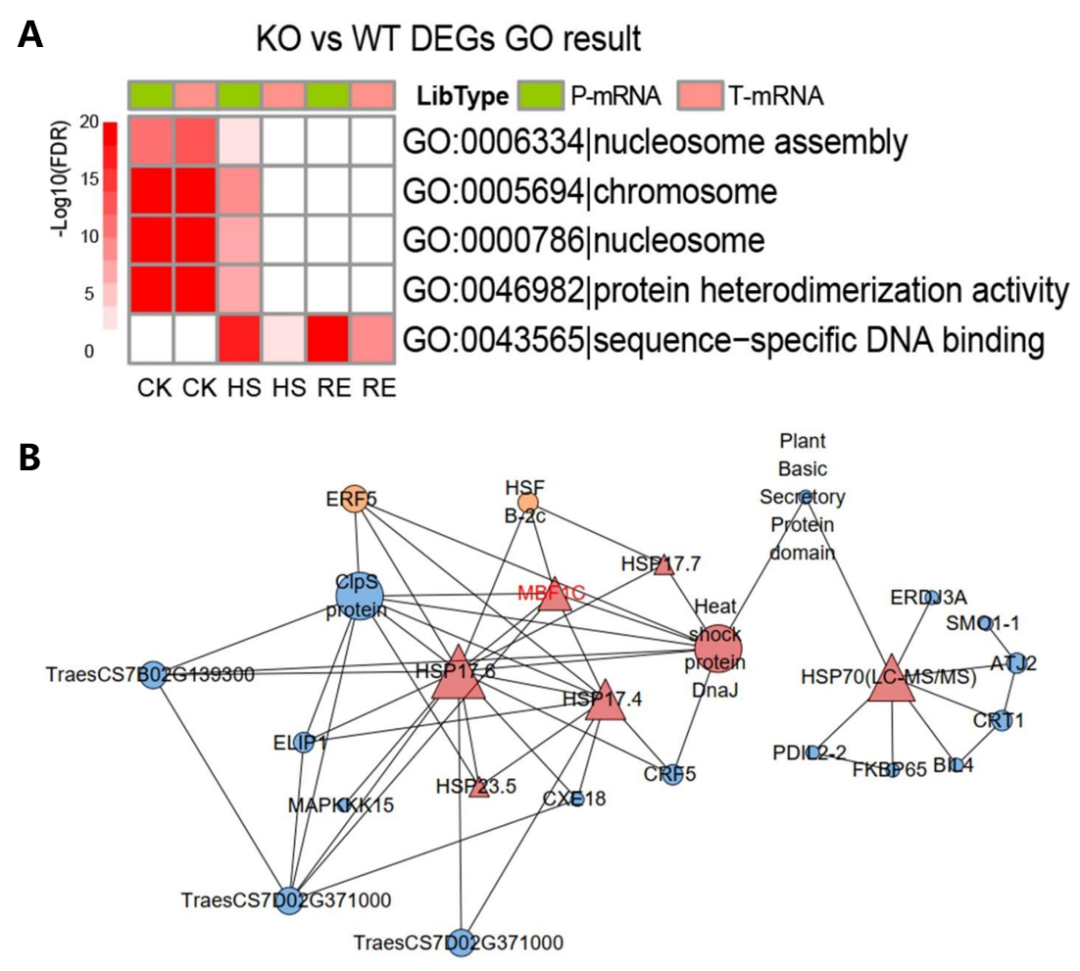
图8 GO富集分析和基因表达网络分析(Tian et al., 2022)。(A)TaMBF1c的编辑突变体下调基因的GO富集分析。选择了十个显著富集的GO term进行绘图。FDR:多重检验阳性发现错误率;CK:正常生长条件;HS:高温胁迫条件;RE:恢复条件;(B)基因表达网络分析。
镁(Mg)是植物生长和发育的必需营养元素,参与叶绿素合成、光合作用、能量转移、维持离子平衡等多个生理过程。植物在缺镁条件下会出现黄化表型(叶绿素降解)等症状,影响植物的生长发育,导致植物的产量下降。越来越多的研究表明在胁迫条件下,转录水平和翻译水平存在不同步性(Lee et al., 2011)。目前对植物缺镁的研究主要集中在转录水平,对缺镁条件下植物翻译调控的研究较少,但是蛋白质才是生命活动的直接承担者,从遗传信息传递到产生具有实际功能的蛋白质,翻译过程起着决定性作用。因此,在翻译水平上研究植物对缺镁胁迫的响应具有十分重要的意义。
2021年9月,美国伊利诺伊大学陈利清课题组在Journal of Inregrative Plant Biology题为“Combined analyses of translatome and transcriptome in Arabidopsis reveal new players responding to magnesium deficiency”的研究论文,作者通过比较缺镁情况下基因转录水平(RNA-seq)和翻译水平(TRAP-seq)的差异,鉴定出仅在翻译水平被调控的26个新基因,并通过构建突变体验证了部分基因在低镁胁迫响应中的功能,该研究结果强调了翻译调控对植物响应营养胁迫的重要性,并为研究植物响应低镁胁迫的翻译调控机制奠定了基础。
首先,作者为了研究拟南芥的茎和根在翻译水平上对缺镁的反应,构建了拟南芥转基因株系,该株系组成型表达带有His6-Flag标签的核糖体蛋白RPL18,RPL18位于核糖体60S大亚基的溶剂侧(Solvent side),用于TRAP-seq实验。获得的拟南芥转基因株系在正常或缺镁条件下培养。