







菊石的复原是兼具严谨和幻想的。一方面,它拥有举世无双的丰富化石库存,外壳又直接暴露在外,无软组织包被,在硬体方面复原可信度很高;另一方面,从未有任何一块软体印痕被发现,这使得对习性,生态更为重要的软体部分只能靠猜测和幻想。
在菊石部分的最后,本篇将是对前文的总结——如何复原一枚菊石?
目录:
1:自然形态
硬体:
壳形:吃错时代了吗?
颜色:提前入棺了吗?
口盖:戴错帽子了吗?
软体:
腕:九十条、十条还是?
眼:鹦鹉螺还是蛸?睁眼还是闭眼?
漏斗:半闭,全闭,还是?
喙:角质喙不是祖传的?
收缩肌:怎么缩进壳里?
2:交互动作
捕食:
食性:韭菜都吃点啥?
游泳:薯条能游多快?
被捕食:
天敌:都有哪些食客?
反抗动作:游走还是缩回去?
被攻击位置:怎么人道吃菊?
被攻击后状态:被咬后浮起还是下沉?
寿命与寄生虫:塞翁失马,焉知祸福
“菊落”:空壳里的蜗居
3:奇思妙想
引颈就戮型
想当海兔型
无法形容型
硬体:
壳形
让我们先从最少争议的外壳开始。
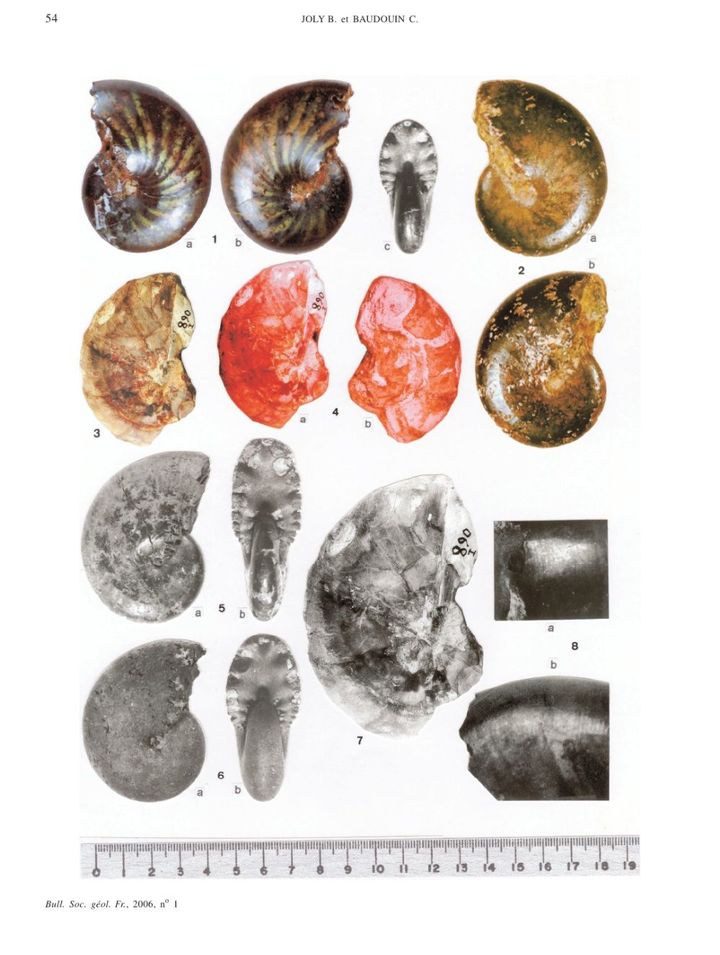
大小:
菊石的大小随着时代增大。泥盆记,石炭纪的菊石不超过7.5cm,二叠纪约15cm,三叠纪20cm左右,大型可达80cm;侏罗纪个体50—80cm的很多,白垩纪可达1.2m,最大的达到2.55m。
材质:
菊石外壳由四层组成:最外侧是壳朊层,内部为纤状层—珍珠层—纤状层,均由文石组成。为适应游泳生活,壳壁很薄,1.5mm的壳厚20~30um,在5cm的壳中仅厚0.1~0.2mm,乃至数米长的直壳鹦鹉螺也只有1cm厚。
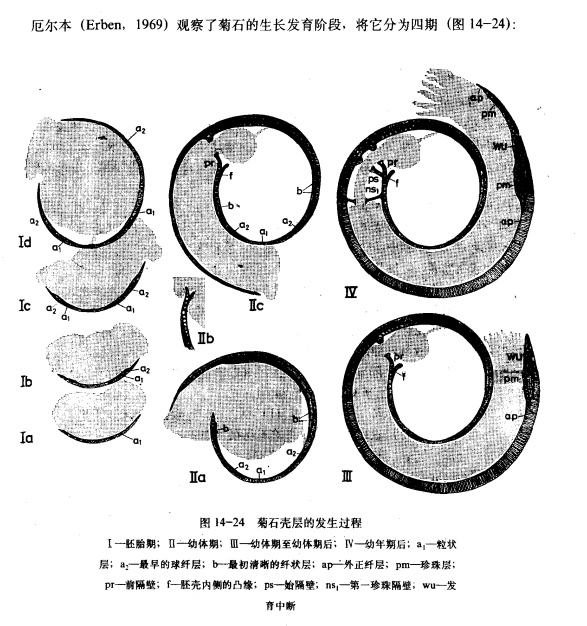
形状:
各个时代的菊石有着不同的壳体特征。泥盆纪的无棱菊石有光滑或带有生长纹的壳体,或是类似于棱菊石的四分球形壳;海神石壳表光滑,各旋环宽度相同,无脐孔;棱菊石多四分壳,但也有不明显者;齿菊石和真菊石形态多样,在白垩纪大量异形壳出现。


