





 本文是MIT团队关于代谢工程的综述,探讨了代谢通路解析、优化和设计,以及代谢工程在宿主选择、产量提升等方面的应用。研究强调了代谢工程在理解和控制微生物代谢网络中的重要性,通过理性设计和组合技术提高产品产率。未来代谢工程将受益于基因组学和新型宿主的开发,以实现更高效的大规模生产。
本文是MIT团队关于代谢工程的综述,探讨了代谢通路解析、优化和设计,以及代谢工程在宿主选择、产量提升等方面的应用。研究强调了代谢工程在理解和控制微生物代谢网络中的重要性,通过理性设计和组合技术提高产品产率。未来代谢工程将受益于基因组学和新型宿主的开发,以实现更高效的大规模生产。

Department of Chemical Engineering, Massachusetts Institute of Technology, Cambridge,
Massachusetts 02139; email: woolston@mit.edu, sedgar@mit.edu, gregstep@mit.edu
Annu. Rev. Chem. Biomol. Eng. 2013. 4:259–88
这是MIT团队较早的一篇代谢工程综述。SE随后又参与了MIT团队的一篇NBT(2015),
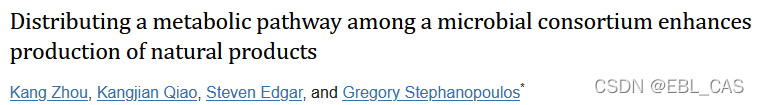
在这篇研究论文中,该团队解决了这样一个问题,即把多个合成通路整合到一个宿主中,往往面临着某些反应不能最优化的情形。这种问题在自然界就是功能互补,个体通过成为群落功能过程的一部分大家都实现最优化。研究正是利用了这种思想,把一个通路不同环节分装到两个模式宿主通过共培养实现终产物taxanes。产率达到了33 g/L。
2016年该团队以SE为一作发表了taxanes关键合成酶的突变改造。通过改造该团队提升了taxanes前体taxol的合成选择性。(ACS Synthetic Biology 6(2) DOI:10.1021/acssynbio.6b00206—)
SE和GS2018年在PNAS探讨了Isoprenoid合成通路问题。Isoprenoid是一类调控非常复杂的二次代谢产物,该团队构建了相关通路并测试了合成前体的效率。
总的来讲,以MIT的软硬件条件,这些伙计不算高产,但干货满满。向他们学习!
正文意译
从人类第一次成功将一个外源基因导入细菌细胞,我们就意识到微生物细胞可以成为众多化工产品的微型工厂。但是很快人们发现,尽管复杂如胰岛素的分子可以通过一个基因的过表达生产,但简单如乙醇的小分子生产却涉及众多基因的调控。就在人们热衷于利用此技术生产医药分子的同时,1980年代,也有研究关注了这项技术背后的代谢复杂性及其调控问题,如代谢通路的热力动力学,代谢步骤的动力控制,遗传通路的设计等等。这些活动有两篇代表性论文,其很关键性地推动了代谢工程的建立。(两篇论文如下)
Stephanopoulos G, Vallino J. 1991. Network rigidity and metabolic engineering in metabolite overproduction. Science 252(5013):1675–81
Bailey JE. 1991. Toward a science of metabolic engineering. Science 252(5013):1668–75
代谢工程与遗传工程不同,前者关注整合的代谢通路和调控网络,后者更多的关注单一基因或酶的操纵。
代谢工程从诞生时起就有显著的工业目标,因为其一开始就意在满足更加有效的催化反应用于生产感兴趣的工业品,如能源,化工品和药品。使用生物催化生产一个产品很容易,但是实现比率、产率和速率的最优化是很难的。这种困难导致代谢过程不得不努力发展代谢通路的设计、构建和最优化。
