



 研究通过对大量弥漫大B细胞淋巴瘤(DLBCL)患者的转录谱分析,揭示了四种主要的淋巴瘤微环境(LME)类别,与生物学性能和临床行为相关。这四种类别包括:生发中心样(GC-like)、间充质样(MS)、炎症性(IN)和耗竭型(DP)。GC-like和MS-LMEs与较好的临床结局相关,而DP-LME与较差的预后和高肿瘤细胞增殖、DNA低甲基化及免疫逃逸相关。LME的细胞组成和功能分析显示,ECM成分、细胞因子和免疫细胞的动态变化在疾病进展中起关键作用,为靶向治疗提供了新的方向。
研究通过对大量弥漫大B细胞淋巴瘤(DLBCL)患者的转录谱分析,揭示了四种主要的淋巴瘤微环境(LME)类别,与生物学性能和临床行为相关。这四种类别包括:生发中心样(GC-like)、间充质样(MS)、炎症性(IN)和耗竭型(DP)。GC-like和MS-LMEs与较好的临床结局相关,而DP-LME与较差的预后和高肿瘤细胞增殖、DNA低甲基化及免疫逃逸相关。LME的细胞组成和功能分析显示,ECM成分、细胞因子和免疫细胞的动态变化在疾病进展中起关键作用,为靶向治疗提供了新的方向。
文献阅读 Clinical and Biological subtypes of B-cell lymphoma revealed by microenvironmental signature
Abstract
弥漫大B细胞淋巴瘤是一种生物学及临床异质性很强的疾病。对DLBCL转录谱和基因组的特性的研究增加了对疾病内在发病机制的理解并且提供了潜在的治疗靶标。但是,DLBCL生物学上的微环境角色目前还不是特别清楚。在本篇文章,我们对4655例来自于多独立队列的DLBCL患者进行转录谱的分析并且描述了四种主要的淋巴瘤与生物学畸变和临床行为相关的微环境种类。我们仍然发现癌细胞为逃避淋巴瘤生长的微环境限制而部署的遗传和表观遗传机制的证据,支持了在特定DLBCL患者中实施DNA低甲基化剂的基本原理。除此之外,我们的工作仍然发现了在细胞外基质的生化组成部分中新的治疗可能性,这种治疗方式可以在临床前治疗中降低DLBCL细胞的增殖。
Significance:
基于转录相关的转录谱的分类,我们将微环境作为B细胞淋巴瘤生物学上的主要组成部分并且将其与DLBCL临床行为相关联,为靶向治疗提供新的机会。
Introduction
弥漫大B细胞淋巴瘤,在成人中最常见的淋巴系统恶性肿瘤,在超过60%的患者中使用基于抗-CD20抗体联合免疫治疗是可以医治的。在过去的几年中,几种新型的靶向药物已经开始应用,但是还是有小部分复发的患者死于淋巴瘤或出现并发症。疾病的进展或治疗的结果依赖于阐明(elucidating)这种异质性疾病临床行为的基础生物学。基于细胞起源(COO)分类的基因表达谱的分析可以从细胞来源上辨别两种主要的DLBCL的亚型:包括ABC(activated B-cell)and germinal center B-cell(GCB)。这两种亚型拥有不同的生物学行为以及分子特征,反应了不停的发病机制。最近,关于弥漫大B细胞基因谱alterations包括基因突变,体细胞拷贝数变异以及染色体结构的变异方面的研究可以鉴定COO亚型中不同的genetic subtypes。在ABC-DLBCL亚型中,MYD88和CD79B突变co-occuring或者harboring NOTCH1突变可能是较其他的ABC-DLBCL亚型的患者来说拥有较差的预后。同样的,在GCB亚型中携带EZH2突变或者BCL2 translocations的淋巴瘤患者常常结局更差。同时,在GCB亚群中携带MYC和BCL2和/或BCL6 rearrangements预示着GCB亚群具有明显的攻击的生物学行为,也称之为双打击或三打击高级别B细胞淋巴瘤。HGBL-DH/TH。
DLBCL的生物学和临床行为结局不仅仅来自于DLBCL细胞的分子突变,同样来自于微环境之间的联系。来自淋巴瘤患者和动物模型数据表明,在淋巴瘤的生态位中,微环境和细胞外基质提供的外部刺激有助于疾病的发展,进展和对治疗的反应。因为淋巴瘤细胞和为微环境的联系是双向的(bidirectional),预示着DLBCL微环境将表现出尚未定义的分子和功能的复杂性。尽管在DLBCL生物学上越来越意识到微环境的重要性,关于疾病的分子和治疗的研究也主要是集中在DLBCL细胞的特性作为一个分离的实体(as an isolated entity)
在这篇文章中,我们基于大量患者微环境细胞和过程转录足迹对DLBCL微环境进行了表征。我们发现四种主要反应不同的生物学性能和临床行为的微环境的组成。这些新描述的类别与遗传上相似的DLBCL的不同临床行为相联系,并为创新的治疗干预提供了一系列新的潜在靶点。
Results
The lymphoma microenvironment is characterized by four distinct category
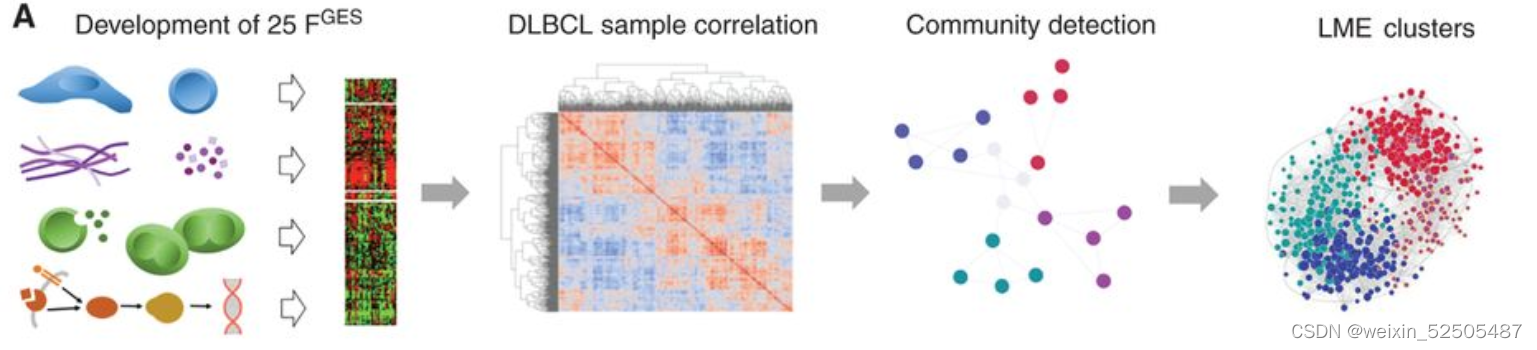
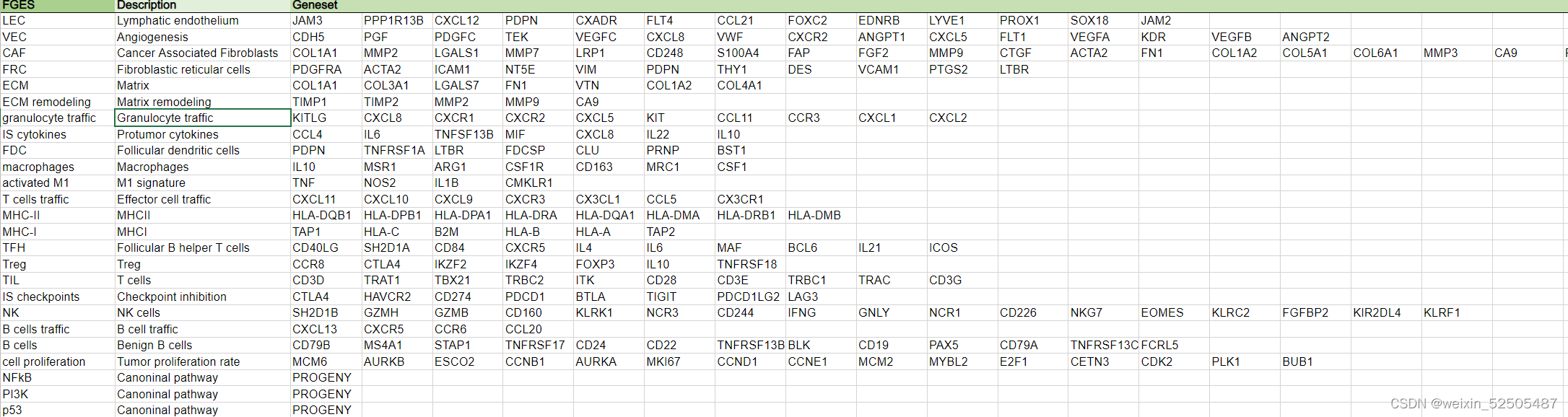
我们的目标是根据数千名患者的基因表达谱所赋予的统计能力(statistical power)来定义DLBCL微环境组成和功能的多样性。作为实现目标的第一步,我们总结了25种Functional Gene Expression Signatures(FGES),包括反应不同的和肿瘤微环境相关的细胞亚型(例如肿瘤相关的成纤维细胞和肿瘤浸润淋巴细胞),肿瘤微环境的非细胞组分(例如免疫抑制细胞因子和ECM),分子过程(例如,分泌和增殖),癌症信号通路激活(例如,PI3K和NFκB)Fig. 1A; Supplementary Fig. S1A–S1C; Supplementary Table S1
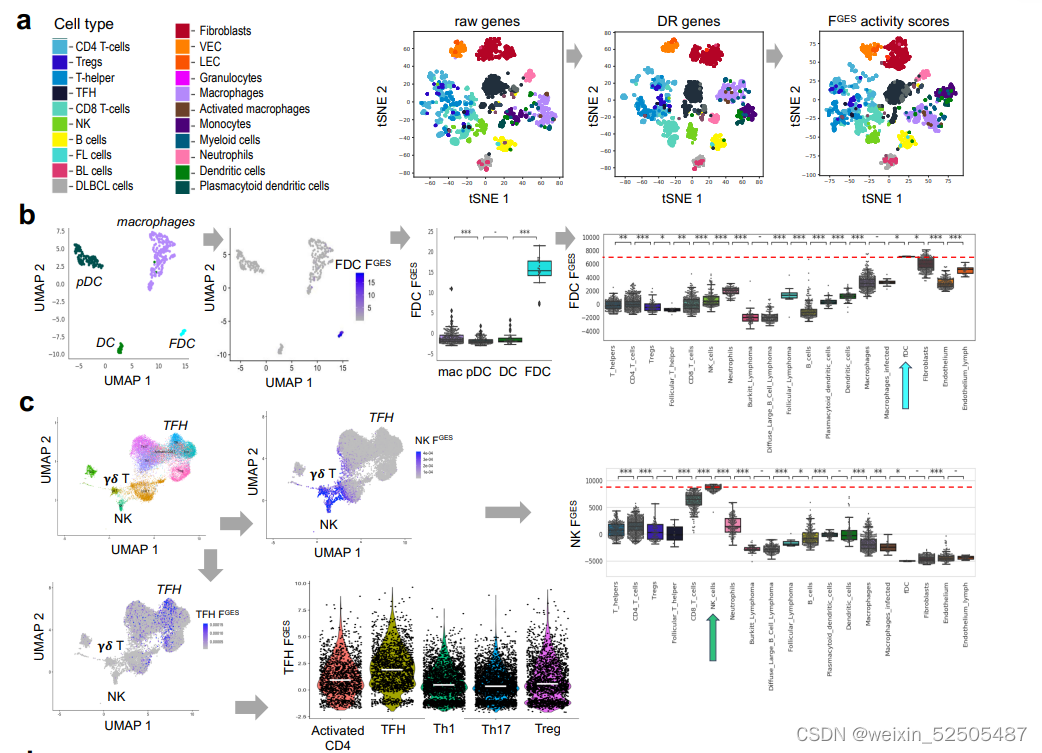
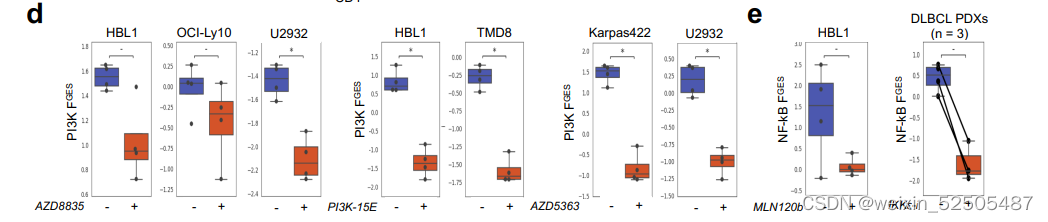
