





研究的重要性:热带森林受到人类活动的威胁,这些活动导致了森林砍伐和退化。然而,多种土地利用和土地覆盖的转变正在热带景观中发生,我们不知道这些转变的速度和对生态系统的影响有何不同。我们表明,在巴西亚马逊河流域,砍伐森林用来放牧是最普遍和影响最大的转变,尽管其他不太普遍的转变也造成了生物多样性和碳储量的减少,并改变了土壤性质。在我们研究的所有生态系统属性中,生物多样性受所有土地利用和土地覆盖转变的影响最大。我们表明,考虑多种转变和生态系统特性对于理解热带森林景观的现状和未来非常重要。
摘要:人类活动对热带森林的生物多样性和生态系统服务构成了重大威胁。尽管对森林砍伐的影响已经进行了充分的研究,但在热带地貌中出现了多种土地利用和土地覆盖的转变(LULCTs),我们不知道LULCTs在其速率或对关键生态系统成分的影响方面有何不同。在此,我们根据从巴西亚马逊地区310个地点收集的18个变量,量化了18个LULCTs对三个生态系统组成部分(生物多样性、碳和土壤)的影响。在所有LULCTs中,生物多样性是受影响最大的生态系统组成部分,其次是碳储量,但变化的幅度在LULCTs和个别变量之间有很大差异。将森林砍伐为牧场是最普遍和影响最大的转变,但我们也发现其他具有高影响,但普遍性较低的LULCTs(例如,从森林到农业)。我们的研究表明,考虑多个生态系统组成部分和LULCTs对于理解人类活动在热带景观中的后果非常重要。
相关工作
热带森林拥有所有陆地生物多样性的三分之二(1),占陆地生产力和蒸散量的三分之一(2),并储存了一半的陆地碳(3,4)。尽管热带森林具有全球重要性,但它们正受到人类活动的严重影响(5)。砍伐森林是变化的一个主要驱动力--在过去的40年里,超过1亿公顷的原始热带森林被转化为农业和造林业(6,7)。许多剩余的原始森林也在退化。2000年至2005年期间,至少有20%的热带森林被选择性采伐(8),同时其他人为因素和极端干旱正在增加森林火灾,20世纪90年代每年有5400万公顷被烧毁(9)。甚至被砍伐的地貌也在发生变化;农业废弃是次生林重新生长的主要驱动力,现在这些森林至少占全球所有热带森林的一半(10),包括亚马逊地区28%的被砍伐土地(11)。其他地区正在经历农业集约化,牧场被转化为耕地(12,13)。因此,许多热带景观现在是由无林土地使用地、再生次生林和不同程度退化的原始森林组成的镶嵌形(14)。
一些研究已经量化了包括亚马逊在内的热带森林地区不同的土地利用和土地覆盖过渡(LULCTs)(例如,参考文献15和16)。例如,我们知道,森林转为牧场和原始森林的退化是亚马逊的主要LULCTs (15, 16),而与森林砍伐相比,次生林的恢复仍然非常有限(11)。此外,人类改造的热带景观的价值已被详细研究,包括地上碳库存(17)、土壤状况(18)、植被结构(19)和生物多样性(20)。生态学文献显示,转换转变,例如砍伐森林,预计会对森林生态系统造成最大的影响(21),但当原始森林的退化大规模发生时,对生物多样性的危害也会与砍伐森林一样大(22)。其他研究讨论了生物多样性和碳在再生次生林中的恢复速度,例如,表明多达80%的原始森林树种可以存在于40年的亚马逊次生林中(23),而一项元分析表明,即使在一个世纪之后,植物物种多样性也不会恢复到未受干扰的热带森林水平(24)。
尽管科学界一直在为热带森林地区的LULCTs建立坚实的知识基础,但由于四个关键原因,现有的研究并没有完全揭示景观现状的变化。首先,仍然缺乏不同类型的LULCTs(包括森林砍伐、再生和森林退化)的比率汇编,这阻碍了对LULCT动态的全面看法,无法对所有转变进行定量比较。第二,特定区域的研究通常集中在一或两个生态系统的组成部分(24-26),或与未受干扰的森林基线进行比较,而不是评估所有的转变(23,27)。第三,统合分析/荟萃分析集中在 1)单一的生态系统成分(18),2)单一的 LULCT 类型(28),或 3)将所有变化与未受干扰的基线进行比较,而没有探索人类改造的土地利用之间的转变(21)。最后,到目前为止,生态变化还没有与土地覆盖转变的流行程度(例如,以年率衡量)联系起来。以流行程度和对生态系统属性的影响来量化转变,是了解整个地貌变化的相对重要性的关键一步,可以为决策者提供基于证据的情景,以决定如何更好地保护热带森林的生物多样性和生态系统服务并从中受益。