但在外壳形状确定的同时,还有三处仍需注意:
1:补全缺失的体室
由于无气室隔板支撑,绝大多数菊石化石都没有保存较为脆弱的体室,而菊石的体室又比鹦鹉螺大得多,开口结构也多种多样,不能简单的加长了之。一般而言,古生代菊石可以在气室基础上延长四分之一个旋环,中生代延长半个到一圈以上。
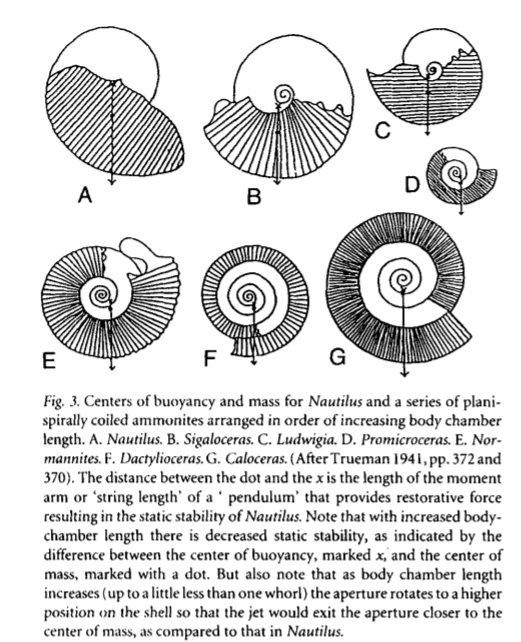
2:口部结构
一般菊石开口处呈椭圆形或圆形,但也有三角形乃至不规则的种类。不过截面在复原中并不重要——毕竟看不到。更为主要的是口部的各种装饰:普遍具有的腹弯,延长腹弯,侧垂(马蹄菊石,镰菊石,饰菊石),包住前一旋环的侧垂(象牙菊石),瘤和刺(瘤菊石)等等。
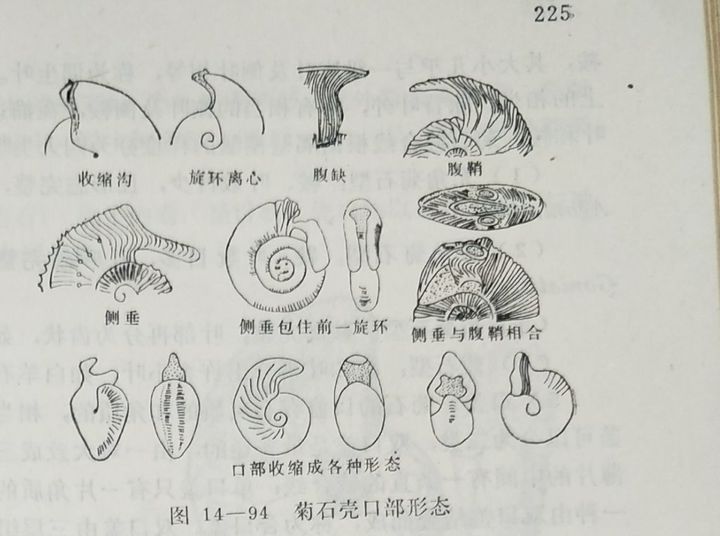
3:补全刺和瘤
白垩纪菊石,尤其是勾菊石的壳上通常有刺,但很难保存,绝大多数只是在壳上留下刺的附着痕迹。但有的时候痕迹很不明显,比如常见的窦维雷菊石(Douvilleiceras),勾菊石亚目—多维尔菊石超科。它的通常状态是这样的:

然而,它的真正样子是这样的:
这样的:

所以,只要发现有哪里颜色不太对,就可以添个刺试一下…
4:缝合线不要露在表面
缝合线是气室与外壳相接处的形态,不会露在壳表。
颜色:
与一般认为和复原的红白相间不同,菊石更可能是黑白二色(Keupp 2000),这种颜色模式已经在Craspeditidae科和叶菊石的化石中得到证实。当然,对于其他菊石而言,更加花哨的复原也是完全可行的——毕竟没人见过,而且很难想象多样性如此丰富的菊石只有一种配色。
当然,直接画个石头壳上去还是算了吧,这样无异于给昆虫穿化石外骨骼……

口盖:
菊石口盖(operculum)有四种:单口盖(anapthychi),双口盖(Aptychi),合口盖(synapthychi)和三分口盖(triapthychi)。其中,单口盖见于古生代,三分口盖只在晚二叠纪的华南有发现,双口盖和合口盖见于中生代。但在现在,双口盖(或包括合口盖)通常被认为是菊石的喙,单口盖和三分口盖也有当成喙的;
因此,中生代菊石的复原图通常不带有口盖,取而代之的是一张血盆大口(误,在外面看不到)。古生代理论上是单口盖,不过绝大多数复原图都画成鹦鹉螺式……

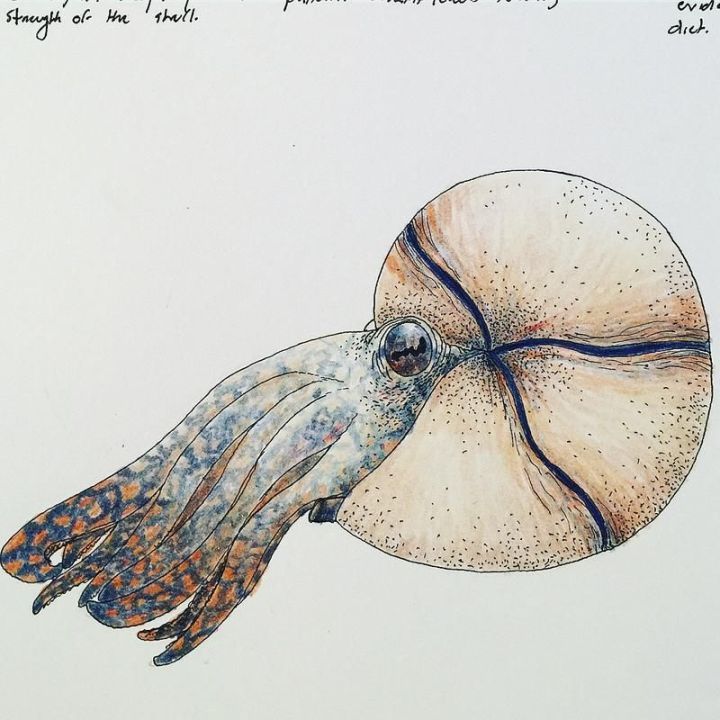
软体:
腕:
鹦鹉螺和蛸的胚胎都是十腕(Kröger et al. 2011),鹦鹉螺腕在成长中分裂,最终形成雄性60条,雌性90条的腕。因此菊石的腕也应当是十的倍数,至于到底是几十条……那就随便了。
但鉴于菊石和蛸同属新头足纲,是蛸类的近亲,而蛸类刚刚出现的原鱿就是十腕,所以一般还是十条比较好。
但实话来说,菊石和蛸的结构并不太一样。起码来说,蛸类化石的特征结构:墨囊和腕钩始终没有在菊石中发现;这么说的话,在复原腕的时候大可随心所欲。一般的复原都有吸盘,但腕钩是绝没有的。

触腕:
随缘。十腕总目的触腕平时缩在触腕囊内,和其他腕等长,因此在复原时画不画都是一样……在2016年的文章中,建议复原时带有触腕:“触腕对于外部有壳的头足类动物在远处捕捉食物非常有用;因为对这些软体动物来说,快速向前冲刺可能很困难。”

