






萜类化合物是人类药物的关键成分,是最大的天然产物家族。真菌是萜类化合物的重要来源,但许多相应的生物合成基因簇(BGCs)在实验室条件下是沉默的。同源激活和异源表达等策略通常用于激活单个簇,效率低下。
为解决这一问题,武汉大学刘天罡教授团队使用米曲霉作为底盘开发了一种自动化和高通量(auto-HTP)生物基础工作流程,能够有效地进行基因组挖掘、BGCs的表征和生物活性真菌萜类化合物的鉴定。相关研究成果(Efficient exploration of terpenoid biosynthetic gene clusters in filamentous fungi)发表在Nature Catalysis上。

天然产物的研究范围已经从探索可以从生长的培养物中分离出来的高丰度分子扩展到开发和使用生物信息学分析和计算工具,如antiSMASH和SMURF,这些工具有助于揭示大量转录沉默或神秘的生物合成基因簇(BGCs),激活这些BGCs并开始生产其生物活性化合物的策略对于释放自然界化学资源的潜在力量至关重要
BGC重构提供了一种这样的方法,剥离了对代谢产物生物合成途径的天然调节。其中异源表达是BGC重构的主要方法,能够对由此产生的聚酮、肽和萜类化合物进行机制研究。其他新兴的策略,包括生物信息学和计算框架、人工染色体、基于代谢评分的方法和HEx(异源表达)平台,进一步提高了我们挖掘隐性BGC的能力。然而,现有的由大肠杆菌和酿酒酵母构建的底盘不太适合生产真菌天然产物,因为它们表达膜定位的细胞色素P450酶(CYP450)较差,缺乏必要的氧化还原伴侣蛋白,并且不包含识别和剪接丝状真菌富含内含子基因的适当机制,因此,这些底盘主要在萜烯环化酶的表达、合成核心碳氢化合物骨架方面取得了成功
丝状真菌米曲霉(AO)通常被认为是安全的,并为探索真菌天然产物提供了改进的细胞环境。事实上,它是通过CRISPR–Cas9介导的基因编辑从丝状真菌中挖掘单个萜烯环化酶或整个BGC的理想底盘。然而,即使在AO中,前体(例如,异戊烯基焦磷酸和二甲基烯丙基焦磷酸)的有限供应也使得获得足够的萜类化合物用于生化和功能研究具有挑战性,尤其是对于需要多个生物合成步骤的后期衍生物。
在此,研究团队使用AO NSAR1作为v.1.0底盘,开发了一种自动化和高通量(auto-HTP)生物基础工作流程,作为一种新的基因组挖掘策略。
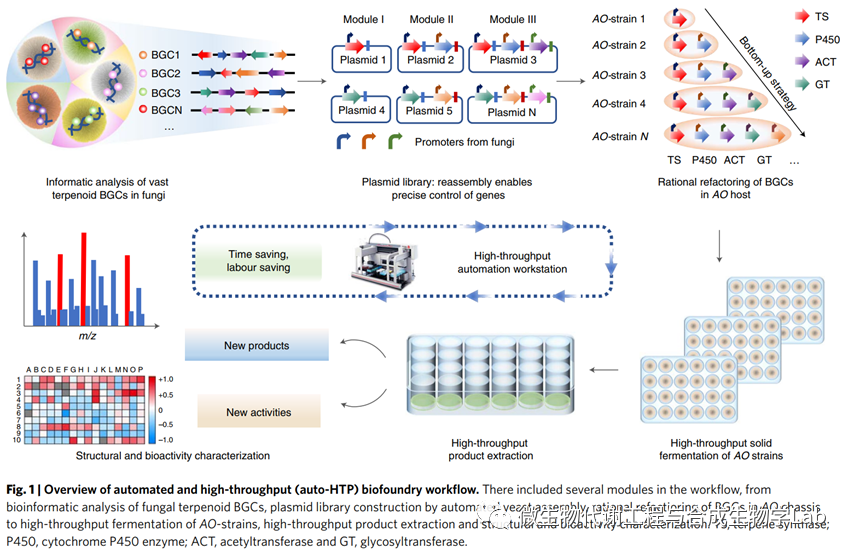
研究团队使用auto-HTP生物流体工作流程重建了39个萜类BGCs,并评估了208个工程菌株产生的萜类化合物的活性和结构新颖性。功能筛选揭示了一种抗炎化合物,芒果醇J,在体外和体内都具有活性,而其工程菌株为快速鉴定参与芒果醇J及其家族成员合成的酶提供了一个简单的框架。最后,我们开发了一种代谢前体滴度提高的第二代AO底盘(v.2.0),它是一个通用而有效的平台,可以过量生产萜类化合物,如芒果醇J。这种组合策略代表了丝状真菌中萜类化合物的大规模基因组挖掘的有效方法,并且还为开采其他天然产品提供了一种通用方法。