在此,我们量化了巴西亚马逊地区18种LULCTs的流行程度和生态影响,以对主要LULCTs的相对风险进行综合评估。我们重点研究了以下问题。1)LULCTs的速度是多少?2) LULCTs对不同的生态系统属性(如生物多样性、碳和土壤)有什么影响?3)哪些转变对生态系统造成的变化幅度最大?生态学数据包括在巴西亚马逊两个不同地区的310个地点采样的18个变量,我们将其归入反映政策或管理杠杆的三个生态维度:生物多样性(维管植物( 注:具有木质部和韧皮部的植物)、鸟类和三个无脊椎动物组)、碳库和土壤属性。这些生态系统组成部分在七个土地使用或土地覆盖等级中进行了调查:未受干扰的原始森林、砍伐过的原始森林、砍伐及烧毁的原始森林、古老的次生林(荒废后>20年)、年轻的次生林(≤20年)、牧场,和机械化农业。然后,根据2006年至2014年(29)或2006年至2019年(15)期间估计的LULCT年速率,评估18种LULCT对生态系统变量的影响,从而了解巴西亚马逊地区土地利用和土地覆盖变化的规模和范围。
结果
LULCTs的速率。根据2006年至2019年的土地利用变化图(15,30)和2006年至2014年的森林退化图(29),LULCTs的最高速率是从牧场到年轻次生林(17,780±2,840 km2y-1),其次是从年轻次生林和未受干扰的原始森林转变到牧场(分别为12,834±1,657和5,981±2,711 km2y-1;图1)。此外,2006年至2014年期间,4,877±1,218 km2y-1的未受干扰的原始森林被砍伐(图1)。另外两个转变超过了2000平方公里-1y:年轻的次生林到古老的次生林(2752±536平方公里1)和牧场到农业(3291±1263平方公里-1y;图1)。事实上,牧场向农业的转变率是反之的三倍(农业向牧场:1004±286 km2y1)。原始森林以及年轻和古老的次生林向农业的转化率分别比向牧场的转化率低67、26和63倍(图1和SI附录,图S2A)。
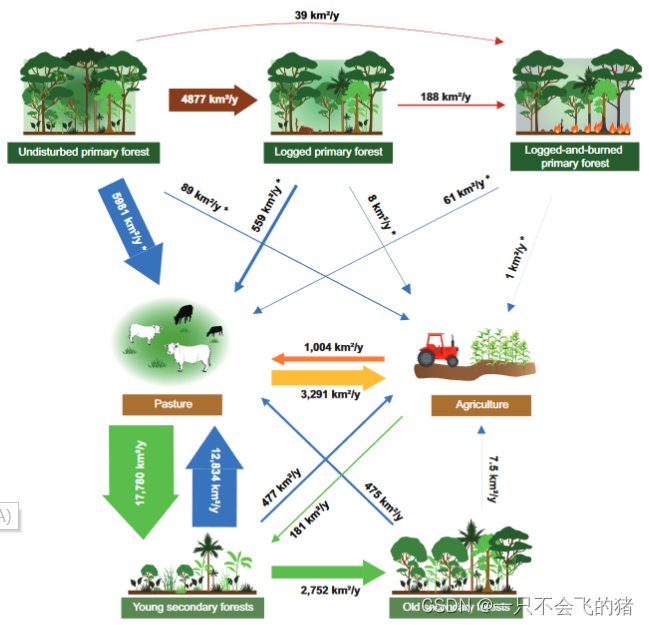
图1. 巴西亚马逊地区的LULCT速率。年平均LULCT速率是根据土地利用变化图[2006至2019年(15)]和森林退化图[2006至2014年(29)]计算和估计的(*)。原始森林即从未被砍伐过,而次生林是再生林。年轻的次生林是指树龄<20年,古老的次生林是指树龄≥20年。农业包括多年生作物和临时作物。箭头的宽度与转变的年平均速率成正比。
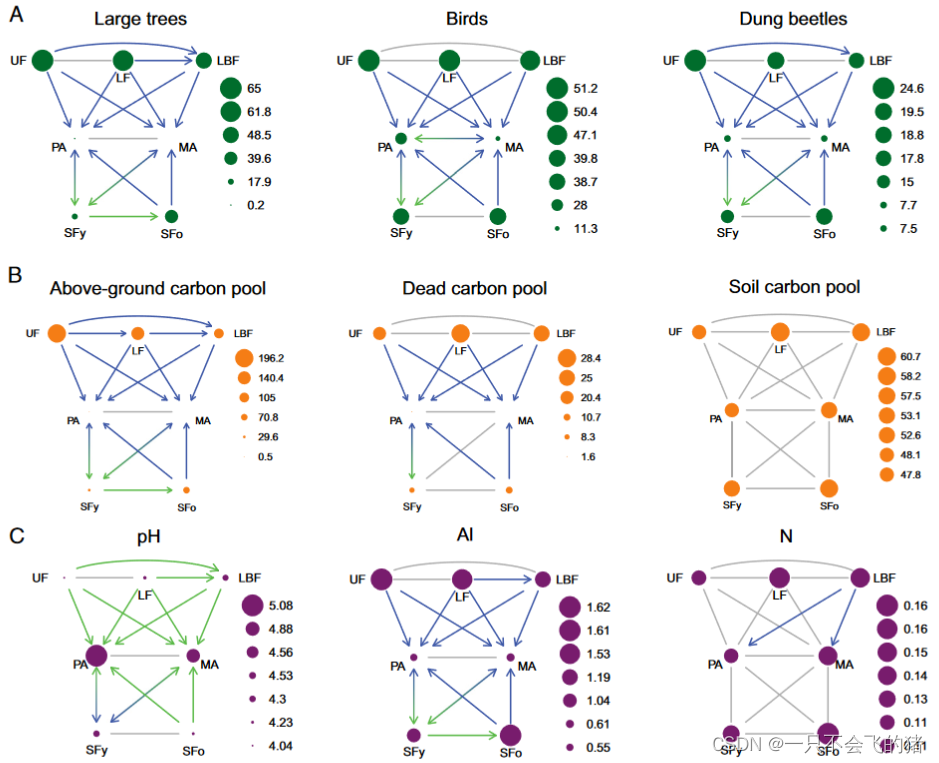
图2. 18种LULCTs对巴西亚马逊地区(A)生物多样性、(B)碳库和(C)土壤属性的影响。生物多样性以物种丰富度表示,碳库以碳的Mgha-1表示,Al(铝)的单位是mmolcdm3,N(氮)的单位是土壤质量百分比。箭头表示转变及其影响,其中灰色表示没有影响,绿色是明显增加,蓝色是明显减少。符号的大小代表了基于21个未受干扰的原始森林(UF)、68个伐木的原始森林(LF)、65个伐木烧毁的原始森林(LBF)、72个牧场(PA)、26个机械化农田(MA)、33个年轻次生林(SFy,树龄<20年)和25个古老次生林(SFo,树龄≥20年)的平均数,分布在巴西东部亚马逊的两个地区。
LULCTs对生物多样性、碳和土壤的影响。随着原始森林和次生林向牧场或机械化农业的转化,几乎所有生物多样性群体的物种丰富度(即分类学多样性,以下简称多样性)都下降了18%至100%(每个变量的平均值和变化百分比分别见SI附录的表S3和S4)。值得注意的例外是蚂蚁和兰花蜂的多样性,在未受干扰的原始森林和古老的次生林转变为牧场后没有变化,而兰花蜂的多样性在任何原始森林或古老的次生林转换为机械化农业后没有变化(图2A和SI附录,图S2)。从未受干扰的原始森林转变到伐木且烧毁的原始森林,大树和蜣螂的多样性分别减少了25%和27%。当伐木原始森林转变到伐木焚烧的原始森林时,大树、小树和藤本植物的多样性下降(分别下降21%、17%和21%),当牧场转为机械化农业时,蚂蚁和鸟类的多样性分别下降30%和59%(图2A和SI附录,图S2和表S4)。最后,大树的多样性增加了一倍,小树的多样性增加了55%,以应对从幼龄次生林到老龄次生林的转变(图2A和SI附录,图S2)。对森林物种的单独分析产生的结果基本上反映了总物种丰富度的模式(SI附录,图S3和S4)。
