






摘要:小脑后部在人类的社会和情感学习中发挥着关键作用。有三个系统及其相关神经网络为小脑的这一功能提供支持:作为扩展感觉运动整合网络一部分的生物动作观察系统;由心理化网络支撑,用于理解他人心理和情感状态的心理化系统;以及支持核心情感(不)愉悦和唤醒过程的边缘网络。在这篇综述中,我将阐述这些系统和网络如何借助在小脑后部与大脑新皮层之间起始和终止的功能性互惠连接,来支持社会和情感学习。有人提出假设,认为小脑后部的一个主要功能是识别和编码事件的时间序列,这或许有助于优化社会和情感学习并使其自动化。我将探讨运用神经影像学和非侵入性刺激的相关研究,这些研究为小脑时间序列功能的假设提供了越来越多的证据,同时也会探讨关于小脑后部在这些社会和情感过程中作用的其他潜在功能解释。
1. 引言
人类的社会思维和情感体验在很大程度上依赖于我们对他人高层次思想和情感的理解,这使我们能够更好地预测他人即将做出的行为,并控制自己对这些行为的反应。理解他人的心理状态,即心理理论(或心智化),是人类高度发达的一种能力,它需要对一个人的意图、情感、信念、偏好和特质进行抽象或高层次的推断。这些心理状态可以从人际交流、互联网上的公开信息以及文学、戏剧、电影、音乐或歌剧等众多艺术形式中直接获取。例如,当一个人朝另一个人走去时,被接近的人可能会想,这个走近的人的潜在动机是什么(意图),他对自己有什么想法或期望(信念),从他的面部表情来看对自己有什么感觉(情感),以及这个人给自己留下了什么印象(人格特质)。
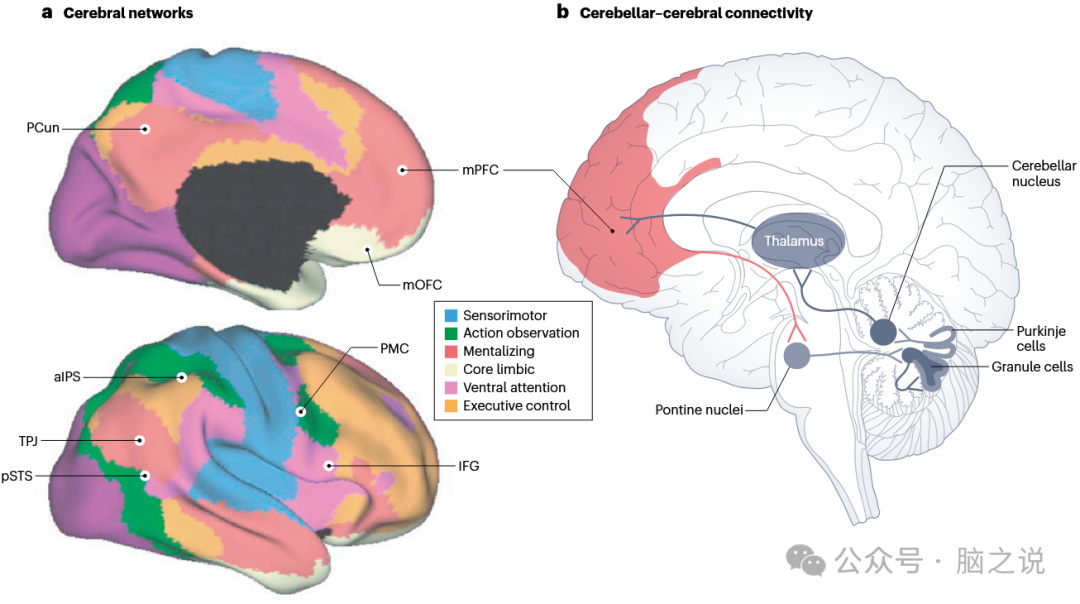
图1 支持社会和情感学习的大脑网络以及小脑与大脑之间的互惠连接
社会和情感学习也得益于对他人生物运动和情感表达的低层次动作观察。动作观察是指通过观察他人运动的方向和终点状态,对他人的目标进行直接的低层次推断。例如,当有人把一个物体伸向某个人而不是其他人时,人们会不假思索地明白,这个被递出的物体的预期接收者是谁。同样地,情感理解(或情感识别)是指通过身体和面部表情,自发地推断出愤怒、厌恶、恐惧、快乐、悲伤和惊讶等基本情感体验。
这些过程中的每一个都由大脑新皮层中不同的皮质回路或网络支持(图 1a)。人类高层次的社会和情感心智化能力由心智化网络支持,这是默认模式网络的主要功能之一。心智化网络有助于识别特定的心理想法,以及他人和自己的情感。这使得人们能够从社会行为、交流、自身的本体感觉,或他人的面部和身体表情(如意图、情感或信念)中理解其意义。关于人类生物运动的低层次社会推断由动作观察网络支持(以前称为镜像网络,是扩展感觉运动整合网络的一部分)。通常认为,动作观察网络通过将观察到的他人动作与自身类似动作的记忆表征相匹配,帮助人们识别他人动作的目标(例如收到礼物时)。低层次的情感过程,包括自我的情感感知和观察他人面部表情时的情感识别,由核心边缘网络支持,该网络代表内脏运动身体状态。核心边缘网络的一个功能是产生愉悦或不愉悦以及唤醒的核心情感基调。虽然理解他人的情感是情感心智化的一个主要组成部分(见证并同情他人的情感或痛苦),但在这篇综述中,我主要探讨理解他人情感的社会层面。
近 200 年来,小脑一直被认为对运动协调至关重要,但小脑也通过与大脑新皮层的互惠连接参与社会和情感学习(图 1b)。除了对小脑疾病患者非运动过程的临床观察外,实验观察也表明,小脑有助于非运动的认知和情感过程。此外,对小脑神经功能和连接性研究的大规模元分析也强调了小脑在社会过程中的关键作用,这些研究使用了传统的社会动作观察和心智化任务。
在这篇综述中,我将介绍小脑在人们理解他人社会和情感状态方面的重要作用。首先,我将讨论小脑图谱,以及支持从低层次到高层次社会和情感学习的动作观察、心智化和核心边缘小脑网络。我还将探讨这些小脑网络如何通过动作观察、心智化和核心边缘网络的小脑与大脑区域之间独特的互惠连接来实现其功能。接下来,我将描述小脑识别时间序列的假设,该假设如何解释小脑在社会动作观察、社会心智化和情感理解中的功能,以及它与其他候选机制的对比。在这篇综述中,我主要关注人类的社会和情感学习,因此主要涵盖使用神经影像学和神经刺激技术的人类研究。不过,我偶尔也会引用动物研究,以说明同源过程,并讨论在人类研究中无法深入探讨的主题,如解剖连接和神经回路。