眼:
基本都是蛸式。头足类的眼睛经历了小孔成像眼(鹦鹉螺)—开眼—闭眼(眼眶外有膜)—眼睑的演变,想用什么基本随缘。
一般来说,平旋菊石和远洋型异形菊石(杆菊石,船菊石,钩菊石)不带眼睑,塔菊石的那堆像腹足类的复原成章鱼样子。

漏斗:
鹦鹉螺的漏斗为半闭式,左右两漏斗管不完全愈合,喷水动力较弱;蛸类则为全闭。
由于发现了软骨痕迹,菊石和蛸类一样具有漏斗锁定装置(或“外套膜--漏斗锁定装置“),由两对软骨组成,身体两侧各有一对,在每一对中,其中一部分位于漏斗的底部,另一部分连接在外套膜上(Bizikov 2008)。在此之外,另有两条肌肉带(漏斗收缩肌)附着到闭锁装置,肌肉带的另一端附在壳侧的“侧部连接肌痕”上。这个装置的存在表明菊石应是全闭漏斗。
但外套膜在壳内的扩张空间是有限的,这必然导致菊石难以快速运动。鹦鹉螺没有闭锁装置,通过伸缩头部来增大空间;那对于有闭锁装置的菊石呢?
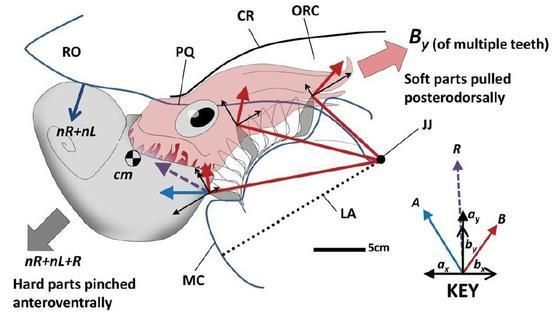
桑德斯和沃德(1994)提出了另一个假设:菊石的主要动力来源不是外套膜的收缩,而是通过压缩大而肌肉发达的漏斗实现的。在这种情况下,必须将软组织的动能转移到壳体。这就需要一个组织将漏斗和外壳连接起来。如果没有,漏斗的反作用力只会将软组织压入壳内,造成浪费。因此,漏斗闭锁装置的外套膜侧应当有组织附着在壳体上,如图所示:

大漏斗占满了开口的下侧,外套膜部分用于封闭壳口,并将动力转移到壳上。这张图便是目前相对正统的复原。

喙:
菊石的喙与蛸类截然不同。其颚主要由两部分组成,外侧和蛸一样为角质,而内侧为钙化结核( calcified concretion), 分别称为上下喙嘴石( rhyncholites and conchorhynch,也即口盖),下颌包被上颌,和鸟喙的结合方式相反。其中上颌与其他软体动物的口同源,下颌发生在上颌之后,由特化细胞分泌。齿舌7列,位于下颌。Lehmann认为菊石下颚宽大的侧壁起到了鳃盖的作用,而TANABE等认为该结论有待商榷。
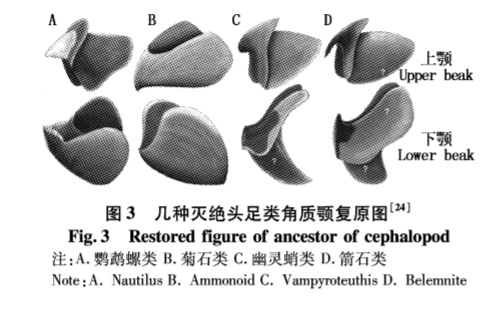
收缩肌:
菊石有三对收缩肌和一条中心肌肉,用于将身体拉回壳内。三对分别为颈收缩肌(附着在背部肌痕上,收缩闭锁装置上部)、头收缩肌(背部肌痕,头和腕)和漏斗收缩肌(侧部肌痕,收回漏斗),中心肌肉作用尚不明确。
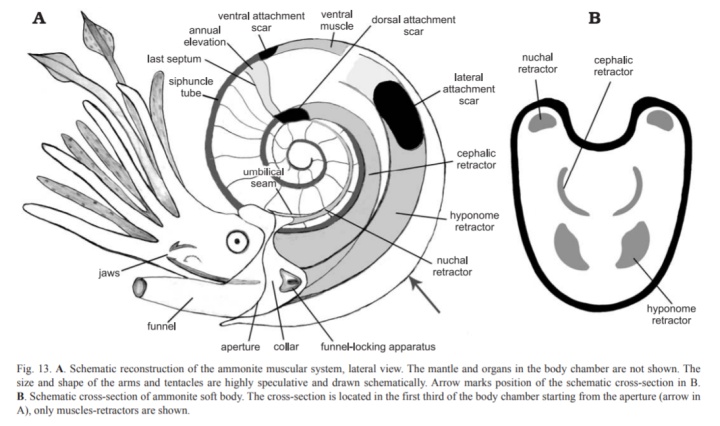
勾菊石的漏斗收缩肌退化,与适应漂游生活相应,但保留了头颈收缩肌以便收回壳内。
2:交互动作
捕食:
食性
虽说菊石被称为海洋薯条,但确确实实保存下来的作为食物和捕食食物的记录都不是很多。
大部分菊石的主食是底栖无脊椎动物。一块杆菊石标本(AMNH66253)的喙中保存了等脚类(isopod,)、和腹足类幼体等无脊椎动物的结构残余。另一块钩菊石的壳内部竟然保存了海百合……没错,就是海百合。
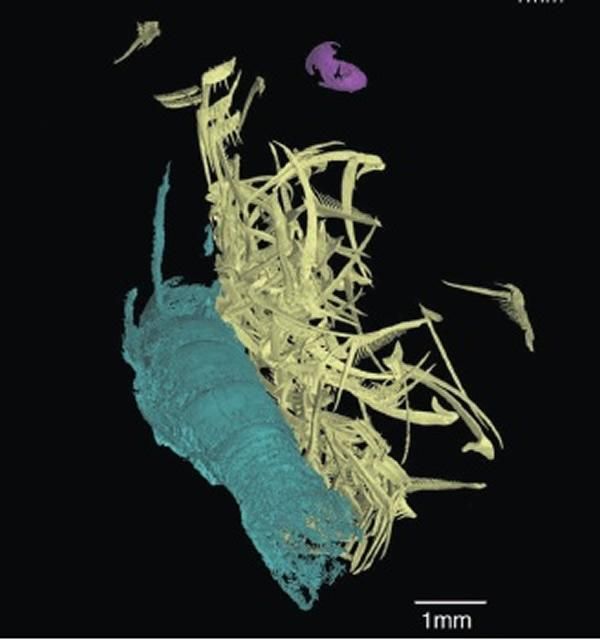
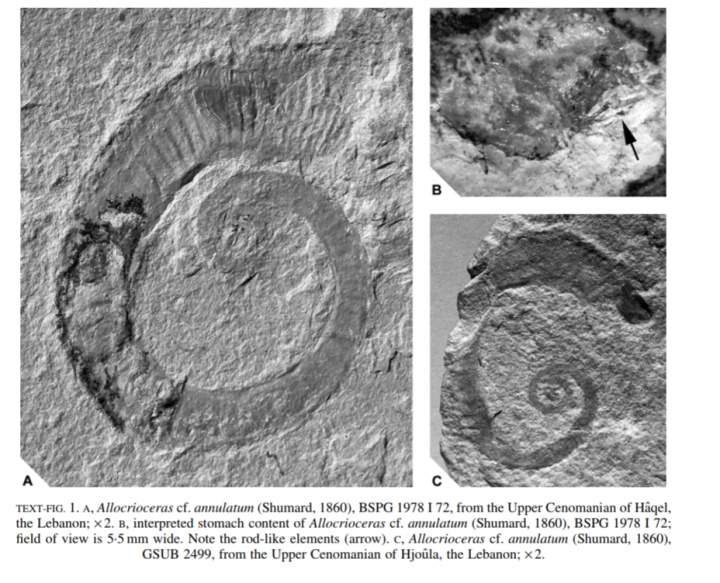
另外,一些菊石壳表的长条状伤痕表明它们以螃蟹为食。有螃蟹就有虾,有海百合就有海星,有腹足类就有双壳类……(疯狂胡编……啊这,为啥不说有浮游生物就有沧龙呢?)
游泳
菊石主要是垂直移动的生物。蛸类在游泳时外套膜收缩提供动力,同时改变漏斗朝向,配合鳍和腕调节方向。由于同时使用漏斗作为动力来源和转向装置,菊石可能难以像现代蛸类一样正向游动,而是始终保持倒游状态(鹦鹉螺就基本不会正游)。
在十腕目做长距离游泳时,十腕紧闭以减小阻力;菊石可能也以此姿势游泳。但在自然博物馆的复原中,菊石像章鱼一样张开腕划水移动……总之,一无二随,一无二随嘛。
菊石快速种类速度约为30~40cm/s,略低于同等大小的鱼类;慢速种类低于12cm/s。