今天的代谢工程,已经不仅仅是生产贵重的稀有的化工品,而是能够放大尺度生产大宗化工品。未来的代谢过程将大幅度提升生产能力和应用范围。一方面,基因合成技术和异源表达越来越成熟,另一方面,组学的发展为更全面了解微生物的代谢地图提供了技术准备,这包括模式生物也包括稀有微生物。组学提供的地图和基因多样性信息将为代谢工程提供无限可能。
第一部分
过去和现在
解析和评估代谢通路:实现底物向产品的转化
确立生产某个化合物之后,第一个问题是什么?寻找该化合物的合成通路。对合成化学家一样。该化合物必须对生物体是无害的,并且相关酶能够在异源宿主洪表达。考虑到与化合物生产有关的反应通路涉及广泛,我们必须细数通路的个数并且评估通路。1990年代人们首次完整确定了赖氨酸的可能合成通路,评估找到了最高产率的可能通路,克服了瓶颈一种更为通用的发现这些通路的算法,叫BINCE,biochemical network integrated computational explorer,在2000左右出现。该方法考虑了底物和产物的化学结构和酶学分类系统,实现了通路的从头设计。分析芳香氨基酸的合成通路表明,从底物出发可以合成众多化合物,并发现了合成苯丙氨酸,酪氨酸和色氨酸的新通路。通过基团贡献理论可以估算酶反应的自由能变化,这种热力学可行性评估被用于评价这些通路。(相关文献如下)
Hatzimanikatis V, Li C, Ionita JA, Henry CS, Jankowski MD, Broadbelt LJ. 2005. Exploring the
diversity of complex metabolic networks. Bioinformatics 21(8):1603–9
MavrovouniotisML. 1990. Group contributions for estimating standard Gibbs energies of formation of biochemical compounds in aqueous solution. Biotechnol. Bioeng. 36(10):1070–82
Mavrovouniotis ML. 1991. Estimation of standard Gibbs energy changes of biotransformations. J. Biol.Chem. 266(22):14440–45
理解代谢通量和代谢控制
获得通路图后,工程改造生态型的下一步是分析该通路的反应速率和控制结构,在此基础上提出理性修改靶点。代谢通路分析可以提供不同条件下代谢通量偏离对照条件下的程度,然后确定动力学限速步骤。
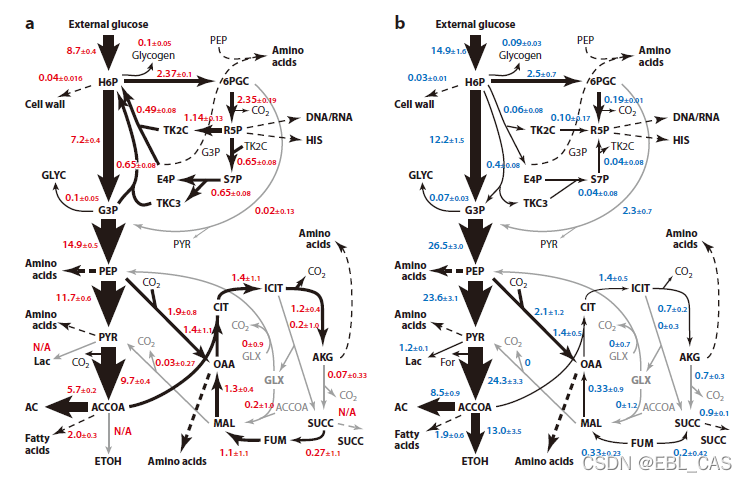
基于碳13获取的大肠杆菌好氧和厌氧条件下生长的碳通量。黑箭头的大小与该步骤通量与蔗糖吸收速率的比值成正比。
代谢图和通量分析获得的信息最终将用于选择基因调控的靶点,这些信息的核心不是通量本身,而是偏离对照的值和通路的可塑性。
最近发展的基因组尺度代谢计量比模型,主要是为了取代c13代谢通路测定方法。同位素方法虽然准确,但是价格昂贵。模型手段比实验手段优越的地方还有:实验数据需求较少,可以通过理论模型设计敲除靶点,一些组学数据已经存在,另外通量平衡分析往往不能得到产物通量,而是得到最大生物量产率。最近的一个基因组代谢图预测了大肠杆菌的2077个反应和1039个代谢产物。基因组代谢图已经有很多模式和非模式生物具备,如
Saccharomyces cerevisiae (36), Bacillus subtilis(37), Clostridium acetobutylicum (38), Corynebacterium glutamicum (39), Arabidopsis thaliana (40), and even humans (41).
