








摘要
玉米的高度由节点数量和节间长度决定。节点数量受插生分生组织的形成驱动,而节间长度则由插生细胞的伸长决定。然而,调控节点建立和节间生长的机制尚不清楚。我们筛选了EMS诱导的玉米突变体,并鉴定出一个与**终端耳基因1(ZmTE1)**的单一碱基变异相关的矮化突变体zm66。详细的表型分析表明,zm66(zmte1-2)具有较短的节间和增加的节点数,这分别是由细胞伸长减少和插生分生组织形成紊乱引起的。转录组分析显示,zmte1-2中的生长素信号基因发生了失调,细胞伸长和细胞周期相关基因也受到影响。这表明ZmTE1调控生长素信号、细胞分裂和细胞伸长。我们发现ZmWEE1激酶能够磷酸化ZmTE1,从而将其限制在细胞核内,并可能抑制细胞分裂。相反,ZmPP2Ac-2磷酸酶促进ZmTE1的去磷酸化及其在细胞质中的定位,同时促进细胞分裂。综合来看,ZmTE1是植物高度的重要调节因子,负责维持节间分生组织的有序形成和快速的细胞伸长。ZmWEE1和ZmPP2Ac-2可能通过平衡ZmTE1的活性,控制细胞分裂和伸长,从而维持玉米的正常生长。
引言
植物高度的变化会影响作物产量,从而影响粮食安全。矮性或半矮性对作物有一定的优势,因为它可以提高抗倒伏能力、增加植株密度和提高收获指数,这对生产具有积极作用(Zhang et al., 2019)。营养生长顶端分生组织(SAM)支持植物的生长和发育。SAM包含一池未分化的细胞,这些细胞可以生成地上结构的原基,包括叶片。在营养生长过程中,叶片由叶原基发育而来,并遵循一种保守的双列叶序。这意味着新叶会在与前一片叶片相对的位置从分生组织的另一侧开始形成(Jackson and Veit, 1994)。玉米的茎由一个植物单元组成,包括叶片、叶节点、节间和腋生分生组织单元(Zhang, Sun, et al., 2018)。茎的生长主要是由节点数量的增加和节间区域的伸长造成的。节点细胞和节间细胞来自同一个细胞池,即SAM(Tsuda et al., 2017)。
近年来,已分离出多种在节间伸长方面存在缺陷的突变体。水稻D50是一种假定的肌醇多磷酸5-磷酸酶(5PTase),研究表明它通过调控细胞分裂方向、细胞壁果胶的沉积以及微丝的组织,促进居间分生组织(即在成熟组织间发育的分生组织)形成(Sato-Izawa et al., 2012)。水稻的Os-GRF1(Oryza sativa-GROWTH REGULATING FACTOR1)参与了**生长素(GA)**诱导的茎部伸长(van der Knaap and Kim, 2000)。在玉米中,也分离出一些与生长素相关的突变体,包括矮秆1(d1)、d3、d5、雄穗耳1(an1)以及显性突变体D8和D9。这些突变体都影响玉米生命周期中的节间伸长。矮秆突变体gif1的窄叶和短节间与叶片和茎部未分化细胞的减少相关(Zhang, Sun, et al., 2018)。矮秆突变体brevis plant1(bv1)的短节间和发育缺陷的居间细胞与生长素运输不足有关(Avila et al., 2016)。类似地,矮秆突变体brachytic2(br2)的下节间缩短是由于从茎尖和根尖分生组织中减少了生长素外流(Knoller et al., 2010; Zhang et al., 2019)。BLH12/14是KNOTTED1(KN1)辅因子,通过与KN1相互作用维持居间分生组织,并防止节间过早分化(Tsuda et al., 2017)。
生长素作为一种重要的植物激素,调控着细胞分裂、细胞伸长和细胞分化,几乎在植物生长和发育的所有阶段发挥作用(Ma 和 Grones, 2018)。生长素的极性运输是通过生长素外流载体(如PINFORMEDs,PINs)和生长素内流载体(如AUXIN/LIKE AUXINs,AUX/LAXs)介导的,这一过程涉及植物的多种生长和发育(Carraro et al., 2006)。PIN1作为一种生长素外流载体,是分生组织(SAM)中生长素分布的主要调节因子(Carraro et al., 2006; de Reuille et al., 2006)。在初期叶原基处ZmPIN1的表达下降,导致叶片发生延迟、分生组织扩大以及异常叶序的变化,这些现象都与异常叶序突变体(abph1,也称为abphyl1)有关(Lee et al., 2009)。此外,生长素信号因子(ARFs)通过上调或下调特定的靶基因,促进细胞膨胀、细胞分裂和细胞壁重构(Chandler, 2016)。小型生长素上调RNA(SAUR)基因家族包括植物特有的细胞效应因子,能够影响生长素水平和生长素极性运输(Ren 和 Gray, 2015; Stortenbeker 和 Bemer, 2019)。大多数SAUR基因,包括AtSAUR10、AtSAUR19/63和AtSAUR32/50,已被充分证明通过增强H±ATPase活性和细胞伸长来促进拟南芥的胚轴生长(Bemer et al., 2017; Spartz et al., 2014; Stortenbeker 和 Bemer, 2019)。