2. 大脑和小脑的功能网络
小脑与服务于不同功能领域的多种大脑新皮层相互连接。重要的是,这些互惠的解剖连接通过多个突触在小脑和大脑新皮层之间传递神经信号,大脑-小脑连接通过脑桥,小脑 - 大脑连接通过丘脑(图 1b),并且对于不同的功能领域,如运动和社会行为,这些连接大致不同。连接的功能特异性互惠结构至关重要,因为这意味着神经信号在两个结构中相似的功能区域之间传递,因此小脑的不同功能区域接收来自大脑新皮层相同功能区域的输入,并向其提供反馈。
与早期认为这些互惠解剖连接由大量平行的多突触一对一轴突连接(称为闭环)的观点相反,最近的动物研究揭示了新皮层 - 小脑回路中明显的发散和单向(开环)连接性。例如,来自脑桥的轴突由多个分支组成,这些分支通常连接到双侧半球的几个小脑小叶。然而,在学习过程中,小脑区域的特定区域显示出越来越多的互惠连接结合。有人提出,包含广泛发散和选择性汇聚连接的新皮层 - 小脑回路,以任务依赖的方式灵活组织互惠连接的功能网络。这表明,最初在新皮层和小脑区域中不同的活动模式,在反复配对后可以收敛到一组共享的神经活动,从而实现神经信号更灵活的功能整合。
人类小脑中互惠连接的地形排列决定了其各种功能,并有助于其复杂的功能网络结构(图 2)。此外,神经信号在同一小脑网络内沿着通过小脑蚓部跨越半球边界的传递纤维进行交换。这导致小脑的功能结构在两个半球上大致对称。
接下来,我将深入探讨小脑在社会和情感功能中的关键作用。首先,我将介绍通过小脑图谱和元分析发现的小脑主要网络,并回顾这些网络与社会和情感过程的关系。其次,我将讨论支持小脑社会和情感功能的互惠连接,主要基于人类研究,因为对他人行为、思想和情感的元表征理解是人类高度发达的能力,在动物中差异很大,包括其潜在的小脑功能和结构。
2.1 神经网络的小脑图谱
在过去十年中,通过识别与大脑新皮层中相同的典型网络,人们对小脑功能的理解有了很大进展,这些网络包括支持社会和情感学习的动作观察、心智化和核心边缘网络。这些主要网络在小脑中的位置和功能可以通过功能图谱可视化(图 2a,b)。虽然最初的图谱是基于静息态功能磁共振成像(fMRI)数据(在扫描仪下静思;图 2a),但最近的功能图谱是从广泛的心理任务中获得的 fMRI 数据中提取的(图 2b),或者是静息态和任务态 fMRI 数据的结合。静息态和任务态功能图谱之间的大量一致性表明,心智化网络位于小脑Crus 1、Crus 2 和蚓部小叶 IX,而其他一些网络的一致性则较低(图 2a,b)。尽管如此,功能性脑图谱,尤其是 Buckner 等人的静息态小脑图谱,如今在研究中经常被用于识别小脑中潜在的功能回路。
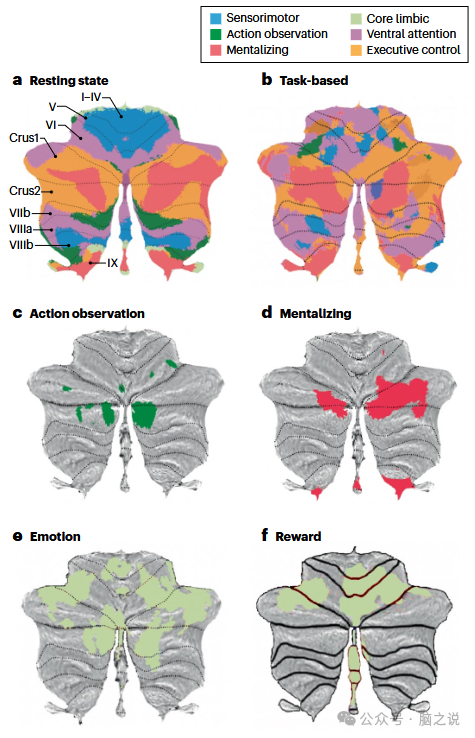
图2 小脑支持许多不同的过程,包括社会和情感学习
然而,所有的功能性脑图谱都有一个缺点。通过采用 “赢者通吃” 的计算方法,它们为最主要的功能提供了证据,但未明确重要的次要过程,鉴于前面提到的小脑互惠连接的斑块状排列,这些次要过程很可能存在。另一个潜在的缺点是个体差异,在绘制人类小脑心理活动与运动活动的图谱时,这种差异可能更大。此外,反复接触后的学习和自动化过程可能会改变小脑募集的程度和定位。因此,我现在转向近期关于社会和情感过程的神经影像学元分析证据,这些证据更多地依赖于特定任务,较少依赖于个体先前的接触,并且不使用 “赢者通吃” 的方法,从而更好地涵盖了重要的次要功能。这些元分析并不总是证实小脑图谱中所代表的网络的解剖位置。
2.1.1 社会动作观察
通过观察生物运动来理解社会(社会动作观察)主要由 Crus 2 支持,Crus 2 包含心智化网络的下部(图 2c)。这一元分析观察结果与作者最初的假设相矛盾,即小脑中的扩展感觉运动整合网络像在新皮层中一样,支持社会动作观察以识别观察到的人类运动中隐含的目标。在原始出版物中未使用的小脑二维平面表示(扁平图谱)清楚地表明,这种解释是错误的(图 2c)。值得注意的是,早期关于社会动作观察的元分析在这个问题上无法提供太多信息,因为它们要么存在方法学上的局限性,如将社会动作观察和社会动作执行混淆,要么报告小脑在单独的社会动作观察中参与度极低。
2.1.2 社会心智化