F¨ orster J, Famili I, Fu P, Palsson BØ, Nielsen J. 2003. Genome-scale reconstruction of the Saccharomyces cerevisiae metabolic network. Genome Res. 13(2):244–53
de Oliveira Dal’Molin CG, Quek L-E, Palfreyman RW, Brumbley SM, Nielsen LK. 2010. AraGEM, a genome-scale reconstruction of the primary metabolic network in Arabidopsis. Plant Physiol. 152(2):579–89
代谢工程的理性设计
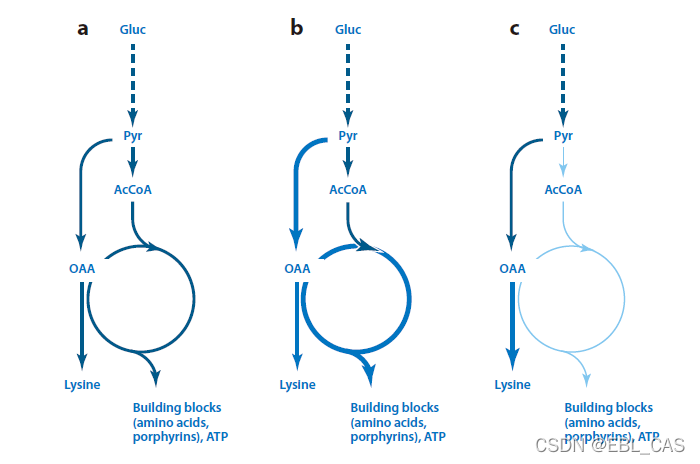
赖氨酸的代谢工程是理性设计的典型案例。A代表野生型,b是pyc过表达菌株,c是ask过表达菌株。过表达aspartate kinase增加了赖氨酸的产率,提升1.5倍,同时维持了与对照的同等生长速率。
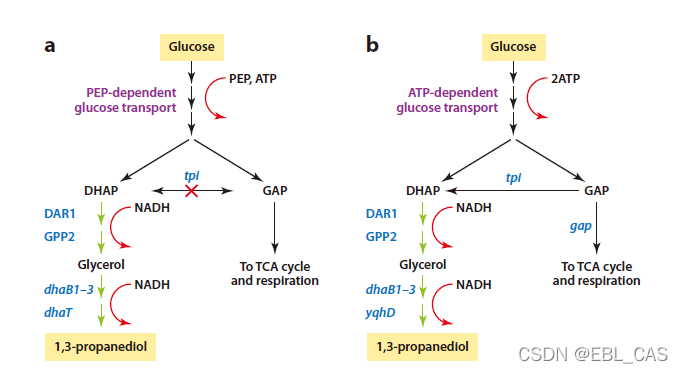
另一个例子是通过蔗糖生产丙二醇的代谢工程。A是前体菌株,b是改造菌株。首先,敲除非产物反应避免丙三醇进入中心碳代谢,其次,glucose phosphotransferase敲除掉。最后一些酶进行了优化。这些改进导致目标产物产率为51%,速率为3.5g/liter/h,titers 135 g/liter
再来看一个改造酵母的例子,酵母本不能同化木糖,代谢工程改造使得其能够通过木糖生产乙醇。通过表达环糊精运输蛋白和胞内葡糖苷酶,酵母实现了二糖的直接同化。
目前大部分代谢共同通路优化都是基于在联合表达组成性基因的基础上,优化部分目标步骤,通过过表达,敲除或移除抑制调控子。现在的进步主要在基因合成和操作技术,也包括生信的完善,如启动子,核糖体结合位点,终止子等等都已经有了清单。
代谢工程的组合技术
大多数情况下,上述技术手段可以实现较好的代谢工程结果。组合技术有利于解决一些复杂的通路。早期的技术包括随机突变,如盘尼西林的生产效率可以达到50 g/liter,约是自然菌株的4000倍。2000左右诞生的逆向代谢工程,拓展了理性再造菌株的思路。该途径使用广泛,非常类似蛋白定向进化。有三个因素影响逆向代谢工程,高通量筛选,高质量表型库,快速鉴定哪个表型对应遗传突变。
不断进步的工具和生物学的基础研究进展,在进一步推动着代谢工程的发展,分子生物学、合成生物学、系统生物学越来越先进,代谢工程的产品也越来越多样。那么,未来代谢工程如何发展?