尽管已有一些玉米基因被鉴定为调控插生分生组织形成和节间细胞扩展的因子(Ballesteros et al., 2013; Tsuda et al., 2017; Zhang, Sun, et al., 2018),但其背后的分子机制仍然不明确。因此,我们对EMS诱导的玉米突变体库进行了高通量筛选,识别出了一个侏儒突变体zm66,其表现为较短的节间和增加的节数。进一步的研究表明,zm66的侏儒表型是由ZmTE1基因的突变引起的,而ZmTE1已被证明是调节玉米高度的关键因子(Veit et al., 1998)。ZmTE1蛋白未被充分表征,但它与酿酒酵母的减数分裂诱导因子2(Mei2)高度相似,Mei2编码一种含有三个保守RNA识别结构域(RRMs)的RNA结合蛋白。Mei2已被报道通过促进前减数分裂的DNA合成来促进酵母细胞的减数分裂(Watanabe 和 Yamamoto, 1994)。相反,Pat1激酶通过磷酸化Mei2,抑制有丝分裂向减数分裂的过渡(Watanabe et al., 1997)。在本研究中,我们确认了ZmTE1在维持插生分生组织形成、节间细胞伸长和植物高度调控中的决定性作用。ZmPP2Ac-2和ZmWEE1可能通过调节ZmTE1的磷酸化和亚细胞定位,从而影响细胞分裂、细胞伸长和植物高度。
结果
ZmTE1对玉米植物高度至关重要
玉米的高度由节点数和节间长度决定,这与插生分生组织的形成和节间细胞的伸长密切相关(Tsuda et al., 2017; Zhang et al., 2019; Zhang, Sun, et al., 2018)。为了阐明插生分生组织形成和节间细胞伸长的分子机制,我们筛选了玉米EMS诱导的突变体库,并识别了一个侏儒突变体zm66(图1a, b)。除了侏儒表型外,该突变体还表现为花序雌性化,进一步导致了类似穗的外观和种子在正常终端花序的位置形成(图1c,d),并且每株植物的种子大小和重量明显减少(图1e,f)。该突变体被回交到自交系B73(以下简称WT),以去除与表型无关的变化。zm66 × B73的F2代在植物高度上表现出3:1(高/矮)的表型分离比(图1g),表明zm66的侏儒表型由一个单隐性基因决定。
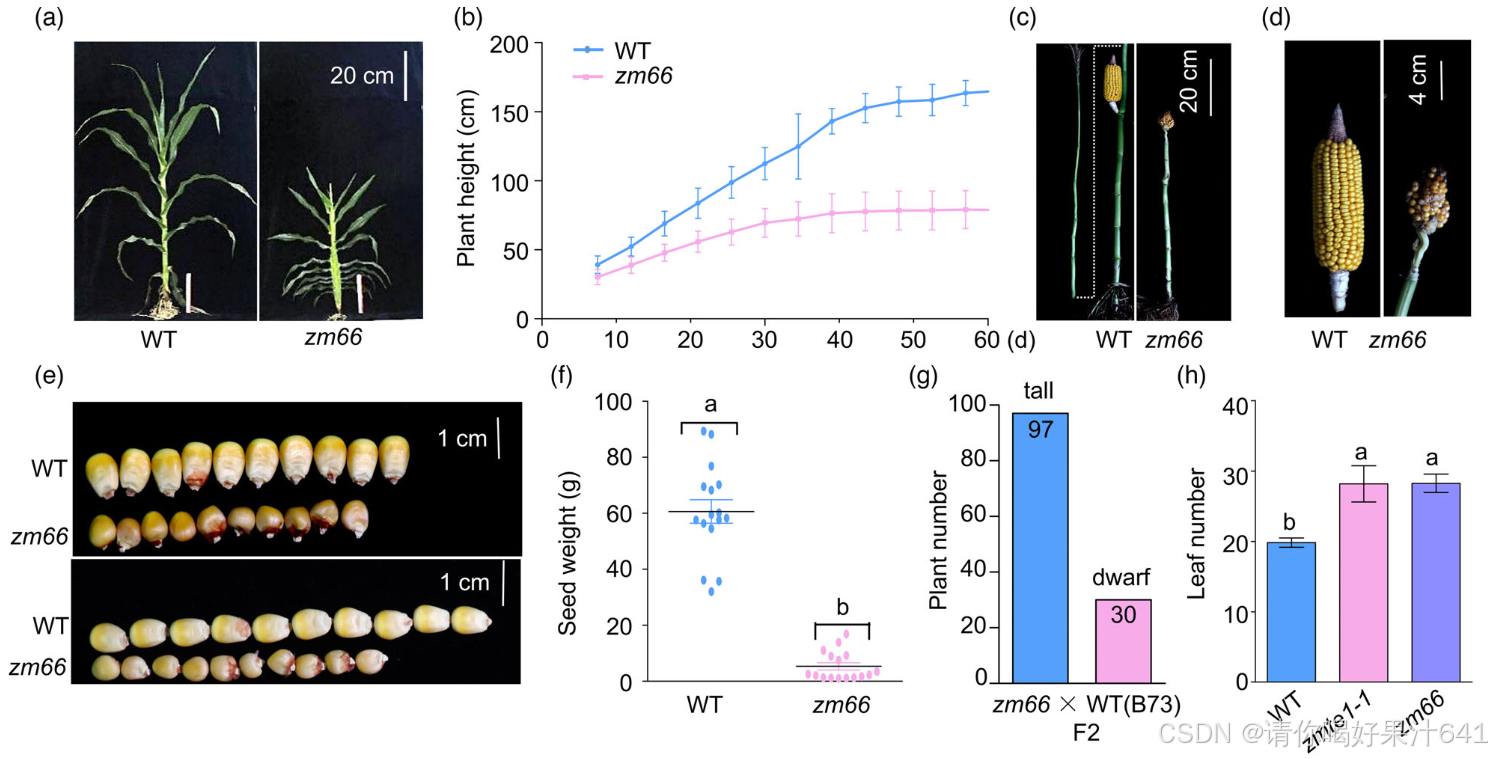
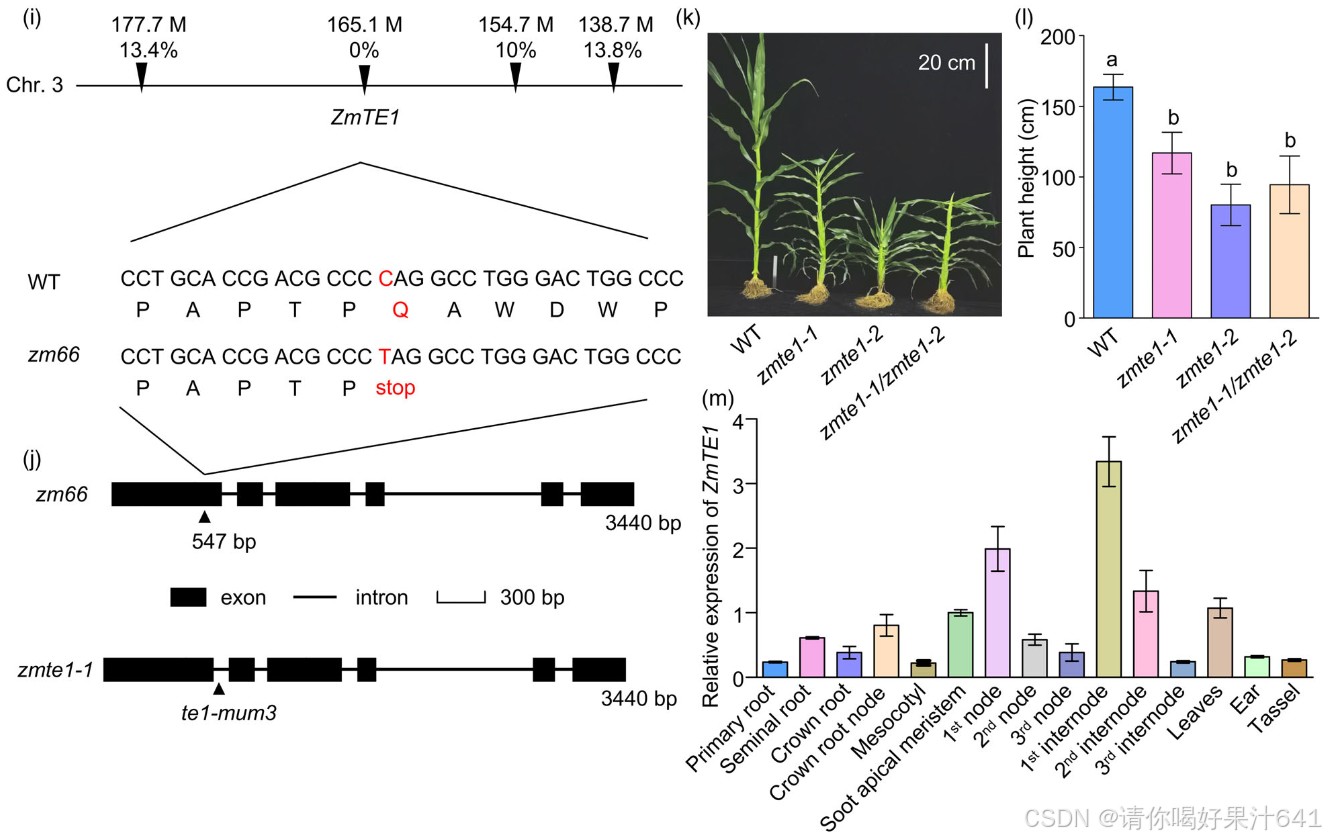
为了确定zm66的突变位点,我们通过基于外显子捕获的MutMap(EcMutMap)分析(Lu et al., 2018),从40株表现出突变表型的zm66 × B73 F2代幼苗中获得了28,594个单核苷酸多态性(SNPs;Excel S1)(图1i)。筛选后,识别出10,687个过渡突变(即G-A或C-T),其中包括315个具有高质量评分的SNP(次要等位基因频率≥90%)。其中,四个SNP(SNP177676403、SNP-165174753、SNP-154716744和SNP138739421)对应于染色体3上编码区的非同义突变,与突变体相关,重组频率分别为13.4%、0.0%、10.0%和13.8%(图1i)。SNP-165174753被定义为ZmTE1(GRMZM2G085113)基因,其重组频率为0%,并且负责侏儒表型。ZmTE1基因第一外显子第547碱基位点的C-T突变将一个谷氨酰胺替换为一个提前的终止密码子(图1i,j)。ZmTE1之前已被报道在防止叶片过早起始和发育中起着重要作用(Veit et al., 1998),这一表型与zm66突变体中叶片数量的增加一致(图1h)。因此,我们将zm66重新命名为zmte1-2,继之前识别的zmte1-1之后(Veit et al., 1998)。我们将zmte1-1等位基因与zmte1-2进行交配(图1k)。zmte1-1/zmte1-2的F1植物也表现出类似于zmte1-1和zmte1-2的侏儒表型(图1k, l),表明zmte1-2的植物高度表型是由ZmTE1功能丧失引起的。
ZmTE1是一种RNA结合蛋白,包含三个RNA识别基序(RRM1、RRM2和RRM3),并与酿酒酵母(Schizosaccharomyces pombe)Mei2、稻米(Oryza sativa)LHD2/PLA2/OML1和拟南芥(Arabidopsis thaliana)AtTEL1/AtTEL2表现出高度相似性(Jeffares et al., 2004; Kawakatsu et al., 2006)。为了更好地理解Mei2样蛋白的进化关系,我们基于酿酒酵母、玉米(Zea mays)、稻米和拟南芥中的Mei2样蛋白构建了一个系统发育树。我们在玉米中发现了九个Mei2样蛋白,在稻米中发现了七个,在拟南芥中发现了八个(图S1)。尽管玉米中有九个Mei2样蛋白,但ZmTE1的缺失会导致侏儒表型(图1a,b),这表明ZmTE1在植物生长调控中的关键作用。