理解他人的心理状态主要由小脑后部的心智化网络稳健地支持,该网络涵盖 Crus 1 和 Crus 2 的主要部分(图 2d)。在这里,元分析和功能图谱数据趋于一致(图 2a、b 和 d)。在啮齿动物中,心智化的前身是通过回忆先前的相遇来识别熟悉和陌生的同类。对小鼠蚓部小叶 VII(部分与人类 Crus 区域同源)的神经活动进行光遗传抑制,破坏了小鼠对熟悉同类的社会偏好,但对非社会物体的偏好没有影响。这表明,跨物种的同源心智化功能可能通过相似的小脑区域进行处理。需要注意的是,在小脑层面,社会动作观察和心智化过程似乎在心智化网络(Crus 2)中共享一个共同的解剖基质。相比之下,大脑皮层在解剖上主要有不同的区域来实现这两种功能,如动作观察中的前顶内沟和前运动皮层,与心智化中的楔前叶和内侧前额叶皮层,尽管有些区域非常接近,可能部分重叠,如后上颞沟与颞顶联合区。
2.1.3 情感
小脑支持情感和奖励过程。最近关于情感和奖励的元分析(图 2e,f)显示,小叶 VI 的腹侧注意网络有大面积激活,这可能有助于将注意力和行为导向显著的情感或奖励刺激。此外,这些元分析还揭示了小叶 I - V 的感觉运动网络中有小的激活区域,这可能反映了接近或避开这些(不)愉快刺激的行动策略。情感,而非奖励,还会募集 Crus 1 和 Crus 2 中的大部分心智化网络,这表明小脑情感网络中存在大量高层次的概念成分,涉及对自身感觉或他人(面部和身体)情感反应的理解。重要的是,情感和奖励反应也占据了小叶 IX 蚓部区域的很大一部分。多个网络的募集与 Lindquist 和 Barrett 提出的情感的模块化网络或建构主义方法一致,该方法认为情感是由更基本的心理过程组合而成的心理状态。这种情感构建得到广泛分布的功能网络的支持,如我在本综述中重点关注的核心边缘、动作观察和心智化网络。
Ferrari 及其同事进行的一系列重要研究,通过对执行情感识别任务的人类参与者的小脑后部进行非侵入性经颅磁刺激(TMS),进一步支持了小脑在情感方面的因果作用。TMS 会在底层脑组织中诱导电流,导致神经元去极化并产生动作电位。在这些研究中,Ferrari 等人应用 TMS 提供短抑制脉冲,干扰参与者识别或区分情感面部表情、身体姿势,或隐性识别情感面孔的尝试。因此,令人惊讶的是,没有已发表的小脑功能图谱显示出有力证据,证明存在参与小脑低层次核心情感处理的边缘网络(图 2a),而在大脑新皮层的功能图谱中可以清楚地观察到核心边缘网络(图 1a)。
总之,小脑功能图谱、元分析和非侵入性刺激研究为小脑参与社会和情感功能提供了有力的实证支持。小脑图谱和元分析证据在网络表征位置上缺乏明确的一致性(除了小脑后部 Crus 区域的心智化网络),这是一个重要问题(潜在解释见 “神经网络的小脑图谱” 小节)。然而,元分析关于低层次动作观察和情感网络在小脑表征的发现(与小脑图谱不同)表明,它们在 Crus 区域有共同的表征,而情感在(旁)蚓部也有独特的表征。此外,研究表明,心智化与情感在小脑有共同的表征,这是相当合理的(它们都基于对一个人心理状态的高层次社会推断以及相关的大脑区域)。相比之下,动作观察与心智化有共同的表征则不太合理。在确定了代表每个网络的小脑区域后,我现在转而探讨它们在情感和社会学习过程中如何与大脑区域相互作用。
2.2 心智化过程中小脑与大脑功能相连
小脑通过小脑和大脑结构之间的持续交互作用来调节社会和情感过程。如前所述,这种交互作用的特点是功能性互惠连接,这些连接在执行相似功能的网络结构之间传输神经信号,因此小脑中的功能区域接收来自大脑新皮层相同功能区域的输入,并向其发送信号。
在解剖层面,Sokolov 等人确定了一条可能促进小脑与后上颞沟(pSTS)之间通信的通路。在功能层面,动态因果模型(DCM),一种用于确定脑区之间功能连接的先进方法,已被用于识别许多支持大脑与小脑之间神经信号交换的功能连接。DCM 分析还揭示了社会动作观察、社会心智化和情感识别过程中的互惠连接(图 3)。接下来,我将更详细地讨论这些功能性互惠连接。
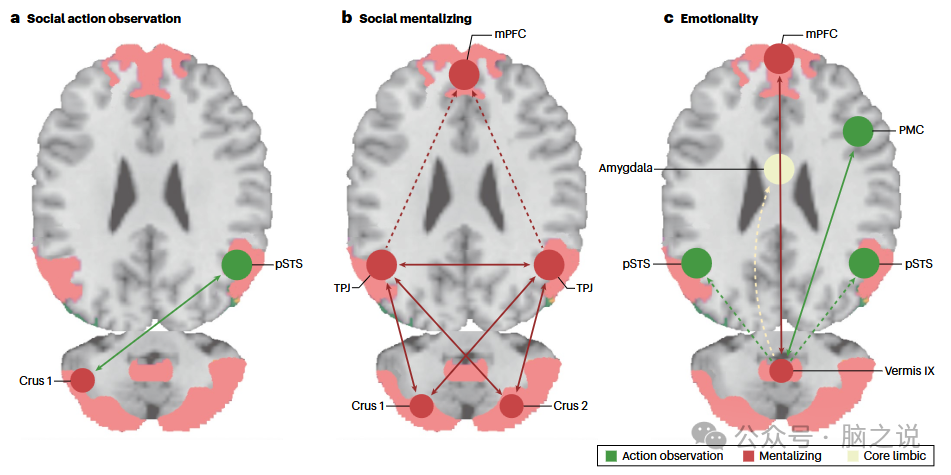
图3 社会动作观察、心智化和情感过程中,小脑与大脑皮层之间的互惠功能连接
2.2.1 社会动作观察