对于每一种FGES,我们计算活性指数(与特定效应在细胞群中存在的程度直接相关),例如,PI3K和NFκB FGES都是足够敏感可以检测到在DLBCL细胞系和小鼠模型中特异性抑制时在通路活性上的改变。
(Supplementary Fig. S1D and S1E)
FGES的活性分数揭示了它们衍生的细胞类型的预期富集,以及潜在新型功能相互依赖的关系。例如,尽管ECM FGES几乎与成纤维细胞相关,但是"ECM remodeling" FGES也与活化的巨噬细胞相关,表明了这些细胞在ECM微结构中的潜在作用。同样的,它们可能暗示了细胞状态,例如,生物学过程中‘cell proliferation’中FGES大部分在非增殖的循环B细胞中缺乏并且progressively higher从低增殖的FL到DLBCLs到高增殖的burkitt lymphoma的BCL中。 Fig. 1B
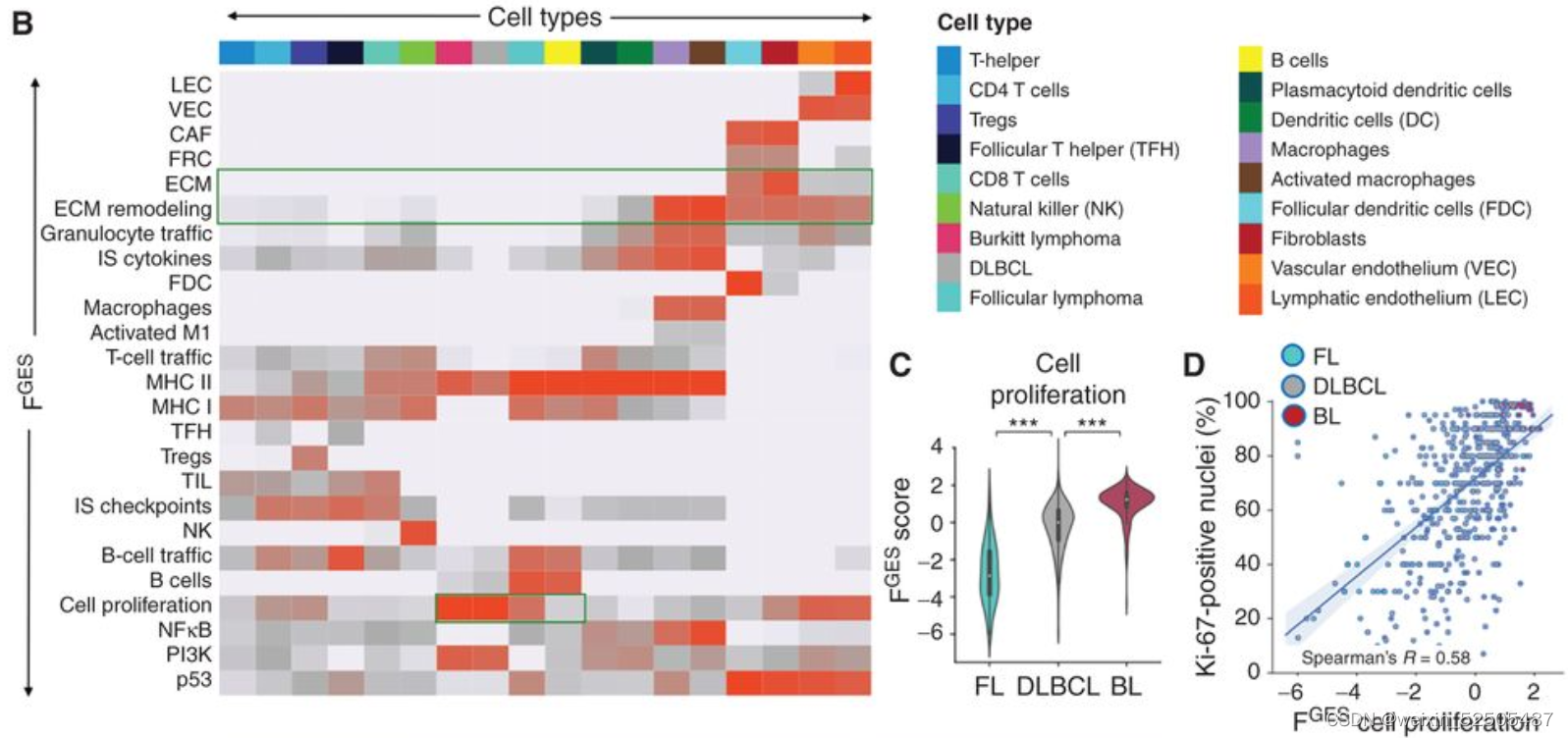
为了去验证来源于混合细胞类型的组织的基因表达谱时FGES是否还具有informative,我们分析了“cell proliferation” FGES的活性分数在4984例B细胞淋巴瘤的基因表达谱中(FL,DLBCL,BL以及发现的相似的单个细胞), FGES的活性分数直接与报道过这些疾病的增值率相关。Fig. 1C
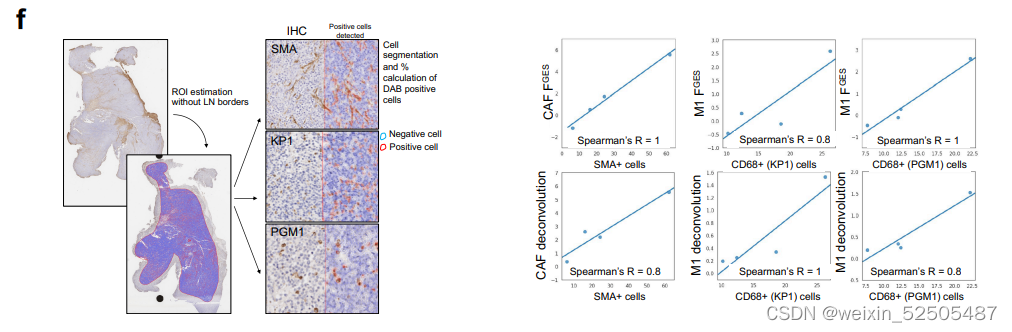
更重要的是,作为一个额外的独立的验证,细胞增殖的FGES与来源于532B细胞淋巴瘤测定的增殖率的黄金标准Ki-67 IHC相关Fig. 1D。最后,我们证明了被FGES评估的成纤维细胞和活化的巨噬细胞的比例与从匹配的DLBCL组织中抗SMA和抗CD68免疫染色定量获得这些细胞的估计相关。Supplementary Fig. S1F
我们利用25种FGES去虚拟重建4580例DLBCL样本(收集与于公共的基因表达谱的数据库以及75例 local RNA-seq)的淋巴瘤微环境(LME)(Supplementary Fig. S2A)
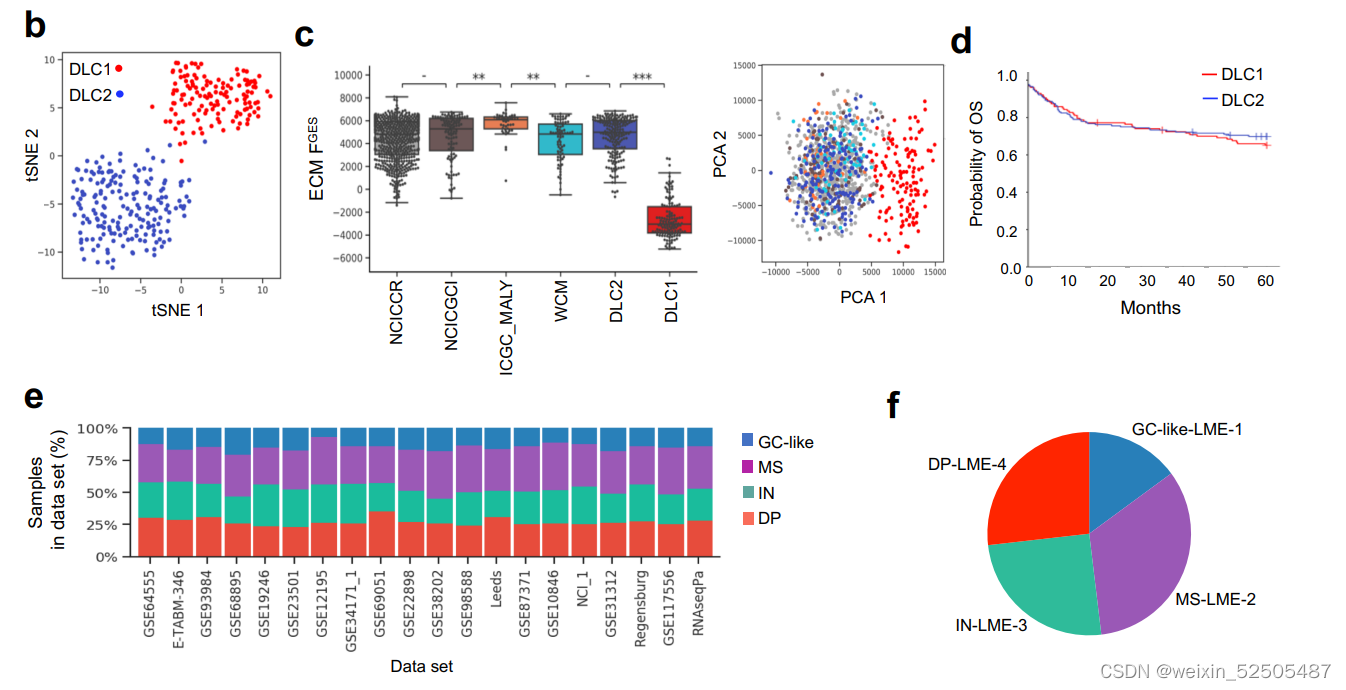
我们利用unsupervisd community detected algorithm分析了不同样本之间FGES的相关性并且获得四种主要的LMEs。这四种LMEs反应了不同的FGES和“communities”关系。并且在所有样本中15% (LME-1), 33% (LME-2), 25% (LME-3), and 27% (LME-4) 并且在所有测试的数据集中呈现相似的分布 (Fig. 1E; Supplementary Fig. S2B–S2F)
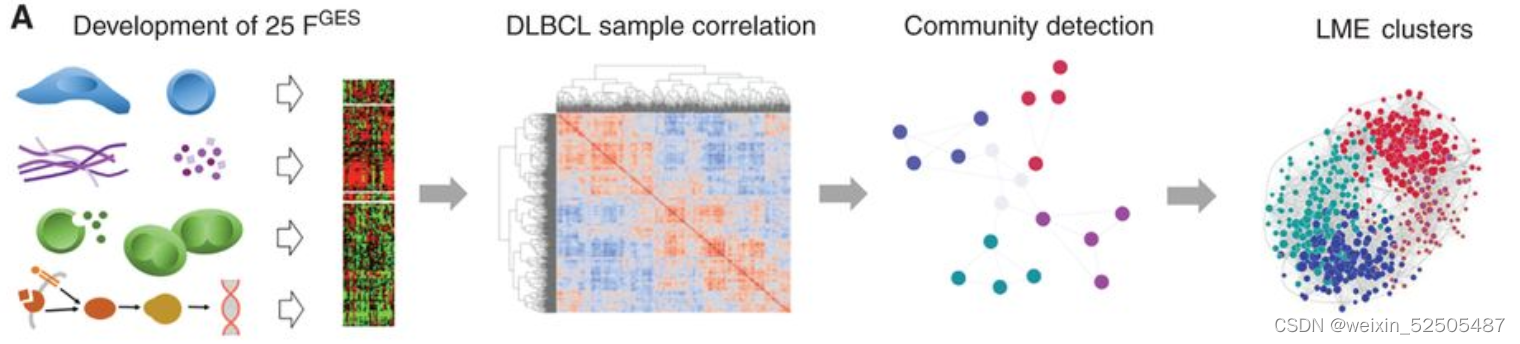
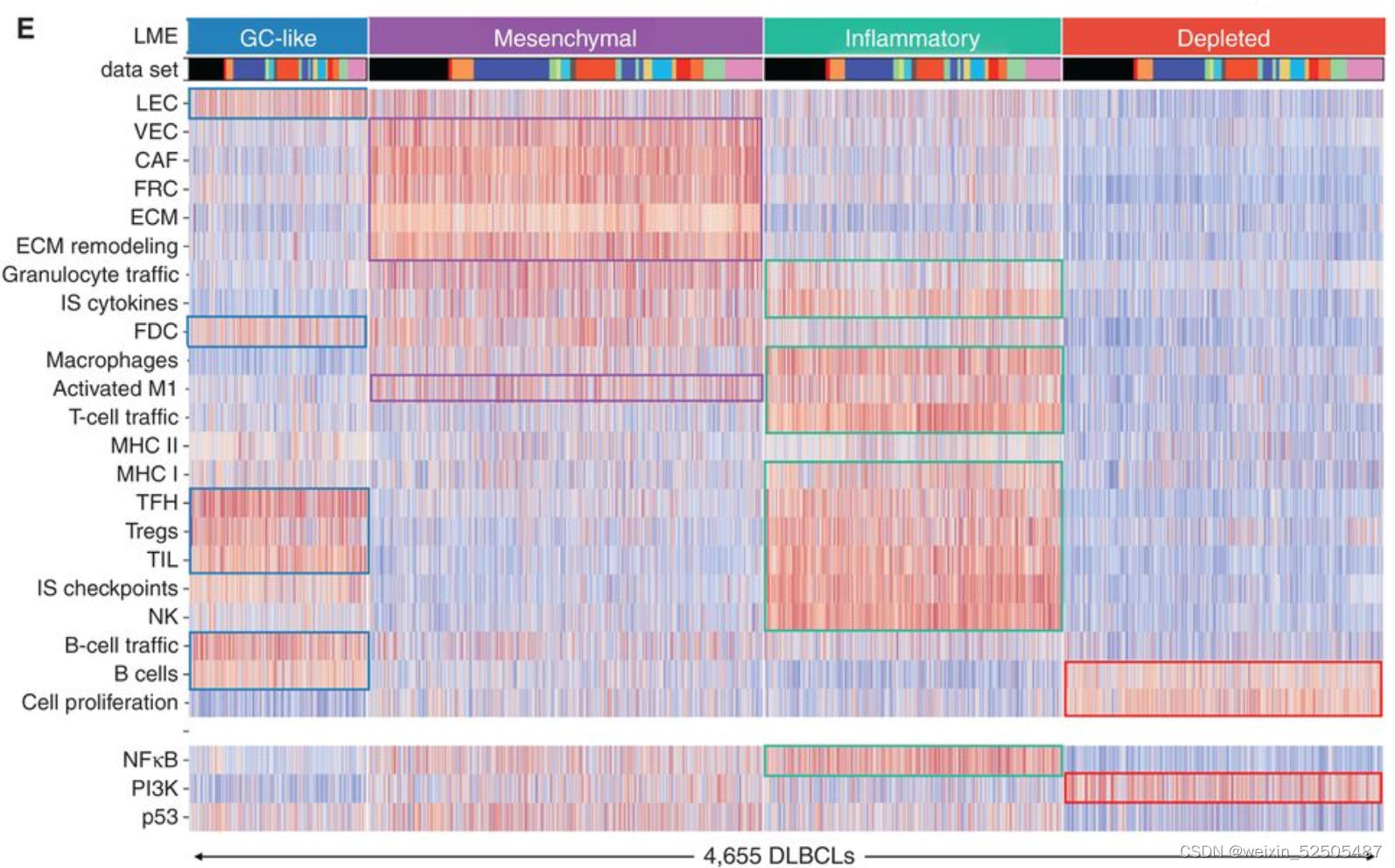
根据每一个微环境中富集FGES的群落,这四种LMEs被分别的termed,germinal center-like其特征在主要源自于普遍在生发中心被发现细胞类型的FGES的存在(GC)。mesenchymal(MS;间充质)其特征主要是源自于基质细胞和ECM通路的FGES的缺乏。“inflammatory”(IN)其特征主要是源自于炎症细胞和通路FGES的存在;最后,depleted” (DP;耗竭)模式,与其他LME相比,其特征在于微环境来源的FGES的缺乏。
LME Categories Represent a novel gene expression-based DLBCL Classification
我们利用了platform-independent algorithm根据ABC-DLBCL和GCB-DLBCL亚型来分析个体的淋巴瘤。
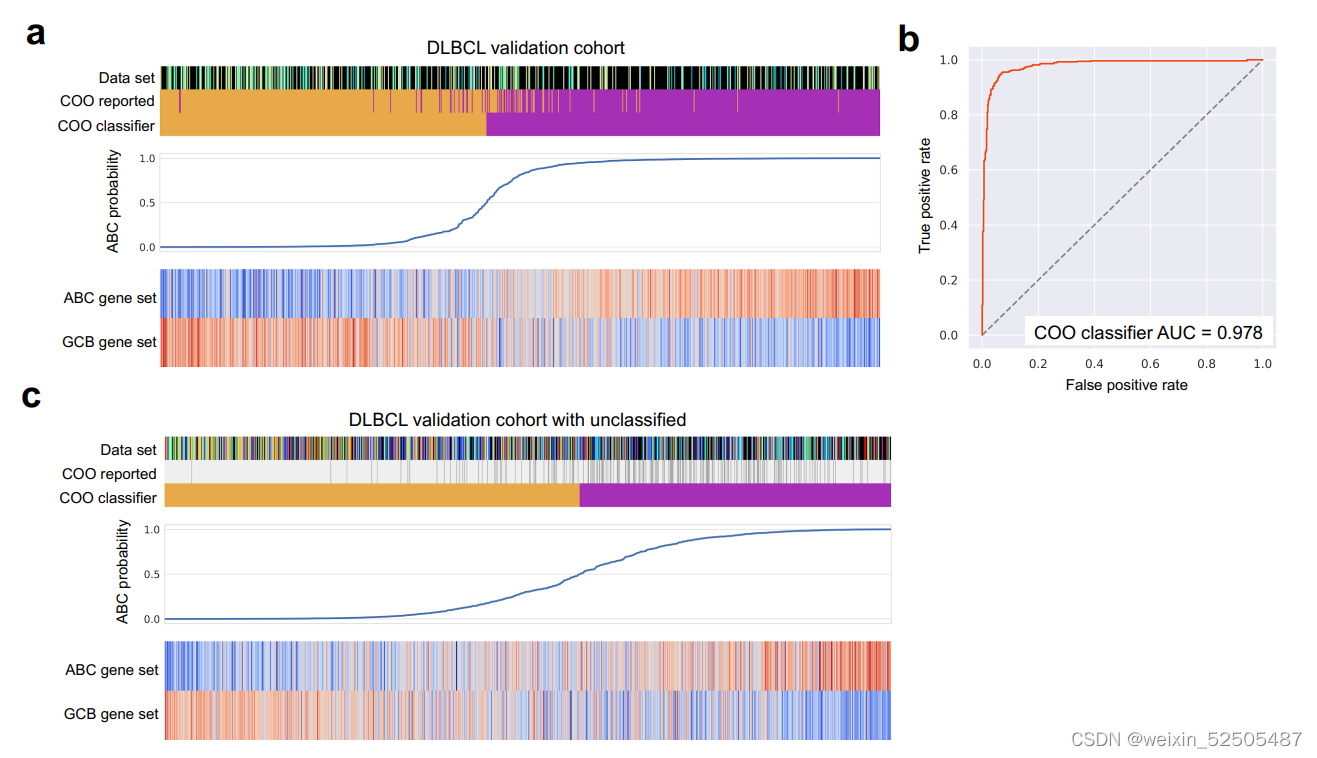
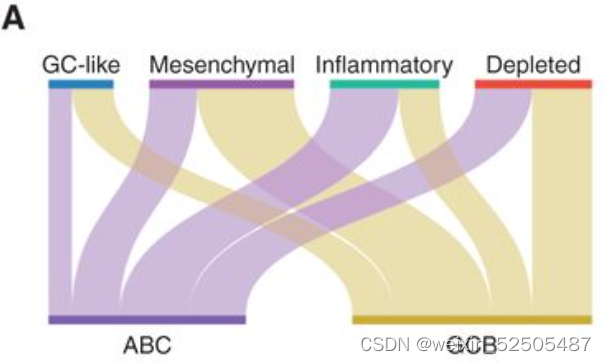
在全部队列中(n=4655),59%都是GCB-DLBCLs和41%是ABC亚型。这些DLBCL亚型在LME关联方面存在差异。例如,inflammatory-LME亚型在ABC亚型中比例更高,GC-like和MS-LMEs则在GCB亚型中富集。(Fig. 2A, P < 0.001, χ2 test, all comparisons) Figure2A
对于subsequent分析需要不同的分子或临床注释,我们使用了具有所需信息的最大子集。Supplementary Fig. S2A
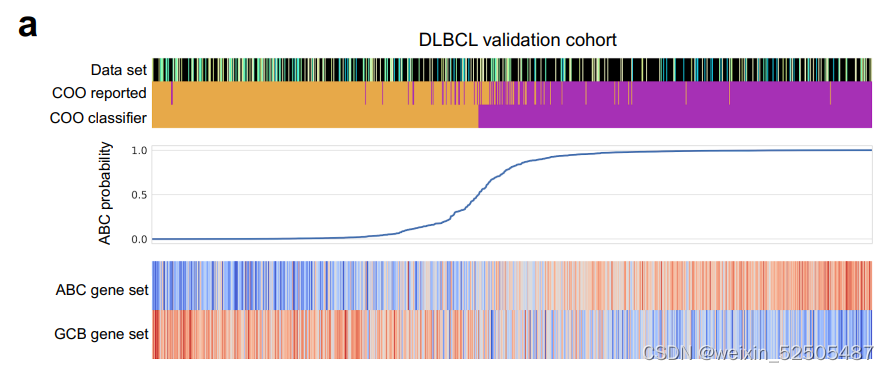
我们investigated LME类别与采用Consensus Clustering Classification(CCC)iyin表达亚型B细胞受体(BCR),氧化磷酸化以及宿主反应的相关性。正如期望的一样,DP-LME和HR(Host response)没有联系,而与BCR和OXPHOS展示了特殊的LME模式**(Supplementary Fig. S3D)。

除此之外,MS-LME主要富集于之前描述的“stromal-1”和“stromal-2”的转录谱signature。Supplementary Fig. S3E
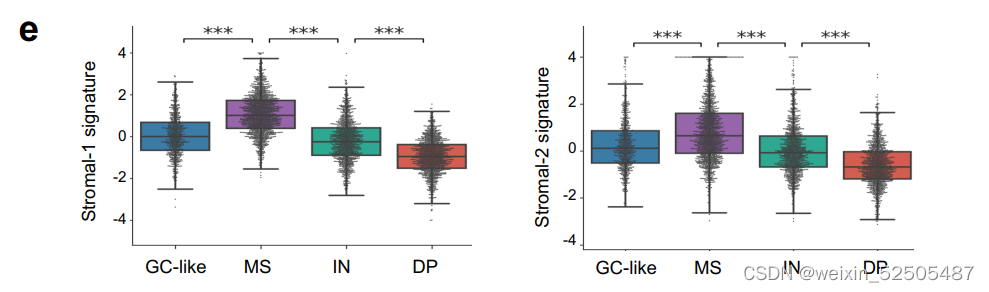
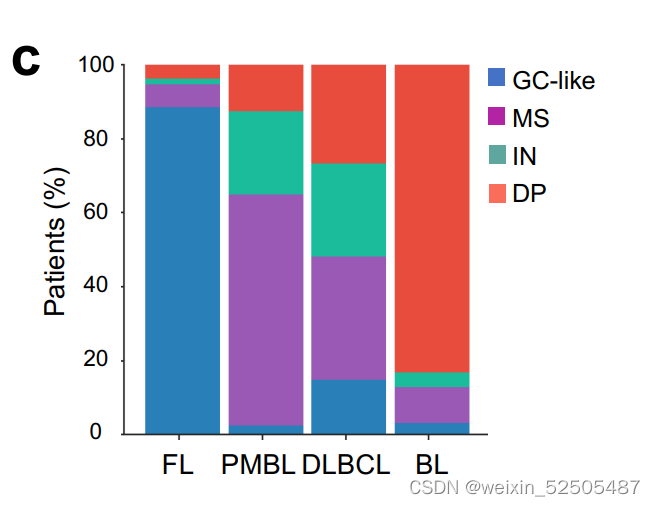
值得注意的是,根据分析1397例可利用的数据,HGBL-DHs主要富集在DP-LME中,其次是富集于GC-LME中。Supplementary Fig. S3D