当原始森林或次生林被转化为牧场或机械化农业时,地上碳库、废弃物和枯木碳库减少了74%到100%(图2B和SI附录,图S5和表S4)。从未受干扰的原始森林过渡到伐木或伐木且烧毁的原始森林,地上碳库减少了28%到46%;从伐木到伐木且烧毁的原始森林,地上碳库减少了25%(图2B)。只有地上碳库随着幼龄次生林向老龄次生林的过渡而增加(增加2.4倍;图2B和SI附录,图S5)。土壤碳库在七个土地利用和土地覆盖类别中没有任何区别。
在评估的七种土壤属性中,从原始森林或次生林到牧场和机械化农业的大部分转变中,土壤pH值和磷、钾、钙+镁的浓度增加了6%到4.2倍,从未受干扰到伐木或伐木且烧毁的原始森林,土壤pH值增加了6%到2.8倍(图2C和SI附录,图S6和表S4)。从伐木到伐木焚烧的原始森林,土壤pH值增加5%,钙+镁增加50%。相比之下,当原始森林或次生林转化为牧场或农业时,土壤铝浓度下降了41%至66%(图2C),但在从年轻次生林到古老次生林的转变中,土壤铝浓度增加了54%(图2C)。土壤钾随着从牧场到机械化农业的转变增加了25%(图2C和SI附录,图S6)。土壤氮的百分比只随着伐木烧毁的森林向牧场和机械化农业的转化而变化(图2C)。最后,土壤钠的浓度在七个土地利用和土地覆盖类别中没有任何差异(SI附录,图S6)。
LULCTs的生态系统变化的程度。根据转变的影响大小的中位数排序,生物多样性的最大下降发生在从原始森林和次生林类向机械化农业的过渡中,其次是森林类向牧场的过渡(图3b)。牧场和机械化农业之间的双向转变对生物多样性的影响最小,但对鸟类和蚂蚁多样性的个别影响是中等的(图3)。其次,对生物多样性变化幅度第二小的是在原始森林类型之间的转变:1)从未受干扰的原始森林到伐木或伐木且烧毁的原始森林,以及2)伐木和伐木后烧毁的原始森林之间(图3b)。同样,从原始森林类型和古老的次生林向牧场或机械化农业的转变对碳库和土壤属性的影响最大,其次是牧场和机械化农业向年轻次生林的转变(图3)。牧场和机械化农业之间的转变对碳库和土壤属性的影响最小,其次是原始森林类别之间的过渡和从年轻次生林到古老次生林的转变(图3)。从未受干扰的原始森林向机械化农业的转变对生物多样性和土壤生态系统成分的影响最大(图3),而地上碳库在未受干扰的原始森林向机械化农业和牧草的转化中记录了两个最高的个体影响大小(图3)。
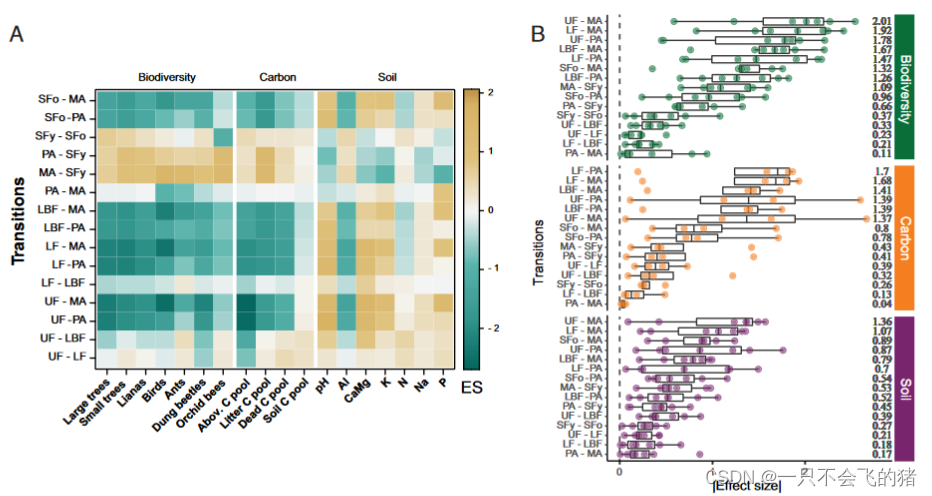
图3. 巴西亚马逊地区LULCTs对生态系统的标准化影响大小。(A) 15个LULCTs(其中三个是双向的,只显示一次。PA-MA、PA-SFy和MA-SFy)对7个生物多样性群体、4个碳库和7个土壤属性的标准化影响大小(ES)。(B)巴西亚马逊地区15种LULCTs对生物多样性、碳库和土壤性质影响大小的绝对值(点)的箱线图。B中的数字,右边的数字是标准化影响大小的中位数,转换为绝对值。标准化效应大小来自混合效应模型,该模型分析了LULCTs对生态系统组成部分内单个变量的影响。请注意,在B中,转换是根据其影响大小的中位数从大到小排序的,因此在不同的生态系统组成部分中是不同的。MA-SFy、PA-SFy和PA-MA的转换是双向的。