ZmTE1在几乎所有玉米组织中都有表达(图1m),这表明其在不同组织中的功能重要性。特别地,ZmTE1在节间和叶片中的表达相对较高(图1m),进一步表明zmte1-2植物中叶片和茎部的发育异常是由于ZmTE1功能丧失所致。
ZmTE1通过加速细胞伸长促进节间伸长。
成熟的zmte1-1和zmte1-2植物的株高分别减少了33%和50%(图1l)。我们发现,zmte1突变体的节间变短,从而导致株高降低(图2a,b)。zmte1-1和zmte1-2的所有节间都比WT短,尤其是第10、第11、第12和第13节间受影响最为明显(图2b;图S2)。此外,zmte1突变体与WT相比,其他重要的农艺性状,如叶片数、叶片大小和节间长度,也发生了变化(图1h,2a-c)。穗叶的最大宽度和长度尤其受到影响,分别减少了47%、36%和32%、46%(图2c-e)。
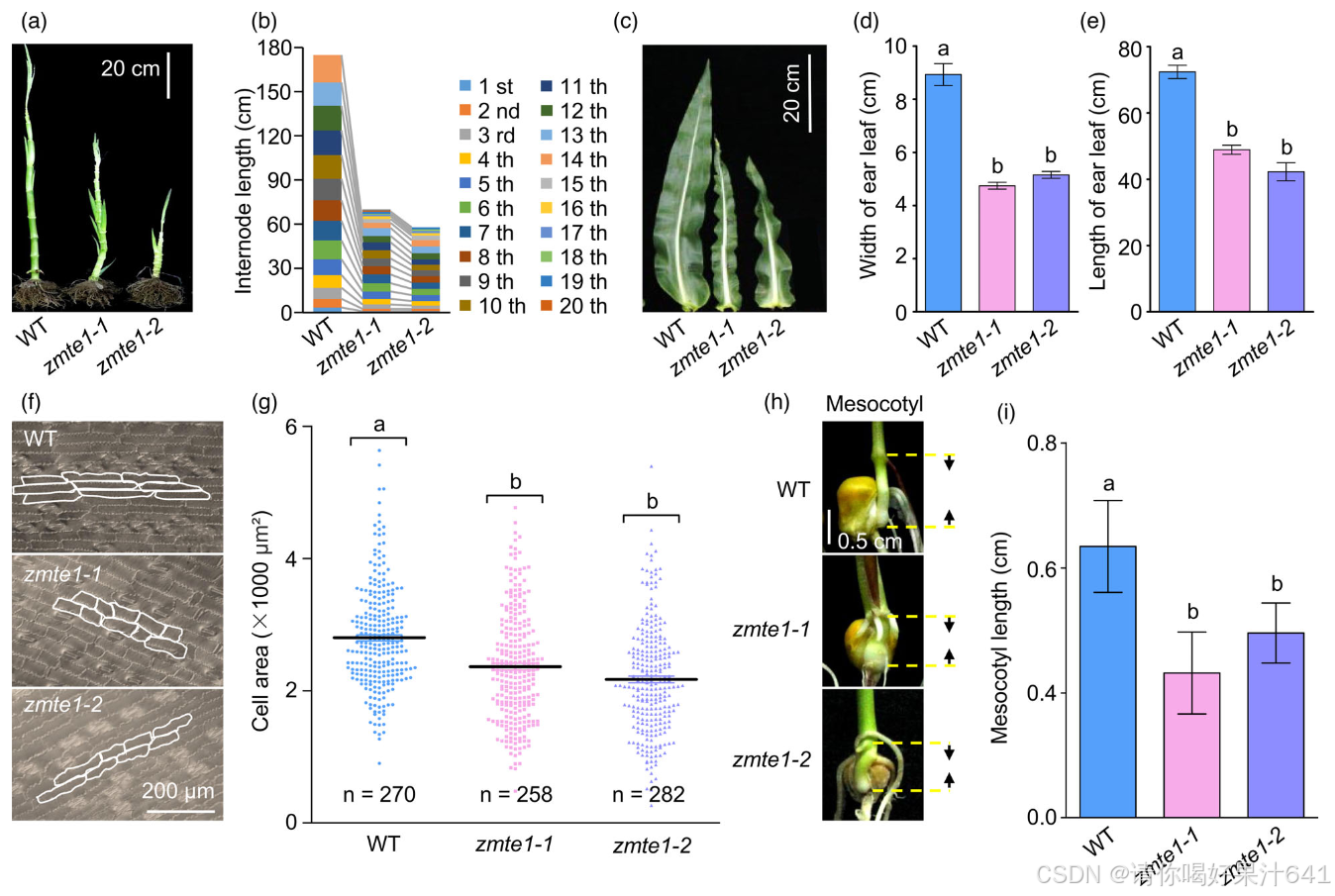
先前的研究表明,节间长度主要依赖于节间细胞的伸长(Tsuda et al., 2017),这暗示ZmTE1可能正向调节节间细胞的伸长。不幸的是,由于节点的纤维化程度较高,我们未能获得良好的茎组织切片,因此无法直接检查插生细胞的长度。由于zmte1-1和zmte1-2突变体的叶片也明显小于WT(图2c-e),我们检查了下表皮细胞的形态(图2f),发现这些细胞在zmte1-1和zmte1-2突变体中比WT更小(图2f,g)。胚芽鞘位于根和茎之间,主要负责将芽推出土壤(Saab和Ho,1995),其生长与拟南芥胚轴相似,即与细胞伸长相关(Kutschera和Wang,2016)。在zmte1-1和zmte1-2植物中,胚芽鞘的发育也明显滞后于WT(图2h,i),进一步确认ZmTE1促进细胞伸长。考虑到zmte1-1和zmte1-2的叶片收缩和胚芽鞘缩短都是由细胞伸长受限引起的,而节间伸长与细胞伸长相关(Tsuda et al., 2017),我们推测ZmTE1通过促进节间细胞伸长来增加节间长度。
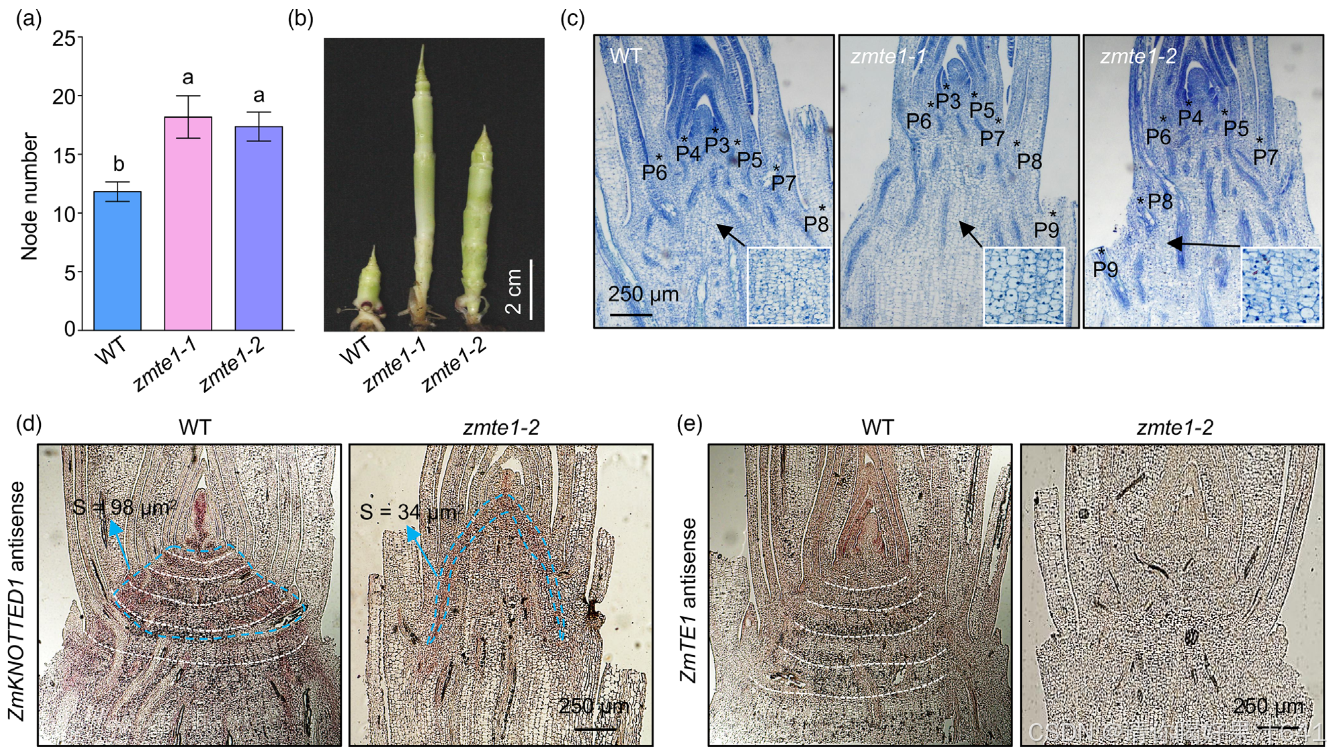
KN1,特异性表达于SAM和插生分生组织中(Tsuda et al., 2017),通常用于确定分生组织的定位和范围。然而,RNA原位杂交结果显示,zmte1-2的分生组织区域明显小于WT(图3d)。我们的数据表明,ZmTE1促进分生组织中的细胞分裂,因为分生组织的大小主要取决于细胞数量(Zhang, Sun, 等,2018)。此外,WT茎部中的规则层次结构逐渐发展为插生分生组织(Tsuda et al., 2017)。相比之下,zmte1-2茎部具有无序的结构(图3d, e),这表明ZmTE1在插生分生组织的有序形成中也发挥着关键作用。因此,zmte1-2植物中ZmTE1调控的缺失导致插生分生组织的任意形成,并随后导致节点的过度形成(图3a)。综上所述,ZmTE1在促进分生组织中的细胞分裂和维持插生分生组织的形成方面起着双重作用,从而确保植物的完整生长潜力。
ZmTE1 正向调控生长素信号通路
为了深入了解ZmTE1介导的细胞伸长和细胞分裂机制,我们对28天龄的WT和zmte1-2植物的节点和节间进行RNA测序(RNA-seq)。与WT相比,zmte1-2的节点中发现了5,546个差异表达基因(DEGs;P值≤0.05,倍数变化≥2)(Excel S2)。京都基因与基因组百科全书(KEGG)分析揭示,DEG数据集中有200个与生长素信号相关的基因(图S3)。考虑到生长素在调控细胞伸长和细胞分裂中的重要作用(Du和Spalding,2020),zmte1-2中的下调生长素簇基因(图4a)可能对这些突变体的发育表型产生影响。
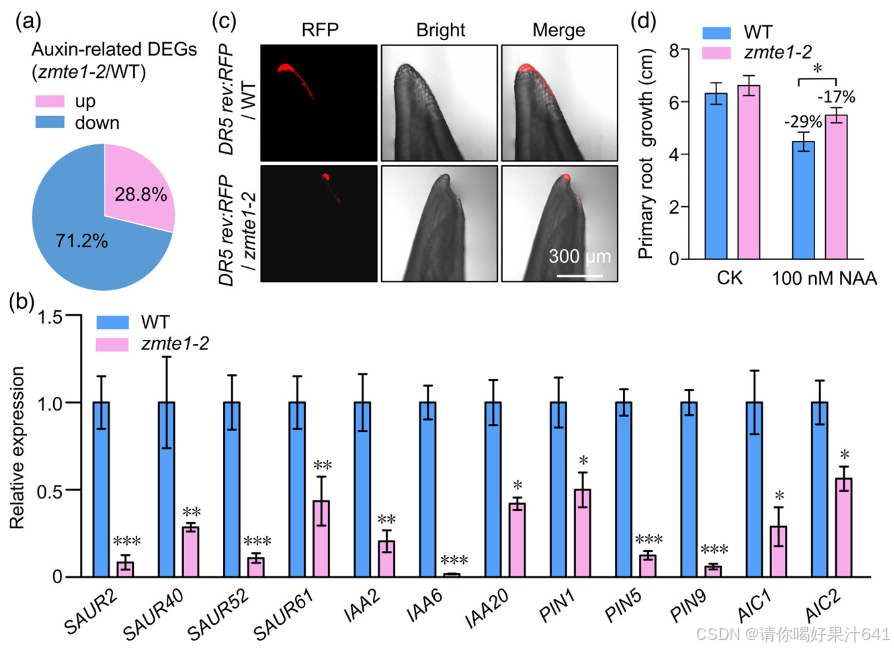
Aux/IAAs 和 SAURs 是生长素迅速诱导的基因(Abel 和 Theologis,1996),通常作为生长素信号反应的标记基因。在随后的详细qRT验证实验中,所有这些基因在zmte1-2中均显著下调(图4b),进一步确认了zmte1-2中生长素信号反应的下降。更直接地,我们将玉米DR5 rev:RFP(生长素响应标记)引入zmte1-2,以在SAM处直观观察生长素信号的强度。结果显示,zmte1-2中的RFP荧光强度显著降低(图4c),提示ZmTE1在维持SAM处生长素信号强度方面发挥着重要作用。