从处理24小时后的植物中提取总mRNA和核糖体相关mRNA,分别进行RNA-seq和TRAP-seq,以分析转录组和翻译组的变化。结果显示无论是在正常生长条件还是缺镁的情况下,基因的转录水平和翻译水平均存在着很大的差异。在缺镁条件下,拟南芥根和茎中有约10%的基因翻译状态发生(|z-scores|>1.5)显著变化(图9),表明翻译调控在植物响应低镁胁迫中起重要作用。
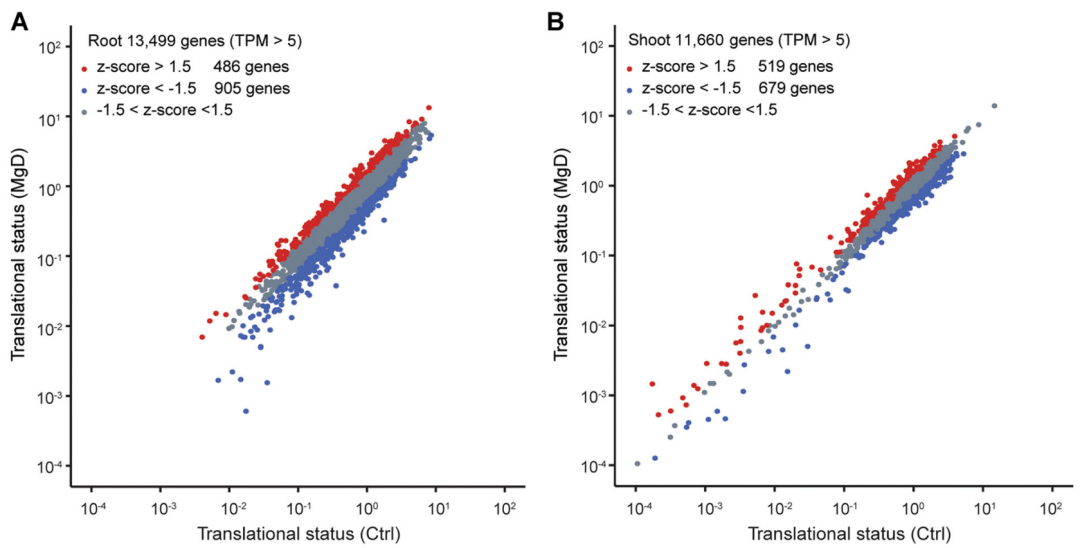
图9 缺镁条件下,转录组和翻译组的比较(Li et al., 2021)。(A)比较根中13499个基因在正常和缺镁条件下的翻译状态;(B)比较茎中11660个基因在正常和缺镁条件下茎的翻译状态。通过计算z值来确定基因翻译状态的变化,z值大于1.5(红点)说明受到缺镁条件的影响基因的翻译状态上调,z值小于1.5(蓝点)说明受到缺镁条件的影响基因翻译状态下调的,z值介于+1.5和-1.5之间(灰点)表示基因翻译状态的变化不太显著。Ctrl:对照组;MgD:缺镁组。
进一步作者分析转录水平和翻译水平差异表达的基因,并在缺镁条件下对这些基因进行突变体表型筛选。其中,编码b-ZIP家族转录因子的基因HY5在缺镁条件下翻译水平下调,其突变体hy5表现出黄化表型(图10A),表明HY5在缺镁条件下起关键作用。编码氢离子/阳离子交换蛋白的基因CAX1和CAX3在缺镁条件下翻译水平上调,其突变体表现出黄化表型(图10B),说明它们参与缺镁条件下的离子稳态调节。编码泛素蛋白的基因UBQ11在缺镁条件下翻译水平下调,其突变体ubq11表现出黄化表型(图10C),表明泛素化途径可能参与镁缺乏响应。这些基因是如何参与响应低镁胁迫的具体分子机制还有待进一步探索。

图10 缺镁条件下,野生型和突变体的表型(Li et al., 2021)。(A)缺镁条件下野生型和突变体hy5的表型;(B)缺镁条件下野生型、cax1和cax3突变体的表型;(C)缺镁条件下野生型和突变体ubq11的表型。
3、植物免疫
植物在病原体胁迫或非胁迫条件下,免疫基因的转录和翻译需要被严格调控,以确保在病原体入侵时能够快速激活免疫反应,同时在没有病原体时避免过度消耗植物的能源。因此,植物需要精细调控免疫基因的表达。翻译重编程在植物免疫调控中发挥关键作用,但其具体机制尚不清楚。