MYC和BCL2的并发重排并不是DLBCL中侵袭性的唯一分子决定因素。两种基因表达的signature,MHG(molecular high-grade)和DHL的signature(DHITsig) 表现为HGVL-DH的特征行为但是却没有特征性的基因的突变。我们根据我们自己的与LME分类相关的数据应用于DHITsig classifier**。Supplementary Fig. S4A–S4G; Supplementary Table S3 ,并且在DP-LME分类中寻找MHG和DHITsig患者significant enrichment寻。(Supplementary Fig. S3D)。相当于在高级别的基础上利用MHG和DHL进行细分)
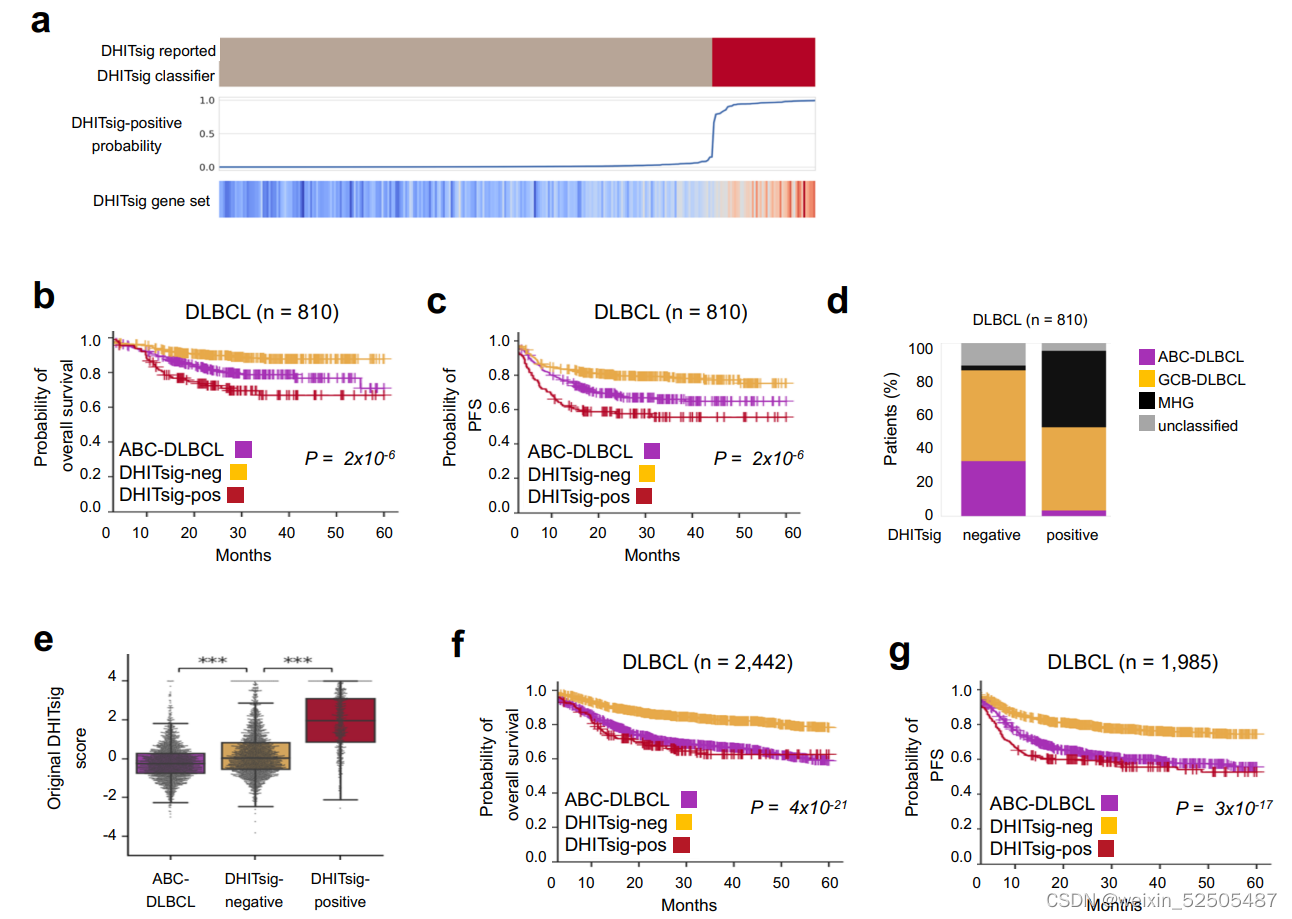
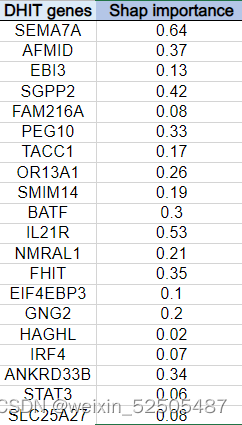
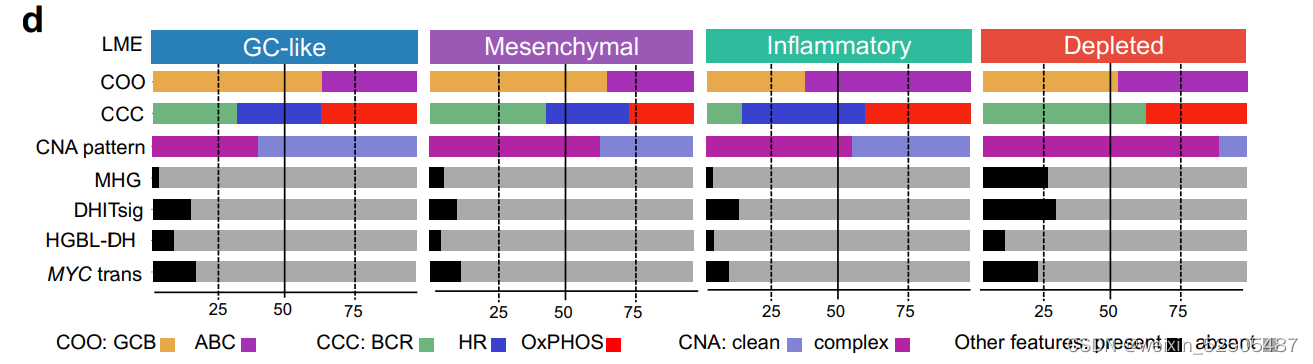
更重要的是,molecular high-grade(MHG)和DHITsig患者高度重叠,很大程度上反应了相似的生物学功能。Supplementary Fig. S4D
最后,在小鼠模型中趋向于在GCB细胞中双表达myc和bcl2,由此产生的B细胞淋巴瘤拥有和DP-LME DLBCLs相似的LME。
DLBCLs最近被重新分类为不同临床特征的遗传定义的实体瘤。其中也与微环境的特征相关。经过一些努力将DLBCL分为MCD亚型(MYD88和CD79B突变的共发生),N1(NOTCH1突变),EZB(EZH2突变和BCL2易位),BN2(BCL6融合和NOTCH2突变),ST2(SGK1和TET2突变)以及A53(非整倍体aneuploid with TP53 inactivion)。这些基因的注释在我们的来自两个队列(NCICCR和DLC2)的737例病例中仍然可以适用并且揭示了四种类别的LME分布不均匀。 (Fig. 2B)
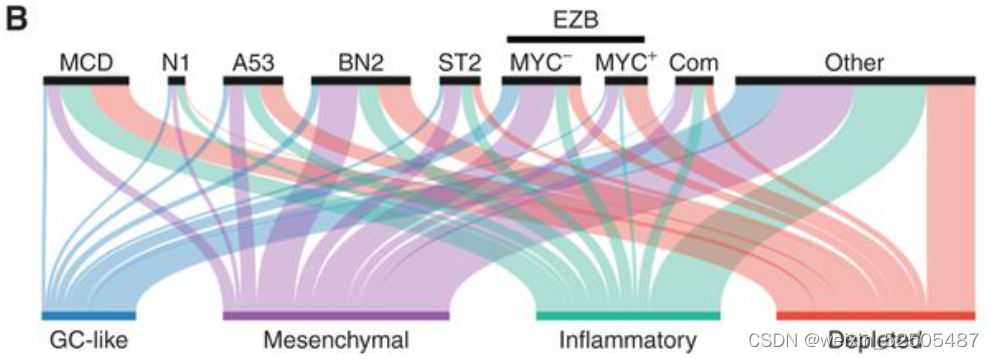
例如,预后较差的MCD(MYD88和CD79B突变的共发生)亚型中有4%GC-like,8%的MS,17.6%的IN和21%的DP-LME (P < 0.001, χ2 test; Fig. 2B)。在LMEs中MCD亚型的分布并不是完全和ABC病例一致。尽管大多数BN2,ST2和EZB的病例有特征性的免疫遗弃性的LME(既包括MS或DP),在EZBs中特殊的免疫遗弃LME分类和MYC的状态相关:EZB-MYC+病中DP占比较多。而EZB-MYC-患者则以MS的LMEs为主。综上所述,包括DLBCL LME分类的病例的结果提供了额外的orthogonal信息是之前关于淋巴瘤分类的报道中没有报道过的。
LME categories associate with specific genomic alterations in lymphoma cells
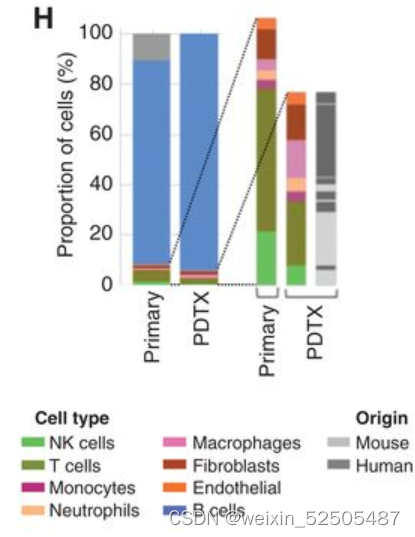
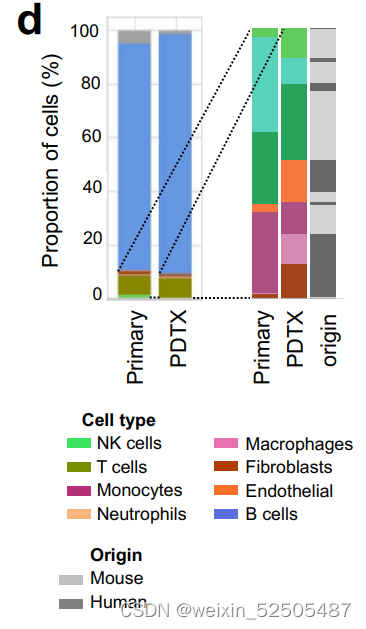
LME依赖于肿瘤与正常的细胞类型之间的动态的联系。为了去更好地理解在driving特殊LME patterning的淋巴瘤细胞的效应,我们通过将患者的DLBCL组织植入免疫抑制的小鼠中,建立了18种患者来源的肿瘤抑制物patient-derived tumor xenografts (PDTX)(PDTX)。我们然后对每一种患者来源的肿瘤抑制物的早期阶段进行RNA-seq并且使用细胞特征性反卷积的方法来确定小鼠的细胞是否以类似于原代人体组织的比例重新填充PDTX。值得注意的是,大多数PDTXs保持与原始原发性人淋巴瘤相似的小鼠LME类别。(Fig. 2C; Supplementary Fig. S5A and S5B)
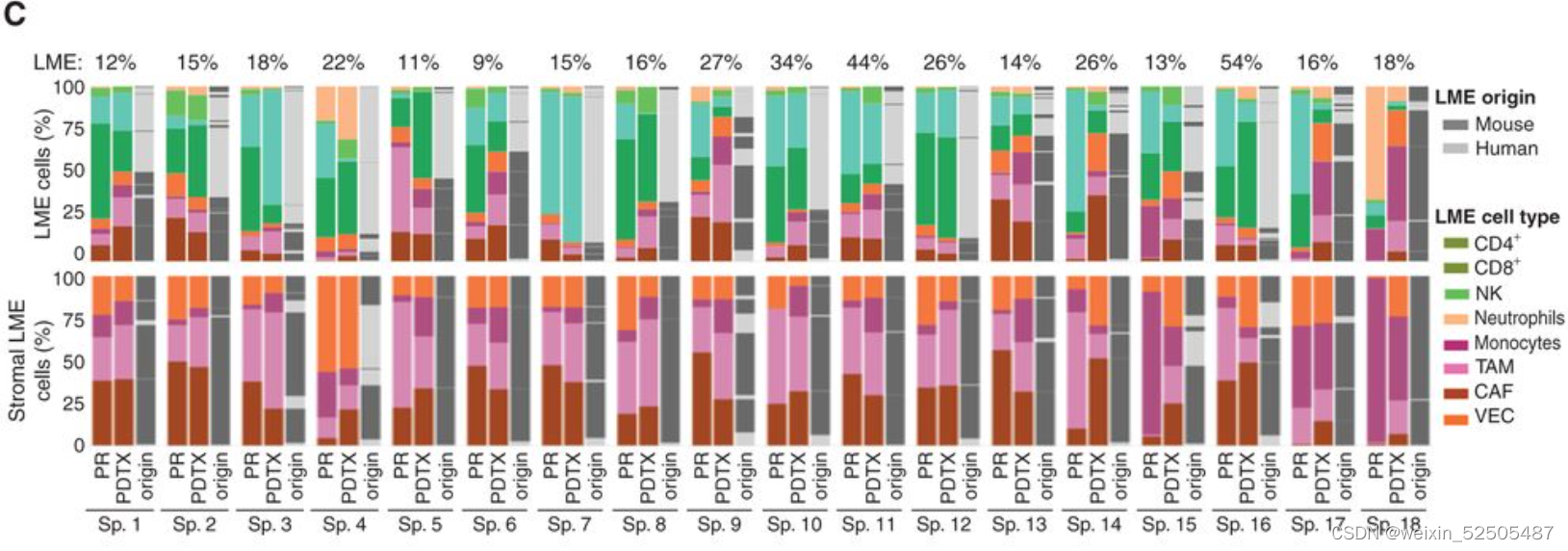
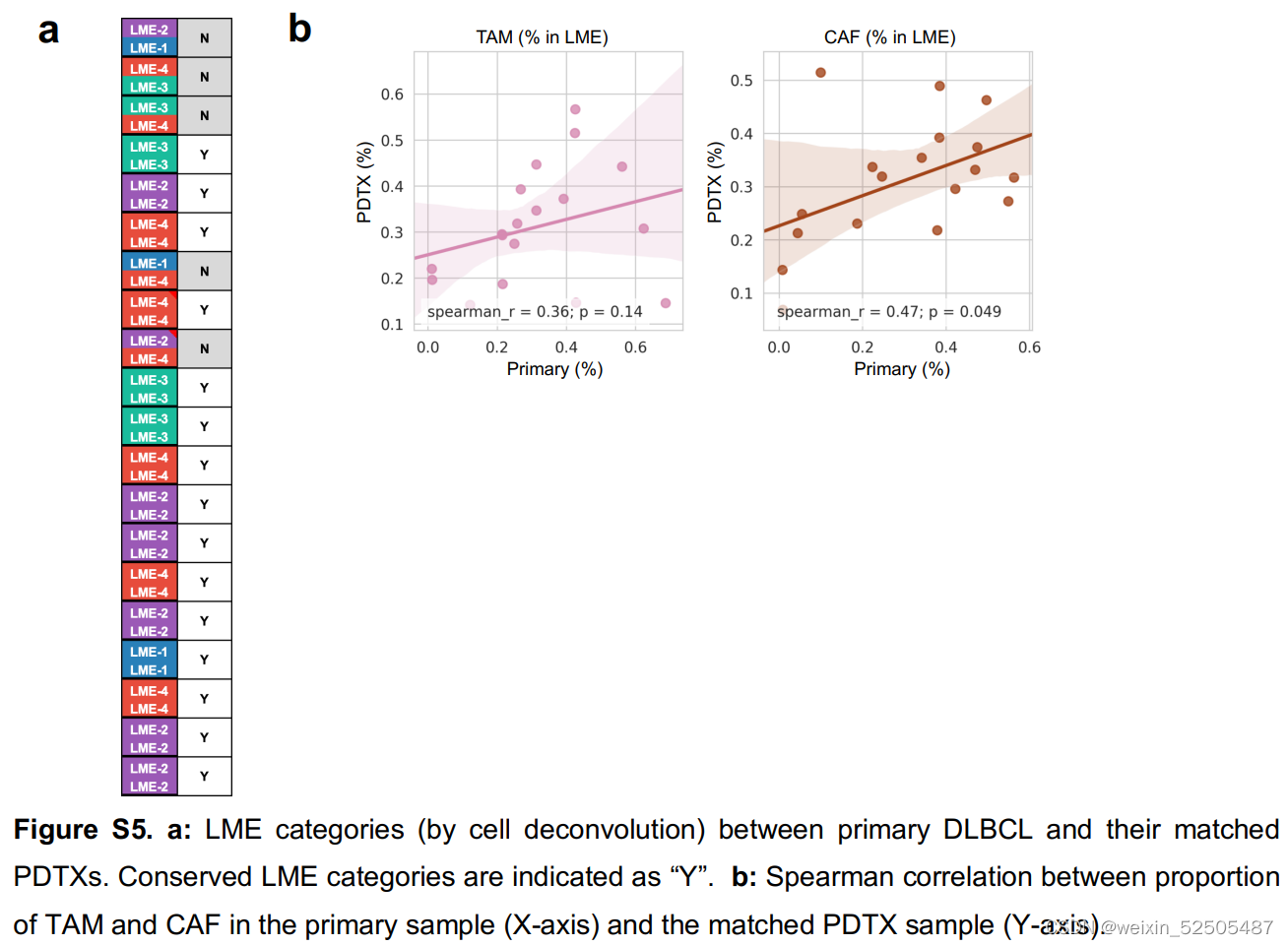
因为受体小鼠是免疫抑制性的,LME免疫组分的维持主要来源于在早期阶段人体原始组织。相比之下,LME基质细胞,例如肿瘤相关的成纤维细胞以及肿瘤相关的巨噬细胞,主要是小鼠的组织招募的。(Fig. 2C)。正如在乳腺癌的PDTX specimens报道的,这表明恶性细胞和LME细胞之间存在潜在的,持久的相互信号通路,这种连接在早期传代PDTX中保持不变。
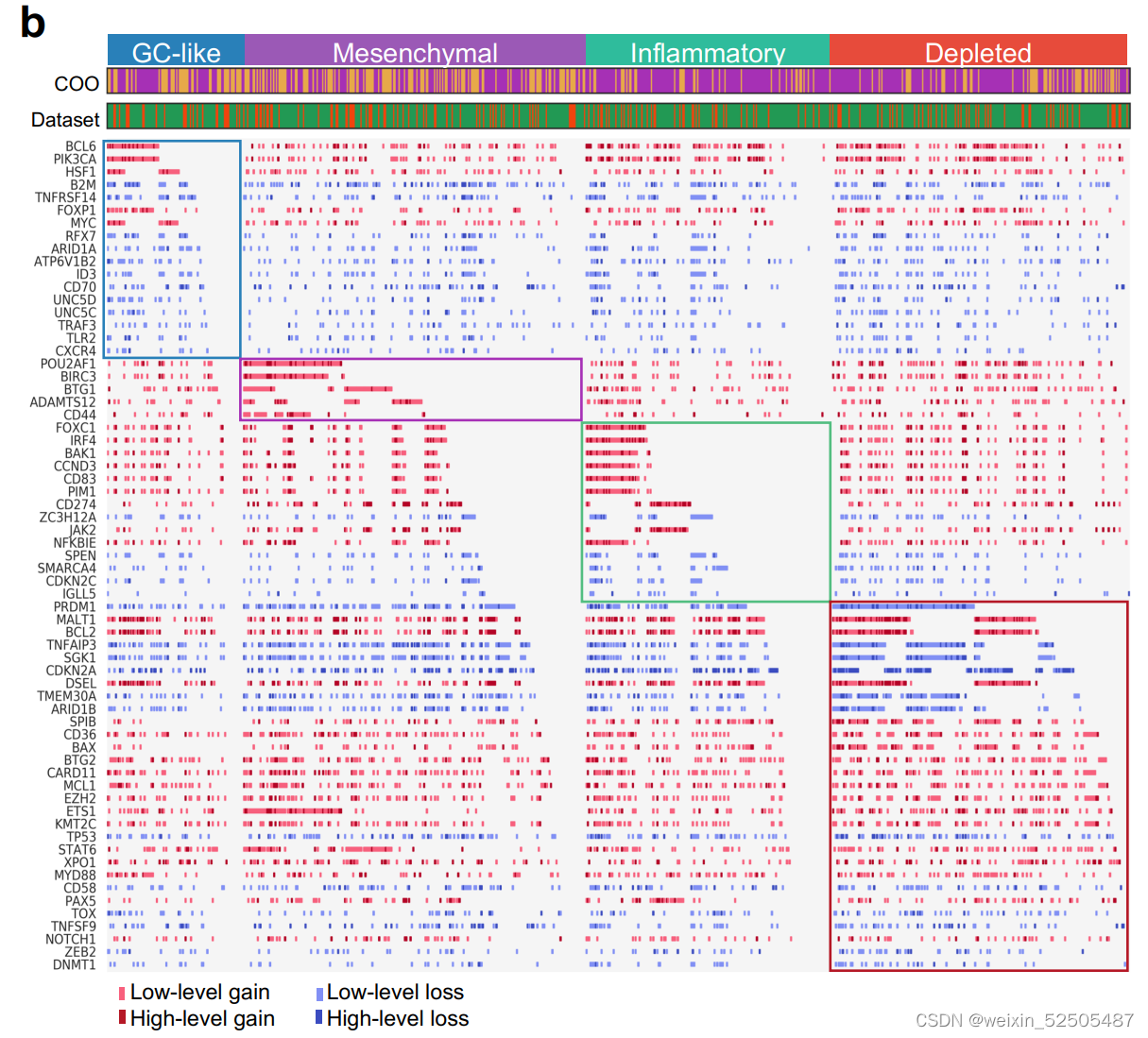
为了探索淋巴瘤诱导的特殊的LMEs的机制,我们比较了突变和拷贝数变异的谱。
突变来自于747名患者全外显子测序以及全基因组测序或靶向测序的结果,而750名全外显子CNA结果是available的,86个最常见的突变的分析以及top65 CNAs分析 (Supplementary Fig. S6A and S6B; Supplementary Tables S4 and S5)显示在特定LME中,有24个突变和22个CNA显著富集并且在LME中分布不均。更重要的是,其中9个分布不均的24个突变和22个CNA中的6个CNAs其LME的类别分布不完全由ABC/GCB来解释。因此,可能与这些突变的直接影响相关。值得注意的是,一些突变和针对动议同一基因的CNAs在特定的LME中coveraged,就像MS-LME中的CD44和DP-LME中的TMEM30A。

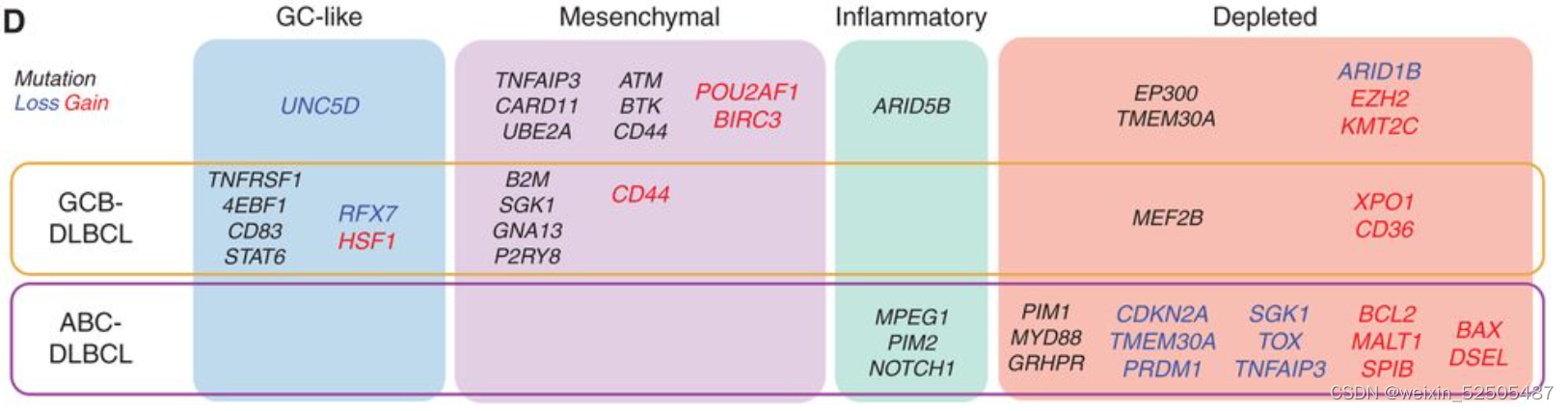
tableS4 mutations
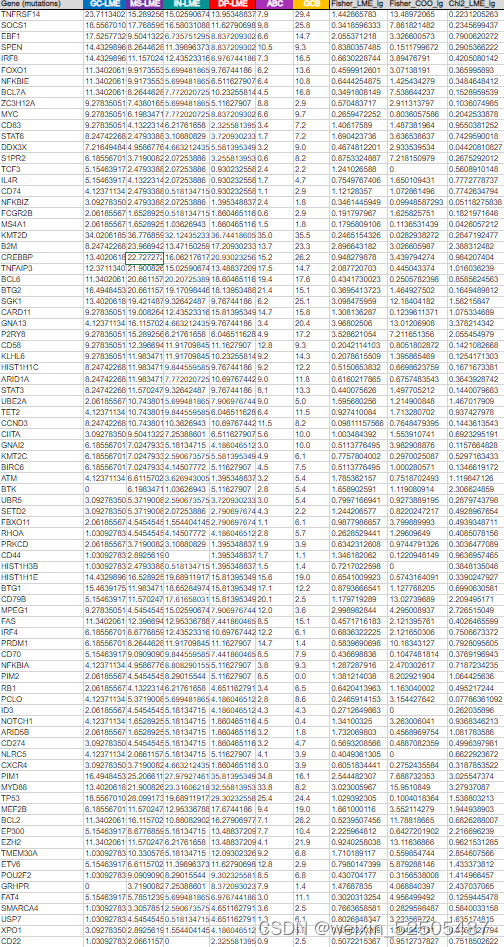
tableS5 CNA
因为CNAs和/或靶向功能相关的基因的突变可能导致相似的表型,因此我们分析与LME类别相关的互补基因组的改变(complementary genomic alterations)。我们认为所有基因组的畸变都可以注释为具有统计学显著性的通路。我们发现在DLBCL的DP-LME组中基因组的突变导致p53活性降低和细胞周期调节的扰动(perturbation)更常见。