替代宿主
目前主要的代谢工程在少数模式生物完成,尤其大肠杆菌和酵母。这与其易于驯化有关,如实验室容易培养,基因操作技术主要在这些生物中发展,基因组测序较完善等。密码子优化技术也较为清晰。
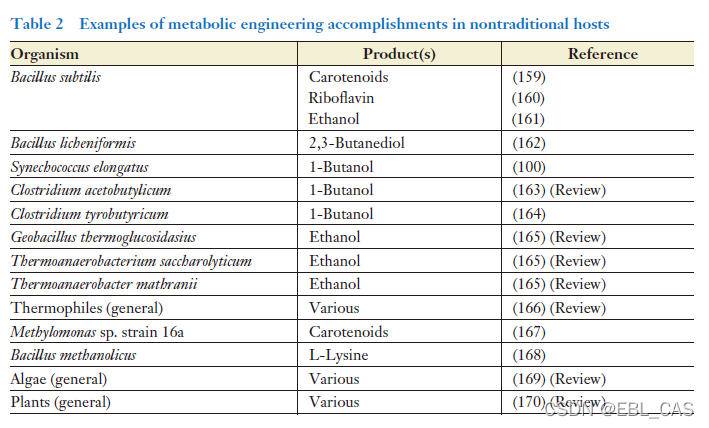
其他生物的代谢工程潜力也有一些研究,尽管不多,但意义重大。尤其一些极端菌,具有重要工业价值。如高温菌,具有在高温条件下保持较高酶活性的能力。高温条件可以排除中温微生物的生长,从而避免杂菌污染。高温也有利于挥发性目标产物的生产,减少蒸馏回收环节。Thermoanaerobacter saccharolyticum and Geobacillus thermoglucosidasius最近被用于乙醇的高温生产,该菌以纤维素为原料生产糖作为其底物。
Shaw AJ, Podkaminer KK, Desai SG, Bardsley JS, Rogers SR, et al. 2008. Metabolic engineering of a
thermophilic bacterium to produce ethanol at high yield. Proc. Natl. Acad. Sci. USA 105(37):13769–74
Cripps RE, Eley K, Leak DJ, Rudd B, Taylor M, et al. 2009. Metabolic engineering of Geobacillus
thermoglucosidasius for high yield ethanol production. Metab. Eng. 11(6):398–408
还有一些微生物如clostriduium可以通过ABE通路acetone-butanol-ethanol生产天然溶剂,有些对大肠杆菌是有毒的,因此这些菌株是生产这些溶剂的良好宿主。
最近一个蓝细菌被用于生产丁醇,底物为二氧化碳,这是首个直接从其他到液体的能源生产平台。
Lan EI, Liao JC. 2011. Metabolic engineering of cyanobacteria for 1-butanol production from carbon
dioxide. Metab. Eng. 13(4):353–63
为什么把一些有用的通路转入常见宿主如此困难?主要原因是在这些宿主中进行遗传操控的工具还不成熟。随着其他源生物基因组被全测序,现在将其基因资源导入易培养菌株的宿主越来越容易。现在促进异源表达的启动子和终止子越来越容易获得,筛选成为可能。PyrF许多生物中存在,可以作为阳性或阴性筛选标记。阻止基因转化的因素如主动限制修改系统active restriction-modification systems,现在也可以通过基因组信息发现并减轻其影响。最近发现允许两个物种中基因横向转移的同源重组成分在所有生物体中都存在,这位异源导入提供了巨大机会。自杀基因就是一种应用。