为了进一步确认ZmTE1对生长素信号的正向调控,我们验证了高浓度生长素抑制根伸长的作用在zmte1-2中是否被削弱。与我们的假设一致,当ZmTE1功能丧失时,生长素介导的根伸长抑制作用显著减弱(图4d),进一步加强了ZmTE1在生长素信号中的重要作用。还值得一提的是,一些生长素转运蛋白基因在zmte1-2突变体中也显著减少(图4b),这暗示ZmTE1可能通过调节生长素转运来增强生长素信号。具体而言,已报道PIN1在SAM发育过程中调控生长素分布发挥着重要作用(Carraro et al., 2006)。在zmte1-2中下调PIN1的表达,提示SAM的发育异常可能是由于生长素分布的干扰和生长素信号不足(图4b)。WT与zmte1-2之间在生长素合成基因的表达上没有显著差异(Excel S2),表明ZmTE1介导的生长素信号调控可能与生长素合成无关。
总之,ZmTE1正向调控生长素信号。
ZmTE1 调控细胞分裂和细胞伸长相关基因的表达
为了进一步研究 zmte1-2 中细胞分裂和细胞伸长缺陷的分子机制,我们再次仔细分析了 RNA-seq 数据。多个细胞周期相关基因,包括 CYCAs、CYCBs 和 CDKs,在 zmte1-2 中显著下调(图 S4a)。这一结果表明,正常的分生组织细胞分裂需要 ZmTE1 的维持,这与在 zmte1-2 中观察到的分生组织发育缺陷一致(图 3d)。与 zmte1-2 中细胞伸长缺陷一致,细胞伸长相关的基因,包括 SAURs、EXPANSIONs 和 EXTs,也在 zmte1-2 中显著下调(图 4b;图 S4b)。此外,一些负向调控细胞伸长的基因,如过氧化物酶(PERs)(Knoller et al., 2010),在 zmte1-2 中显著上调(图 S4b)。这些结果表明,ZmTE1 是细胞伸长的必要维持因子,这与在 zmte1-2 中观察到的节间和胚轴缩短现象一致(图 2a,h)。已有充分的文献表明,生长素通过上调 SAURs、EXPANSIONs 和 EXTs 的表达,并下调 PERs 的表达,促进细胞伸长(Du et al., 2020;Knoller et al., 2010),这支持了生长素信号对 ZmTE1 诱导的细胞伸长至关重要的观点。
总之,ZmTE1 通过生长素介导的细胞分裂和细胞伸长调控,维持分生组织的形成和节间的伸长。
ZmTE1 与 ZmPP2Ac-2 和 ZmWEE1 互作
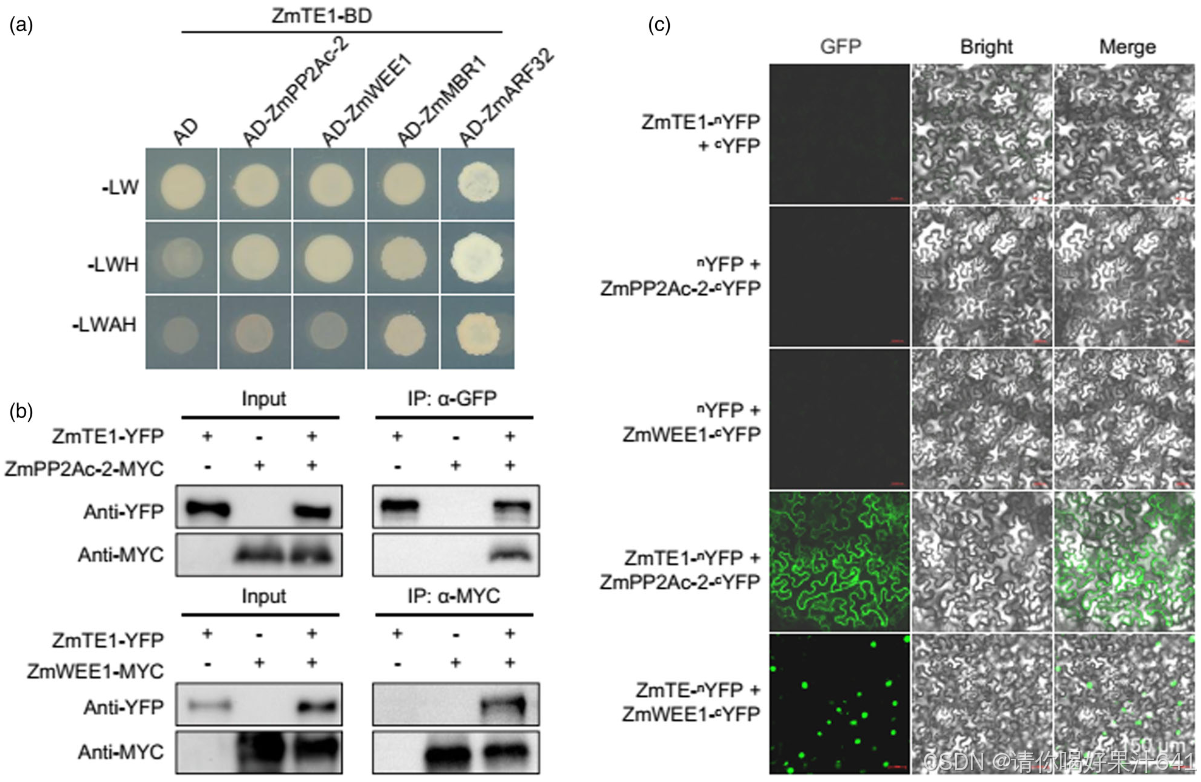
为了进一步阐明 ZmTE1 如何调控细胞伸长、细胞分裂和插生分生组织的形成,我们使用** ZmTE1 作为诱饵蛋白筛选了玉米 cDNA 文库**。我们鉴定出四个与 ZmTE1 互作的蛋白,分别是 ZmMBR1、ZmARF32、ZmWEE1 和 ZmPP2Ac-2(图 5a)。AtMBR1(GRMZM2G165044,在本研究中命名为 ZmMBR1)是 E3 泛素连接酶 AtMED25-BINDING RING-H2 PROTEIN1 的同源物,已被报道参与促进开花(Inigo et al., 2012)。ZmARF32 是转录因子 AtARF17 的同源物,作为 ZmTE1 的结合伙伴。AtARF17 已被报道在花药裂开和花粉壁图案形成中起关键作用(Xu et al., 2019)。这两种蛋白可能与节间细胞伸长或分生组织中的细胞分裂无关。ZmWEE1 是 WEE1 激酶的同源物,WEE1 激酶作为细胞周期 G2/M 检查点,在 DNA 复制应激时抑制细胞周期的进行(Velappan and Signorelli, 2017),并且是 ZmTE1 的潜在结合伙伴之一。此外,我们还发现 ZmPP2Ac-2 基于酵母双杂交分析与 ZmTE1 互作。ZmPP2Ac-2 是与植物磷酸酶 AtPP2AC3 和 AtPP2A-C4 同源的蛋白,通过去磷酸化促进 PIN1 的极性定位,从而促进生长素的极性运输。这些磷酸酶已被报道调控细胞伸长和分生组织的形成(Ballesteros et al., 2013)。我们还通过双分子荧光互补(BiFC)和共免疫沉淀(CoIP)实验在烟草叶片表皮细胞中验证了 ZmTE1 与 ZmWEE1 和 ZmPP2Ac-2 的互作(图 5b, c)。
ZmWEE1 和 ZmPP2Ac-2 调控 ZmTE1 的磷酸化状态和亚细胞定位
