






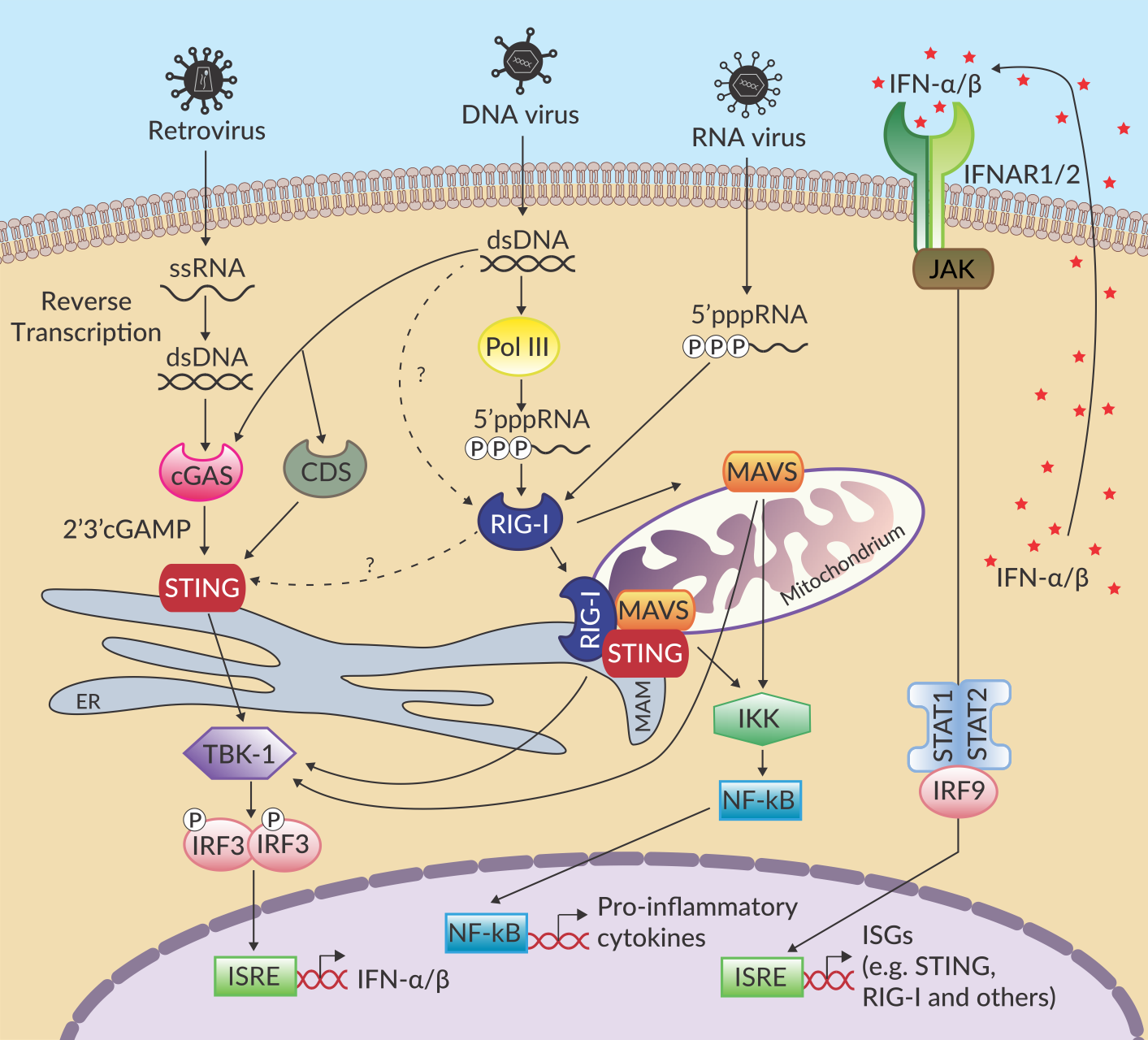
天然免疫系统对于限制病毒感染作用重大。其依靠若干组模式识别受体(PRRs)来识别病毒核酸1。这些PRRs包括胞浆DNA感受器(CDS),环鸟苷酸腺苷酸合成酶(cGAS),和细胞质RNA感受器视黄酸诱导基因I(RIG-I)。这些受体一旦被活化,将诱导不同的信号通路导致一系列抗病毒分子的产生。有趣的是,有证据表明,这些信号通路之间存在紧密联系以增强抗病毒反应。
感受器cGAS和RIG-I通过单独的接头蛋白来识别不同的核酸和信号。cGAS感受细胞质中异常DNA的存在和浓度,而后通过环二核苷酸2’3’-cGAMP2募集干扰素基因刺激因子(STING, or MITA/ERIS/MPYS)。RIG-I检测出5`-二/三磷酸末端未加帽的病毒RNA和短平末端双链部分的病毒RNA后,其与线粒体抗病毒信号蛋白(MAVS, or IPS-1/VISA/Cardif)1结合。它们激活后,cGAS/STING和RIG-I/MAVS将触发共同的信号级联反应导致I型干扰素(IFNs)和促炎性因子的产生。
在不同水平上均有报道称RNA和DNA通路之间存在复杂的相互作用。首先,若干组数据表明在人源细胞中,胞浆中富含AT的DNA以一种RIG-I/MAVS依赖的方式导致了I型干扰素的产生,而在小鼠细胞中则没有3-5。虽然RIG-I和dsDNA之间显示有直接的相互作用6,但是RIG-I的这种识别作用需要poly(dA:dT)通过RNA聚合酶III的转录作用实现4,5。此外,在STING缺失细胞中,应答胞浆双链RNA和病毒感染时I型干扰素产量的减少和免疫共沉淀实验中STING和RIG-I的直接结合共同证明STING参与RIG-I/MAVS应答途径7,8。已发表的数据表明,STING作为共同接头蛋白与MAVS在线粒体相关内质网膜(MAM)形成复合物,从而活化RIG-I7,9。此外,cGAS参与反转录病毒RNA10和胞浆RNA:DNA杂合体的识别11,并在晶体结构研究中显示其与合成RNA结合12。
除调节器与RNA和DNA途径物理上的互作之外,STING和RIG-I表达水平上的协同调节也有描述。事实上,合成的或病毒激动剂激活RIG-I后可诱导STING表达13,反之亦然,STING活化的下游信号通路可诱导RIG-I的表达,有趣的是,上调的RIG-I参与STING表达的负反馈调节以防止过度的免疫反应14。
总之,cGAS/STING和RIG-I/MAVS在生理和功能上是相互联系的。RNA和DNA检测中的互作和随后的信号级联反应在应对微生物核酸的多样性和抵抗病毒逃逸机制上都是至关重要的。此外,RIG-I和STING之间的互作可能有助于平衡细胞的抗病毒反应和自身节律。值得注意的是,RIG-I和STING之间的互作在物种和细胞类型之间差异很大,这可能反映了病毒及其目标细胞间的协同进化作用。核酸感应途径的激动剂或抑制剂的未来发展需要严格检查其对每个核酸感受器的影响。
原文出处:RIG-I 与 STING 的联盟


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