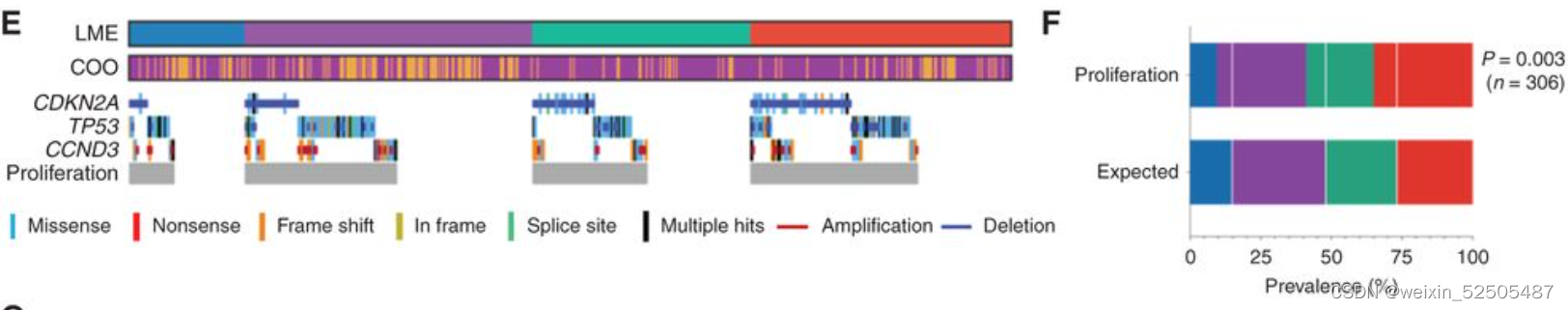
并且可以在67例DLBCLs样本中进行验证,在90%DP-LMEs展现出影响该途径的复杂的CNAs,而其他的LMEs仅仅只有60% (P < 0.04; Supplementary Fig. S3D)。
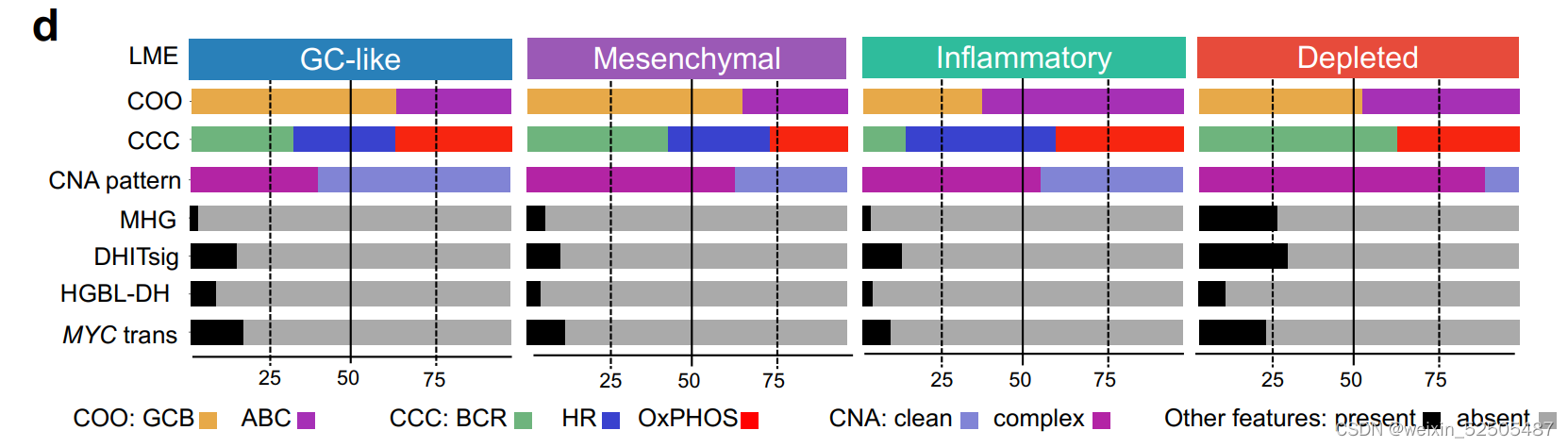
以此为根据,DP-LMEs有更高比例的A53淋巴瘤并且在细胞增殖FGES有很强的富集正如之后描述的那样。
最近的研究显示了MHC I and MHC II antigen presentation genes的表达并且与表达EZH2Y641的小鼠淋巴瘤模型中免疫浸润的相应稀疏性。与这些数据一致,EZH2基因的突变和B2M或CIITA突变一致可以分别影响MHC I or MHC II并且主要富集在MS-LME中。除此之外,MS-LME主要富集在GNA13, GNAI2, and P2RY8

GNA13,/GNAI2和/或P2RY8破坏了Gα信号通路,这条信号通路是对于周围免疫细胞归巢到淋巴结所必须的,这种LME耗尽模式,可能表明淋巴瘤细胞获取了LME非依赖性的生长模式。
LEM categories associate with distinct disease courses
GC-LME和DP-LME分别有最高和最低的国际预后指数 Supplementary Fig. S7A
为了去判断LME分类和R-CHOP化疗反应之间的联系,我们对105名新诊断的DLBCL患者进行RNA测序,这些患者在应答者(55%)和非反应者(45%)之间保持平衡,包括难治性(即从未达到完全缓解)和复发(即 <2年内没有疾病)。responders数量在GC-LME中最高,在DP-LME中最低(Z testsP = 0.030 for GC-like vs. DP and P = 0.022 for GC-like + MS vs. IN + DP)
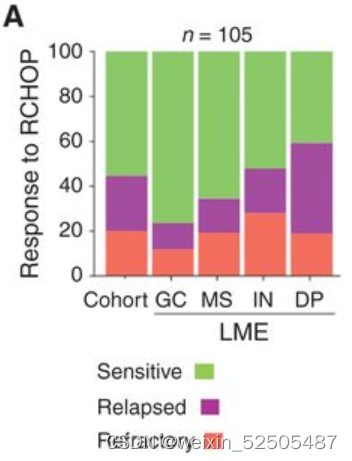
相比之下,DP-LME中non-responders占比最多并且特异性的具有relapsed cases。 (Fig. 3A; 40.5% vs. 16.2% for DP-LME vs. all other LMEs, respectively, Z test P = 0.011)。上述的联系可以在1,349名未经选择的DLBCL的独立队列中得到验证,这些患者经过免疫化学的治疗并且根据RECIL标准进行评估的人群。nonresponders(SD+PD)在GC-like、MS、IN和DP-LMEs中的占比分别为4.5%, 7.3%, 11.1%, and 12.7%
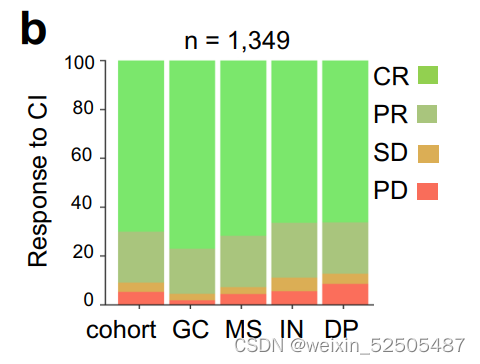
(Supplementary Fig. S7B; Z test P = 5 × 10−4 for GC-like + MS vs. IN + DP-LMEs)
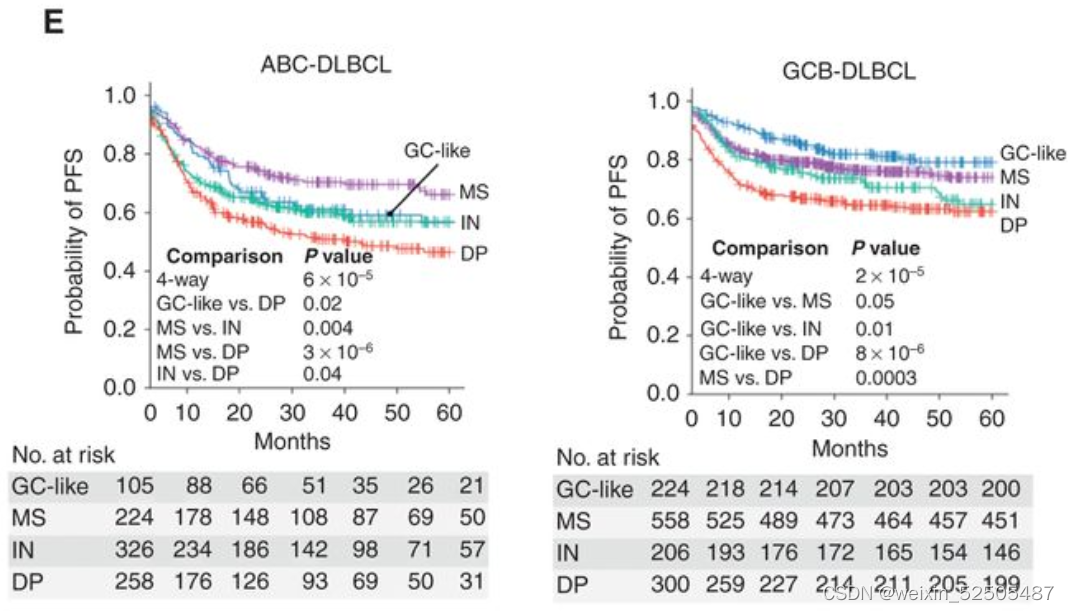
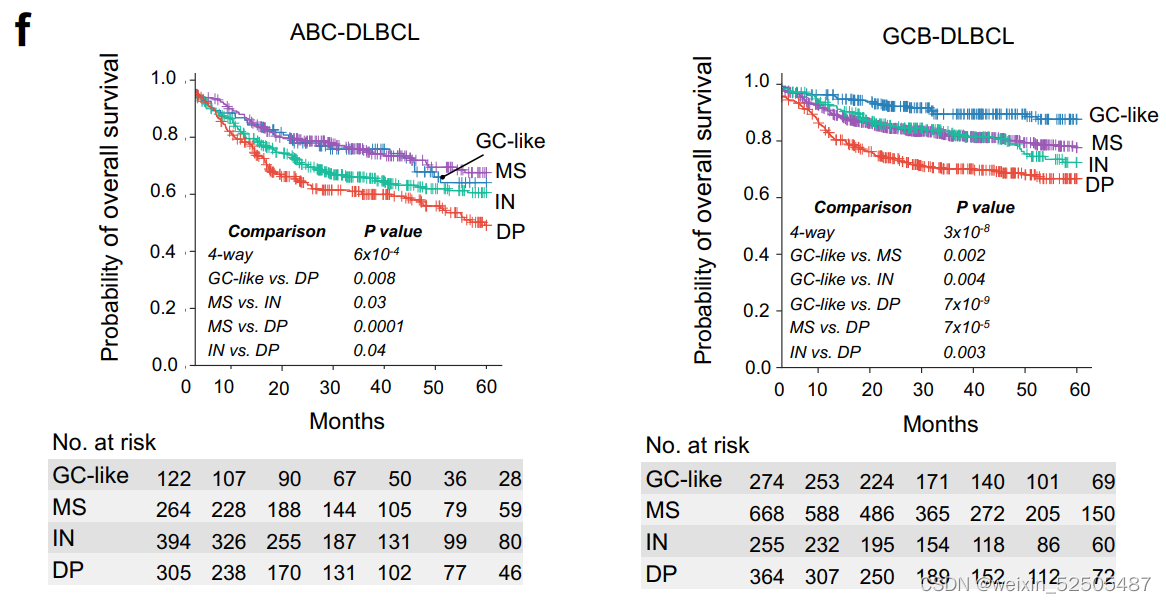
我们在2,646和2,189名DLBCLs患者队列中进一步分别分析了LME类别与OS和PFS之间的关系。所有患者均接受基于利妥昔单抗的化学免疫治疗,导致典型的OS和PFS卡普兰-迈尔曲线。分析LME不同类别的OS从better到poor如下:GC-like, MS (P = 0.03, vs. GC-like), IN (P = 0.0008, vs. MS), and DP (P = 0.007, vs. IN) LMEs。而GC和MS有相同的较好的PFS曲线,其次是DP和IN,呈现出最差的PFS。
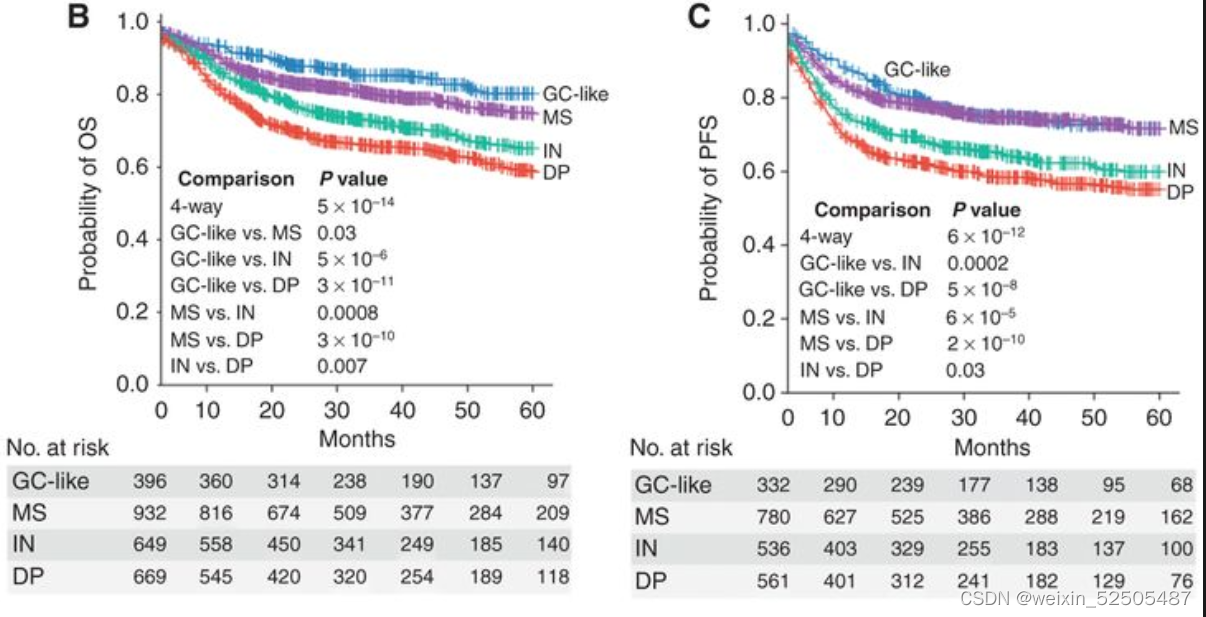
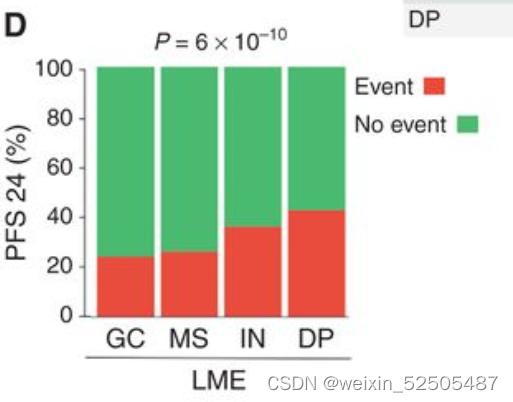
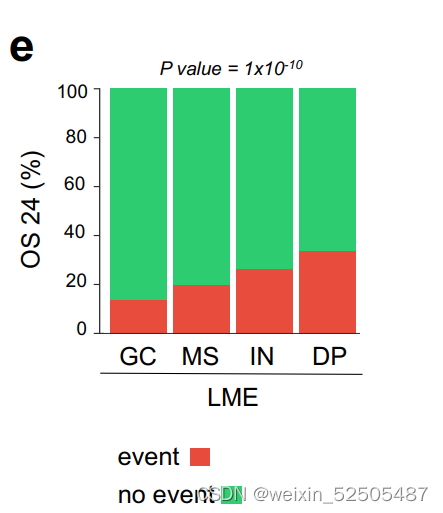
多次测试的Bonferroni correction说明GC-like和MS的OS和PFS处于同一层,对于IN和DP来说PFS也处于同一层。LME分类和24个月时达到OS和PFS的不同比例的病例相关(分别为OS24和PFS24)(OS24 and PFS24, respectively; Fig. 3D; Supplementary Fig. S7E)
当看COO的时候,DP-LME仍然在两种ABC和GCB亚型中OS和PFS最差。但是,在ABC亚型中,最好的PFS和OS都是MS,而在GCB亚型中,最好的LME是GC-like
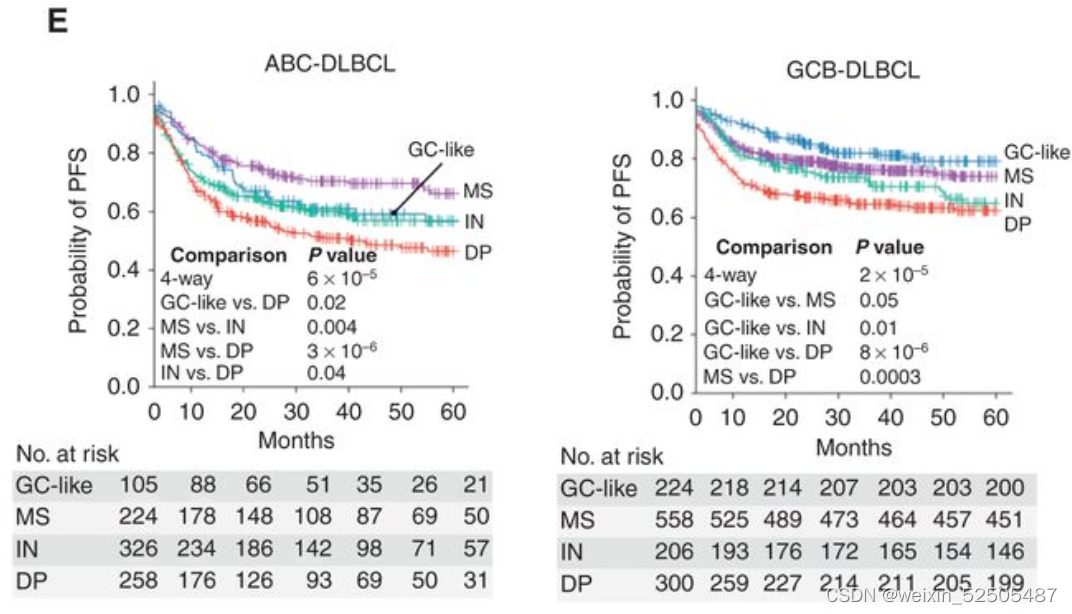
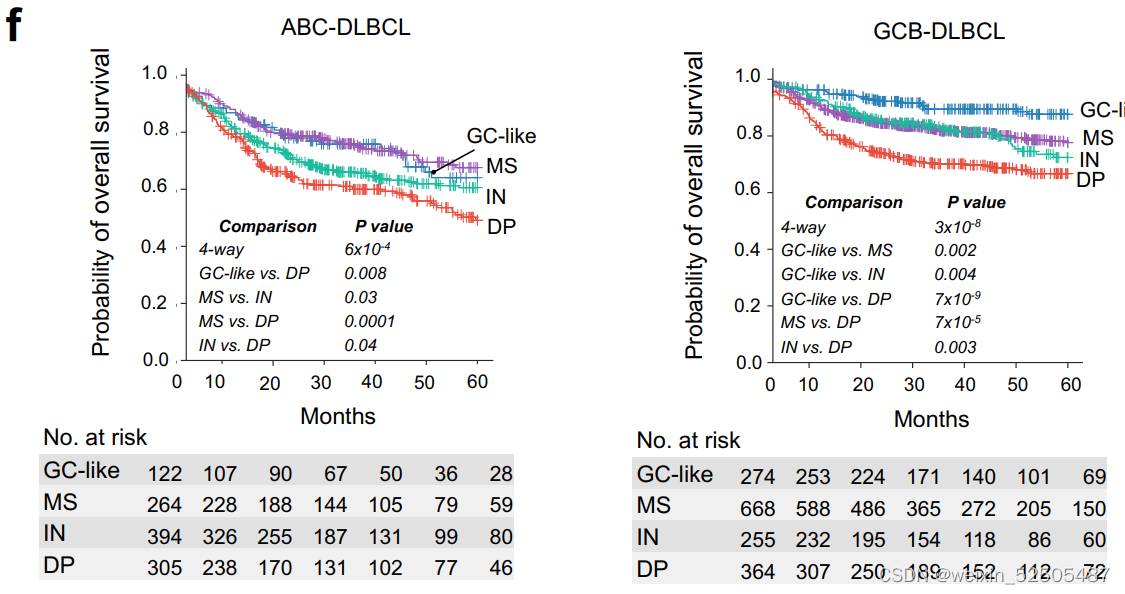
表明了微环境的生物学影响可能依赖于淋巴瘤亚型。以此为根据,单个个体LME细胞亚型在ABC或GCB-DLBCLs中对OS有不同的贡献, (Fig. 3F)
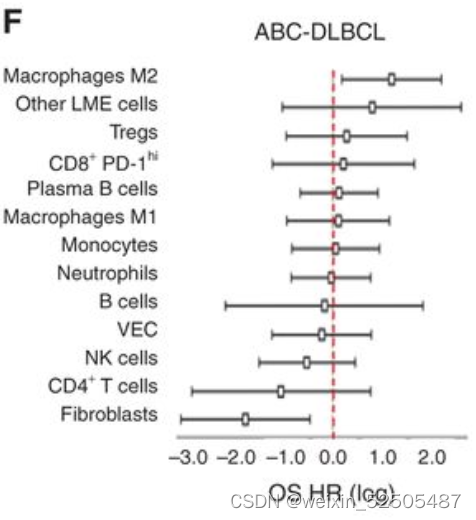
总的来说,GCB亚型中GC-like LME和ABC亚型中DP-LME导致了最好和最坏的临床结局。
针对COO和IPI控制的PFS和OS的影响因素进行多因素回归分析显示,LME亚型仍然具有丰富的信息,GC样和MS样LME具有更好的结局。(Fig. 3G; Supplementary Fig. S7G)

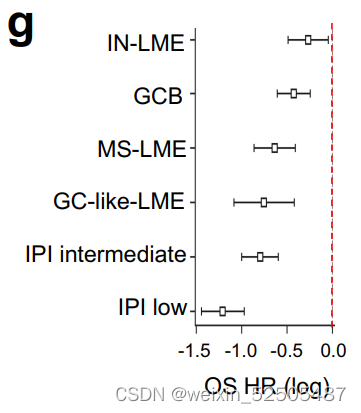
最后,HGBL-DH比MYC或BCL2重排拥有更低的PFS和OS (Supplementary Fig. S7H)。在考虑到LME时候结果可能会有所不同。