DCM 分析显示,在一项视觉动作观察任务中,当点光源显示描绘人类运动时(任务细节见图 4a),左半球 Crus 1 和 pSTS 之间存在互惠功能连接(图 3a)。在其中一项研究中,pSTS 与下额叶 gyrus(IFG)进一步交换神经信号,形成了一个小脑 - 大脑动作观察网络,其中 pSTS 似乎充当连接小脑与大脑新皮层的中间枢纽。
2.2.2 社会心智化
对一项图片排序任务(要求参与者生成社会动作的时间顺序;任务细节见图 4d)中进行信念心智化时的大脑活动进行 DCM 分析,揭示了一个小脑 - 大脑心智化网络。研究发现,Crus 1 和 Crus 2 通过同侧或对侧半球与双侧颞顶联合区(TPJ)相互连接;双侧 TPJ 相互连接,并依次单向连接到内侧前额叶皮层(mPFC)(图 3b)。此外,小脑中的两个半球通过小脑 - 大脑通路间接连接。因此,TPJ 似乎充当小脑与大脑新皮层之间的中间枢纽,类似于动作观察网络中的 pSTS。
在涉及特质推断的社会心智化任务中,观察到了直接连接小脑后部和 mPFC 的互惠功能连接,以及直接连接左右半球 Crus 2 的连接。一系列 DCM 分析在结合来自不同任务背景下涉及特质和信念推断的多项研究数据时,始终发现这种连接模式(任务细节见图 4b - e)。人类连接组项目的一项大规模功能连接分析(使用与 DCM 类似但功能较弱的方法)也证实了这些连接发现。
2.2.3 情感
对参与者编码负面和正面情感图片时的大脑活动进行 DCM 分析。结果显示,小叶 IX 的蚓部与 mPFC(心智化网络的一部分)和前运动皮层(PMC,动作观察网络的一部分)形成互惠连接(图 3c)。此外,作者观察到从蚓部 IX 到双侧 pSTS(动作观察网络)和杏仁核(核心边缘网络)的单向通路。这种连接模式(图 3c)与 Lindquist 和 Barrett 提出的情感的模块化网络(或建构主义)方法一致,表明涉及核心边缘网络(杏仁核)、心智化网络(蚓部 IX、mPFC)和扩展感觉运动整合网络(pSTS 和 PMC,可能是由于参考文献69 中用作刺激的人类动作,如运动或攻击行为)的区域参与。
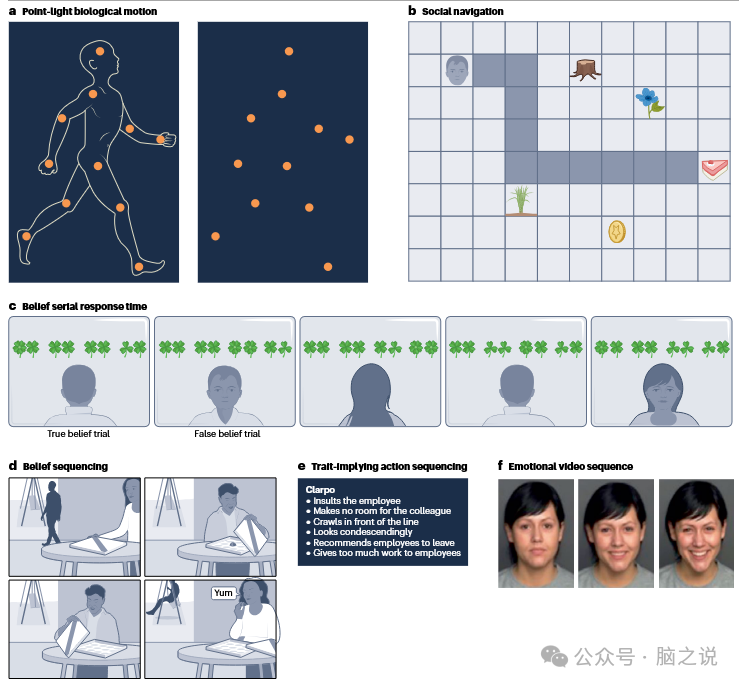
图4 在社会动作观察、心智化和情感过程中,测试小脑序列功能的任务
综上所述,使用 DCM 在社会动作观察、社会心智化和情感理解过程中,已经描绘出了不同的小脑 - 大脑功能性互惠连接(图 3)。相互连接的区域与从小脑元分析(图 2c - f)和大脑网络(图 1)得出的功能一致。然而,也观察到了差异(如零连接、单向连接或双向连接),这可能部分归因于模型中分析的关键区域不同,或者 DCM 的方法学局限性,如模型可识别性(由传递相似信息的通路之间的竞争导致的多重共线性),或对复杂模型而非简单模型的惩罚(需要更多研究来探讨 DCM 结果对这些参数的敏感性)。此外,人类社会动作观察和情感的连接性研究(图3a,c)与啮齿动物社会偏好和灵活行为的连接性研究之间存在显著相似性。这种连接相似性表明,从老鼠到人类的进化过程中,一些反映小脑后部和 mPFC 之间低层次社会和情感神经信号交换的基本连接在很大程度上得以保留。
一个重要的观察结果是,在小脑中,社会和情感信息似乎主要由心智化网络的 Crus 区域处理,但这些功能在该区域的共同表征机制尚未得到充分理解。这或许意味着,社会和情感过程均由一个相似的、专门的小脑系统所支持,该系统整合了动作观察和心智化(对社会行为以及心理和情感状态的处理)。啮齿动物研究也表明,Crus 区域是社会行为的共同小脑基础(方框 1)。小脑与新皮层相连的解剖回路呈现出分散和汇聚的斑块状特征,这或许可以解释大脑中不同的功能网络,如何通过该特征在小脑的同一 Crus 区域汇聚,进而支持社会和情感行为,使这些多元过程能够更灵活地同步运作。要深入了解人类后 Crus 区域中动作观察和心智化系统的共同与独特功能成分及通路,还需要开展更多研究。
总之,在社会和情感学习过程中,小脑会显著激活,且小脑的处理过程与大脑之间的神经信号相互交换紧密相关,这种交换发生在动作观察、心智化和情感认知的特定小脑 - 大脑连接中。这表明小脑在社会和情感学习中犹如一个核心枢纽。那么,与大脑相比,它的具体功能作用是什么呢?下一部分将探讨这个关键问题。
3. 序列识别是小脑在社会和情感过程中的主要功能