被捕食:
天敌:
菊石最著名的天敌便是旋齿鲨。在经典的复原图中,旋齿鲨用内侧的牙抓住猎物的身体,同时用外面的牙把甲壳推出去。不过,旋齿鲨的食性也有争论—有些人认为旋齿鲨是吃珊瑚的。
确切留下证据的天敌并不多。一起是嵌入菊石壳中的弓鲛Planohybodus的牙齿,证明弓鲛咬过菊石;另一次是鹦鹉螺上的沧龙的咬痕,两次攻击都发生在气室。第二只鹦鹉螺上有两种大小不同的穿孔,意味着它曾遭受过来自两个不同沧龙个体的攻击。
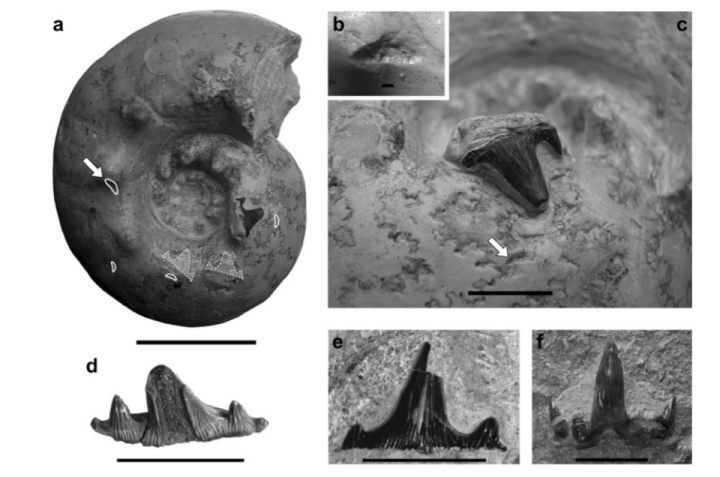
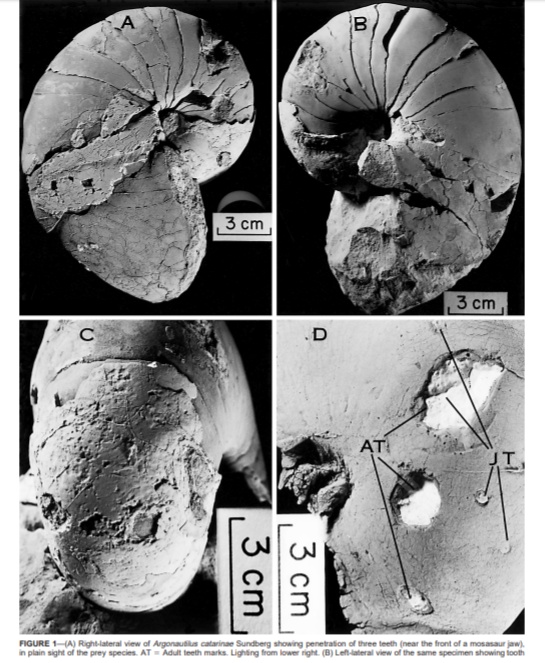
对于第二具化石上的穿孔做个粗略推算的话,1cm的牙齿直径约对应24cm的下颌,沧龙体长2~3米,鹦鹉螺直径13cm,壳体并没有被压碎。这么一看,外壳头足类的壳还算结实?
被攻击位置
虽说确切的两次攻击都在气室,但菊石上的伤疤体室居多,占了80%以上。底栖动物的反抗攻击基本也在体室——毕竟菊石用头对着它们。
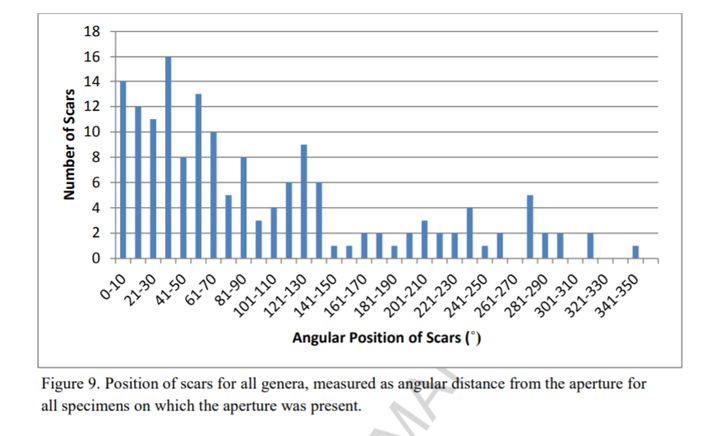
反抗动作
腹足类被攻击时,会缩回壳内;章鱼被攻击时,会加速逃离。那菊石呢?勾菊石放弃了漏斗收缩肌,但保留了头部收缩肌以便收回壳内;它们体室的瘤,肋和棘刺也是用来保护柔软的身体。那么结论就很明显了。被攻击时,鹦鹉螺会缩回身体,用口盖封闭壳口;菊石也应是一样。
不过这么说的话,如果菊石是缩起来的,旋齿鲨怎么咬呢……?
被攻击后状态:
菊石的浮力是远远过剩的。即使部分气室漏气,菊石仍可上浮;只有大部分气室被摧毁,菊石才会失去浮力。加之菊石体管并不直接暴露,而是表面有钙化壳朊包被,气室漏气也不会很大影响体管的调节。
如果菊石没有即死,气室和体室的损伤都可以被修复。但到底怎么修复的呢?我也不知道QAQ
寿命与寄生虫:
菊石由于游泳生活和生命周期短,体表附生物相对较少。但仍有一些蠕虫可以寄生在菊石上:查尔茅斯泥岩组的管生蠕虫最常见的生长方式是附着在幼年菊石的脐带上,随着菊石长大,最终围绕着菊石旋环,开口在壳口后方,保持与水流方向平行;腹部中间的位置使蠕虫能够展开它位于菊石两侧的鳃。另一些蠕虫则是寄生在死后的菊石空壳里。在菊石上生长对蠕虫有益,但对菊石有害—带有表生蠕虫管菊石死亡时的大小与蜗牛管的额外重量成反比。