将影响与转变率联系起来。LULCTs发生的速度与它们对生物多样性(r=0.49,P=0.03)和土壤性质(r=0.49,P=0.03;图4)的影响成反比,但我们发现这些变量与碳生态系统成分之间没有关联(P=0.08;图4)。然而,使用年转变率中位数和影响大小中位数对每个生态系统组成部分的转变率和影响进行分类,我们发现,无论生态系统组成部分如何,从未受干扰或伐木的原始森林到牧场的转变总是被列为高影响、高比率。大多数向机械化农业的转变被归类为高影响、低速率,因为这些转变通常具有最大的影响,但发生的年速率较低(例如,未受干扰的原始森林的转化)。牧场和机械化农业之间的双向转变,以及从年轻次生林到古老次生林和从年轻次生林到牧场的转变,对于三个生态系统组成部分来说,都被归类为低影响、高速度。尽管从未受干扰的原始森林向伐木森林转变的年率很高,但所有生态系统组成部分的影响中位数大小都很低,因此该转变也被归类为低影响、高速率。最后,从未受干扰的原始森林和砍伐森林到被砍伐和烧毁的原始森林的转变被归类为低影响、低速率(图4)。
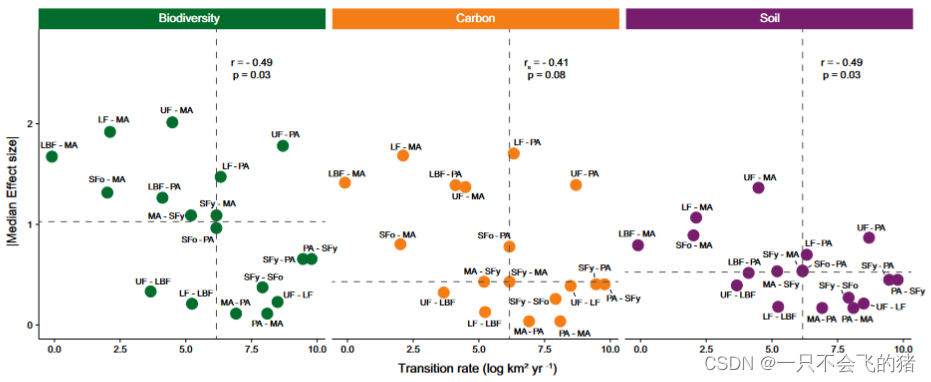
图 4. 巴西亚马逊地区LULCT速率与它们对生态系统成分的影响之间的关系。在每个生态系统成分(生物多样性、碳或土壤)中,虚线代表中位数,分隔出四类转变:高影响、低速率(左上),高影响、高速率(右上),低影响、高速率(右下),以及低影响、低速率(左下)。年平均速率(km2y1)进行了对数转换,影响由评估18种LULCTs对7个生物多样性群体、4个碳库和7个土壤属性的影响的模型得到的标准化影响大小(转换为绝对值)中位数表示。
讨论
我们的研究探讨了LULCTs对热带生态系统的普遍性和影响。我们确定了LULCT速率与对生物多样性和土壤属性的影响之间的负相关关系,这表明最大的影响正在影响巴西亚马逊的较小区域。然而,通过识别LULCTs对生态系统成分的相对风险,我们也强调了那些既具有高影响又普遍的转变,以及那些具有高影响但发生频率较低的转变。通过这样做,我们提供了一种方法,可以应用于为景观规模的保护或恢复工作提供比迄今为止更具体的指导。为养牛而砍伐森林仍然是亚马逊地区最广泛的人为活动(31,32),而从未受干扰或被砍伐的原始森林向牧场的过渡是我们唯一列为高影响、高速度的。从原始森林和次生林转变到牧场的面积达24,000平方公里y-1,导致地上碳库完全丧失,鸟类和无脊椎动物多样性的主要损失在18-69%之间。这些多样性的影响还包括物种替代,即森林物种被开放地区的物种所替代,因此,如果考虑到物种构成,变化的幅度可能会更大(20,22)。从原始森林和次生林向机械化农业的转变影响最大,但发生的频率较低。然而,机械化农业只在亚马逊的一些特定地区盛行,那里为谷物生产和出口建立的生产系统占主导地位(33)。虽然原始森林的退化像砍伐森林一样普遍(29,34),但从未受干扰的原始森林过渡到伐木或伐木及烧毁的原始森林,其影响要小得多,突出了保护受干扰森林不被进一步清除的重要性(例如,参考文献35)。然而,这些转变降低了森林对生物多样性、碳和土壤的价值,显示了限制退化和毁林的行动的相关性,特别是当退化发生在大空间范围内时,退化的累积影响可能与毁林一样有害(22)。最后,两个最普遍的转变是从牧场到年轻的次生林,以及相反的转变(分别为17,780和12,834平方公里y1),反映了广泛牧场的低效率,在巴西,牧场的生产力只有潜在生产力的30%(36)。
生物多样性和碳库是许多转变最敏感的生态系统组成部分。这令人担忧,因为有许多例子表明,生物多样性的丧失和更替会降低关键生态系统服务的效率和稳定性(37,38),并破坏我们为气候和生物多样性危机开发基于自然的解决方案的能力(39)。此外,LULCTs 对植被的负面影响不仅直接影响到碳储量、森林结构和植物多样性,而且对许多其他分类群的多样性也有连带影响(40)。兰花蜜蜂值得特别一提,因为它们对大多数LULCTs不敏感。兰花蜜蜂的觅食距离很长,即使在破碎的地貌中也能分散开来(41),使它们对土地利用和土地覆盖的变化有相对的抵抗力。此外,它们会被一种化学诱饵诱捕,吸引他们跨越非常远的距离。然而,也有可能这两个研究地区的景观中森林覆盖率高(>50%),有助于缓冲LULCTs的影响,但景观中较低水平的生境数量可能会连带产生更大的影响(42)。此外,尽管兰花蜜蜂的多样性对许多LULCTs没有反应,但相关的授粉服务是否也不受影响,还有待观察。