2023年2月16日,武汉大学胥国勇课题组在Nature Plants上发表一篇题为“Plant HEM1 specifies a condensation domain to control immune gene translation”的研究论文,作者通过对拟南芥野生型和敲除突变体HEM1进行了Ribo-seq、RNA-seq和多组学联合分析,结果发现HEM1突变会增加促死亡免疫基因的翻译能力,证实了HEM1可限制免疫基因的过度翻译,构成了免疫激活的刹车机制。
作者利用uORF控制的翻译报告基因进行遗传筛选,该报告基因的LUC翻译受到转录因子TBF1基因的uORF的抑制。在之前的研究中,该LUC报告基因的翻译能够被模式触发免疫(PTI)和效应因子触发免疫(ETI)的激活所诱导,可用于筛选具有影响免疫基因翻译活性的基因。实验发现hem1突变体会增强报告基因翻译活性。通过全基因组重测序和转基因回补实验,确定HEM1基因的突变是导致翻译活性增强的原因。这些结果表明HEM1是翻译调控因子,能够抑制免疫基因的翻译,从而在植物免疫反应中起到负调控作用。
作者利用Ribo-seq技术,比较了野生型和hem1突变体在没有免疫激活和ETI激活时的翻译效率变化。在ETI激活时,hem1突变体中一些与细胞死亡相关的免疫基因表现出更高的翻译效率(图11)。这表明HEM1通过抑制这些基因的翻译来控制免疫反应的强度。
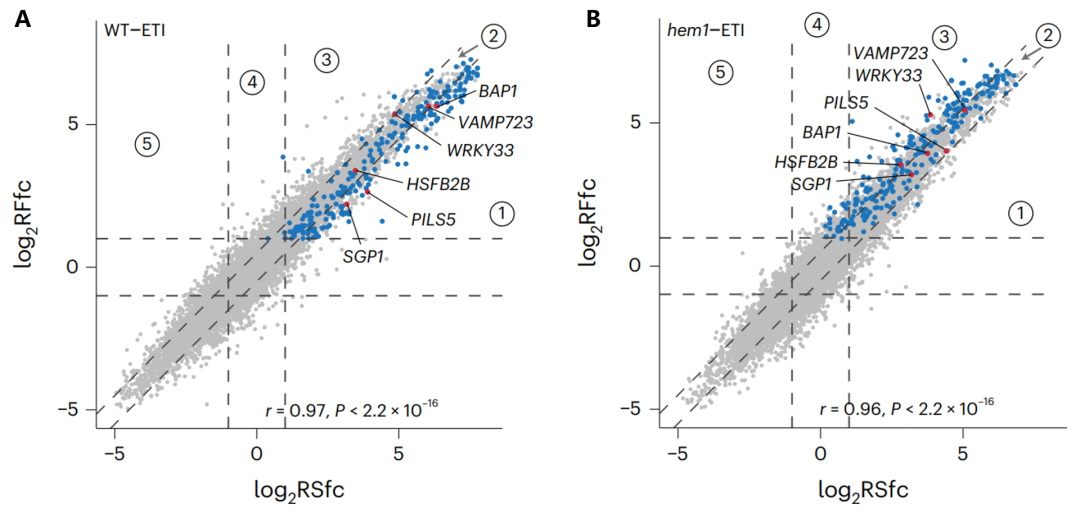
图11 Ribo-seq结果显示野生型(A)和hem1突变体(B)在ETI激活时翻译变化(RFfc)和转录变化(RSfc)之间的关系(Zhou et al., 2023)。蓝点表示hem1中翻译效率(TE)增加的246个基因,其中部分基因用红点突出显示。