蠕虫在一个季节内完全长成,按照蠕虫的生长纹推算,菊石长到全尺寸需要两到三年,因此菊石的寿命应当比蛸类(一年)更长。
另外的寄生虫是蔓足类。斯特雷门虾密集地附生在菊石壳上,有一只菊石身上甚至有三十多只藤壶,包括祖孙三代一家老小。藤壶和壳体一同保存下来,甚至保存了不同的动作状态,显示它们是被活埋的。相对于管生蠕虫,藤壶体型较小,并不很干扰菊石的生存。
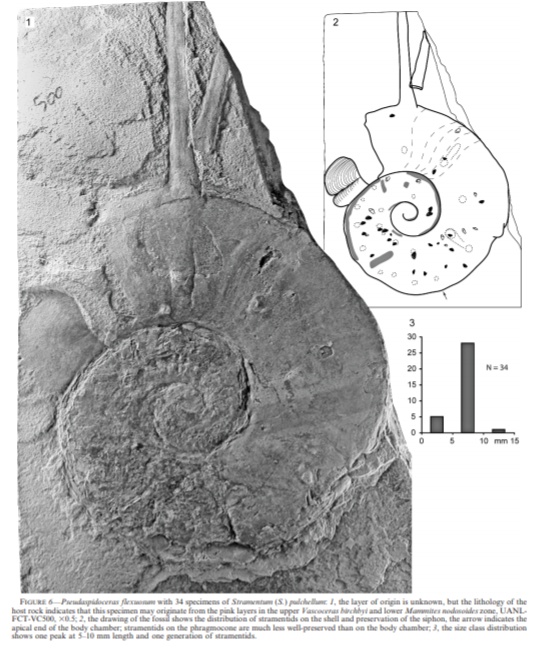

有甲壳类就有软体动物。双壳类,如牡蛎Liostrearoemeri也在活体菊石上寄生,同样扶老携幼,尊卑有序,老者居中,小者拱列……等下,为什么寄生还搞这一出啊?
牡蛎当然不晓得礼制。实际上,原因是菊石中心(幼体时)的牡蛎是最先附着上的……外侧旋环上的牡蛎或是它的后代,或是后附着的牡蛎幼体,当然小了。
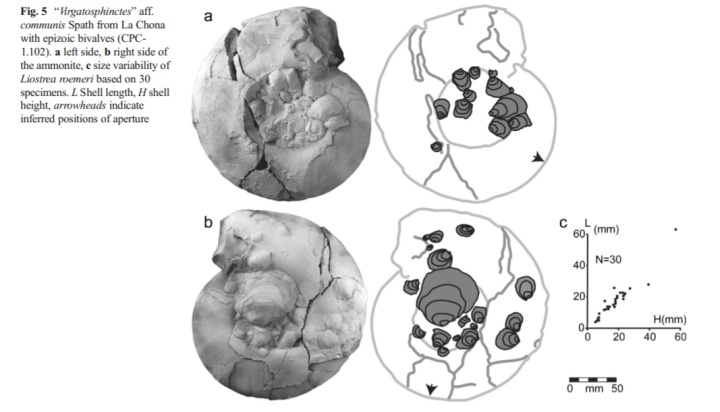
“菊落”
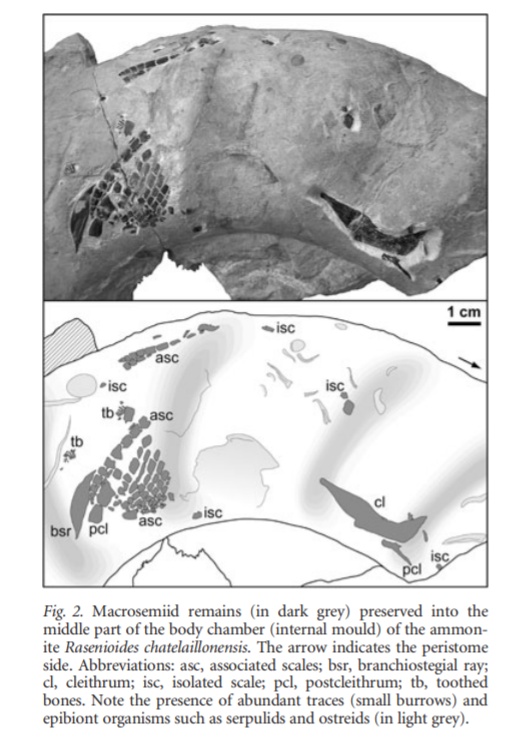
菊石沉到海底的空壳也成为海底生物的乐园。除了上述所叙的寄生虫在死后继续利用外,在菊石空壳内还发现过硬骨鱼类和镰龙虾。