萜类BGCs的生物信息学分析及重构原理:
AO NSAR1(niaD−,sC−,ΔargB,adeA−)是一种营养缺陷型菌株,已被广泛用于丝状真菌基因的异源表达,被选为挖掘在五种内部基因组测序丝状真菌中发现的萜类BGCs的宿主,即Colletotrichum gloeosporioides ES026、Alternaria alternataTPF6、Fusarium graminearumJ1-012,Trichoderma virideJ1-030和Aspergillus flavipes。根据antiSMASH分析,初步预测这五种真菌含有54个萜类BGCs。如果推定的BGC含有缺失已知保守基序的萜烯合成酶(I类的DDXXD/E和NSE/DTE基序和II类萜烯合成酶的DXDD基序)或具有不完整的功能域,则排除了它们,剩下39个BGC被预测产生倍半萜、二萜和倍半萜。
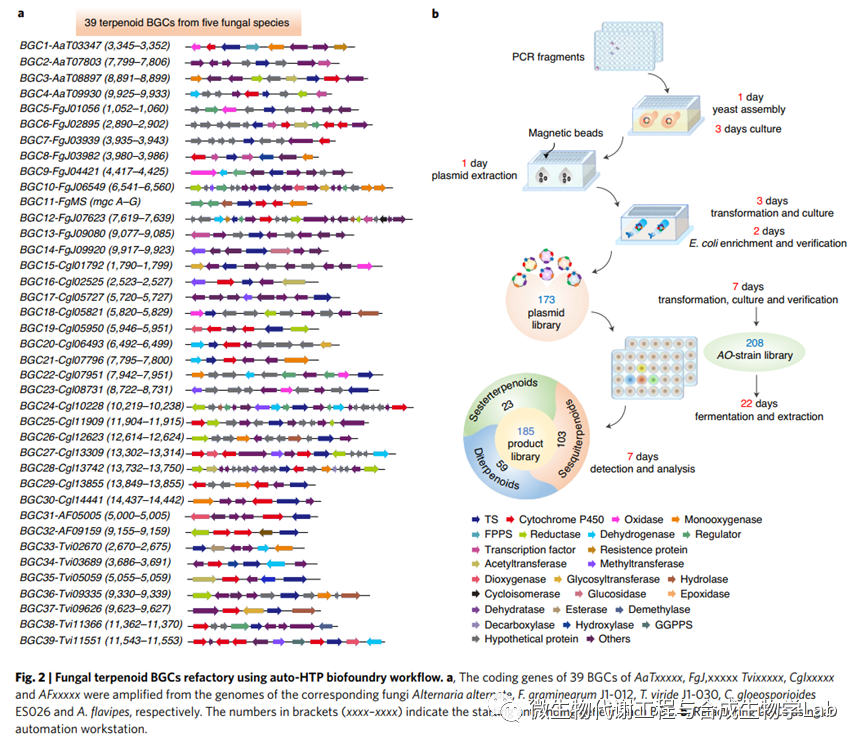
接下来,研究团队构建了一个重构39个基因簇的计划。然而, 萜类BGCs在AO中的异源表达经常受到识别BGCs的天然启动子的挑战的阻碍,导致启动子的随机选择而不考虑如何精确控制功能基因的表达。在这里,我们表征了一系列可以用来代替天然序列的启动子。我们使用β-葡萄糖醛酸酶作为读数来确定这些启动子的强度,顺序为hlyA>oliC>amyB>glaA>enoA>gpdA>agdA>trpC>alcA。接下来将BGCs中的功能基因分类为上游萜烯合酶模块,其中低聚异戊二烯的环化产生具有多个立体中心的多环烃或醇,中游氧化模块,其中核心被氧化形成双键,羰基和醇基,以及下游模块,其中任选的后官能化步骤(例如酰化、糖基化等)形成最终的复合物结构。结合这些结果提供了一种替代调节基因的方法,将功能基因置于强组成型(hlyA)和诱导型(amyB,glaA)启动子的控制之下。在自动HTP生物试剂的帮助下,将同一模块中的基因放置在单独的质粒中,或共同构建到质粒中,以进行所有可能的组合。在组合来自不同模块的质粒后,我们将BGC重构为菌株库,其中包含萜烯合成酶基因作为基本起点,然后单独添加来自同一BGC的其他下游基因。这种方法不仅可以根据产生的化合物来破译每个(部分)簇的功能,还可以确定每个新酶的具体作用,并检查产生的萜类化合物的生物活性。
萜类BGCs的自动化和高通量生物基础工作流程:
生物基广泛用于DNA组装、酶进化和菌株工程。它们在天然产物生物合成中的应用可能将使人们能够以自动化和高通量的方式挖掘大量隐蔽的BGCs。自动HTP生物流体工作流程使用先进的液体处理技术来加速PCR扩增、使用酵母同源重组的质粒构建和AO原生质体的转化以重构萜类BGC的过程(图2b)。使用Biomek FXP实验室自动化工作站(Beckman Coulter)作为自动平台进行PCR扩增、PCR产物回收和质粒提取,以及大肠杆菌、酿酒酵母和AO原生质体的转化。使用分子装置Qpix 460来采集菌落。使用自动HTP生物基础工作流程,我们在两轮后获得589个片段(98%的设计序列)自动化PCR扩增;这些包括来自BGC的质粒骨架、启动子、终止子和功能基因。剩下的9个片段可能受到长基因片段和引物特异性低的限制,手动扩增。通过同源重组将片段在酿酒酵母中组装成173个质粒。将组装好的质粒转化到大肠杆菌中进行复制,然后自动提取。接下来,我们将不同的质粒组合到AO原生质体中,合并不同组和顺序的基因,产生208个不同的菌株。
因此,在自动化生物基础工作流程的帮助下,将39个BGC重构为足够的组合,一次完全显示每个BGC的代谢中间体和最终产物。其中,经过两轮构建获得166个(96%)质粒和195个(96%的)AO菌株。剩下的7个质粒和13个菌株分别含有多个基因和大质粒,是人工构建的。从PCR扩增到AO菌株的最终构建的整个过程需要17天,而每一步都进展顺利。最后,将工程化AO菌株在含有固体大米培养基的24个深孔板中在30°C下孵育2周。之后使用丙酮和乙酸乙酯从培养物中提取两次化合物,并浓缩以产生粗提取物。使用气相色谱-质谱(GC–MS)和高分辨率电喷雾电离-质谱(HR–ESI–MS)检测了208株AO菌株中萜类化合物的产生。39个BGC中的26个共检测到185种不同的萜类化合物(1–185),其中47种仅通过GC–MS检测到,84种仅通过HR–ESI–MS检测,54种同时通过GC–MS和HR–ESI-MS检测到(扩展数据图4)。此外,103、59和23分别为倍半萜、二萜和倍半萜(表1)。与先前纯化的化合物的质谱分析和国家标准与技术研究所图书馆(https://webbook.nist.gov/chemistry/)的比较表明,22种萜类化合物是先前报道的化合物。通过PCR扩增和参考对阴性结果的菌株进行复核,以消除实验过程造成的任何影响。

AO底盘细胞2.0,用于mangicol J的高产:
AO底盘1.0是挖掘各种BGC并进行生物活性初步探索的理想起点。然而,对特定化合物的详细研究需要更有效的生产;例如,菌株AO-Y51和AO-Y52分别只积累0.66 mgl-1的mangicdiene和0.08 mgl-1的mangicol J,这不足以进行后续研究。先前的研究表明,优化甲羟戊酸(MVA)途径可以明显提高萜类化合物的滴度,其中tHMG1和Idi是限速酶。为了扩大化合物的生产,我们系统地设计了AO的MVA途径,以建立一个有效的底盘2.0,用于过量生产mangicdiene和mangicol J。