接着,作者通过CRISPR-Cas9技术敲除了HEM1的LCD结构域,并分析了其对翻译调控和免疫反应的影响。结果表明HEM1的LCD结构域赋予其相分离能力,这种能力在ETI激活时被增强。敲除LCD结构域会导致ETI过程中发生更多的细胞死亡,表明LCD结构域在抑制免疫激活中起着关键作用。最后,作者通过免疫共沉淀和质谱分析HEM1与翻译过程中的多个组分的相互作用,结果显示HEM1能够与翻译因子相互作用,并在ETI激活时形成凝聚体。这种凝聚体可能限制了翻译因子发挥作用,从而抑制了免疫基因的翻译。HEM1通过其LCD结构域介导的相分离来控制免疫基因的翻译,平衡植物的组织健康和疾病抗性。
小远叨叨
伯小远今天为大家介绍了翻译组学技术及其技术案例,如果大家对这些技术感兴趣,后续会针对单个翻译组学技术的实验流程和实际应用进行详细地介绍。随着科技的进步,我们拥有了更多更先进的技术手段来开展科学研究。同时,新技术的出现往往也预示着新的突破和机遇,在这样的时代背景下,学习永无止境。我们不仅要掌握现有的技术,更要保持对新技术的敏感和好奇心,持续学习和无限探索都是我们前进的动力。
References:
Guttman M, Russell P, Ingolia NT, Weissman JS, Lander ES. Ribosome profiling provides evidence that large noncoding RNAs do not encode proteins. Cell. 2013 Jul 3;154(1):240-51.
Hsu PY, Calviello L, Wu HL, Li FW, Rothfels CJ, Ohler U, Benfey PN. Super-resolution ribosome profiling reveals unannotated translation events in Arabidopsis. Proc Natl Acad Sci U S A. 2016 Nov 8;113(45):E7126-E7135.
Inada T, Winstall E, Tarun SZ Jr, Yates JR 3rd, Schieltz D, Sachs AB. One-step affinity purification of the yeast ribosome and its associated proteins and mRNAs. RNA. 2002 Jul;8(7):948-58.