传统观点认为,小脑对运动协调至关重要。例如,在指鼻测试中,神经正常的人能够闭眼快速且准确地将手指指向鼻子,而小脑功能障碍的人则无法做到。该测试结果表明,这一运动学习过程需要小脑参与,它能逐渐将由视觉反馈控制的肢体运动转变为无需视觉反馈的自动运动。实际上,根据 Ito 的理论,运动皮层充当肢体运动的控制器,经过反复练习和学习后,小脑内的内部模型会取代运动皮层的作用,使肢体运动更加自动化、快速、精确和流畅,即便没有视觉反馈和意识思考也能如此。关键在于,通过内部模型实现的运动自动化,使小脑能够对计划或观察到的运动及其本体感受和外部结果进行预测,从而快速察觉运动中的错误或环境变化,并及时、恰当地调整正在进行的运动。这种快速的运动调整能力,是神经正常的人即便在对方手指移动时,也能有效触摸到对方手指的基础。
鉴于小脑也参与诸如社会和情感学习等心理活动,运动领域中关于内部模型实现自动化和快速纠错的理论,也被延伸应用到了这些非运动领域。小脑的表面积较大(约为大脑新皮层的 80% ,包含了人类大脑中约 80% 的神经元),这为小脑提供了充足的计算资源,使其能够为运动和心理活动编码大量的内部模型。
为描述小脑通过内部模型实现的主要协调功能,人们提出了多个概念,如计时机制、错误检测与自动化、预测与准备、动态状态监测或信息的感觉预处理。在大脑功能的预测框架内,受 Ito 早期研究的支持,Leggio 和 Molinari 提出了序列检测假设,即小脑的核心功能是随时间检测运动或心理动作的时间序列(它们的连续顺序或时间顺序)。Molinari 和 Masciullo 指出,这种序列识别不仅支持大脑的预测能力,而且尽管涉及多个脑区,但小脑在序列识别中起着核心作用。根据序列检测假设,识别和编码(学习)运动和心理动作的时间序列,对于小脑实现各种运动和心理过程的自动化、微调以及在预测出现偏差时快速调整至关重要。
在某些领域,如计划或观察运动中,以毫秒精度编码序列的精确时间可能很重要,但在社会心智化等认知领域,其重要性相对较低。在社会领域,时间序列可在不同的时间尺度和层次上发挥作用,从肢体运动产生(动作)和观察(动作观察或情感识别)的毫秒级,到社会心智化或故事理解的秒级或分钟级,再到对课堂或工作中未来行为预期的月级。动作观察涉及时间序列,因为人类的生物运动需要动态变化和肢体位置的序列。像准备浪漫晚餐这样更为复杂的有意行为,涉及从较低层次的肢体运动(伸手拿刀)到较高层次的动作组合(准备甜点)的层次结构,每个层次都需要在不同的行为层面上遵循正确的时间序列。心智化同样需要编码关键动作的序列。例如,要理解游戏中一个主体的目标,人们通常需要跟随主体在轨迹中的行动步骤(包括转弯以避开障碍物和竞争目标),直到到达轨迹终点,进而推断出其预期目标(图 4b)。同样,要理解故事内容,就需要记住事件展开的时间顺序,以全面理解主人公的目标、兴趣以及争议或共识的原因。情感处理也需要对序列的认知。例如,面部表情从中性变为愤怒通常比从愤怒变为中性更具威胁性。在更高层次上,看似无端的攻击性拳击,通常比针对他人攻击行为的防御性拳击,更让人感觉具有攻击性。
接下来,我将重点介绍支持小脑在社会动作观察、心智化和情感处理中识别时间序列作用的实证研究。需要注意的是,为评估序列识别在以下研究中的作用,通常会将序列学习与包含静态刺激(无序列)的对照条件进行比较。
3.1 社会动作观察中的小脑序列学习
在比较人类肢体运动视频片段与包含无序列运动静态图片的对照条件,或不太严格的静态对照条件(注视点或休息;这些对照条件的结果没有明显差异)的研究中,对生物运动序列(动作观察)的观察需要小脑 Crus 区域的支持(图 2c)。Sokolov 及其同事最近的神经影像学研究,通过使用点光源显示描绘人类运动(图4a),证实了序列在生物运动动作观察中的作用,研究发现左半球 Crus 1 被激活。此外,左半球外侧小脑病变(包括 Crus 2)的患者,对由点光源显示呈现的身体运动的视觉敏感性存在缺陷,而小脑前蚓部病变对人类运动的视觉感知影响不大。一项使用经颅直流电刺激(tDCS)的非侵入性神经刺激研究,通过亚阈值调节神经静息电位,自发改变大脑活动和兴奋性,为小脑在观察动作序列中的因果作用提供了更确凿的证据。在这项研究中,Oldrati 等人向参与者展示描绘人类动作(抓取苹果)的视频片段,这些动作以明确的目标结束(自己吃或递给别人)。刺激小脑后部调节了参与者根据情境线索预测人类动作正确目的的能力,而对非社会形状的预测能力未受影响。