许多土壤特性--包括土壤碳、氮和钠--似乎对森林砍伐不敏感,而土壤pH值、钙+镁、钾和磷增加,可能反映了化肥和石灰的应用或牲畜的投入(43,44)。尽管在人类改造的生境中,土壤pH值和养分浓度的增加有可能在再生过程中促进竞争性快速生长树种的建立[例如,在恢复性种植中(45)],但几乎没有证据表明这种情况在高土地使用强度下发生:在以前被牧场和机械化农业占据的地方,次生林的恢复速率往往比较低(46)。此外,营养丰富的农业系统似乎很可能不是目前正在再生的次生林,因为这些生产性土地不容易被遗弃。此外,鉴于目前对热带地区土壤过程的了解有限,尚不清楚一些观察到的土壤理化性质的变化如何通过土壤微生物群级联并影响土壤功能,特别是在长期内(47)。
将古老的次生林(≥20年)转化为牧场或农业,对生态系统成分的影响与原始森林的转化相似。尽管次生林不能替代未受干扰的原始森林,但我们的研究结果强化了古老次生林的高潜在价值,特别是那些在保留50%以上森林覆盖率的地区自然再生的次生林(48);这种森林具有生物多样性(23)、碳储存(24)和土壤条件,可能与原始森林类似(27)。因此,我们的研究结果为立法保护古老的次生林不被清除提供了强有力的经验支持[例如,在帕拉州(23,49)],特别是考虑到最近亚马逊地区次生林清除的加速(50)。不幸的是,从年轻的次生林转变到古老的次生林的速度很低,这表明次生林在重新生长的头20年内就被清除了。因此,保护这些年轻的次生林并使其成熟,也可以为生物多样性和缓解气候变化带来巨大的好处。
牧场和农业之间的双向转变很广泛,影响到4,295平方公里y-1,而且在2006年至2019年期间,这一比率一直在增加(SI附录,图S7)。在一些地区,由于避免进一步砍伐森林的激励措施,从牧场到机械化农业的过渡正在取代2000年代初发生的从森林到机械化农业的转变(36)。尽管牧业向机械化农业转变对生物多样性、碳和土壤的影响最小,但我们只分析了其对陆地生态系统的影响。水生生态系统可能特别容易受到涉及农业集约化的转变的影响(51)。溪流中的磷和氮负荷、浊度和温度都会因农业集约化而增加,并可能导致对生物多样性和生态系统服务的一连串影响(51)。
我们的研究结果强调了某些生态系统成分对不同LULCTs反应的巨大差异(图3B),其中碳库之间的差异尤其明显。例如,尽管地面碳库随着未受干扰的原始森林转化为牧场而减少了99%,但土壤碳库却没有变化。根据LULCT的不同,不同生物多样性群体的反应也有明显的不同。例如,所有植被群体的多样性都随着伐木原生林向伐木焚烧原生林的转变而减少,但没有任何动物群体受到影响。可以想象,开放栖息地物种的殖民化弥补了森林物种的损失(52),而对森林物种进行更严格的定义(例如,森林专家)可能会揭示出火灾和伐木的更强影响(22)。除了我们研究中包括的不同变量的内在本质外,这种高变化性也可能是由于转变发生的强度、频率和时间的不同,以及景观的具体环境[如周围森林覆盖的程度(53,54)]。
对政策和实践的影响。确定生态系统中最重要的转变和敏感属性是对热带森林的现状和可能的未来有更全面了解的第一步。我们认为我们的结果对政策有五个重要的影响。首先,它们提供了令人信服的证据,表明砍伐原始森林以创造牧场仍然是巴西亚马逊地区最重要的高影响、高速度的土地使用转变。因此,我们强调了打击亚马逊地区森林砍伐的至关重要性,该地区的森林砍伐率在2020年达到了十年来的最高水平,并且在未来可能还会增加(55)。第二,通过将LULCTs的普遍性和影响联系起来,我们的分析有助于确定地方和区域的近期和长期行动,突出应该优先处理的转变。例如,限制我们确定的高影响、高速度的转变需要在生物群落尺度上最直接的做出努力,但遏制高影响、低速度的转变也可以在地方尺度上优先处理(33)。第三,我们的研究结果表明,改善生态状况还需要采取一些措施,而不仅仅是追踪和减少森林砍伐,因为涉及二次再生和原始森林退化的转变会影响一系列生物多样性、碳和土壤指标。综合景观管理将需要改进对土地覆盖转变的监测和报告,以支持激励积极转变的新政策,限制最具破坏性的LULCTs的发生,或减轻其生态影响。鉴于使用农用化学品对淡水生态系统的潜在风险,从牧场到耕地的转变值得特别关注,我们在这项研究中没有记录。第四,我们表明,生物多样性是受土地使用和土地覆盖变化影响最大的生态系统组成部分。虽然世界上对热带森林景观的兴趣主要是围绕着碳排放,但我们表明,生物多样性需要更多的关注,并希望在即将举行的生物多样性缔约方大会第十五届会议和其他全球会议上强调这一点。最后,我们表明,理解人类对热带森林的影响需要超越对生物多样性状况的单一成分和二元评估--通过这样做,我们希望我们对更全面的转变的评估为景观管理开辟新的机会。