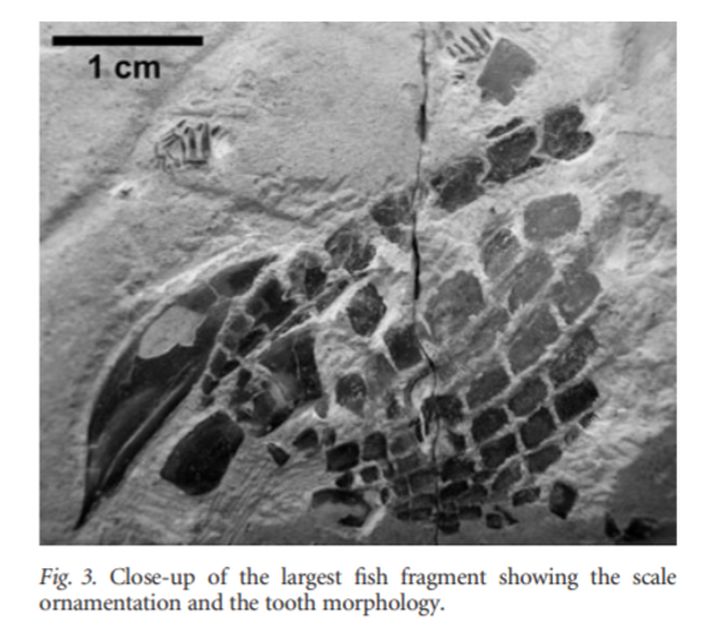
在侏罗纪菊石的壳内发现过一只辐鳍鱼类Macrosemiidae,它出现在菊石的体室内,骨骼完整,排除了被捕食的可能。外壳内壁上的表生生物的发育表明这条鱼在沉积物完全充满体腔之前进入了住室。它可能是使用空菊石壳作为避难所,或可能用于产卵和孵卵。

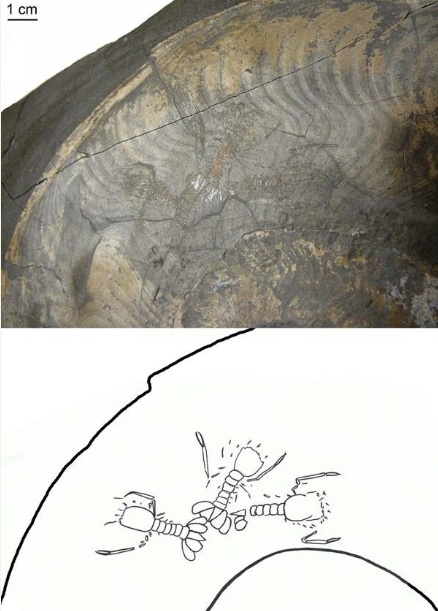
在德国的侏罗纪Posidonia页岩的一块镰菊石化石内,René Fraaije博士透过菊石半透明的、薄的壳(periostracum)观察到其内的镰龙虾。菊石壳体内的龙虾呈尾部互相对着的状态,并组成一个圈形,Klompmaker说,为何这些龙虾会呈现这样的保存状态目前仍不清楚,它们可能曾在这里暂时藏身以躲避天敌,或是一直栖居于此。
3:奇思妙想
引颈就戮型(参见小传18)
菊石和蛸类的近亲关系、许多中生代菊石有侧面的重叠或前部孔腹缘的投影(Arkell1957)表明具有壳分泌能力的外套膜可以延伸到壳口之外、异形菊石Ptychocerus显微结构的研究也表明一些外套膜能够远远超出开口边缘(Doguzhaeva&Mutvei 1989)。
综合以上信息,Jacobs, D.K. & Landman, N.H. 在1993年的文章中推测菊石具有蛸式游泳方式,部分外套膜暴露在壳外,如随便p的图所示:

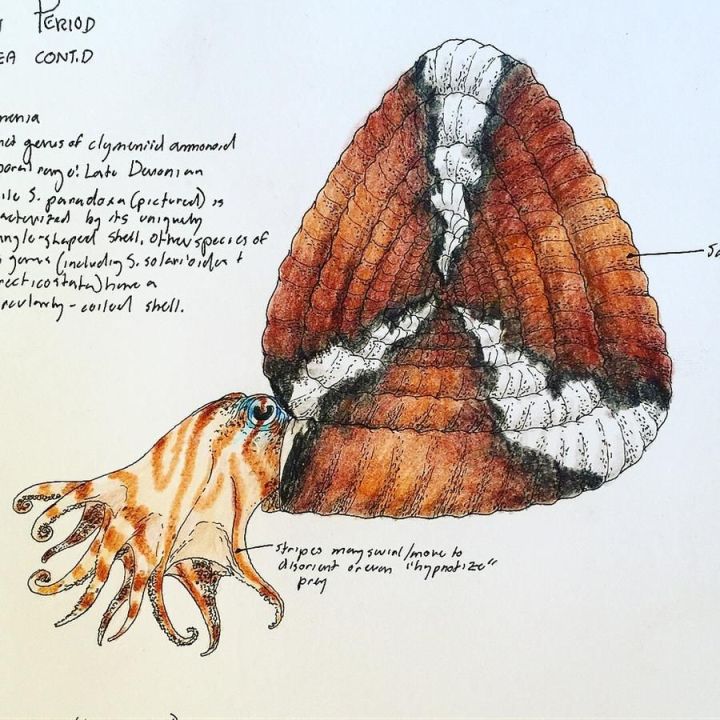
想当海兔型
螺旋菊石成体的螺旋部分变得很小,成了可有可无的装饰;在重建中,异形菊石的身体通常比鹦鹉螺大得多,与章鱼非常相似再加上1993年的文章中推测菊石具有蛸式游泳方式,部分外套膜暴露在壳外。
因此,螺旋菊石可以复原成这样(谁都没见过就是可以为所欲为)

无法形容型





最后,附上在大连贝壳博物馆照的菊石:
(去看菊石还是要贝壳馆,普通地质博物馆里并不多)

