由于 ZmWEE1 激酶和 ZmPP2Ac-2 磷酸酶都与 ZmTE1 互作,我们推测 ZmWEE1 和 ZmPP2Ac-2 可能通过分别调节磷酸化和去磷酸化来影响 ZmTE1 的功能。为了验证这一假设,我们在拟南芥原生质体中进行了体内磷酸化实验。共转化 ZmWEE1 和 ZmTE1 后,出现了一个额外的蛋白条带,经过小肠碱性磷酸酶(CIAP)处理后,该条带被去除(图 6a),表明这个蛋白是磷酸化的 ZmTE1。值得注意的是,ZmWEE1 介导的 ZmTE1 磷酸化也被 ZmPP2Ac-2 消除(图 6a),这表明 ZmTE1 可以分别通过 ZmWEE1 和 ZmPP2Ac-2 进行磷酸化和去磷酸化。这些数据证实了这些因子对 ZmTE1 磷酸化具有相反的调节作用,并可能通过调控这一翻译后修饰的相反作用来调节 ZmTE1 的功能。
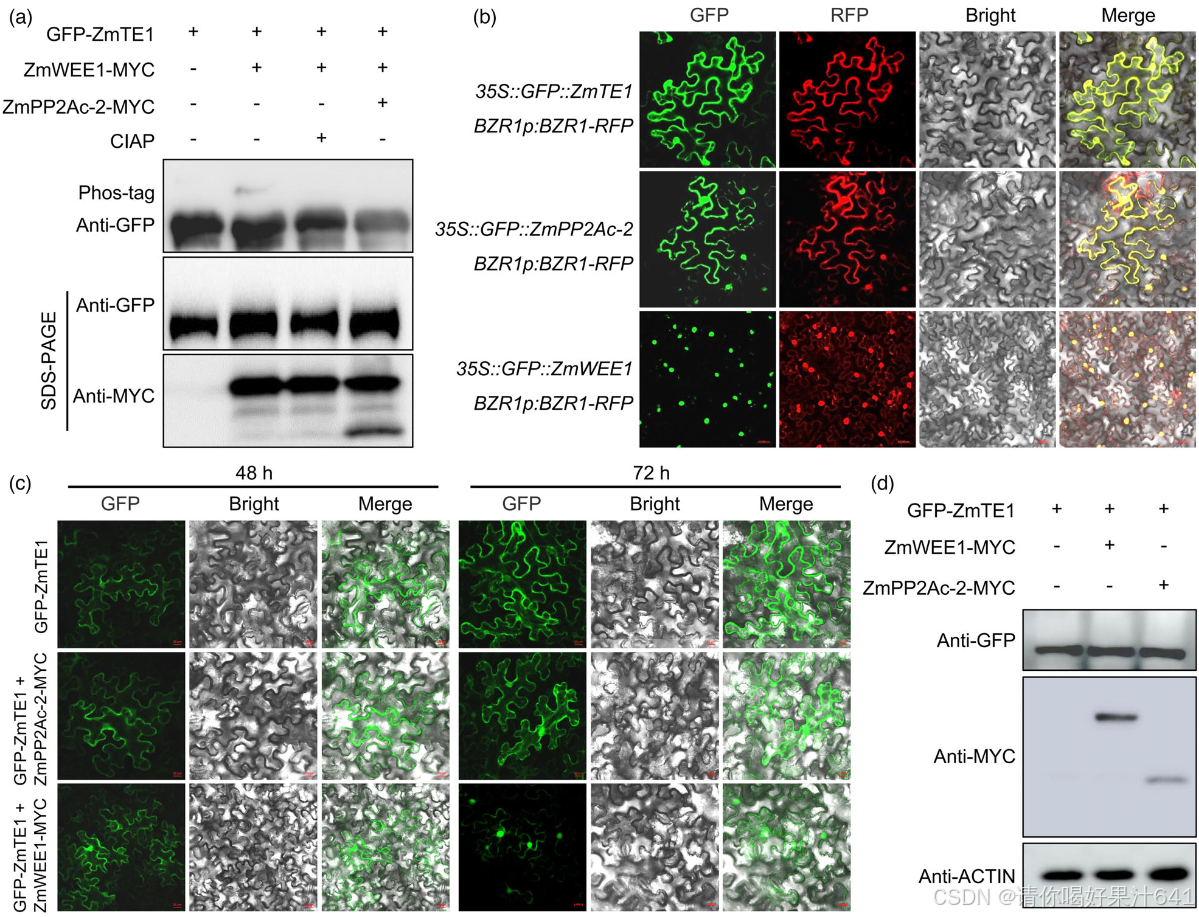
磷酸化修饰可以影响蛋白质的稳定性、活性、亚细胞定位或与其他蛋白质的相互作用(Yu et al., 2019; Zhang et al., 2021)。鉴于 ZmTE1-ZmPP2Ac-2 的荧光在细胞质和细胞核中都有观察到,而 ZmTE1-ZmWEE1 的荧光则专门出现在细胞核中(图 5c),我们推测 ZmTE1 的磷酸化可能改变其亚细胞定位。在随后的亚细胞定位分析中,我们发现 ZmTE1 和 ZmPP2Ac-2 都定位于细胞质和细胞核,而 ZmWEE1 专门定位于细胞核(图 6b)。然而,在共转化实验中,ZmWEE1 几乎完全将 ZmTE1 限制在细胞核内(图 6c),这意味着磷酸化的 ZmTE1 更倾向于定位于细胞核。与此假设一致,在 ZmPP2Ac-2 存在的情况下,去磷酸化的 ZmTE1 倾向于定位于细胞质(图 6c)。此外,ZmWEE1-ZmTE1 或 ZmPP2Ac-2-ZmTE1 的共转化并未影响 ZmTE1 的蛋白质水平(图 6d),表明 ZmTE1 的磷酸化主要影响其亚细胞定位,而不是蛋白质的稳定性。考虑到 ZmPP2Ac-2 和 ZmTE1 在调控生长素运输、细胞伸长和分生组织形成中的一致作用(Awotunde et al., 2000; Ballesteros et al., 2013; Yue et al., 2016),以及 ZmWEE1 和 ZmTE1 在调节细胞周期中的相反作用(Velappan et al., 2017),ZmPP2Ac-2 介导的 ZmTE1 去磷酸化和核输出可能促进生长素信号传导、细胞分裂、细胞伸长和分生组织形成,而 ZmWEE1 则可能抑制 ZmTE1 的功能。
讨论
植物高度对倒伏有重要影响,因此是决定作物产量的关键因素。这一点在玉米和水稻中特别明显,因为植物高度主要取决于节间数和节间长度(Tsuda et al., 2017;Zhang, Sun, et al., 2018)。为了阐明调控插生分生组织形成和插生细胞伸长的分子机制,我们筛选了一个 EMS 诱变的玉米突变体库,并鉴定了一个矮秆突变体 zm66,该突变体表现为节间较短和节点数增多。zm66 的矮秆表型是由 ZmTE1 基因突变引起的,ZmTE1 已知在玉米高度调控中起着关键作用(Veit et al., 1998)。我们的研究进一步表明,ZmTE1 通过调控生长素信号、细胞分裂、细胞伸长和插生分生组织的形成,来调节玉米的高度。
由于 ZmPP2Ac-2 和 ZmTE1 都调控生长素信号、细胞伸长和分生组织形成(Awotunde et al., 2000;Ballesteros et al., 2013;Yue et al., 2016),我们进一步研究了这一磷酸酶以及与之对立的激酶 ZmWEE1(Velappan et al., 2017)的作用。我们的数据表明,ZmPP2Ac-2 和 ZmWEE1 可能通过直接的蛋白质相互作用对 ZmTE1 的功能产生正向和负向的调控效应。此外,ZmWEE1 将 ZmTE1 特异性限制在细胞核内,而 ZmPP2Ac-2 和 ZmTE1 则促进其在细胞质中的定位(图 6b,c)。因此,ZmTE1 的细胞核与细胞质定位可能是调控其功能的关键。通过调控这一亚细胞定位,ZmPP2Ac-2 和 ZmWEE1 可能影响 ZmTE1 促进细胞分裂、细胞伸长和分生组织形成的能力。细胞分裂和细胞伸长对于植物生长至关重要,这意味着 ZmTE1 在正常情况下可能会从细胞质转运到细胞核。然而,由于 ZmWEE1 与 DNA 损伤时关闭细胞周期以允许 DNA 修复相关(Velappan et al., 2017),因此 ZmWEE1 媒介的对 ZmTE1 的抑制作用在 DNA 损伤修复过程中尤为重要。随着 DNA 损伤修复并恢复植物生长,ZmTE1 的磷酸化被 ZmPP2Ac-2 去除,亚细胞定位的限制也会得到解除(图 6a)。