材料和方法
LULCTs的速率。为了估计巴西亚马逊地区LULCTs的比率,我们使用了两个来源的数据。首先,对于从原始森林和次生林到牧场或农业,以及从农业和牧场到次生林的转变,我们使用了基于MapBiomas Collection 5数据集(1985年至2019年;mapbiomas.org)的土地利用变化图(15)。我们应用Smith等人(30)所述的方法,将MapBiomas模式分为四类(森林、牧场、耕地和其他),并使用变化检测算法,将森林类进一步分为原始森林和次生森林(30)。我们进一步将次生林分为两类。如果它们的树龄≤20年,则为 "年轻";如果它们的树龄>20年,则为 "古老"。由于MapBiomas的时间序列始于1985年,任何在此日期之前开始生长的次生林都包括在原始森林类别中。使用20年的阈值,我们在2005年之后才开始检测古老的次生林。第二,对于从未受干扰的原始森林到被砍伐或被砍伐烧毁的原始森林,以及从这些原始森林类型到被砍伐的土地利用类型的转变,我们使用了1992年至2014年巴西亚马逊的森林退化图(29),其中数字对象分析框架和数字光谱分析与实地数据进行了校准,以绘制由伐木、火灾、边缘效应和隔离造成的森林干扰(29)。基于这两个数据集,我们获得了17个LULCTs的信息(SI附录,图S2),并计算了它们在2006年至2014年(森林退化图)和2006年至2019年(土地利用变化图)期间的年平均速率(km2y1)。我们重点关注2006年以后的LULCTs,因为这一年是亚马逊土地利用变化方面社会政治背景的一个突破点(56),但转变速率变化的完整时间序列见SI附录,图S7和S8。由于第一个数据集没有区分未受干扰的和已退化的原始森林,第二个数据集没有区分原始森林被砍伐的土地用途,我们估算了这些年速率。为此,我们用所有原始森林向牧场和农业的转化率(0.985转化为牧场,0.015转化为农业)来计算从未受干扰的原始森林、伐木的原始森林和伐木烧毁的原始森林向牧场和农业过渡的年率,基于它们的总体砍伐率(SI附录,图S2B)。例如,如果我们根据退化图(29)计算出6,070 km2y1的未受干扰的原始森林砍伐率,我们估计有5,978.95 km2y1被转换为牧场,91.05 km2y1被转换为农业。因此,我们评估了18个LULCT率,用于将过渡期的普遍性与影响联系起来(将影响与转变率联系起来)。
LULCTs对生物多样性、碳和土壤的影响。研究地点和采样设计。我们从位于巴西帕拉州的亚马逊东部的两个地区的310个地点收集数据:Santarem、Belterra和Mojuı-dos-Campos市(以下简称STM)和Paragominas市(以下简称PGM)。我们所有的研究地点都位于最初被陆地菲尔梅森林覆盖的地方。在21世纪初,这两个地区都经历了森林砍伐的增加,这与机械化农业的扩张有关。这些地区目前的景观由未受干扰和受干扰的原始森林、农业用地和次生林组成(48)。
我们将这两个地区分成三阶或四阶排水流域(每个流域约5,000公顷),然后在每个地区选择18个代表森林覆盖梯度的流域。在每个流域,我们设置了8至12个站点(横断面或平面),它们之间至少相隔1.5公里,以避免空间自相关。动物群的采样是沿着300米的样带进行的,而植物群和土壤属性是在250×10米的地块(0.25公顷;SI附录,图S1A)内调查的。关于场地和采样设计的更多细节,见Gardner等人(48)。
我们的310个地点分布在七个不同的土地利用和土地覆盖类型(SI附录,表1):未受干扰的原始森林;被砍伐的原始森林;被砍伐和烧毁的原始森林;古老的次生林(>20年);年轻的次生林(1至20年);牧场;和机械化农业。不同类型的土地利用和次生林的年龄是通过实地评估,结合卫星图像的时间序列分析(1988年至2010年至2011年之间)进行分类。完整的细节,见Gardner等人(48)。