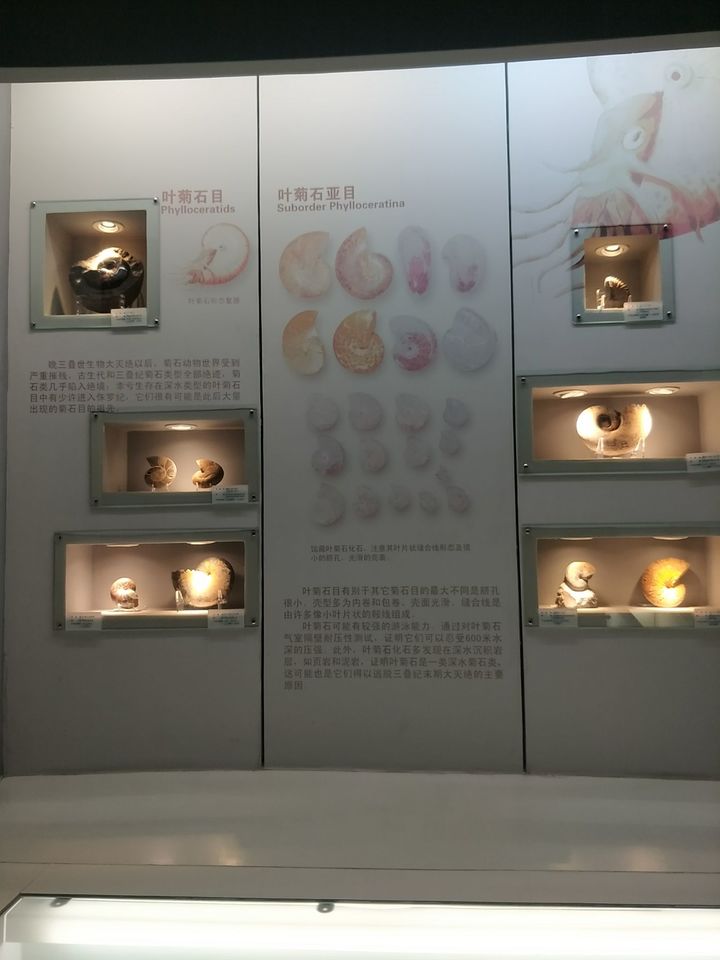



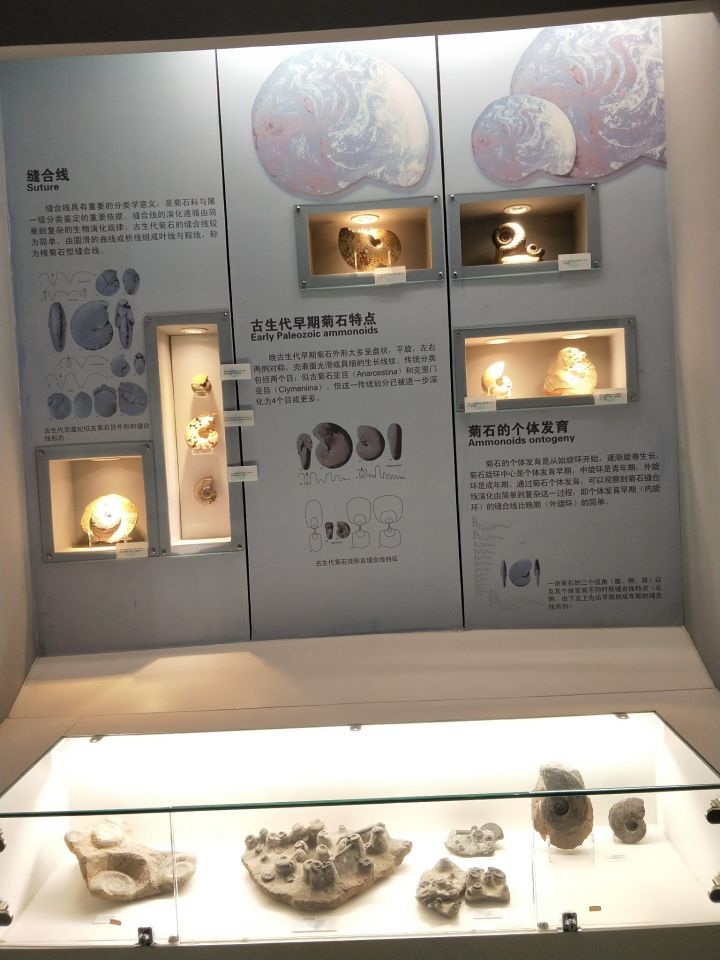


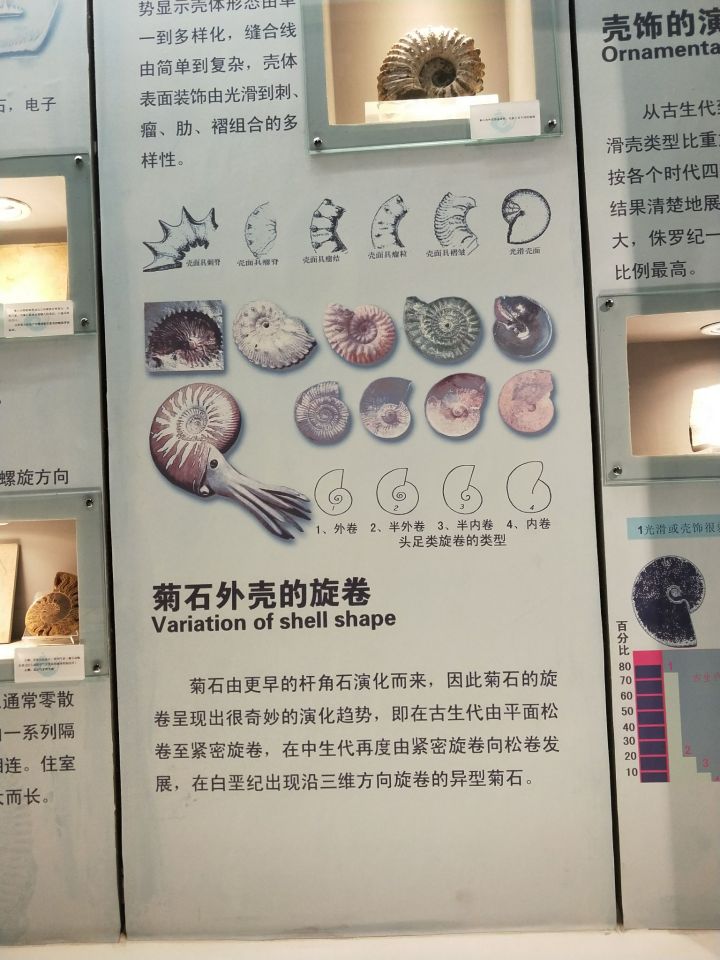

然后是自然博物馆:
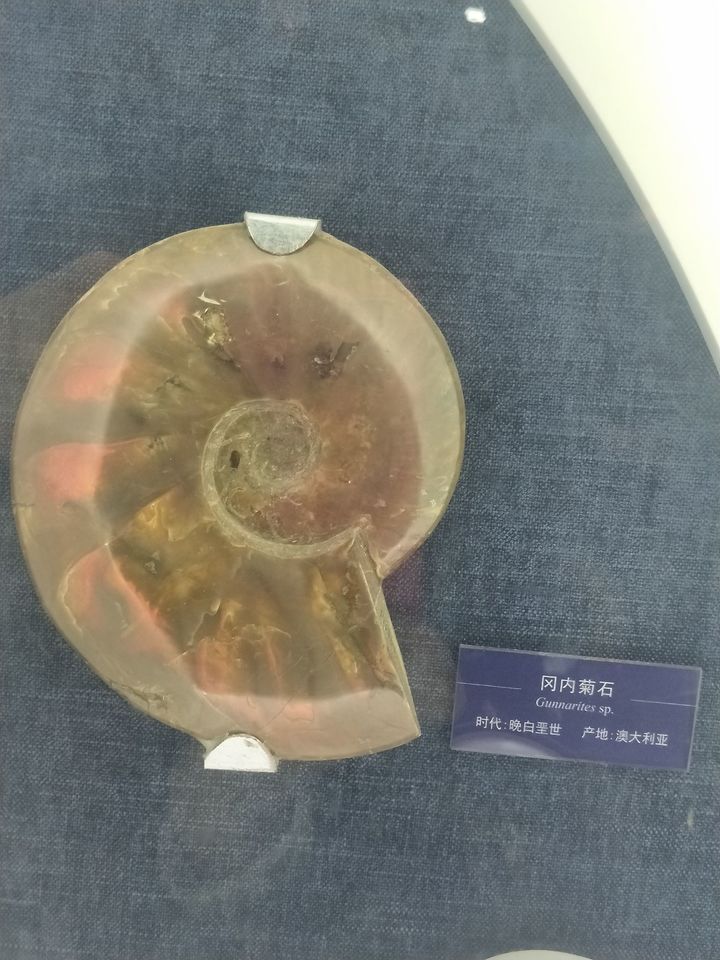