Mmi1 是一种 RNA 结合蛋白, 在有丝分裂细胞周期中至关重要,能够去除有害的减数分裂特异性 mRNA(Yamanaka et al., 2010)。有研究报道,ZmTE1 的酵母同源基因 Mei2 可以通过抑制 Mmi1 的功能,在进入细胞核后暂停有丝分裂并启动减数分裂(Mukherjee 和 Futcher,2018)。这表明,细胞核中的 Mei2 会减少细胞分裂。如果细胞核中的 ZmTE1 也抑制细胞分裂,那么在 zmte1-2 中相关基因的表达应该显著增加,但实际情况并非如此(图 S4a)。因此,我们假设,含有 RNA 识别结构域的 ZmTE1 转移到细胞质中,并不会去除与细胞分裂相关基因的抑制,而是将这些基因的 mRNA 转运到细胞质中,加速其翻译。遗憾的是,由于 mRNA 检测方法的敏感性较低,我们未能从在拟南芥原生质体中表达的 ZmTE1-GFP 蛋白中分离出 ZmTE1 结合的 mRNA。
酵母双杂交实验表明,ZmTE1 与七个 ZmARF 成员相互作用(图 5A 和 S5),这些成员是与生长素信号传导密切相关的核心转录因子(Chandler, 2016)。考虑到 ZmTE1 功能丧失导致生长素信号传导显著下降(图 4a-c),ZmTE1 可能在正向调控生长素信号传导。因此,ZmTE1 与 ZmARF 之间的相互作用可能通过增强生长素响应基因的表达,从而调控相关生理过程,这一具体机制有待在未来的研究中进一步探索。
总之,提出了一个工作模型,以说明 ZmTE1 如何通过调控插生分生组织的形成和节间细胞的伸长来促进玉米植物的高度(图 7)。ZmTE1 可能间接控制与细胞周期、细胞伸长和生长素信号传导相关基因的转录,从而增加这些 mRNA 在细胞核中的浓度。ZmPP2Ac-2 可能通过去磷酸化维持 ZmTE1 从细胞核向细胞质的转移,同时将这些结合的 mRNA 带入细胞质,增强这些 mRNA 转化为功能性蛋白质的翻译。相反,ZmWEE1 对 ZmTE1 进行磷酸化,限制这些结合的 mRNA 留在细胞核。然而,ZmTE1 的确切直接靶标 mRNA 以及 ZmWEE1 和 ZmPP2Ac-2 对 ZmTE1 活性的精细调控平衡仍需在未来进一步研究。
材料与方法
矮秆突变体筛选与 ZmTE1 基因鉴定
约 12,000 个通过 EMS(化学诱变剂)诱导的玉米突变体,来源于 Lu 等人(2018)的研究,经过田间表型筛选,包括植物高度。我们筛选到了一株矮秆突变体 zm66(zmte1-2),并将其与野生型 B73 进行回交。F1 代植物自交,F2 代植物在田间生长,并测量 80 天龄植物的植物高度。我们还将 zmte1-2 和来自突变体库的 zmte1-1 进行交配(https://www.maizegdb.org/),通过 F1 植株的表型来检查是否存在等位基因关系。
基于外显子捕获的 MutMap(EcMutMap)(Lu 等,2018)是一种改进的 MutMap 方法,通过突变体与野生型交配,快速鉴定基因,这一方法已在水稻中得到应用(Abe 等,2012;Takagi 等,2013)。简言之,提取 40 个显示矮秆表型的 F2 种子ling 的 DNA,采用 CTAB 方法提取,并将其按等摩尔比例混合,然后使用 GAIIx 设备(Illumina)进行 EcMutMap 测序。通过 MAQ 软件(Li 和 Ruan,2008)将过滤后的读数映射到野生型参考序列上。通过对每个 40 个模板的扩增子进行测序来验证多态性,并计算交叉率(Zhang, Lu 等,2018)。当变异位点与 G/A 或 C/T 峰对相关时,推断在一条同源染色体上发生了交叉;单个峰对应 G 或 C 时,表示两个同源染色体上都发生了交叉,而单个 A 或 T 峰则表明没有发生交叉。计算每个突变体的交叉率(G/G+A 或 C/C+T)。不与矮秆表型相关或位于不同染色体上的变异位点预计会以 1:1 的比例分离;与致病位点完全连锁的变异则不应与表型分离。位于同一染色体上的变异(本研究为第 3 染色体)与致病位点连锁的变异预计会部分连锁。通过对每个突变体进行重测序,获得 ZmTE1 的等位基因状态,使用的引物见表 S1。
外源NAA处理对主根生长的响应
野生型(WT)和zmte1-2种子通过5%(W/V)NaClO消毒30分钟,并用无菌水洗涤三次。随后,种子在28°C/25°C(昼/夜)温控箱中,湿度约60%,光照周期为16小时光照/8小时黑暗(光照强度约100 μmol m²·s⁻¹)条件下水培生长。培养液为0.5×Hoagland液体溶液[0.51 g/L KNO3,0.82 g/L Ca(NO3)2,0.49 g/L MgSO4⋅7H2O,0.136 g/L KH2PO4,0.6 mg/L FeSO4,2.86 mg/L H3BO3,1.81 mg/L MnCl2⋅4H2O,0.08 mg/L CuSO4⋅5H2O,0.22 mg/L ZnSO4⋅7H2O,0.09 mg/L H2MoO4⋅4H2O],每两天更换一次培养液。三天龄的幼苗主根长度约为5 cm时,使用100 nm的NAA进行48小时处理。