生态学变量。我们对18个生态变量进行采样,以代表三个不同的生态系统成分:生物多样性、碳和土壤。我们根据2010年和2011年期间对七个组的调查来评估生物多样性。植物的多样性根据茎的大小或功能生长类型分为三组。1)大树,2)小树,和3)藤本植物,这些都是在0.25公顷地块的植被调查中确定的(SI附录,图S1B)。为了捕捉动物群的多样性,4)鸟类和5)蜣螂在每个样带相隔150米的三个点上采样(SI附录,图S1B),6)蚂蚁在相隔50米的六个点上采样,7)兰花蜂在相隔50米的五个点上采样。关于每个分类群的采样技术的更多细节可以在SI附录,方法和Gardner等人(48)找到。除兰花蜂外,所有变量都在两个地区取样,兰花蜂只在PGM取样。我们计算了每个采样点每个生物多样性组的总物种丰富度和的森林物种丰富度(那些在原始森林地点至少出现一次的物种)。
我们遵循政府间气候变化专门委员会的标准(57),将生态系统碳储量分为:1)地上碳库;2)枯木库;3)废弃物碳库;4)土壤碳库。为了获得碳储量,我们按照Berenguer等人(17)的方法估算生物量,计算的全部细节见SI附录《方法》。在植被调查期间(2010年和2011年),我们测量了所有活体树木、棕榈树和藤本植物的个体,以及胸径≥10厘米的立枯树和棕榈树。我们首先计算每个采样个体的生物量(SI附录,方法),然后将所有活树、棕榈和藤本植物的数值相加,得到每个采样点的地上生物量总量。地面碳库被假定为代表每个地点活体生物量总量的 50%(57)。我们还根据每个研究地点的五个0.01公顷的子地块(5×20米)的测量结果,计算出粗大的木质碎屑(至少在一个端部直径≥10厘米)的生物量。然后,我们将立枯树和棕榈树的生物量与粗木屑的生物量相加,得到枯木碳库,占每个地点总枯木生物量的50%[全部方法和所用公式见Berenguer等人(17)]。我们通过对在另外五个面积为2×5meach的子地块中发现的所有碎片进行加权估计细木质碎片(至少在一个端部直径为2至10cm)的生物量(SI附录,图S1B),而垃圾生物量是从每个地块的10个0.25m2的四分区中收集的样品估计的,这些样品后来被烘干至恒定重量。废弃物碳库被计算为细小的木质碎屑和落叶的碳含量。土壤碳库是在0-30厘米深度测量的碳储量(详情见下文)。所有数值都被转换为每公顷的碳储量。
为了确定土壤碳储量和土壤属性,我们在每个采样站的5个采样点(相隔50米)以10厘米为单位采集土壤样品(SI附录,图S1B),并将其批量化,以获得每个地点和深度的一个复合样品。土壤采样是在2010年和2011年的碳储量调查期间进行的。我们用每个深度增量的两个完整的土壤核来测量体积密度,这些核是在每个地块的中心用一个体积测量环收集的。随后使用干燥的土壤按照标准方法(58)对pH值、氮、磷、钾、钙+镁、钠和铝进行分析,并按照Ellertl和Bettany(59)对土壤碳含量和化学性质进行分析。然后,通过将每一层的碳含量乘以该层的厚度(10厘米)和土壤的体积密度来计算土壤碳储量,并对其进行调整,以比较不同土地使用之间的土壤当量。Gardner等人(48)和Durigan等人(60)给出了关于土壤采样和分析的进一步细节。
最后,为了在分析LULCTs时考虑土壤类型和地形的混杂影响,我们使用密度计方法获得每个土壤样品的粘土含量(61),并使用数字高程模型获得每个站点周围100米缓冲区的平均海拔和坡度(48)。
数据分析。所有的数据分析都在R4.1.0版本中进行(62),R代码和数据(63)可在Zenodo(64)获得。根据我们在七种土地利用和土地覆盖类型中的18个生态变量,我们评估了18个LULCTs,其中三个是双向的(即从牧场到机械化农业,以及从牧场和机械化农业到年轻次生林)。为此,我们首先使用 "尺度 "函数对所有响应变量和解释变量进行了中心化(减去每个值的变量平均值)和尺度化(每个值除以变量的SD)。然后,我们使用lme4软件包(65)中的'lmer'函数运行线性混合模型(LMMs),将每个生态变量作为响应变量,将土地利用和土地覆盖类别(有七个级别的分类变量)作为解释变量,将粘土含量、海拔和坡度作为协变量。