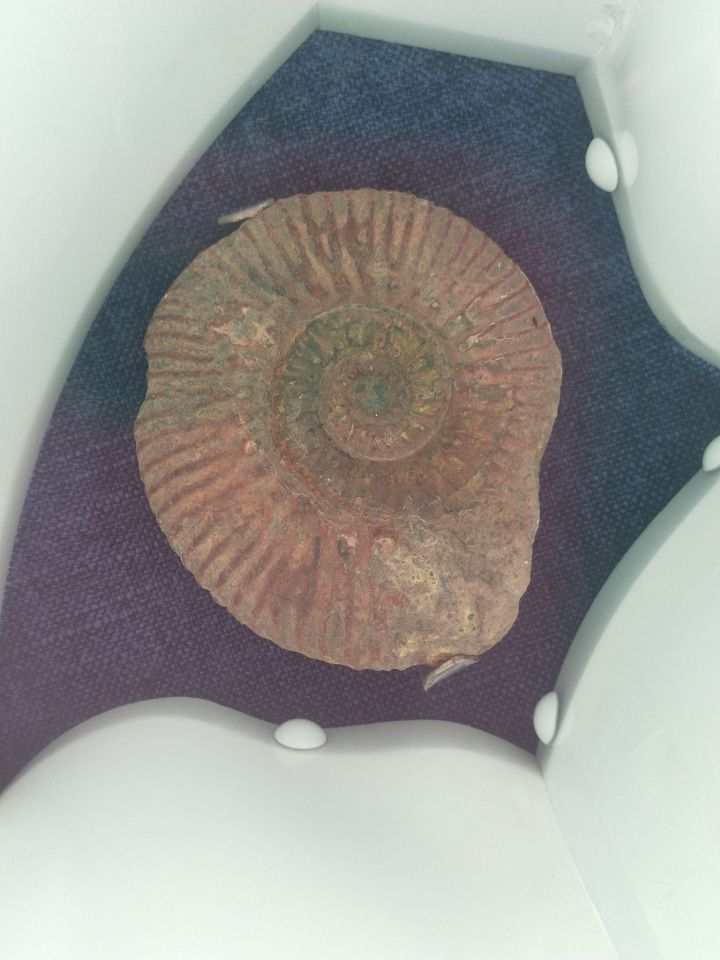






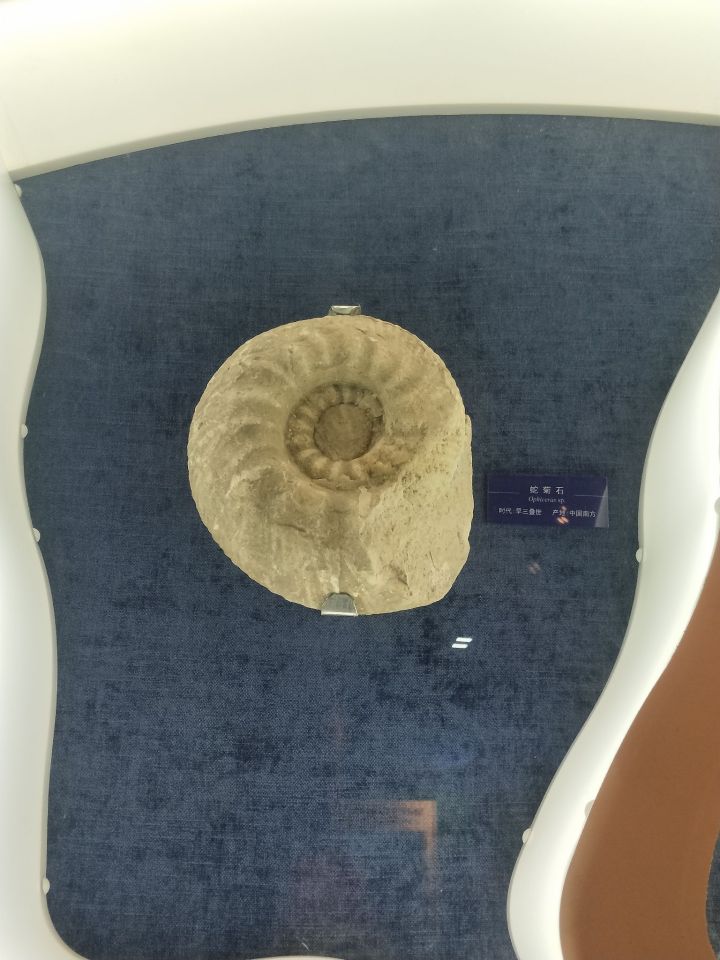








参考文献:
很多
(实际上:弄完忘了保存,再找找不到了...(* ̄0 ̄)ノ)


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