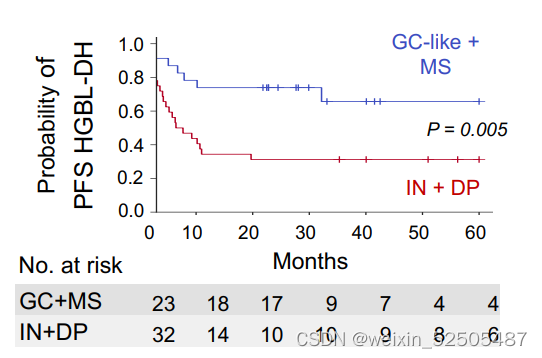
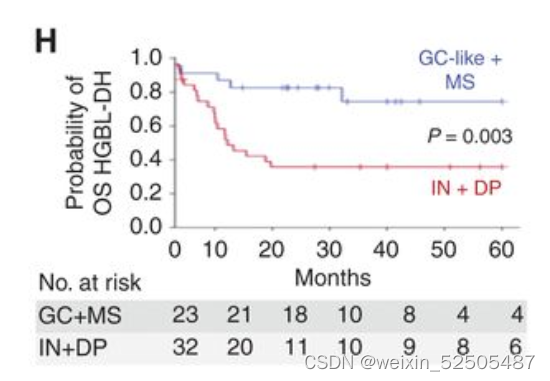
HGBL-DH当处于预后较好的如GC-like和MS-LME的时候比HGBL-DH处于预后较差的IN-LME和DP-LME时OS和PFS要更好。(Fig. 3H; Supplementary Fig. S7H)**
针对COO和IPI以及MYC或BCL2重排控制的PFS和OS进行多因素回归分析显示,LME亚型仍然具有丰富的信息,GC样和MS样LME具有更好的结局。(Supplementary Fig. S7I)
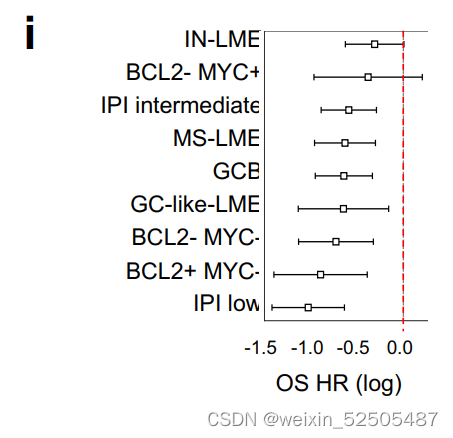
在DHITsig-positive DLBCLs的病例中同样也是适用的,即预后较差的GCB-DLCL具有高级分子特征,包括同时进行的MYC和BCL2重排**(Fig. 3I and J; Supplementary Fig. S7J and S7K)**
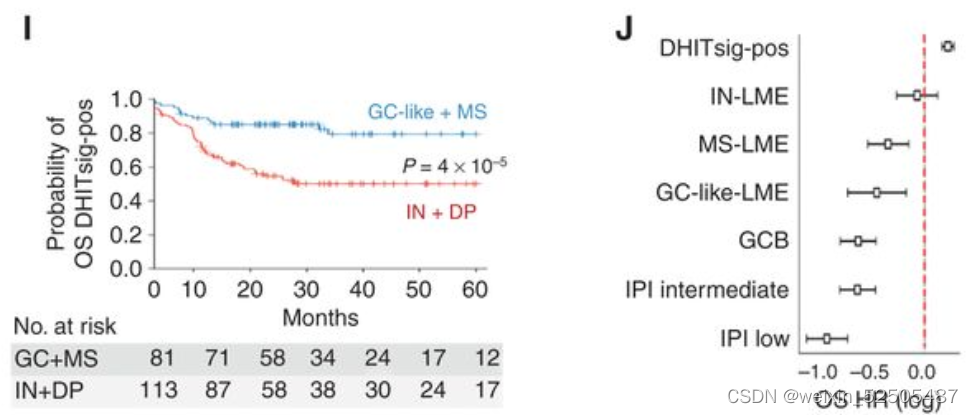
总的来说,以上结果强调了LME category对结果的影响,无论是COO亚组或遗传亚型如何,这表明宿主因素对淋巴瘤生物学和临床表现做出了真正关键和独立的贡献。
LME categories are composed of distinct cellular communities
为了更深层次的了解每一种LME类别的特性,我们通过使用RNA反卷积法计算LME中细胞百分比,扩展了对FGES的community analysis。上述的方法允许我们更加精确的定义亚群以及评估细胞的丰度。我们通过比较和对比在活性扁桃体上进行质谱分析的结果来验证这种细胞组成分析的方法**(Supplementary Fig. S8A)。
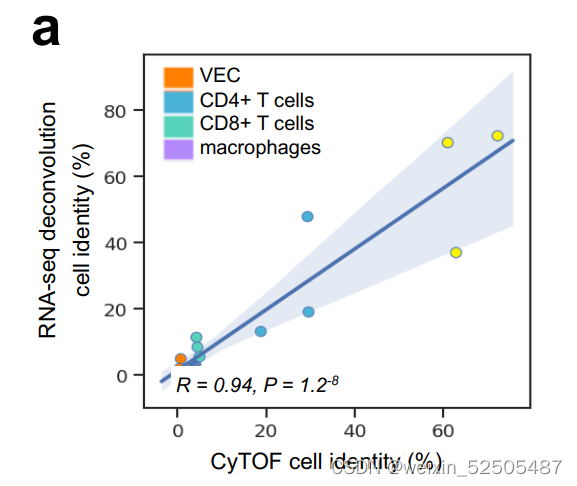
另外对于细胞组成,我们还研究了LME依赖性信号传导和转录途径缺氧诱导因子(HIF),TGFB/SMAD, JAK/STAT, and TNF。
GC-like LME像GCB微环境中的细胞组成,因为其滤泡树突状细胞,淋巴内皮细胞,总T细胞以及几种CD4+ T细胞的亚群包括Treg细胞和Th细胞的含量较高。(Fig. 4A and B)**
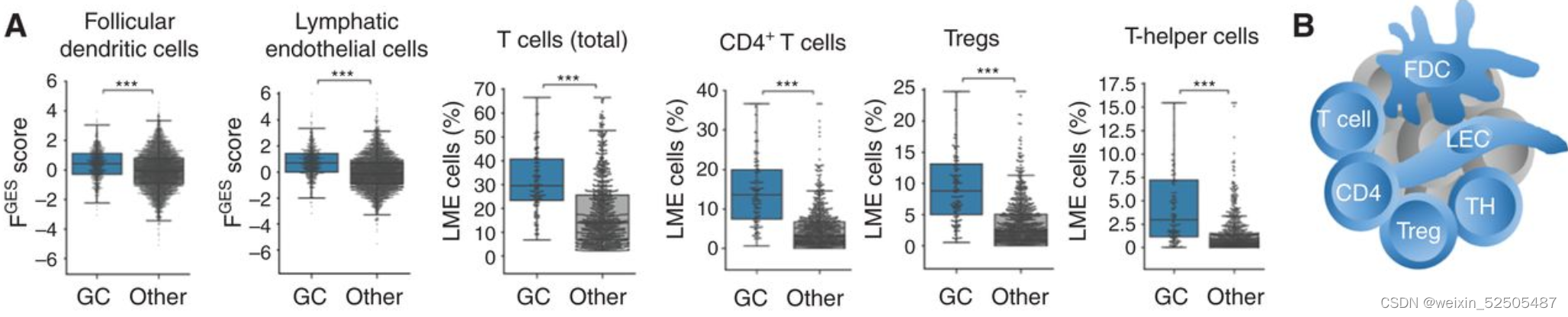
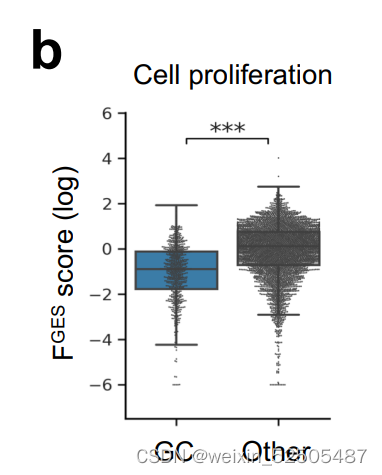
恶性细胞与正常B细胞的比低于其他的亚型。与其他LME类别相比,主要捕捉恶性细胞活性的细胞增殖相关的的FGES较低(Supplementary Fig. S8B)
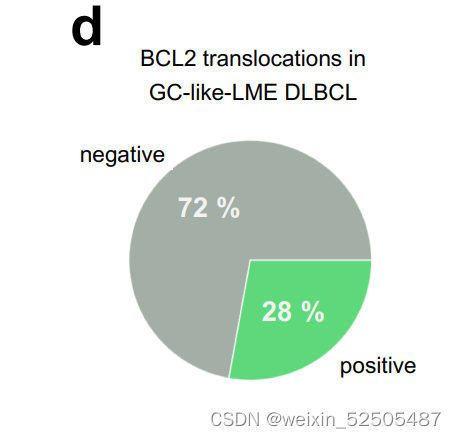
这些淋巴瘤可能代表恶性惰性GC衍生的FL的转化变体,进一步支持这一观点的是28%的GC样的LME病例中含有FL的hallmark BCL2的易位,132个FL的队列中90%的LME也是GC样的**(Supplementary Fig. S8C and S8D)**
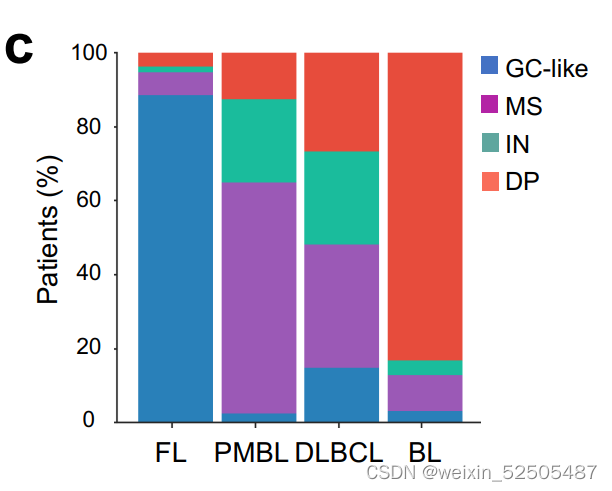
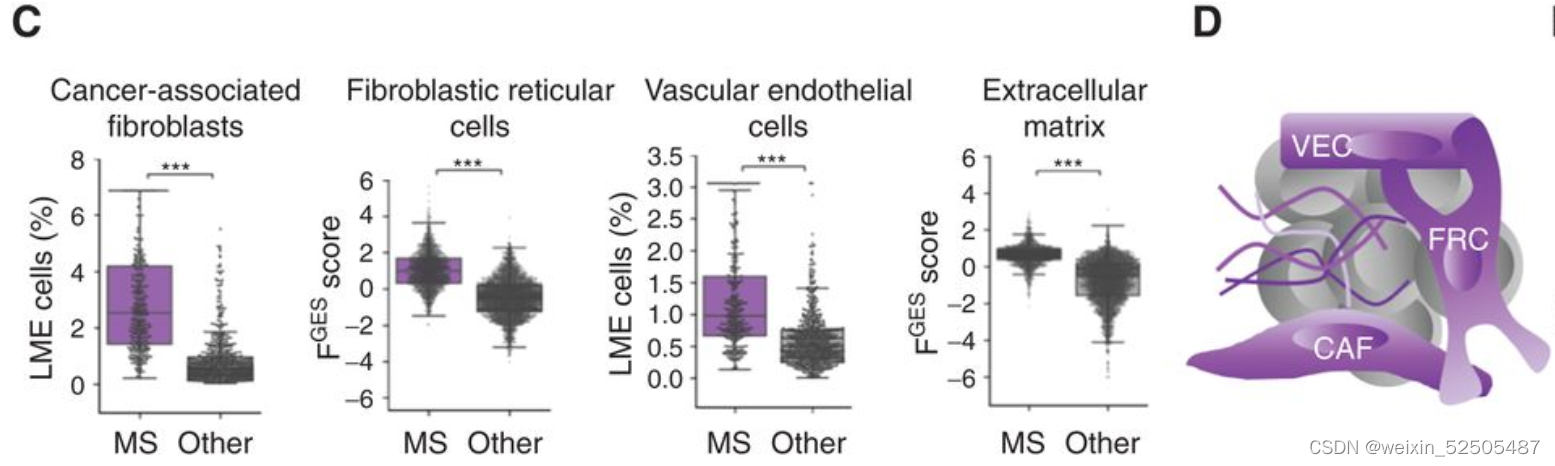
对MS-LME分析显示揭示了血管内皮细胞,CAF,成纤维细胞网状细胞以及ECM占比较高。(Fig. 4C and D)
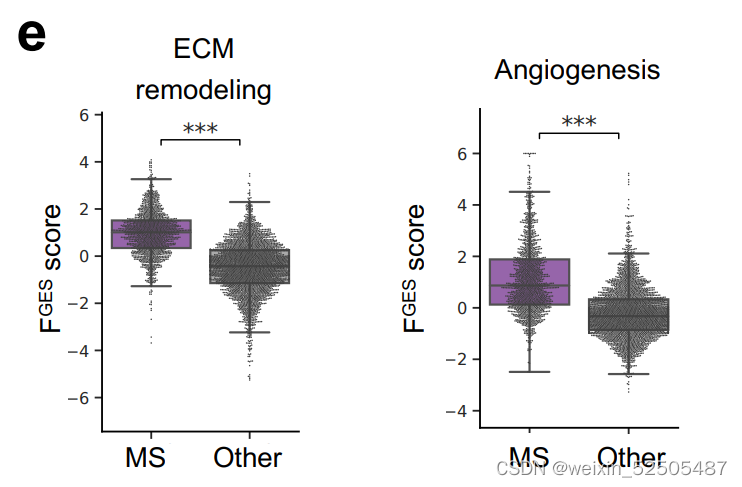
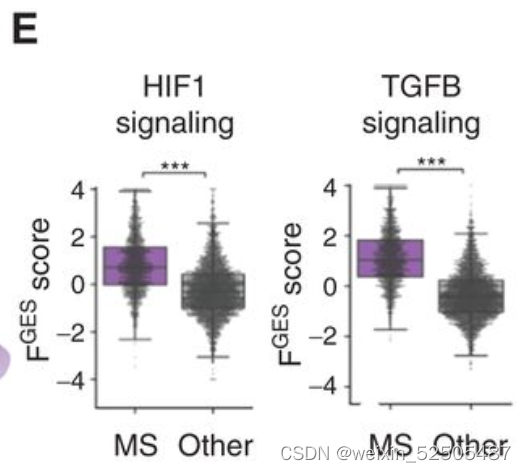
除此之外,MS-LME的FGES主要富集在血管生成和ECM重塑以及TGFB/SMAD 和HIF通路上。(Supplementary Fig. S8E) (Fig. 4E)。上述的通路在DLBCL中预示着预后较好
值得注意的是,与其他LME相比,原发性纵隔大B细胞淋巴瘤的MS-LME category富集较多。(n = 287 total cohort, P = 0.003; Supplementary Fig. S8C)
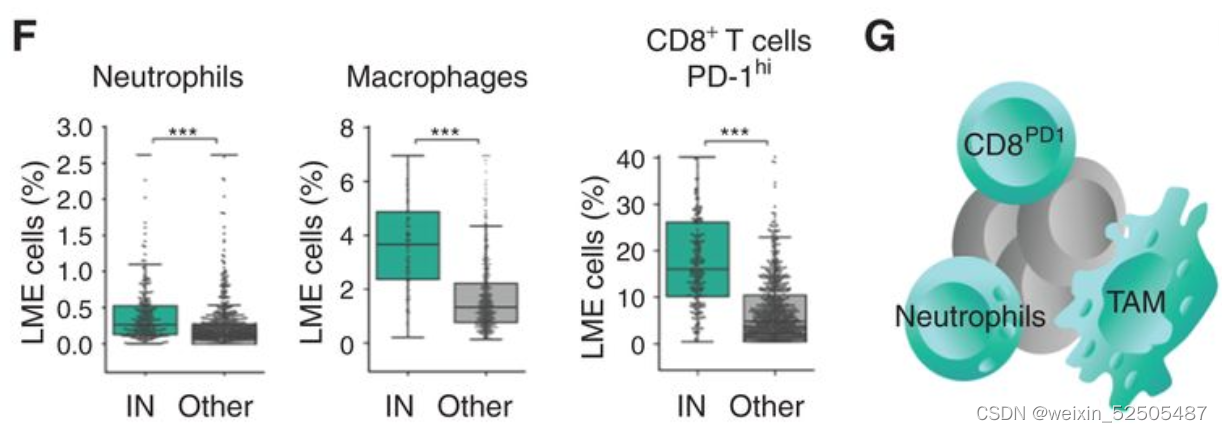
在不利的LME类别中,IN-LME富集在中性粒细胞FGES,常常与较差的临床结局,巨噬细胞,CD8+ T cells(超过总的T细胞或超过CD4+ T细胞)Supplementary Fig. S8F,并且CD8+ T细胞亚群常常有PD-1高表达。(Fig. 4F and G)
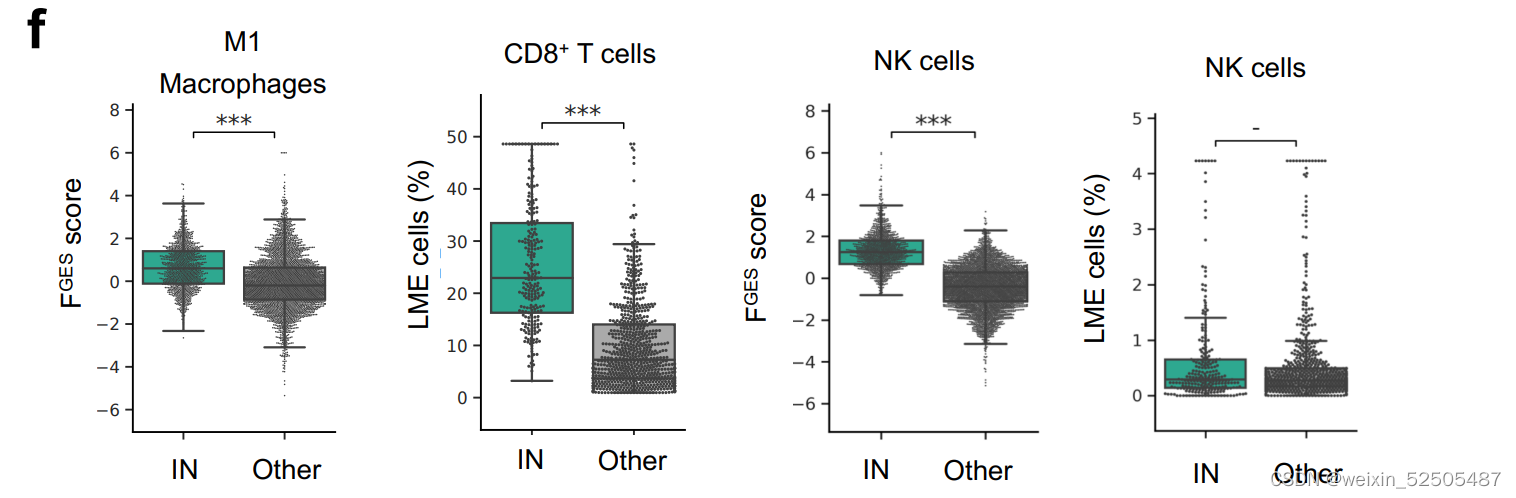
NK细胞的占比在不同的LMEs categories中并没有明显的不同。但是,由FGES介导的NK细胞的活性在IN-LME中明显较高。
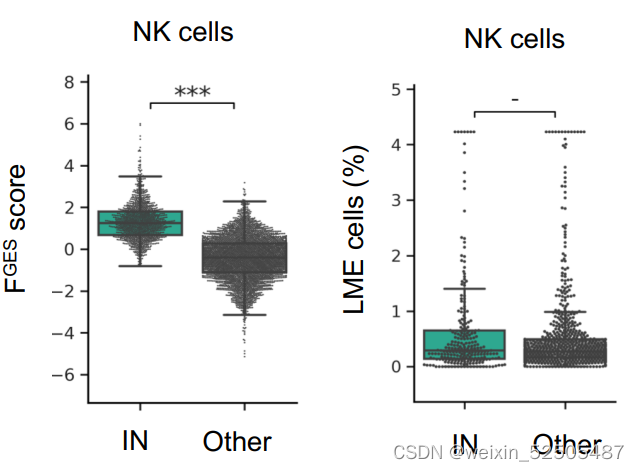
这一发现加上由细胞溶解评分确定的细胞毒性活性富集和T细胞活化标志物的存在表明具有一定程度的抗淋巴瘤的免疫**(Fig. 4H)**
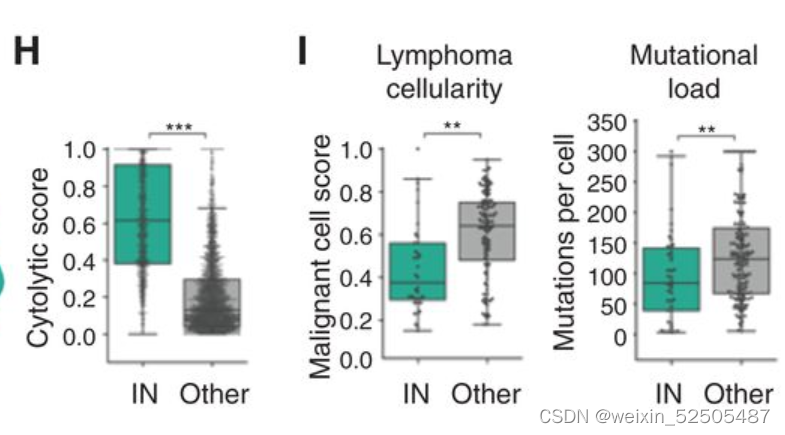
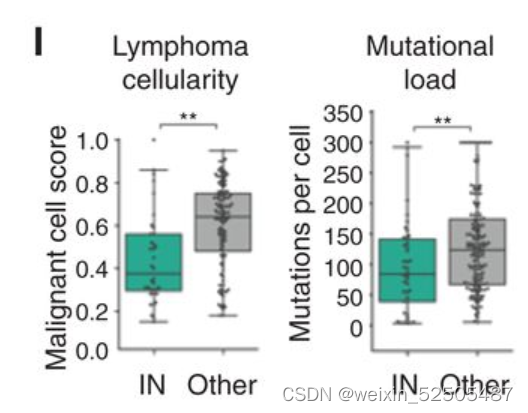
事实上,IN-LME代表最低数量的恶性细胞和最低的突变负荷。(Fig. 4I)可能代表可以逃避肿瘤免疫的淋巴瘤细胞。
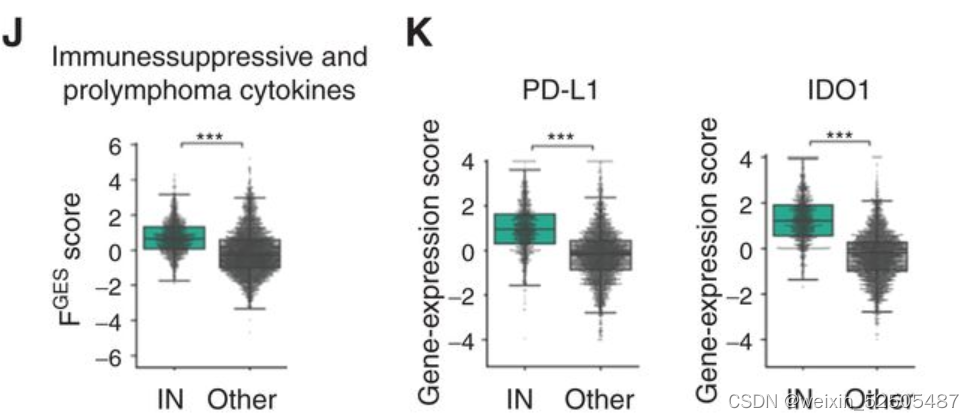
IN-LME可能有助于降低淋巴瘤免疫特征例如免疫抑制和前淋巴瘤细胞因子的富集(包括嗜中性粒细胞引诱剂CXCL8,IL10)(Fig. 4J) 以及免疫检查点分子PD-L1和色氨酸分解代谢酶吲哚啉2,3-二加氧酶1(IDO1)(Fig. 4K)