总之,有力的证据表明,小脑后部的动作观察回路支持对他人社会运动序列的识别。
3.2 社会心智化中的小脑序列学习
一系列新开发的带有序列成分的心智化任务,评估了序列在社会心智化中的作用。在这些研究中,参与者在推断他人心理状态的同时,生成或记忆暗示这些推断的动作的时间顺序。与非序列或非社会对照条件相比,生成或记忆这些序列激活了 Crus 区域。心智化推断涉及一系列从低层次到高层次的社会判断,包括社会目标导向导航(图 4b)、他人的信念(图 4c,d)、人格特质推断(图 4e)或对未来社会行动的预测。此外,在进行社会预测时,这些研究报告称,小脑心智化网络的另一个区域 —— 小叶 IX 也会额外激活。重要的是,这些任务均未涉及对人类生物运动的观察,以排除动作观察过程。相反,这些研究使用语言描述或简单的视觉刺激,如卡通形象,或从一个位置到另一个位置的简化移动,通常描绘没有肢体运动或特定面部表情的简化类人形象(比利时卡通人物蓝精灵)。
心理状态推断的时间序列通常是内隐学习的。例如,当一个人与他人打招呼时,他们通常会遵循并期望一系列特定的动作、想法和表情的时间顺序(如打招呼、微笑、被问及近况),这些在反复的社交接触后内隐学习,成为自动化的常规行为。这些内隐的心智化序列同样由Crus 区域支持。为评估内隐学习,参与者在不知情的情况下,以相同的重复顺序接受真实和错误信念试验(类似于经典的刺激 - 反应 - 时间任务;图 4c),研究发现重复的试验顺序会增加 Crus 区域的激活。偶尔以随机顺序呈现试验,会打乱自动化的顺序,这不仅会减慢反应速度,还会导致 Crus 区域强烈激活。这一结果支持了反复的心理推断序列可在后小脑内隐学习并自动化的观点。
先前的研究表明,打乱心智化推断的自动化顺序可能会减慢反应速度。那么,当隐含的社会推断而非序列不一致时会发生什么呢?一些社会违规行为包括:一个一直表现诚实的人突然开始说谎;一位电影明星在垃圾桶里找食物;一个人见面时给对方送不受欢迎的礼物;或某人闯入禁区。在这些情况下,为理解社会不一致性,需要调整最初根据行为对主人公做出的特质或刻板印象推断(这个人其实并不那么诚实,或者电影明星失业了)。尽管序列未被打断,但有证据表明,与常规和一致的行为相比,这类高层次的社会违规行为会导致 Crus 区域的激活增加。这表明,小脑能够识别不同层次的不一致性,无论是序列上的还是社会层面的,这可能是因为不同层次的不一致性都需要在社会互动中进行调整。这凸显了小脑在减少错误和准确预测后续行为方面的作用。这也得到了先前关于基于已知人物心理状态(特质、信念或偏好)预测未来社会行动的研究的支持,这些研究表明,当需要明确未来行动的时间序列时,适应性预测会募集后小脑(以及心智化网络的小叶 IX)。
关于小脑在社会序列识别中的因果作用的更多证据,来自信念序列研究。与神经正常的人相比,小脑功能障碍患者在图片排序任务中,仅能解决约一半的错误信念故事(图 4d),但在关于机械事件和社会常规的对照故事中,两者没有差异。Heleven 进行的一项非侵入性低频重复经颅磁刺激(TMS)神经刺激研究,在参与者执行相同的图片排序任务前,对其后小脑进行刺激。尽管低频TMS 通常会降低兴奋性,但该干预措施缩短了参与者在任务中生成社会和非社会事件正确时间顺序的时间。
综上所述,有充分的证据支持这一假设,即小脑的心智化网络与社会序列的学习和自动化,以及其中潜在的不一致性处理密切相关,这与小脑在时间学习中作用的理论模型一致。
3.3 情感观察中的小脑序列学习
研究情感中序列识别的一种方法,是比较呈现动态情感刺激(如现实生活中体验到的变化的面部表情)的视频片段与静态图片(类似于动作观察研究中,比较变化的肢体运动视频片段与静态照片)。然而,相关证据并不十分确凿。虽然一项元分析显示,动态情感刺激会导致更广泛的大脑激活,但只有少数研究直接比较了动态和静态情感面部表情,且没有研究发现小脑有额外激活。不过,一项非人类灵长类动物研究表明,在动态面部情感处理中,小脑的激活更强。此外,一些非侵入性神经刺激研究的证据表明,识别动态和静态情感面部表情可能涉及不同的过程。Malatesta 等人使用高频经颅随机噪声刺激(tRNS)刺激小脑后部,这种刺激通常会产生兴奋效应。在参与者观看从中性表情转变为快乐或悲伤的静态和动态面部表情时(图 4f),高频 tRNS 小脑刺激降低了识别静态悲伤面部表情的准确性,但提高了识别动态悲伤面部表情的准确性,而对转变为快乐面孔的情况没有影响。