研究人员通过改良菌株,改进了mangicdiene核心架构的生产。通过构建了含有整个内源性MVA途径、额外三个拷贝的tHMG1和mgcD的质粒,并将它们随机插入AO NSAR1的染色体中,以产生菌株AO-S81至AO-S84。同时,在先前开发的CRISPR–Cas9方法的基础上,开发了CRISPR–Cas9介导的位点特异性整合系统,并通过同源重组以85%的效率整合了14kb的表达框(来自pSC247的片段)。对于位点特异性整合,使用高表达基因座(hot spots(HS)),HS201、HS401、HS601和HS801作为我们的靶位点,以促进功能基因的表达。具体而言,通过整合了整个MVA途径,并在AO的不同HS位点额外整合了四个tHMG1拷贝,以产生AO-S95。然后我们将mgcD的一个拷贝整合到AO-S95的HS801位点中以产生AO-S96。与亲本菌株AO-Y51相比,mangicdiene的滴度在AO-S96中从0.66 mgl-1增加到27.38 mgl-1,增加了41倍,在AO-S84中为87.84 mgl-1增加到133倍(图6b、c和扩展数据图9)。这些滴度明显高于在大肠杆菌和蜡样芽孢杆菌中检测到的倍半萜,为丝状真菌来源的萜类BGCs的高通量基因组挖掘提供了理想的平台。
接下来,使用功能基因的随机插入和位点特异性整合创建了用于过量生产mangicol J的菌株。对于随机插入,将两个拷贝的mgcE随机插入AO-S84的染色体中以产生菌株AO-S94。对于位点特异性整合,我们将一个拷贝的mgcD和两个拷贝的mgcE整合到AO-S95的HS801位点中,以产生AO-S97。与菌株AO-Y52相比,mangicol J的滴度在AO-S98中从0.08增加了112倍,达到8.93 mgl-1,在AO-S94中增加了151倍,达到12.09 mgl-1(图6c、d和扩展数据图9)。
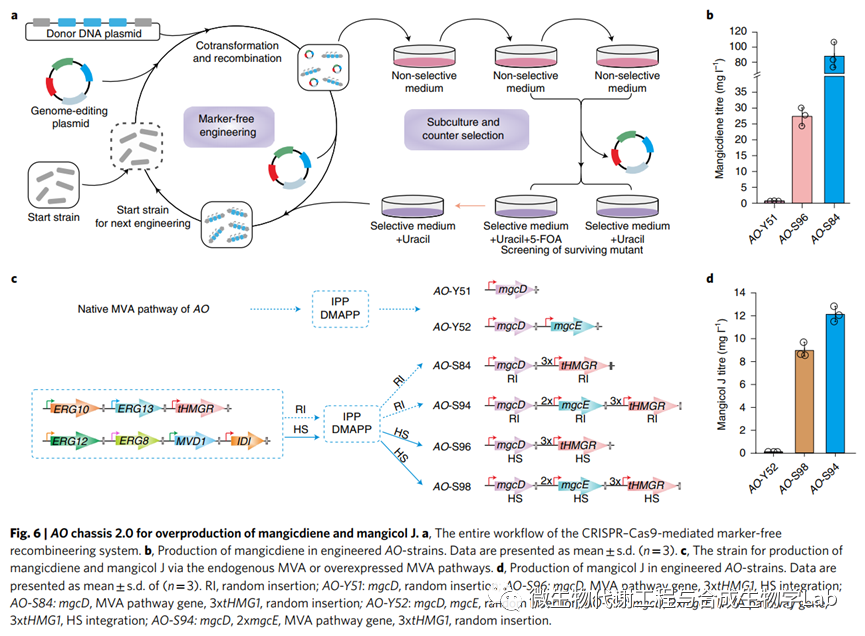
最后,从菌株AO-S84中敲除mgcD以产生AO-S85。因此,该菌株作为表征基因和萜类结构的通用底盘2.0,为真菌特异性萜类化合物的过量生产提供了丰富的前体;HS特异性整合菌株AO-S95作为真菌衍生萜类化合物稳定过量生产的有力补充。通过将BGC37-Tvi09626基因簇重构到AO-S95菌株中,证实了这种先进底盘的通用性。与底盘1.0相比,氧化产物和糖基化产物的滴度明显提高。
总之,该研究提供了一种高效的天然产物生物合成基因簇快速挖掘的自动化手段,能够很好的运用到其他天然产物生物合成基因簇的挖掘中。(刘威摘译)
文章来源:
DOI: https://doi.org/10.1038/s41929-022-00762-x
往期精品(点击图片直达文字对应教程)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
















































 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