Ingolia NT, Brar GA, Rouskin S, McGeachy AM, Weissman JS. The ribosome profiling strategy for monitoring translation in vivo by deep sequencing of ribosome-protected mRNA fragments. Nat Protoc. 2012 Jul 26;7(8):1534-50.
King HA, Gerber AP. Translatome profiling: methods for genome-scale analysis of mRNA translation. Brief Funct Genomics. 2016 Jan;15(1):22-31.
Lee MV, Topper SE, Hubler SL, Hose J, Wenger CD, Coon JJ, Gasch AP. A dynamic model of proteome changes reveals new roles for transcript alteration in yeast. Mol Syst Biol. 2011 Jul 19;7:514.
Li Y, Li Q, Beuchat G, Zeng H, Zhang C, Chen LQ. Combined analyses of translatome and transcriptome in Arabidopsis reveal new players responding to magnesium deficiency. J Integr Plant Biol. 2021 Dec;63(12):2075-2092.
Schwanhäusser B, Busse D, Li N, Dittmar G, Schuchhardt J, Wolf J, Chen W, Selbach M. Global quantification of mammalian gene expression control. Nature. 2011 May 19;473(7347):337-42.
Su D, Ding C, Qiu J, Yang G, Wang R, Liu Y, Tao J, Luo W, Weng G, Zhang T. Ribosome profiling: a powerful tool in oncological research. Biomark Res. 2024 Jan 25;12(1):11.
Tian X, Qin Z, Zhao Y, Wen J, Lan T, Zhang L, Wang F, Qin D, Yu K, Zhao A, Hu Z, Yao Y, Ni Z, Sun Q, De Smet I, Peng H, Xin M. Stress granule-associated TaMBF1c confers thermotolerance through regulating specific mRNA translation in wheat (Triticum aestivum). New Phytol. 2022 Feb;233(4):1719-1731.
Wang J, Liu J, Guo Z. Natural uORF variation in plants. Trends Plant Sci. 2024 Mar;29(3):290-302.
Wang T, Cui Y, Jin J, Guo J, Wang G, Yin X, He QY, Zhang G. Translating mRNAs strongly correlate to proteins in a multivariate manner and their translation ratios are phenotype specific. Nucleic Acids Res. 2013 May;41(9):4743-54.
Wang Y, Jiao Y. Translating ribosome affinity purification (TRAP) for cell-specific translation profiling in developing flowers. Methods Mol Biol. 2014;1110:323-8.
Yang Q, Zhu W, Tang X, Wu Y, Liu G, Zhao D, Liu Q, Zhang Y, Zhang T. Improving rice grain shape through upstream ORF editing-mediated translation regulation. Plant Physiol. 2024 Dec 23;197(1):kiae557.
Zhang G, Hubalewska M, Ignatova Z. Transient ribosomal attenuation coordinates protein synthesis and co-translational folding. Nat Struct Mol Biol. 2009 Mar;16(3):274-80.
Zhao D, Zhang C, Li Q, Liu Q. Genetic control of grain appearance quality in rice. Biotechnol Adv. 2022 Nov;60:108014.
Zhao J, Qin B, Nikolay R, Spahn CMT, Zhang G. Translatomics: The Global View of Translation. Int J Mol Sci. 2019 Jan 8;20(1):212.
Zhong J, Xiao C, Gu W, Du G, Sun X, He QY, Zhang G. Transfer RNAs Mediate the Rapid Adaptation of Escherichia coli to Oxidative Stress. PLoS Genet. 2015 Jun 19;11(6):e1005302.
Zhong V, Archibald BN, Brophy JAN. Transcriptional and post-transcriptional controls for tuning gene expression in plants. Curr Opin Plant Biol. 2023 Feb;71:102315.
Zhou Y, Niu R, Tang Z, Mou R, Wang Z, Zhu S, Yang H, Ding P, Xu G. Plant HEM1 specifies a condensation domain to control immune gene translation. Nat Plants. 2023 Feb;9(2):289-301.

















 4826
4826

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