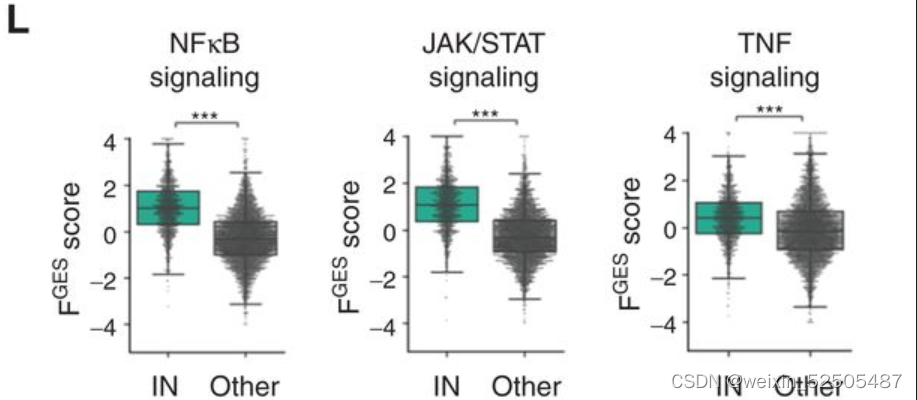
鉴于中性粒细胞和巨噬细胞FGES的存在,这种炎性环境可能预料到。同时在 NFκB,JAK/STAT以及TNF信号通路上相较于其他的LME category的活性更高。(Fig. 4L)
DNA methylation patterning of lymphoma cells contributes to depleted (耗尽) mocroenvironment
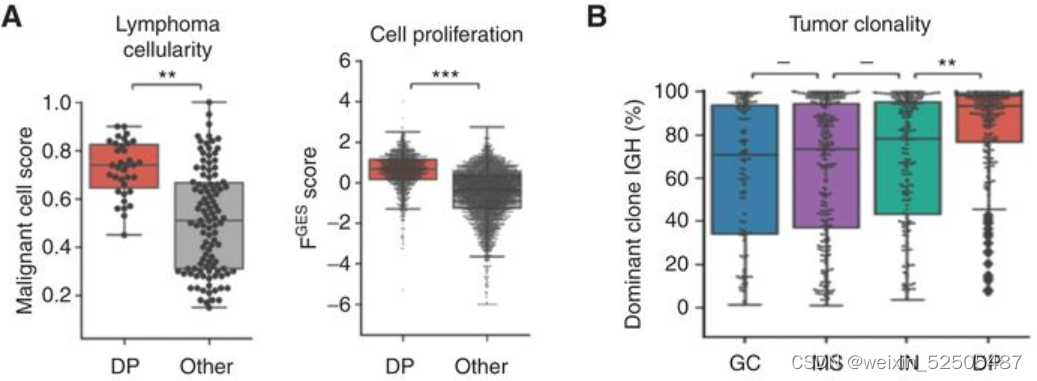
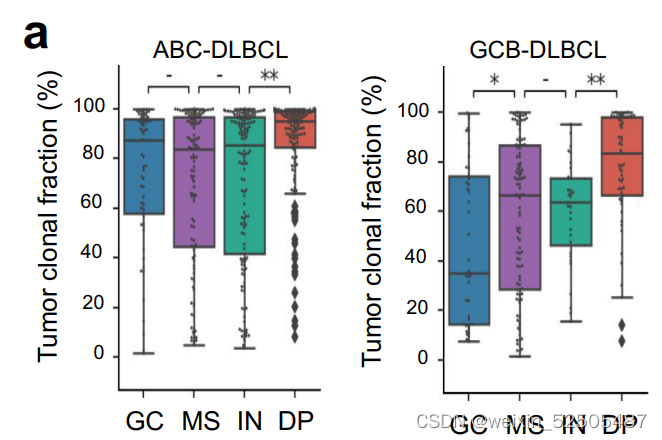
具有DP-LME特征的DLBCL主要以来源于微环境细胞的最小的FGES存在以及更高比例的增殖肿瘤细胞和克隆的肿瘤细胞。 (Fig. 5A) (Fig. 5B; Supplementary Fig. S9A)
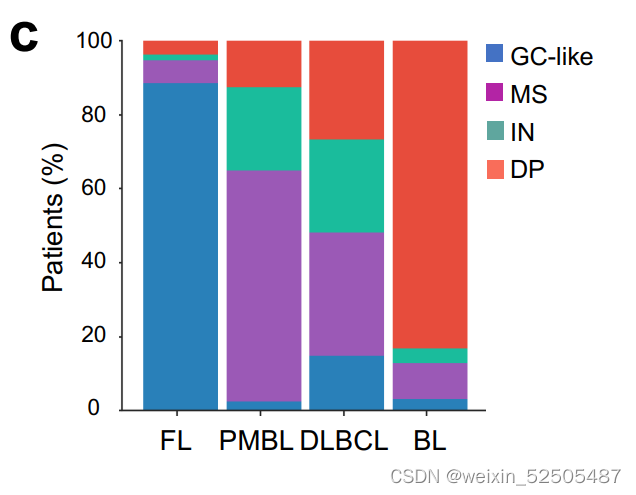
这种LME模式在BL中特别常见**(Supplementary Fig. S8C)**
DP-LME模式以PI3K信号通路的激活为特征 (Supplementary Fig. S9B),表明潜在的治疗的脆弱性。
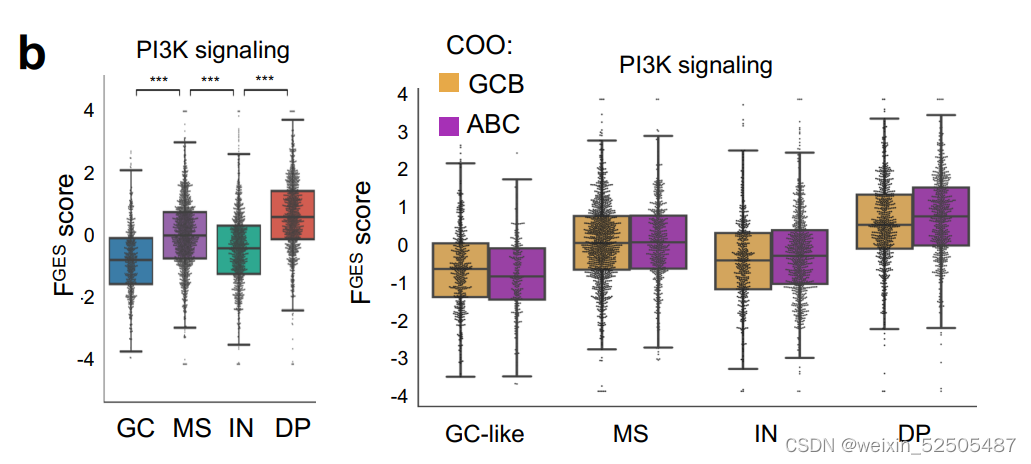
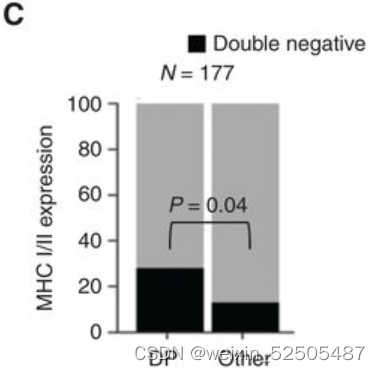
较高的肿瘤细胞的增殖不足以解释来源于微环境细胞的FGES的稀缺性。事实上,DP-LME模式相较于其他的LME category来说大部分的肿瘤细胞MHC Ⅰ和MHC Ⅱ表达阴性。(n = 177, P = 0.04; Fig. 5C)。尽管在该组中没有MHC突变富集。
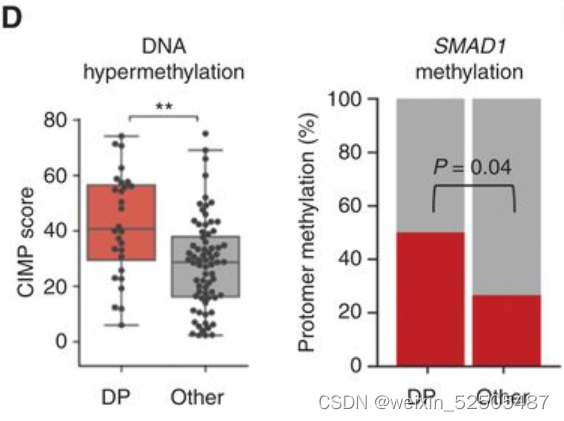
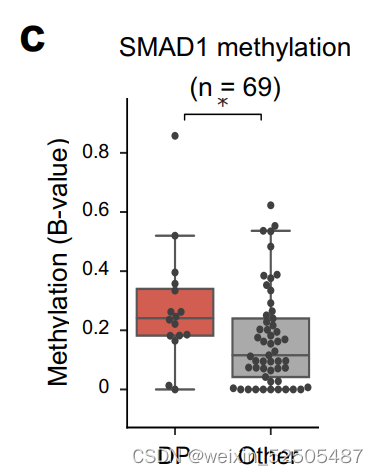
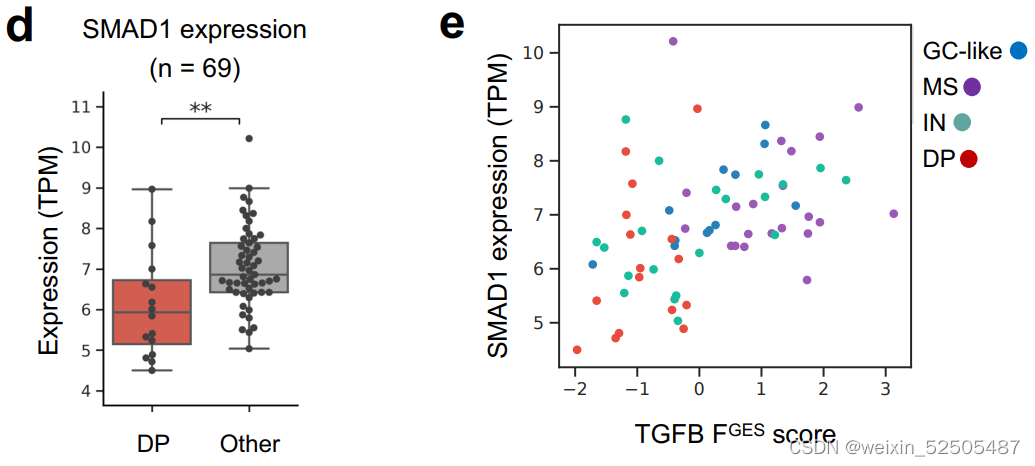
我们和其他先前已经报道的研究证明异常SMAD1启动子 DNA高甲基化允许DLBCL细胞逃避LME驱动的TGFB的配体产生的抗增殖的效应。事实上,对我们的数据以及TCGA数据共100例DLBCLs样本分析显示,global DNA高甲基化以及特殊的SMAD1启动子的甲基化在DP-LME的模式种更高。(Fig. 5D; Supplementary Fig. S9C)
以此为依据,SMAD1表达和TGFB通路活性更低。(Fig. 5E; Supplementary Fig. S9D and S9E)
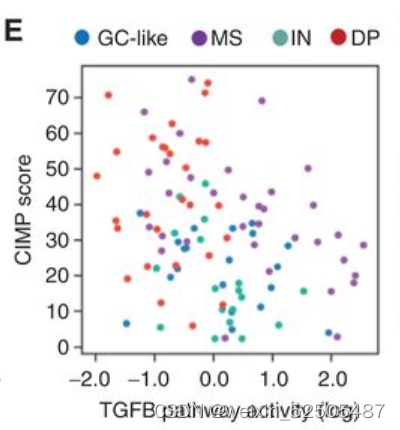
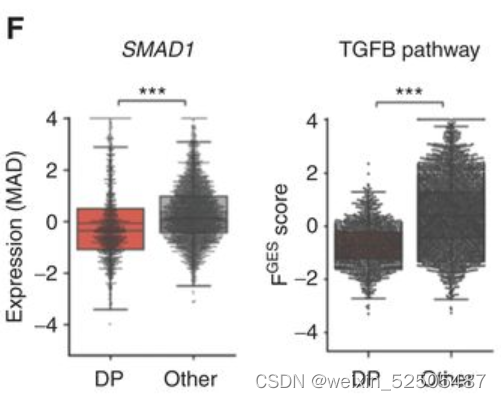
在整个数据集中,DP-LME模式的DLBCLSMAD1表达以及TGFB通路活性更低 (Fig. 5F).
为了判断异常高甲基化对微环境的作用,我们使用 A20 syngeneic murine B细胞淋巴瘤模型,我们将其描述为与DP-LME模式的DLBCL患者具有相似的LME特性的模型。(Fig. 5G)
一旦肿瘤在治疗队列中发展,我们就将小鼠暴露于DNA低甲基化azacitidine或者载体连续四天,然后对肿瘤进行RNA测序和DNA甲基化测序。
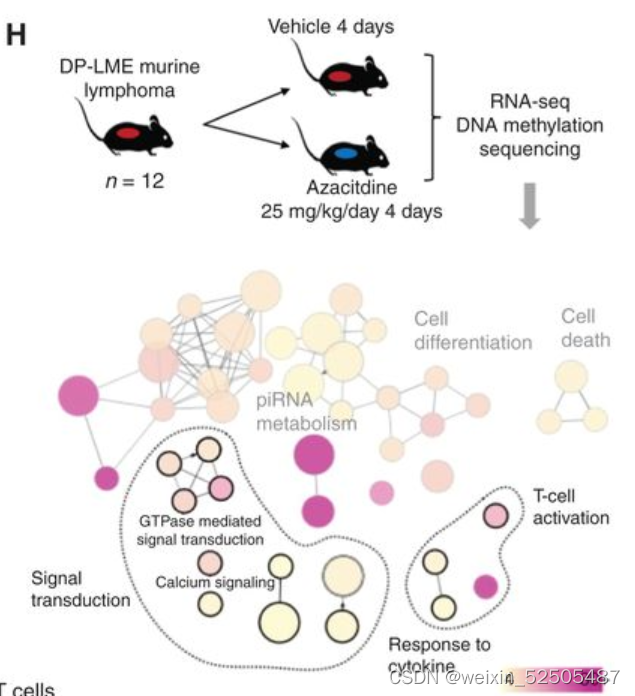
azacitidine的治疗导致基因上调和低甲基化 (Supplementary Fig. S9F and S9G)
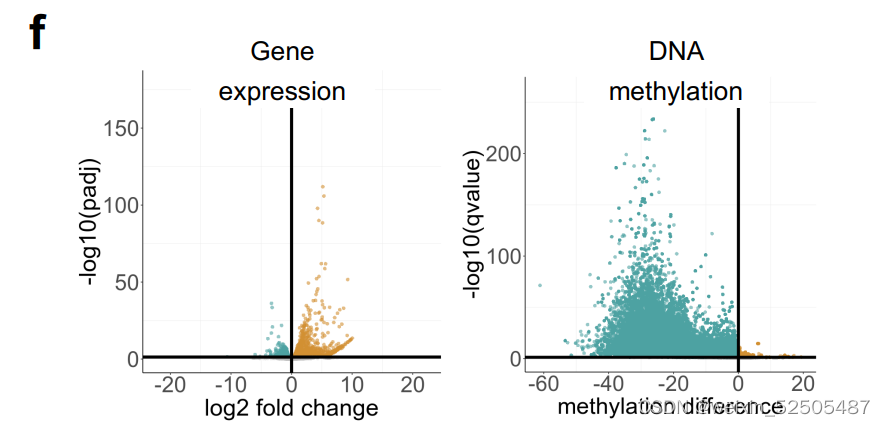
对显著性低甲基化基因启动子以及上调基因进行通路分析,揭示了“cellular reponse to cytokines”,“signal transduction”,and immune response包括“activation of T cells” (Fig. 5H; Supplementary Table S6)
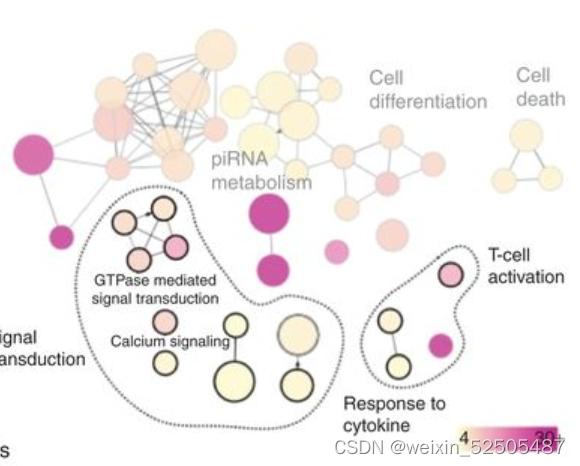
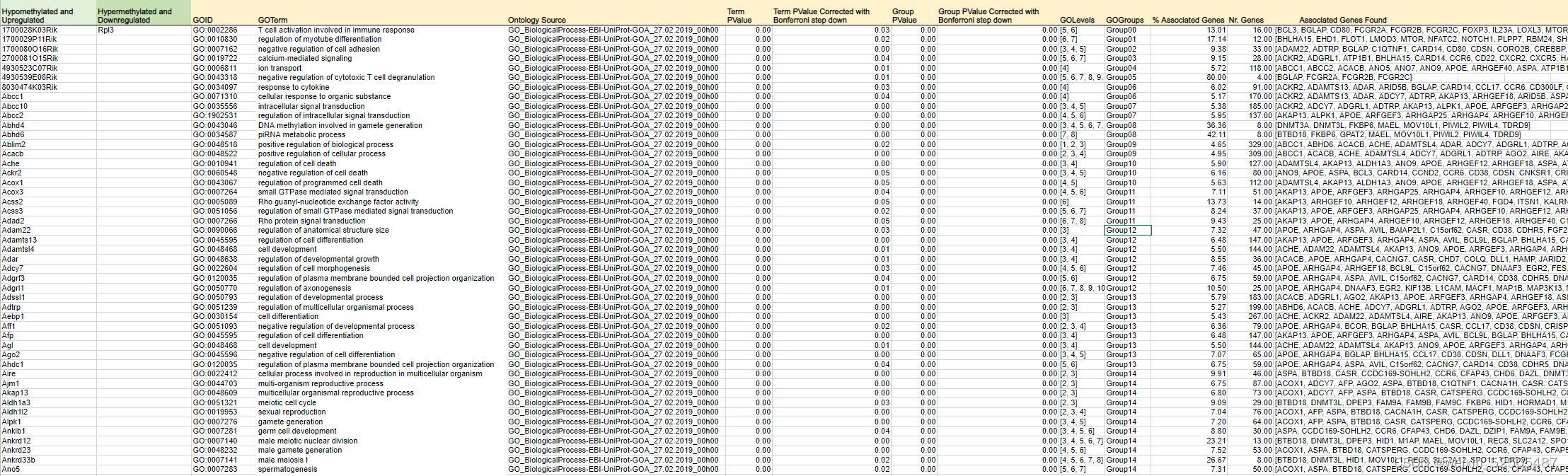
仍然由许多上调的通路例如SMAD和TGFB的信号通路活性。(Fig. 5I)
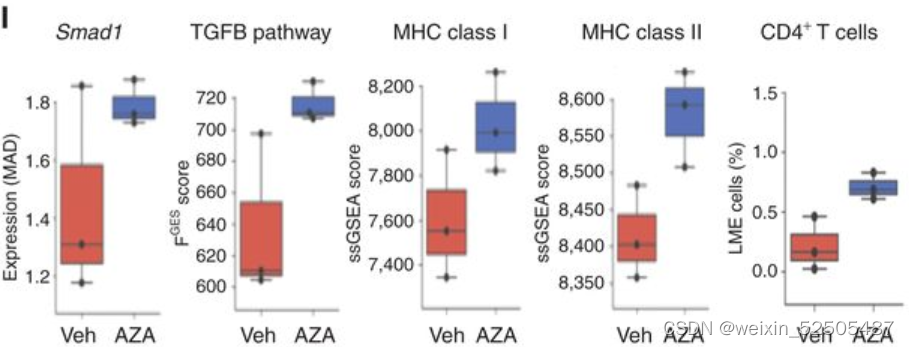
除此之外,通过转录谱反卷积法计算MHCⅠ和Ⅱ表达以及CD4+ T占比增加 (Fig. 5I) 并且通过IHC分析肿瘤浸润性T细胞占比增加[4.7% (q25–75: 2.9–7.3) vs. 18% (q25–75: 14.65–18.9),vehicle vs. azacitidine, P < 0.001 Mann–Whitney test, Fig. 5J]
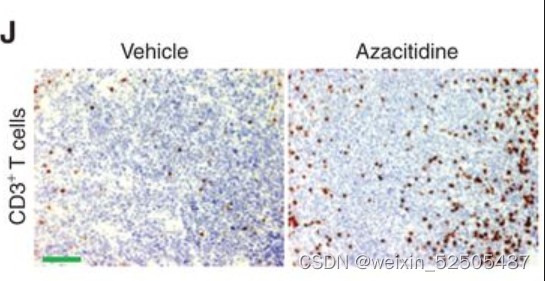
总之,这些数据表明淋巴瘤胞嘧啶甲基化模式可能导致LME耗竭。
LME heterogeneity decreases with disease progression
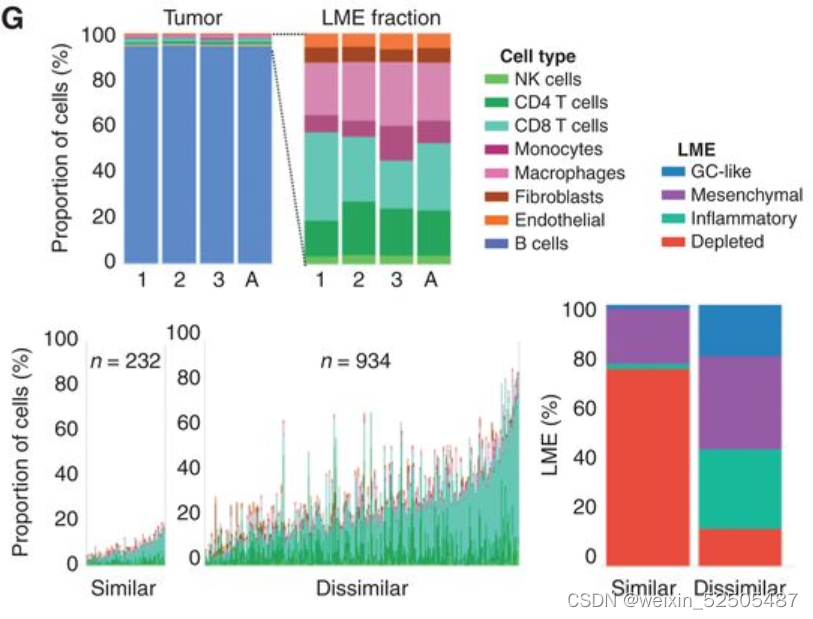
为了去探究在淋巴瘤进程中LME细胞组分改变的概念,我们首先建立了一系列代表淋巴瘤早期和晚期的时间顺序条件。在每一个样本中,我们通过转录谱signature反卷积法构架了ME组成以及通过Shannon entropy metric测量其复杂度,他有两个组件:不同细胞亚型的数量(richness)以及贡献的公平性(evenness)。Shannon entropy metric直接和LME细胞异质性相关。我们获得一例诊断和配对的样本,惰性FL和转化性FL(transformed follicular lymphoma)(n = 5),早期和晚期小鼠淋巴瘤进展(n = 1)以及来自基因工程小鼠模型的癌前淋巴结和淋巴结淋巴瘤,这些模在GCB细胞中表达myc+ BCL2,EZH2突变+bcl2,或KMT2D+ bcl2(n = 3)( Fig. 6A)