总体而言,小脑在情感处理中进行序列识别的功能证据有限,需要更多研究。或许研究不应仅关注小脑在动态情感面部表情中的作用,还应关注其在情感表达的动态个人或社会背景中的作用。
3.4 小脑社会和情感学习的其他解释
尽管有令人信服的证据支持小脑在社会动作序列识别中的作用,但关于小脑如何促进社会和情感学习,还存在其他可能的机制。我将首先回顾与时间序列假设更互补的解释,然后探讨完全不同的理论方法。
3.4.1 基于自适应误差的学习和预测
现代对小脑的解释,聚焦于基于自适应误差的学习和预测,将其视为小脑的核心功能。如前所述,序列假设基于预测框架,但强调时间序列是预测的重要基础。然而,序列识别并非预测的唯一机制,因为社会和情感预测的意外偏差,也会增加小脑的激活和预测误差。
对时间的强调,与基于误差的学习和预测计算模型大致相符。这些模型通过在序列的每个时间步逐渐最小化误差来学习,因此本质上对时间非常敏感。这些计算模型与小脑功能有何关联呢?计算模型包含三个核心要素:输入预处理、反馈信号和自适应处理,这些要素在小脑等生物系统中也存在。在小脑中,输入预处理将苔藓纤维信号转换为适合基于误差学习的颗粒细胞表征。鉴于哺乳动物大脑中一半以上的神经元是颗粒细胞,这是一个重要的、大规模的输入重新编码过程。关于期望结果或奖励的(未)获得情况和大小的反馈信号,由脑干中的下橄榄核计算,并通过攀缘纤维传递到小脑。自适应处理在浦肯野细胞中计算,它将学习记忆保留在突触强度中(类似于计算模型中的连接权重)。这些细胞接收输入信号和误差信号,并将其产生的反应分配到包括新皮层在内的众多脑区。对小脑时间编码最具意义的是,颗粒细胞能够学习维持一到两秒的预期活动,直到攀缘纤维发出奖励传递信号。这使得需要在较长时间间隔内进行学习的认知过程成为可能。然而,这些较长的时间间隔,如何维持小脑对后续运动或认知过程的序列学习,目前尚不清楚。
经典和更先进的计算模型,均支持对时间序列的处理。现代预测学习计算模型,将时间信息作为进行未来预测和基于误差校正的基础。序列学习在网络的双向递归连接权重中积累,并且也在预测未来时间序列的前瞻性预测神经元中进行特定编码,这与小脑的过程非常相似。因此,小脑中前瞻性预测神经元的结构,可能对需要处理序列的任务特别有效,例如在社会互动中协调顺序动作,以及进行心理状态归因和行为。
3.4.2 无序列的小脑学习与自发序列
对序列假设的一个质疑是,在没有明确的序列操作或社会情感违规的情况下,也经常观察到小脑的募集。例如,在前面提到的 Van Overwalle 等人对社会认知的小脑元分析中,很大一部分研究并未明确操纵序列,但仍发现至少 30% 的小脑活动。
社会心智化任务中观察到小脑募集的一个可能解释是,其中一些任务可能引发了自发序列。事实上,在元分析中,小脑并非在所有心智化任务中都均匀激活。在具有视觉元素(但无生物运动)且内在包含序列的故事类刺激中,小脑最常被募集。这可以通过分析不同类别中募集小脑的研究比例来验证。例如,能触发小脑激活(在超过 40% 的研究中)的任务包括:形状表现得像人类且 “目标导向” 的动画(如一个三角形追逐另一个三角形,占 46%);描绘人类有意目标导向行动的 “动作卡通”(如结局取决于主人公意图的故事,占44%);邀请对(亲近)他人特质进行推断的 “动作故事”(如主人公 “在上班路上对每个人微笑”,暗示友好,占45%),尤其是当动作 “不一致” 或呈现角色 “面孔” 时(占 83%);以及涉及过去、未来或假设行动或事件的类别,包括自传性记忆和对负面生活事件的反事实 “要是我…… 就好了” 的思考(占 83%)。相比之下,在涉及错误和真实信念或道德判断的故事中,小脑的激活最少(最多在 10% 的研究中),这可能是因为它们涉及一次性的单一行动,或者是由于其典型的语言输入方式。对于情感刺激,类似的解释是,它们在没有序列操作的情况下,内在地引发了导致小脑激活的动作序列。大多数情感理论假设,情感会触发即时的行动倾向,如愤怒时的战斗、恐惧时的逃跑、悲伤时的哭泣、喜悦时的跳跃或厌恶时的退缩,这些都是对情感刺激的面部和身体反应的一部分。这种隐含的动作序列,可能会产生一些轻微的小脑激活。