此外,我们还在所有模型中加入了流域ID和区域(STM和PGM)作为随机因素,但兰花蜂物种丰富度模型除外,该模型只包括流域ID,因为该类群只在PGM区域采样。为了评估哪个LULCT对每个响应变量有影响,我们使用multcomp软件包(66)中的函数'glht'对土地利用和土地覆盖类别的平均值进行了多重比较(15个比较,每个单向转换和三个双向转换中的每一个)。为了限制I型错误,对比采用单步检验,考虑到检验统计之间的相关性,并在考虑多重对比的情况下调整P值。此外,我们还使用'confint'函数计算了每个对比(即每个过渡)的同步CI,考虑了多重比较。在P<0.05时,我们认为过渡期对反应变量有显著影响,并使用β系数来获得关系的方向。所有的模型都经过检查,以确保 [...]
对于有限的模型子集,Q-Q图显示了对正常分布的小偏离以及土地利用和土地覆盖类别之间的不平等变异。这种情况发生在一些模型中,其中一些响应类别(如牧场和机械化农业中的树种丰富度)中的零值占多数,不可避免地导致这些土地利用类别中的方差非常低。尽管与正态分布的偏差很小,但我们还是用量化的广义加性模型(qGAMs)验证了所有模型,以测试我们的结果对正态分布的偏差和不等式的敏感性(68)。我们遵循LMM分析的相同步骤,并对中位数(0.5分位数)进行多重比较以计算效应大小和P值。然后我们进行相关测试,将从qGAMs获得的效应大小与用LMMs获得的效应大小进行比较。这一验证表明,我们的研究结果对偏离正常分布和不平等方差的情况不敏感。首先,所有重要模型的结果都高度相似(r = 0.99),验证了我们的LMMs。其次,在270个配对比较中(18个变量×15个过渡),258个(95.5%)保持在显著性阈值(0.05)的同一侧。在改变其重要性的转换中(SI附录,表S7),大多数(3.7%,n = 10)显示了使用qGAM变得重要的转换,只有0.75%(n = 2)使用qGAM变得不重要了。这些显著性变化没有明显的模式,它们既发生在符合所有良好拟合假设的模型中,也发生在出现偏离正态分布和不等值的模型中。验证分析的完整描述和结果见SI附录,方法和图S12。
生态系统上变化的转变幅度。
影响大小分析。为了了解生物多样性、碳和土壤的变化程度,我们使用了标准化的影响大小。标准化效应大小和它们的CI是由上述手段的多重比较产生的系数和95%的CI。由于我们对所有变量进行了中心化和缩放,LMMs中土地利用和土地覆盖类别的平均估计值的差异可以解释为每个转变(每个比较)对每个变量的效应大小。这些效应大小在原来具有不同单位的不同变量之间也是可以比较的(例如,生态系统组成部分之间)。每个变量和每个LULCT的效应大小(和CI)见SI附录,表S6。
为了比较LULCTs,我们使用了影响大小的绝对值(即把负值转化为正值),因为在这里我们感兴趣的是变化的幅度,而不是其方向。我们将所有变量的影响大小的绝对值按生态系统成分分开的每个转变绘制出来。然后,我们按照影响大小的中位数对转变进行排序--分别对生物多样性、碳和土壤的七个、四个和七个变量的效应的中位数。我们认为每个转变的中位影响大小代表其对生态系统组成部分的 "平均 "效应。
将影响与转变速率联系起来。为了更好地理解LULCTs的生态影响,我们通过运行相关测试('cor.test'函数),分析了每个LULCT的年率与每个生态系统成分(生物多样性、碳和土壤)的中位效应大小之间的关系。对于生物多样性和土壤生态系统成分,我们进行了Pearson相关测试,对于碳成分,我们进行了Spearman等级相关测试,因为碳的中位效应大小不遵循正常分布。此外,我们将数据分为四个等级,代表每个LULCT的影响和速率增加的规模:1)低影响,低速率;2)低影响,高速率;3)高影响,低速率;4)高影响,高速率。为了实现这一目标,我们计算了每个生态系统组成部分的年度LULCT率和效应大小的中位数,并将中位数作为区分低和高比率或影响的分界点。


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