系统发育分析
为了构建系统发育树,使用ClustalX(Yu et al., 2020)对裂殖酵母(Schizosaccharomyces pombe)、玉米(Zea mays)、水稻(Oryza sativa)和阿拉伯芥(Arabidopsis)中的Mei2样蛋白进行比对。基于该比对结果,使用MEGA7中的邻接法(neighbour-joining method)构建了系统发育树。
表型分析与细胞学观察
对zmte1突变体的表型进行了详细分析,记录了植物的高度、节间数和节间长度、叶片的大小和数量、芽鞘的长度以及种子的大小和重量。为了测量细胞大小,从zmte1-1、zmte1-2和野生型(WT)各选取三株植物在耳位的成熟叶片进行取样。使用奥林巴斯BX53显微镜观察叶片中央区域的下表皮细胞。每片叶子观察三个视野,每个视野测量约30个细胞。通过测量五片叶子的细胞平均长度来代表每个基因型的细胞大小,并使用Image J软件计算细胞大小。
RNA-Seq、qRT-PCR与组织表达模式分析
收集四周龄的野生型(WT)和zmte1-2突变体最后一根支根的三个节段,使用TRIzol试剂(Invitrogen,Carlsbad,CA,USA)进行RNA提取。纯化的RNA被送至BGI(深圳,中国)进行RNA-seq分析。通过P值≤0.05和|log2|≥1的阈值筛选差异表达基因。为了在分生组织(SAM)中可视化生长素信号反应,将携带DR5 rev:RFP转基因标记的玉米品系(Gallavotti et al., 2008)与zmte1-2突变体进行杂交,并扫描两周龄的DR5 rev:RFP/zmte1-2和DR5 rev:RFP/WT幼苗的RFP信号强度。
为了分析ZmTE1在不同玉米组织中的表达,从野生型(WT)植物中收集了两周龄的主根、种子根、冠根、冠根间隔、芽鞘、分生组织(SAM)、以及六周龄的支根、三个节段和间隔、叶片、雄穗和穗。使用TRIzol试剂提取不同组织的RNA,并通过qRT-PCR分析ZmTE1在不同组织中的表达。
对于qRT-PCR,从WT和zmte1-2的节段中提取RNA,并使用Transcriptor First Strand cDNA Synthesis试剂盒(Roche,Basel,Switzerland)根据厂家说明进行反转录。qRT-PCR在MyiQTM实时PCR检测系统(Bio-Rad,Hercules,CA,USA)上进行,使用ChamQ SYBR Color qPCR Master Mix(Q411,Vazyme,Nanjing,China)。ZmACTIN和ZmGAPDH基因作为参考对照。qRT-PCR使用的引物列在表S1中。
石蜡切片和原位杂交
将14天龄的zmte1和WT植物的分生组织(SAM)组织固定在含有4%多聚甲醛的溶液中,在4°C下固定16小时,然后用1×PBS洗涤两次,经过一系列乙醇梯度脱水,随后用二甲苯替换,嵌入Paraplast Plus(Sigma-Aldrich)中,切成8μm厚的切片。植物组织切片用托鲁胺蓝染色进行组织学观察,使用奥林巴斯BX53显微镜拍摄切片图像。
原位杂交按之前的方法进行(Zhou et al., 2011)。简而言之,我们构建了ZmTE1和ZmKN1的正义链和反义链RNA探针。使用ZmTE1和ZmKN1的引物对分别扩增461 bp和570 bp的片段,然后将其克隆入pSPT18载体,并分别使用HindIII和EcoRI线性化。正义链和反义链探针分别使用SP6和T7 RNA聚合酶合成,并以地高辛(Digoxigenin-11-UTP, Roche Diagnostics)作为标记。最后,按照之前的研究方法(Zhou et al., 2011)制备标准石蜡切片,并进行原位杂交。
酵母双杂交筛选cDNA文库与蛋白互作验证
从14天龄的WT幼苗中提取总RNA,经过DNase I处理去除基因组DNA。使用SMART cDNA文库构建试剂盒(Clontech)合成cDNA,并送往Takara(大阪,日本)构建cDNA文库。酵母双杂交实验按照厂家手册和Matchmaker GAL4 Two-Hybrid System 3(Takara,大阪,日本)进行。将ZmTE1的编码序列(CDS)克隆到诱饵质粒pGBKT7(BD)中,并按照厂家说明进行酵母双杂交筛选。
为了验证识别到的蛋白–蛋白相互作用,将候选基因的CDS克隆到pGADT7(AD)中。将BD-ZmTE1和AD-候选融合蛋白(空载体作为阴性对照)一起转入酵母Y2HGold中,使用PEG/LiAc方法进行转化。转化后的酵母在缺乏Trp和Leu的合成培养基板(SD培养基,-LW)上培养两天后,再转移到含有SD-Trp-Leu-His(LWH)和SD-Trp-Leu-His-Asp(-LWAH)培养基上继续培养三到四天。此实验使用的引物列在表S1中。