总之,一些以连贯且可视化的故事形式叙述动作序列的社会任务,以及情感表达,即使在未对序列进行实验操纵的情况下,也可能引发自发的序列过程和小脑活动。这一解释可以通过在控制其他关键变量(如熟悉度或自动化程度)不变的情况下,区分可能触发或不触发自发序列过程的刺激,来直接验证。
3.4.3 语言
除了社会和情感学习,语言是另一种似乎在 Crus 区域招募与心智化类似网络的心理过程,但语言似乎也会招募一个独立于心智化网络的小脑语言网络。语言,尤其是言语叙事,通常按时间顺序传达关于社会角色及其心理状态(如特质、信念和动机)的信息。仅有有限的证据表明,纯粹的非社会语言序列会募集小脑后部,如字母序列和非社会语义句子处理。此外,最近的社会序列研究显示,小脑后部会被募集(见 “社会心智化中的小脑序列学习” 小节),这些研究依赖于语言和非语言材料(图 4b - e)。这使得语言不太可能成为小脑社会和情感学习的候选机制。需要更多研究来确定小脑后部对社会和语言材料的选择性,以及这两种小脑过程可能的相互作用方式。
综上所述,关于社会和情感学习的几种替代解释,要么与序列假设兼容(预测性解释),要么可以由叙事性刺激材料引发的自发序列过程来解释,要么在整合语言和非语言序列方面存在不足(语言解释)。然而,要根据小脑的理论解释确定序列识别的具体作用,以及其他互补过程(如自发序列)的具体作用,仍需开展更多工作。
4. 结论
小脑对于理解社会和情感行为的序列动态,以及识别预期社会行为的偏差至关重要。对小脑功能图谱、元分析,以及小脑与大脑互惠连接的研究,揭示了后 Crus 区域和小叶 IX 在支持社会动作观察、社会心智化和情感理解方面的独特地位。仍有许多开放性问题有待探索。
Crus 区域广泛参与从低层次社会动作和情感观察,到高层次社会和情感理解等不同的社会和情感过程。这一共同区域表明存在共同的心智化过程?还是说它服务于动作观察和心智化过程的混合?亦或是支持一个统一的小脑过程,例如识别时间序列,而不考虑其具体的社会或情感内容?未来需要进一步探究的另一个问题是,序列处理在多大程度上是小脑特有的、专门用于预测和自动化的过程,以及小脑是否可能支持与序列无关的其他过程。我们需要更详细的理论来解释小脑如何影响社会和情感行为,这可能会进一步明确预测性、序列性和其他替代解释之间的差异、相似性和局限性,并为未来研究提出相关问题。
如果序列识别是社会和情感理解的关键(但可能并非唯一)组成部分,那么它可能会让小脑功能障碍患者更好地理解自身困难,并且如果在训练计划中加入序列元素,他们可能会从训练中获得更多治疗益处。例如,一些研究报告称,患有自闭症的参与者在解决错误信念序列问题时速度较慢,而自闭症患者中与小脑相关的胎儿或基因缺陷发生率较高。在接受了一个为期六节、侧重于故事理解中的心智化和序列识别的训练计划后,自闭症患者在社会序列和故事连贯性方面有了令人鼓舞的改善。需要更多尝试来诊断患者群体的序列识别困难,并通过训练计划提高他们的序列识别能力。
未来研究的另一个方向是关注大脑中其他可能也负责时间序列处理的皮层下核团,如基底神经节,它们在皮层下层面与小脑存在解剖连接。一种理论方法认为,基底神经节通过传递奖励预测误差来最大化未来行动的价值,而小脑则负责追踪具有最少序列错误的最适应性轨迹,二者之间存在相互作用。出于理论和治疗方面的原因,加深我们对小脑和基底神经节支持的不同学习功能及其相互作用的理解非常重要。
对小脑后部的非侵入性神经刺激研究正在不断发展。尽管目前的尝试前景良好,但用于改善社交技能和情感的设备(如 tDCS 中的电极和 TMS 中的线圈)的最佳设置,以及刺激条件(如刺激的聚焦度和个体的瞬间状态)仍在评估中。这项研究可能会成为改进非侵入性神经刺激、开发未来治疗应用,以及进一步理解小脑后部在支持社会动作观察、社会心智化和情感理解方面作用的重要途径。
参考文献:Social and emotional learning in the cerebellum.


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