**
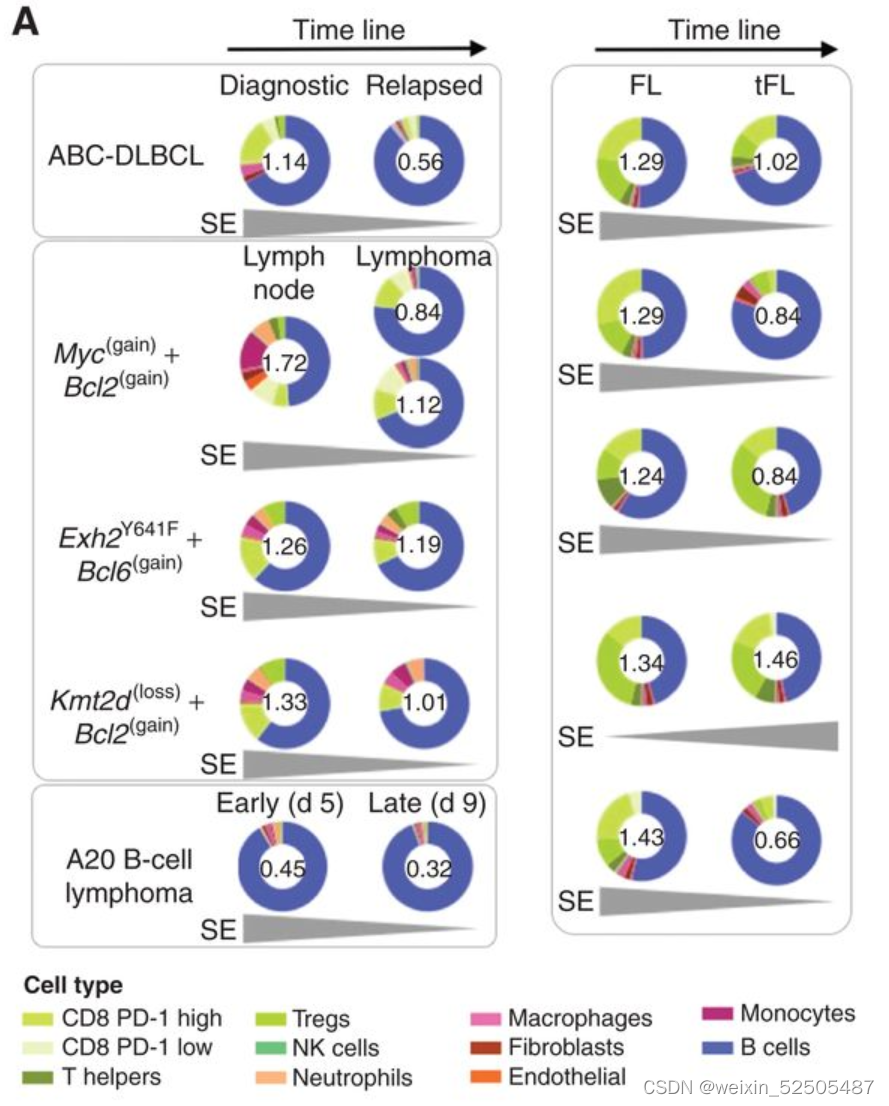
我们观察到LME复杂性(Shannon entropy measured)随着淋巴瘤的发展而降低约40%(95% CI, 20%–60%, P = 0.001, paired t test; Fig. 6A)**
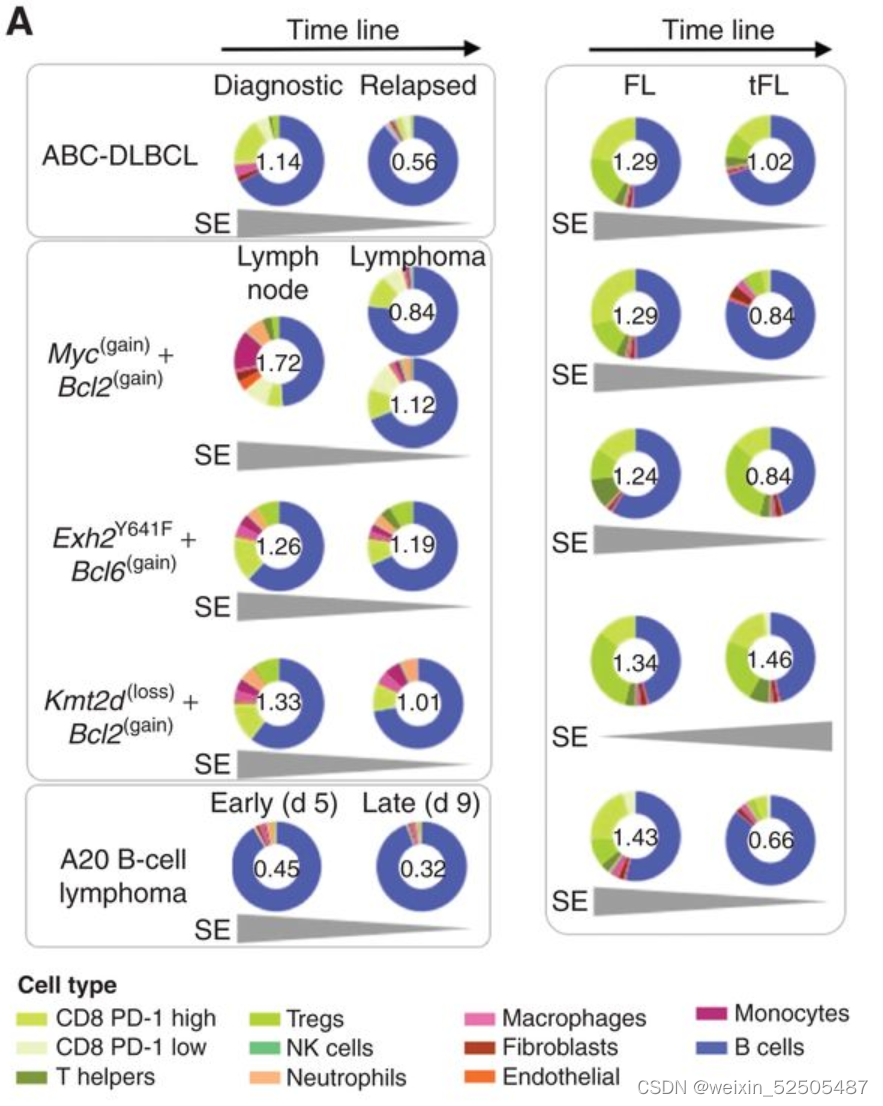
这种效应在T细胞的肿瘤内含量下降方面更加臭名昭著,这与最近的数据一致,该数据与复发的DLBCLs相比,新诊断中的肿瘤T细胞浸润显著减少。为了表征LME中免疫desertion趋势作为跟更大尺度的淋巴瘤进展的函数,我们应用一种伪时间算法来重建LME时间动力学,使用单个样本作为该轨迹中的点。利用25种FGES我们预测2,128 ABC和2,867 GCB样本
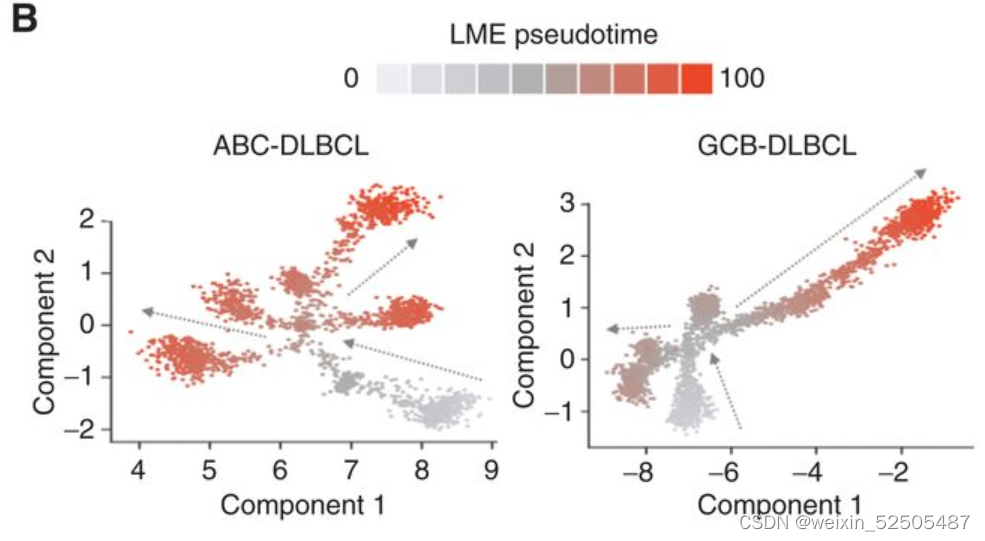
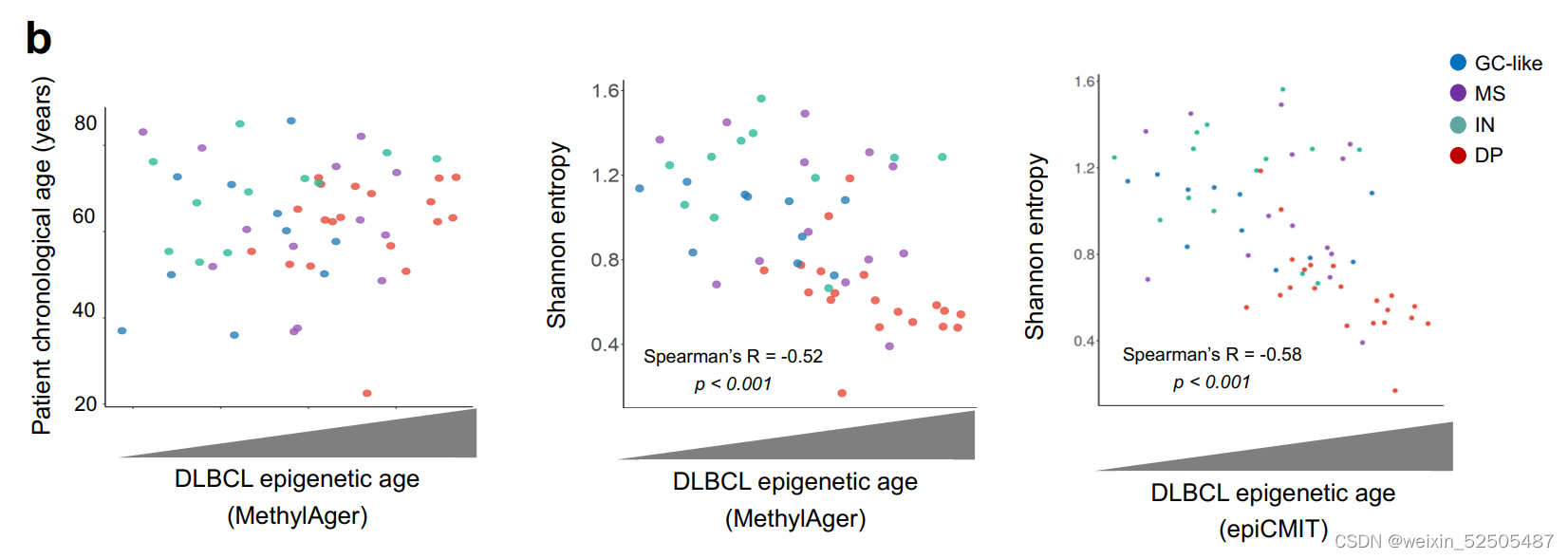
类似于GC免疫微环境的细胞构成的免疫富集的GC-LME被认为是起点。对于单个DLBCL样本的LME Shannon entropy显示出与伪时序和淋巴瘤表观遗传的阶段相反的联系**(Fig. 6C; Supplementary Fig. S10A, R = −0.51, P < 0.001)。 (Supplementary Fig. S10B, MethylAger R = −0.52, P < 0.001 and epiCMIT R = −0.58, P < 0.001)
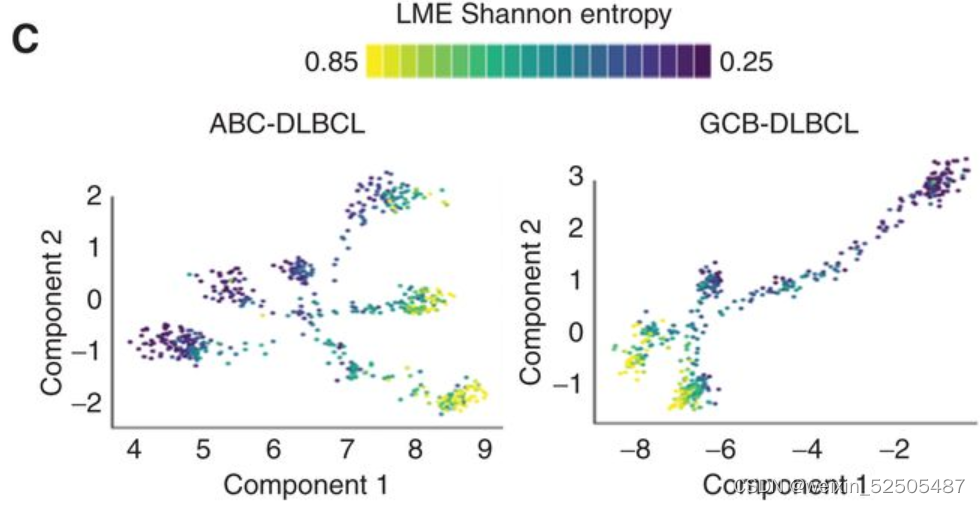
来自两个队列中的FL,转化性FL以及新诊断的DLBCL在GCB-LME拟时序分析显示在低LME复杂性的更高的LME拟时序之间的联系。(Fig. 6D)**
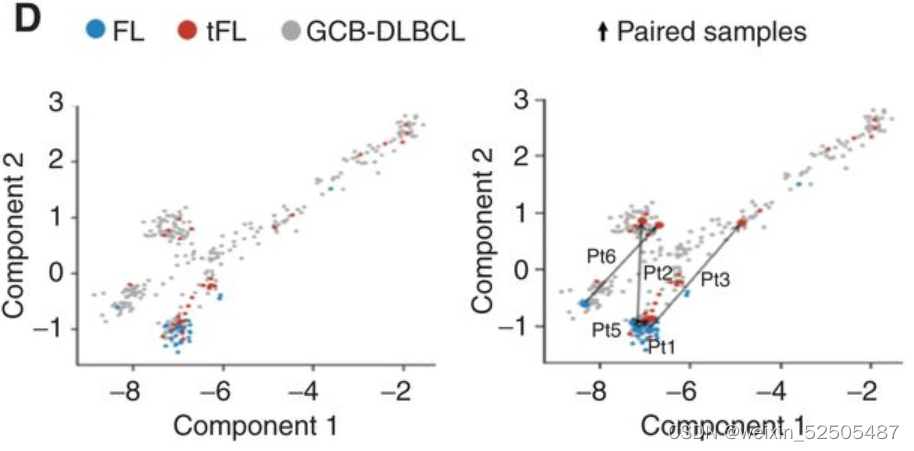
此外,五个配对FL和tFL样本的投影也显示出这种趋势。和肿瘤配对样本相似,LME拟时序的进展和肿瘤浸润T细胞的减少相关。 (Fig. 6E)
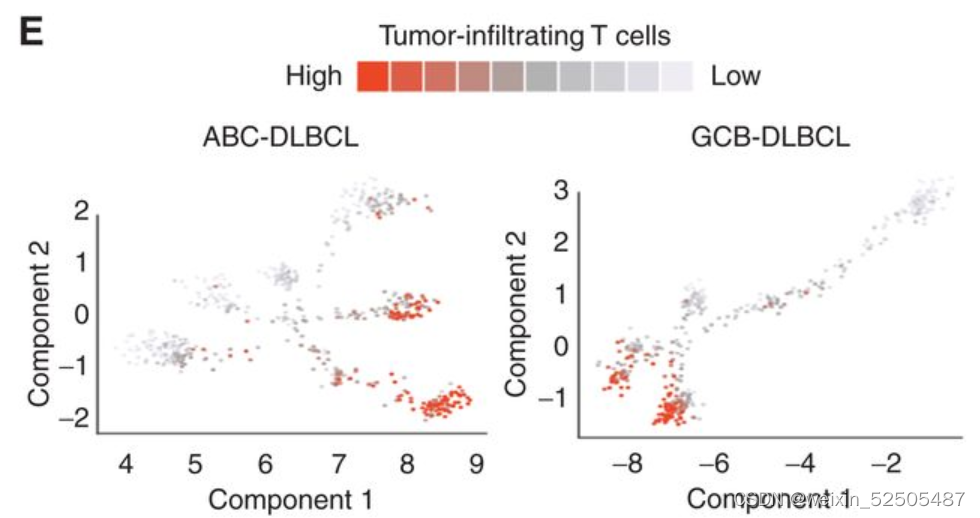
上述的notation表明了淋巴瘤细胞可能获得或选择浸润性T细胞减少的机制,导致了DP-LME
CAFs produce a lymphoma-restrictive ECM(细胞外基质)
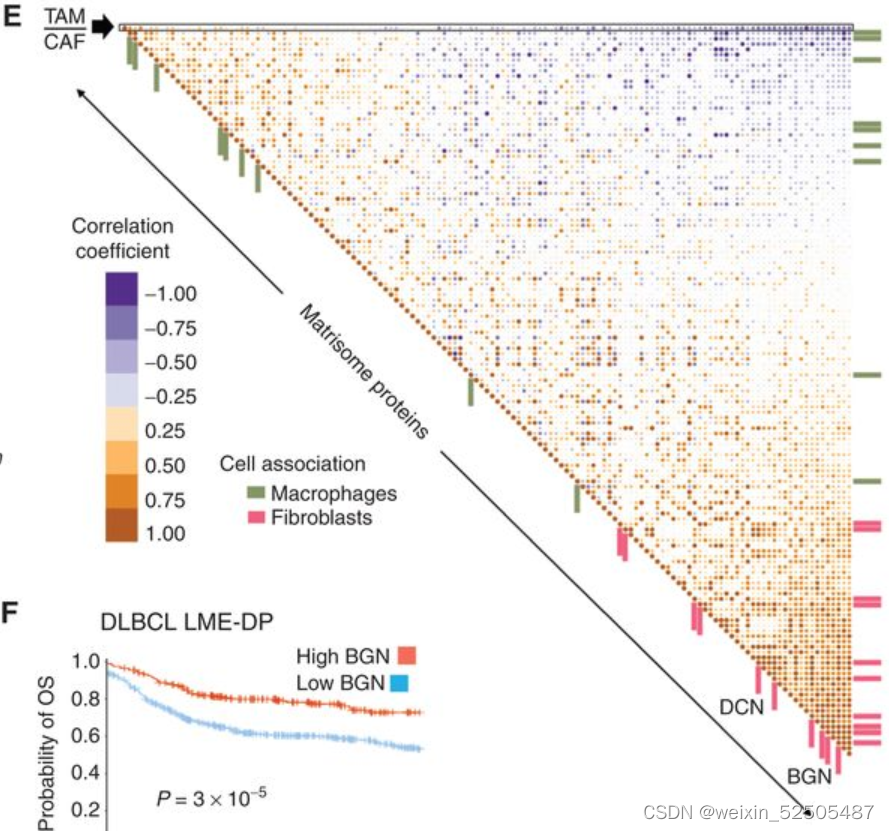
在肿瘤微环境中,ECM主要是由TAM和CAFs的平衡活性活动产生和重塑。我们探究TAM FEGS和CAF FGES的比是否会影响预后。TAM/CAF和高比例的死亡相关(HR:9.1),在所有LME的亚型中观察到一定程度的效应,但是在DP-LME中特别强烈 (HR:4.4)。(Fig. 7A)

为了去确定ECM组分中最可能与CAFs/TAM的活性最相关,我们将发炎的扁桃体的基质体作为对照组,对18个原发性DLBCLs的ECM进行了无偏倚蛋白分析。(matrisome; Fig. 7B)
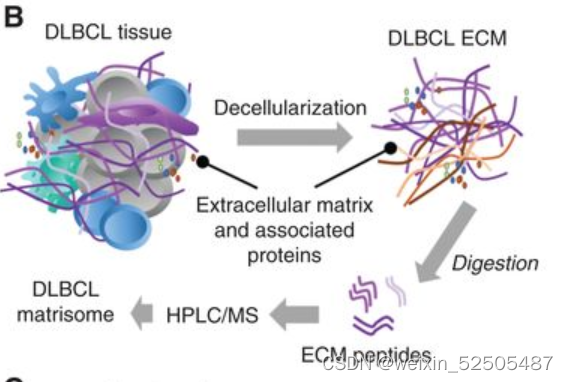
然后我们使用这些DLBCLs样本中组织量充足的14个样本进行RNA测序来评估TAM/CAF,LME以及COO。我们在DLBCL基质体中鉴定了131种蛋白质,这些蛋白质分布在糖蛋白,EVCM调节剂,胶原蛋白,ECM附属物,分泌的细胞因子和蛋白聚糖,富集在三种母体型中 (Fig. 7C)。
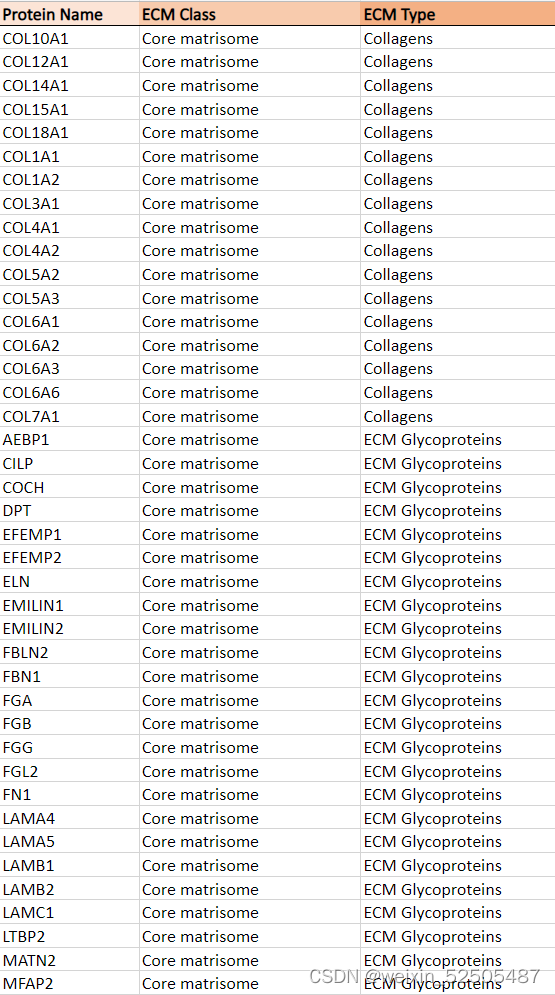
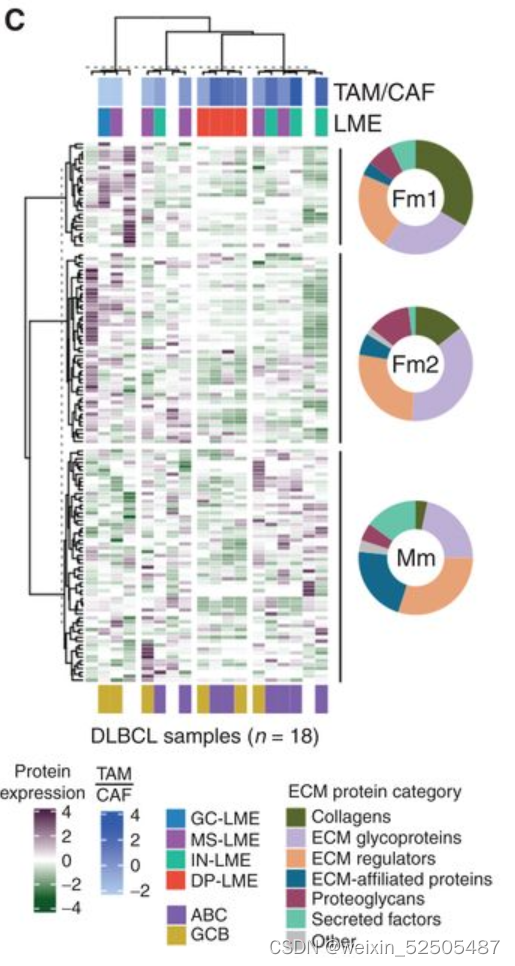
两种母体型 (termed Fm1 and Fm2)与更高水平的CAFs和更高水平的胶原蛋白和蛋白聚糖相关 (Fig. 7C),而含有较高比例的ECM附属物和分泌蛋白的第三种母体型具有较高的TAM的表达(因此称之为Mm),反应FGES,DP-LME DLBCL拥有更低的ECM丰度。为了去了解DLBCL ECM的细胞组成,我们将基质体基因与他们在15个LME subtypes和DLBCL细胞中的中值表达相关联,以获得根据其细胞类型表达的概率**(Supplementary Fig. S11A; representative examples)**
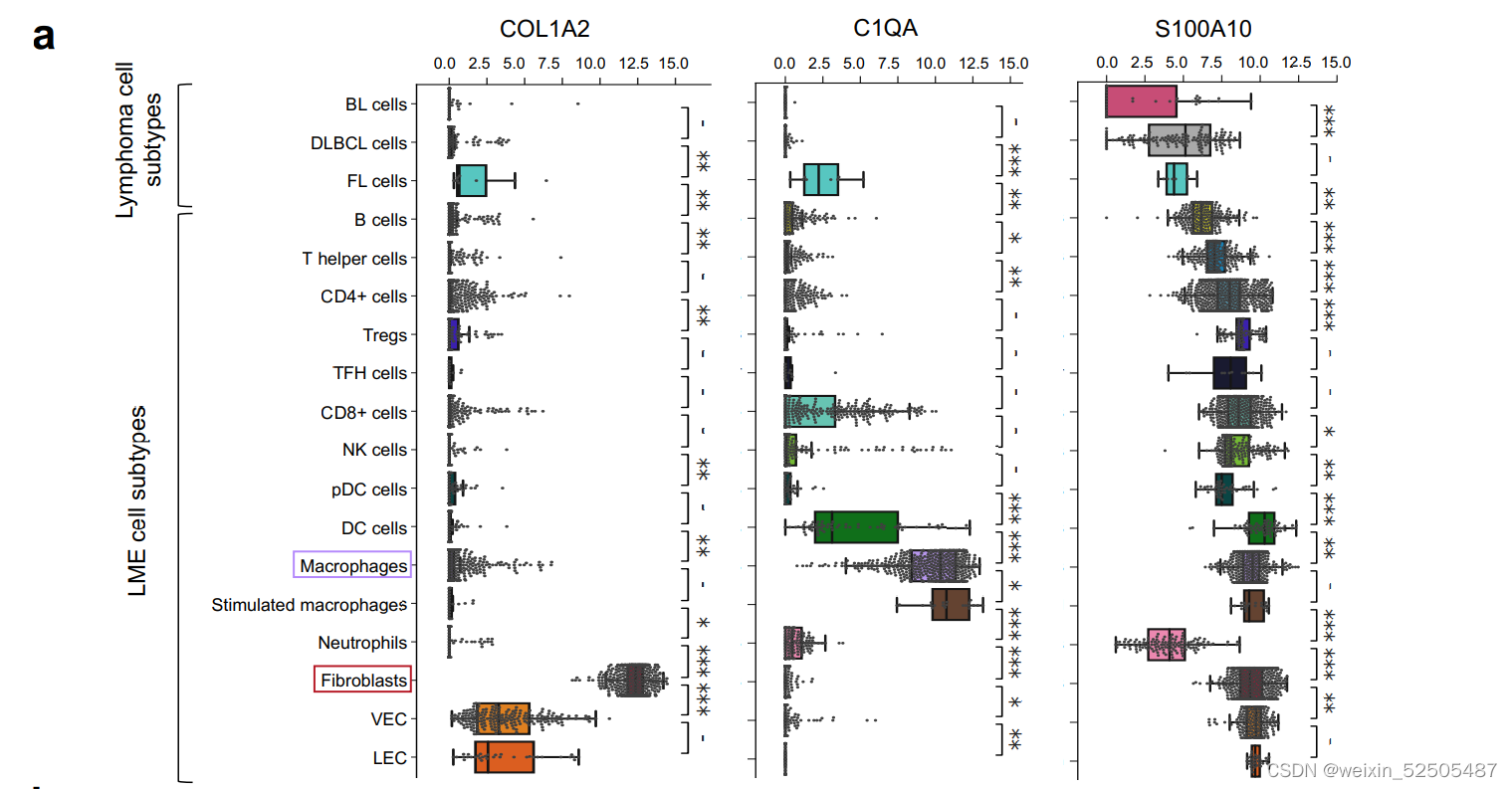
DLBCL基质体基因从成纤维细胞与巨噬细胞中表达的最高概率进行排名,以获得它们对ECM代表性的贡献。(Fig. 7D; Supplementary Fig. S11B)
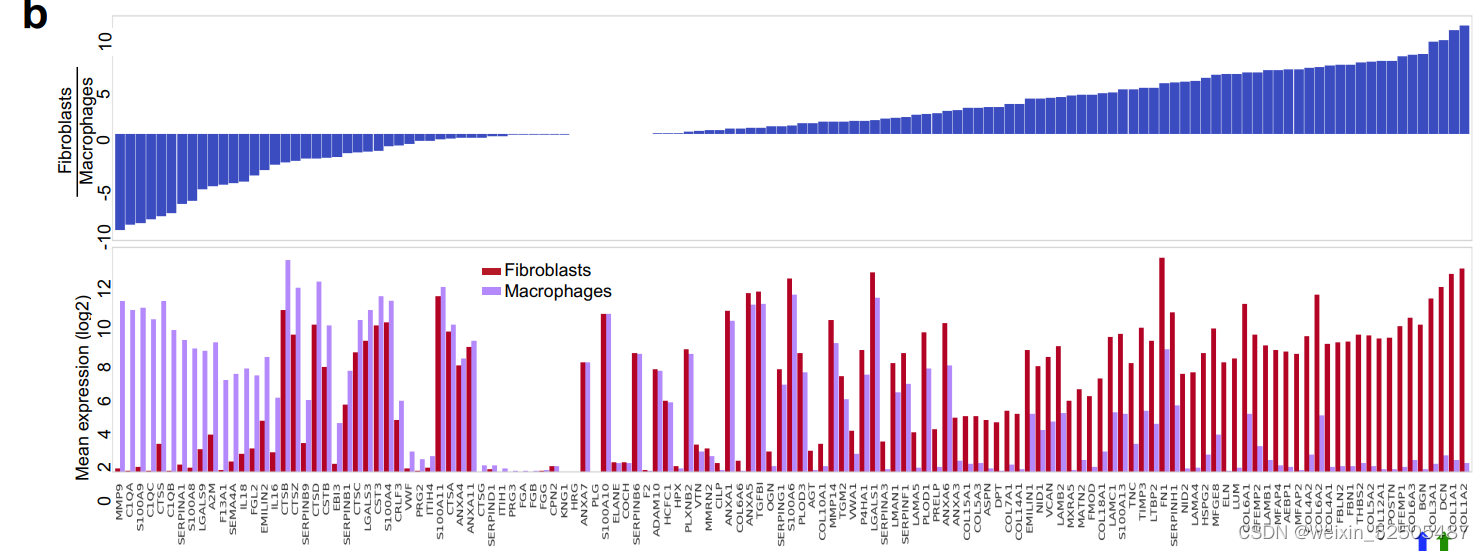
胶原蛋白与蛋白聚糖酯(DCN)以及大聚糖(BGN)的表达与CAFs具有较强的联系**(Fig. 7D)。相反,这些胶原蛋白以及蛋白聚糖的ECM蛋白丰度与FGES对CAFs的估计以及成纤维细胞比例显著相关(R = 0.9, P < 0.001, n = 9 patients; Supplementary Fig. S11C)**
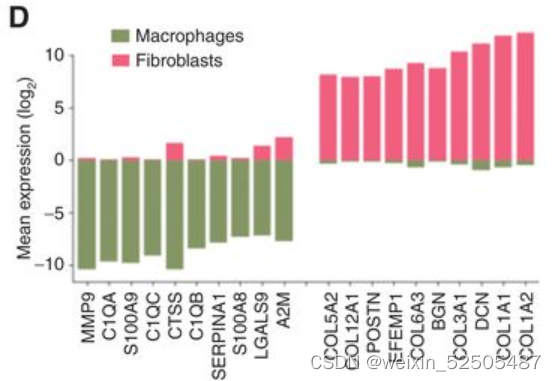
以此为根据,因此,在14例患者中,CAF相关蛋白在DLBCL基质体中富集,该部分与较低的TAM / CAF比率相关**(图7E)。
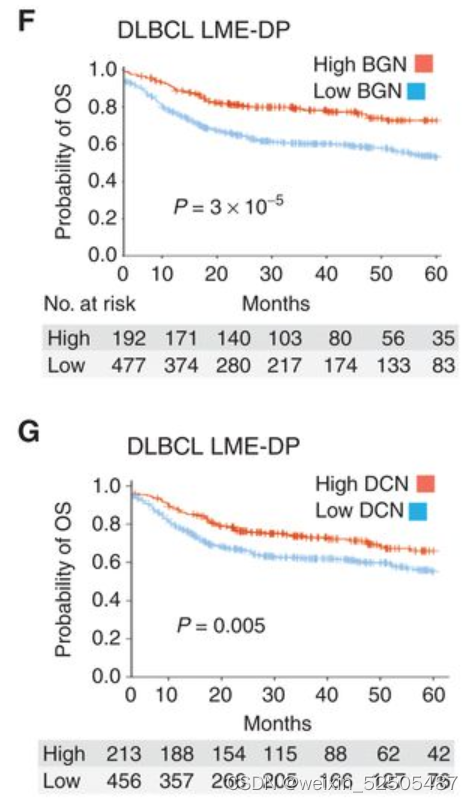
最后,和CAF FGES相似,CAF中ECM的蛋白聚糖DCN和BGN主要与好的预后相关,特别是在DLBCL LME-DP的患者中。 (Fig. 7F and G).