共免疫沉淀(Co-IP)
将ZmWEE1、ZmPP2Ac-2和ZmTE1基因克隆到pCAMBIA1390-7Myc-6HIS或pEarleyGate 101-YFP载体中,分别构建35S::ZmTE1-Myc、35S::ZmTE1-YFP、35S::ZmPP2Ac2-Myc和35S::ZmWEE1-YFP质粒。然后,将这些构建体转化到拟南芥叶肉细胞中进行瞬时蛋白表达。共免疫沉淀(Co-IP)实验按照之前的研究方法(Lv et al., 2020)进行。简要来说,收获拟南芥叶肉细胞并在细胞裂解缓冲液(0.5 mM EDTA;10 mM Tris-HCl;pH 7.5;0.5% NP-40;1 mM PMSF;150 mM NaCl)中冰上裂解30分钟,每10分钟吹打一次。裂解液离心后,收集上清液并与MYC-Trap磁性琼脂糖珠(Chromotek,目录号ytma20,德国)在4°C下孵育2小时。然后,用稀释缓冲液(10 mM Tris-HCl;pH 7.5;150 mM NaCl;0.5 mM EDTA)洗涤珠子三次,最后重悬于SDS加样缓冲液中。重悬的珠子煮沸10分钟后,通过Western blot检测,使用抗MYC抗体(Abclonal,目录号AE010,武汉,中国)或抗GFP抗体(TransGen Biotech,目录号HT801-02,北京,中国)进行检测。
双分子荧光互补(BiFC)和亚细胞定位分析
为了构建用于双分子荧光互补(BiFC)实验的载体,分别扩增并克隆ZmPP2Ac-2、ZmWEE1和ZmTE1的全长CDS,将其分别克隆到p2YN和2YC载体中,通过PacI和AscI线性化,与YFP的N末端和C末端融合。将这些质粒转化入农杆菌GV3101株,随后将农杆菌注射到四周龄的烟草(Nicotiana benthamiana)叶片中,使用MMA培养基(50 mM MES,10 mM MgCl2,20 μM乙酰茴香酸,pH 5.6)进行瞬时蛋白表达(Yu et al., 2019)。然后,使用Zeiss LSM880激光共聚焦显微镜(德国)在488 nm波长下对烟草表皮细胞进行成像。每种组合分析了三次生物学重复,并且使用空载体组合作为阴性对照。
对于亚细胞定位分析,我们将ZmPP2Ac-2、ZmWEE1和ZmTE1的CDS克隆到pB7WGF2载体中,使用LR反应生成35S::GFP-ZmPP2Ac-2、35S::GFP-ZmWEE1和35S::GFP-ZmTE1构建体,随后转化到GV3101中。然后,将携带35S::GFP-ZmPP2Ac-2、35S::GFP-ZmWEE1或35S::GFP-ZmTE1的GV3101与携带BZR1p:BZR1-RFP(作为细胞核和细胞质定位标记)的GV3101一起转化到烟草叶片中。使用Zeiss LSM 880激光共聚焦显微镜(德国)评估荧光信号。
体内磷酸化和去磷酸化实验
为了确定ZmWEE1和ZmPP2Ac-2是否影响ZmTE1的磷酸化状态,分别将GFP-ZmTE1、ZmWEE1-MYC和ZmPP2Ac-2-MYC转化到拟南芥叶肉原生质体中。收获原生质体,并在冰上用细胞裂解缓冲液(0.5 mM EDTA;10 mM Tris-HCl;pH 7.5;0.5% NP-40;1 mM PMSF;150 mM NaCl)裂解30分钟,每10分钟吹打一次。裂解液离心后,收集上清液,将ZmTE1与ZmWEE1一同用小牛肠碱性磷酸酶(CIAP)在30°C下处理30分钟。最后,将上清液加热至99°C,煮沸10分钟。通过phos-tag分离和检测磷酸化与去磷酸化的ZmTE1(Yu et al., 2019)。使用抗GFP抗体(TransGen Biotech,目录号HT801-02)检测ZmTE1的蛋白水平和磷酸化状态;ZmWEE1-MYC和ZmPP2Ac-2-MYC的蛋白水平则使用抗-MYC抗体(Abclonal,目录号AE010)进行检测。
统计分析
统计分析使用Student’s t检验进行(*P < 0.05,**P < 0.01,***P < 0.001),或使用单因素方差分析(ANOVA,P < 0.05;LSD和Duncan检验)。所有实验至少重复三次,数据以均值±标准误(SE)表示。
数据存取号
RNA-seq数据已在基因表达综合数据库(Gene Expression Omnibus)中提供,数据存取号为GSE181794。


















 1762
1762

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