如何选择宿主?是转入传统宿主,还是转入更适合生长但技术了解较少的宿主?主要原则是考虑待操作的性状在非标准生物的复杂性。如人们曾尝试生产自养大肠杆菌,这样就可以从二氧化碳直接生产化工产品。但通路的复杂性表明在蓝细菌中改造更为可行。但当一个性状可以被简化为一个单一基因或简单基因簇时,则具有可行性。
提升生产浓度的技术
优化反应酶的空间格局是提升通路通量的有效方法。这一方法可以降低中间产物抑制因子,减少中间产物向非目标产物通路转化,并帮助克服扩散限制因素。有三种办法可以共定位通路酶蛋白:产生二聚体融合蛋白,将蛋白结合为框架,或者将其定位到亚细胞器。
最近有研究将酰基辅酶连接酶与对称二苯代乙烯合成酶融合,并连接到一个三氨基酸序列,获得了白藜芦醇生产效率的15倍提升。动力学和结构研究表明,这种操作改进了酶活性位点空间接触性并没有改进酶活性。
通过将甲羟戊酸通路酶形成框架,其效率提升了77倍。同样的策略整合三个通路酶,d型葡萄糖二酸的生产效率提升了5倍。框架本质上提升了通路酶的互作效率,而不仅仅是实现了酶直接反应步骤的串联。这种策略也用于构建自然界没有的通路。
目前最复杂的共定位方法是方法是区隔化。微生物没有真正的亚细胞器,但是微型区隔是有的。某研究将Halothiobacillus neapolitanus的十个二氧化碳固定基因异源表达在大肠杆菌中,实现了活性再构。当前细胞器改造的主要工作集中在氮端序列靶向定位,如过氧化物酶,液泡,线粒体和叶绿体。最近有研究表明,细胞器定位技术可以实现植物萜类在酵母中的合成,效率提升了8到20倍,可能的机理是提升了前体的可利用性。细胞器定位策略真正的潜力是再造细胞器而不是解码现有细胞器。但迄今为止没有真正全新的合成细胞器报导。通过直接注入、入侵等手段,最近有研究表明蓝细菌可以在动物细胞系中存活,意味着细胞器的法则有可能通过合成细胞器改变。

系统层次控制下的全新通路设计
受体-调控因子动态系统的构建是本领域的重要进展,最近被用于了生物燃料脂肪酸乙酯的生产。
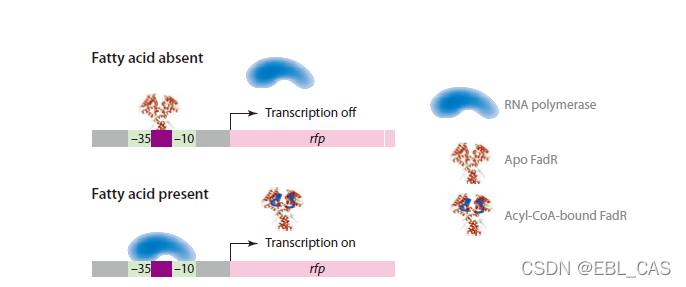
通路基因受统里前体acyl-coa的调控,该调控使得通路酶的表达与底物可利用性成相关性。没有前体时,fadr与聚合酶结合并阻止转录。过量的前体则于fadr结合,减少阻碍并允许通路酶结合。

















 6738
6738

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