通过短程和远程效应结合,微环境中得蛋白聚糖具有影响肿瘤生长得潜力。为了去判断其在DLBCL中得影响,我们首先在DP-LME ABC-DLBCL中开发了两种PDTX模型,这些模型在小鼠中保留了上述得特征。(Fig. 7H; Supplementary Fig. S11D)**
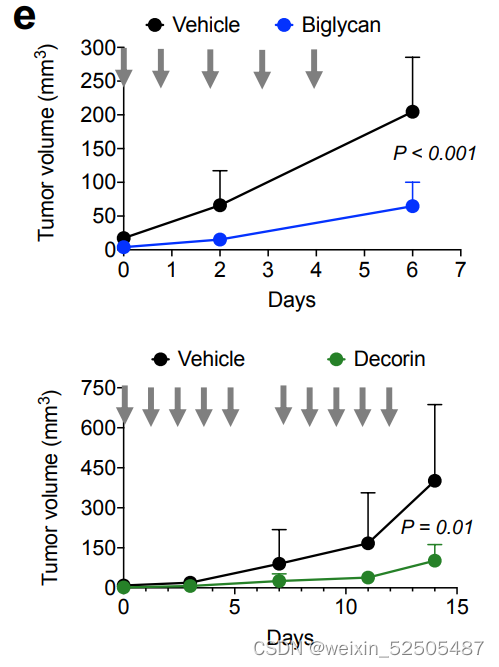
一旦PDXs完全建立,我们就用载体,DCN和BGN腹腔注射小鼠,并遵循肿瘤生长曲线。我们发现与载体相比,两种蛋白聚糖显著的延缓了肿瘤的进展**(P = 0.009 and P = 0.039 for BGN and DCN, respectively, in PDTX1 and P = 0.0004 and P = 0.01 for BGN and DCN, respectively, in PDTX2, Fig. 7I; Supplementary Fig. S11E)**
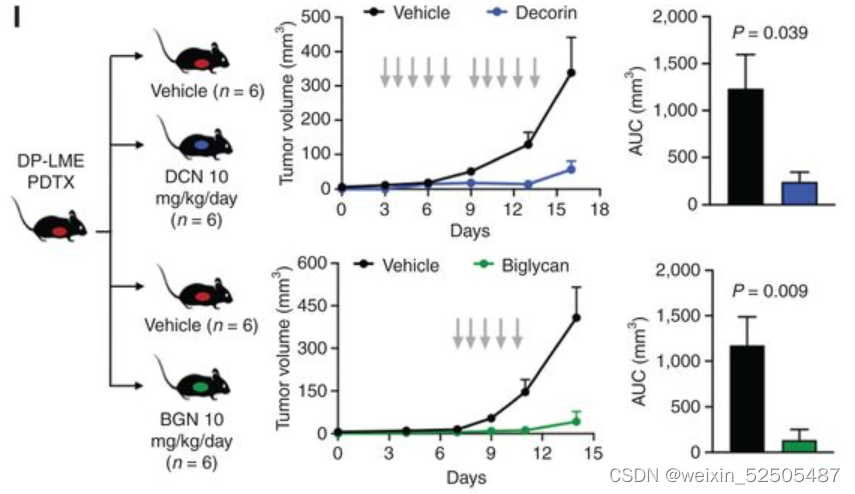
蛋白聚糖在分离的PDX源性的淋巴瘤细胞中没有显示出统计学上显著的抗增殖作用。尽管没有证据表明所施用的蛋白聚糖直接掺入到PDTX ECM中,但是如前所述,BGN给药导致了PDTX2 ECM中DCN含量增加20%(P = 0.032, vs. vehicle) 。总的来说,*上述结果表明了短程和远程效应结合来看蛋白聚糖具有抗肿瘤增殖的效应。 *
Discussion
我们通过分析大量的基因表达谱和技术创新,包括开发微环境衍生的FGES,通过蛋白质组学分析ECM组成以及建立PDTX模型。与细胞反卷积法类似,我们的方法反应了肿瘤微环境中细胞的组成,但也提供了有关其功能的宝贵的信息。这些方法允许我们描述具有不同的临床和生物学内涵的四种基本的DLBCL LME分类。除此之外,通过将淋巴瘤细胞中的突变和表观遗传的突与LME的变化的时间顺序叠加在一起,我们能够推断出癌细胞-LME共同进化的潜在的机制。最后,我们确定了LME中存在的新型的潜在治疗的靶点,包括以蛋白聚糖DCN和BGN为代表的ECM蛋白。
我们的研究代表了在DLBCL中LME的综合性的分析,同时将微环境和恶性细胞群的特征整合到疾病的预后中。开创性的转录组学的研究,检查宿主因子以及DLBCL中基质,确定微环境为疾病生物学的相关组成部分。最初的研究依赖于识别肿瘤样品中基因表达谱的差异。我们的研究通过丰富的淋巴瘤细胞混杂转录组背景中提取微环境的特征来完善这一分析。从而能够定性和定量分析本体组织转录组内的微环境的细胞。我们进一步优化了这一项技术以包括功能特征,这一功能特征包含确定的细胞类型包括T细胞,NK细胞毒性,以及某些与细胞类型不唯一相关的途径,如ECM重塑和细胞因子分泌。这些和功能特征的关联表明了LME中存在功能性细胞的生态系统,并提供了对微环境这种疾病中的作用的全面见解。这种方法提供了比单个细胞群定量中获得的更多的生物学相关的证据。值得注意的是,在LME中反映GC微环境比其他的微环境catrgories拥有更好的预后。这表明,类似于centroblasts中起作用的分子治疗的靶点。微环境可以提供避免淋巴瘤发生的原始机制。相反,淋巴瘤发展和/或选择有助于逃避免疫微环境限制的遗传和表观遗传特征。我们的数据表明了对于最初富含有免疫力的DLBCL,疾病的进展往往与免疫-deserted LME相关。更加广泛的说,这一概念表明淋巴瘤细胞和LME细胞(可能还涉及到非免疫的LME细胞)共同进化在这两个组分中朝着异质性较低的方向发展。随着更多的LME被发现,这一概念仍然有待验证。
除了免疫细胞对淋巴瘤细胞的发展有限制以外,我们的研究表明ECM也具有关键的作用。ECM是一种物理上和生物学上不同的大分子网络。主要为纤维蛋白,蛋白聚糖和糖蛋白组成,它们是组织结构完整性和恶性和微环境细胞行为调节的核心。我们描述具有较高比例的来源于CAFs(即CAF基质型)的ECM组分的LME具有良好的预后,而不是来自TAMs的ECM成分的LME具有良好的预后。一些ECM来源的蛋白,一些ECM衍生的蛋白质,特别是富含亮氨酸的小蛋白聚糖的BGN和/或DCN,发挥短期或长期的效应,最终影响淋巴瘤细胞的增殖,正如我们PDTX实验展示的那样。因为PDTX是在免疫功能低下的小鼠,因此该机制独立于适应性淋巴瘤免疫应答的建立。沿着这些思路,先前的工作已经确认了DLBCL中CAFs有可能与更好的结果相关。此外,通过分泌可溶性因子如TGFB配体的间质基质中,成纤维细胞可能有助于减少淋巴瘤细胞的进展。一种机制,可以通过TGFB换能器如SMAD1的表观遗传抑制在DLBCL中克服,CAFs和TGFB的抗肿瘤作用在DLBCL中与大多数实体瘤中的作用相反,强调了肿瘤特异性方式研究肿瘤微环境的必要性。
在队列研究中,HGBL-DH患者具有BCL2和MYC转位特性以及高级别转录signature的表达特性在一线治疗以后。这通常证明实施强化化疗方案是合理的。但是,在个体水平,确实存在对于标准免疫化疗方案的长期反应者,这表明存在侵袭性较低的淋巴瘤亚组。尽管这一现象可以被额外的基因和表观遗传突变的存在所解释,我们的数据仍然表明了和不同LME catrgories相关的生物学行为。HGBL-DH以及其他的MHG DLBCL在GC-like和MS-LMEs中相比于IN和DP-LMEs中具有更好的预后。鉴于这种与淋巴瘤亚型相关的化学耐药性,新方法正在考虑添加靶向治疗,例如BCL2抑制剂。我们的研究证明,在临床研究中,在临床的试验中,不仅应在特定遗传亚型的背景下考虑靶向药物,还应该考虑LME类别。
总之,我们通过不同的生物学组成和临床行为定义了四种不同的LME类别,补充了基因表达谱的亚型以及遗传的亚型,指导这些患者开发出合理的治疗方法。


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









