






点击蓝字 关注我们
解码免疫T细胞的作用:改善代谢相关脂肪性肝病的新领域

iMeta主页:http://www.imeta.science
综 述
● 原文链接DOI: https://doi.org/10.1002/imt2.76
● 2023年1月18日,北京中医药大学李晓骄阳团队在iMeta在线发表了题为“Decoding the role of immune T cells: A new territory for improvement of metabolic-associated fatty liver disease”的文章。
● T细胞在代谢相关脂肪肝病(MAFLD)中的作用越来越受到全球学者的重视。本研究指出了几种基于T细胞的治疗方法,包括但不限于过继转移T细胞、粪菌移植和药物治疗,这项研究有助于更好地理解T细胞在MAFLD进展中的重要作用以及相应的治疗方向,并为进一步的药物开发提供了潜在的参考。
● 第一作者:刘佳、丁明宁
● 通讯作者:李晓骄阳(xiaojiaoyang.li@bucm.edu.cn)
● 合作作者:白金钊、罗苒艺、刘闰平、曲姣蓉
● 主要单位:北京中医药大学生命科学学院
亮 点
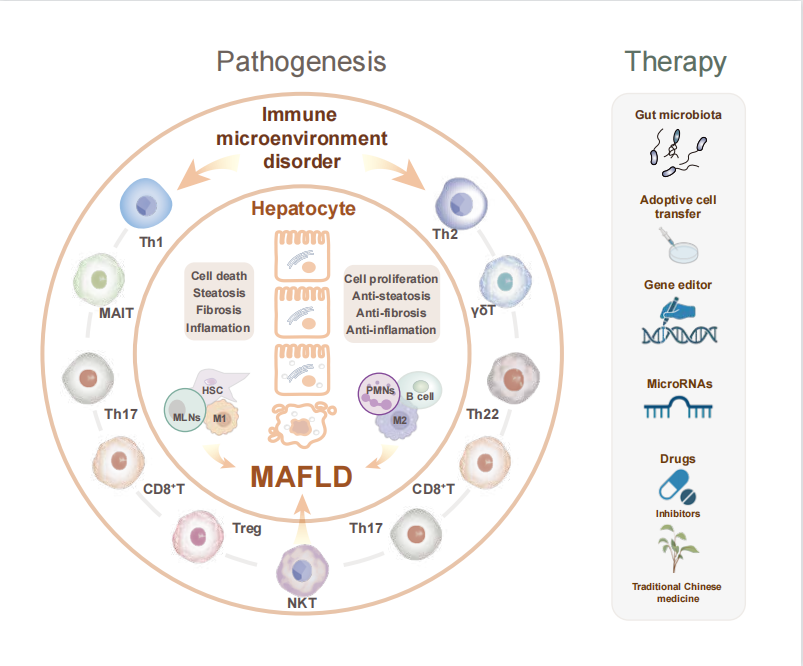
● 代谢相关脂肪性肝病(MAFLD)是由代谢失调引起的最常见的脂肪肝病
● T细胞(CD4/8+T细胞、Treg细胞、γδT细胞、NKT细胞、MAIT细胞)稳态失调会导致严重的肝脏脂肪变性和脂肪堆积
● 肠道菌群调节或中药等药物治疗可以恢复肝脏T细胞的免疫微环境以改善MAFLD
摘 要
代谢相关脂肪肝病(MAFLD)顾名思义是与代谢功能障碍相关的肝病,可以更准确的定义这类肝病而取代非酒精性脂肪肝病(NAFLD)这一旧称。由于饮食变化、代谢紊乱和缺乏一线治疗药物,MAFLD的患病率逐年上升,已经影响到全球四分之一的人口,并成为医疗系统负担的主要经济问题。目前,除了胰岛素抵抗、氧化应激和脂肪毒性等常见致病因素外,免疫细胞,特别是T细胞在MAFLD中的作用越来越受到全球学者的重视。基于免疫T细胞的不同分类和病理生理效应,我们全面分析了它们对肝脏炎症微环境和MAFLD进展的双向调节作用。MAFLD和T细胞之间的这种相互作用也与肝-肠免疫串扰和肠道微生物群稳态有关。此外,我们指出了几种基于T细胞的治疗方法,包括但不限于过继转移T细胞、粪菌移植和药物治疗。特别要强调的是,药物治疗中天然产物和中药复方的重要作用。总之,这项研究有助于更好地理解T细胞在MAFLD进展中的重要作用以及相应的治疗方向,并为进一步的药物开发提供了潜在的参考。
视频解读
Bilibili:https://www.bilibili.com/video/BV1nv4y1k7No/
Youtube:https://youtu.be/sSXilRsBVAA
中文翻译、PPT、中/英文视频解读等扩展资料下载
请访问期刊官网:http://www.imeta.science/
全文解读
引 言
代谢相关脂肪肝病(MAFLD)是遗传、代谢功能障碍引起的一种进展性的脂肪肝病,如果不及时治疗,可能发展为肝硬化甚至肝细胞癌(HCC)。MAFLD是一个代替非酒精性脂肪肝疾病(NAFLD)的新定义,它强调了代谢因素在疾病病因中的作用,而后者不能准确反映脂肪肝疾病的综合机制和严重程度,并在一定程度上阻碍了医学发展的进展。大量的MAFLD临床报告显示代谢功能障碍的肝脏中出现了过度脂肪堆积,而MAFLD会增加脂肪性肝炎和肝纤维化的发生概率。与临床上的活检困难不同,动物模型中MAFLD的诊断标准通常是病理组织切片和脂肪肝血液中标志物的检测,包括丙氨酸转氨酶(ALT)、天冬氨酸转氨酶(AST)、总胆固醇(TC)和甘油三酯(TG)。有数据表明,MAFLD的数量正在持续增加,已影响全球25%的人口,到2030年仅在美国地区病例可能会扩大到1.009亿例,这对经济发展、公共卫生和社会发展等均产生重大的影响。越来越多的证据表明,年龄、不健康饮食、代谢状态、细胞内氧化应激、线粒体功能障碍、炎症细胞浸润和肠道微生物群组成是MAFLD的主要危险因素,并影响MAFLD生理病理过程。尽管如此,MAFLD的发病机制仍不明确且缺乏FDA批准的临床治疗药物。虽然有大量药物被认为是治疗MAFLD的良好候选药物,但大多数药物主要集中于PPARγ或GLP-1受体的激活和胰岛素致敏,并伴有一些严重的副作用。随着研究的深入,发现与MAFLD发病机制密切相关的另一个重要因素是免疫调节T细胞的功能障碍。例如,Van Herck等人发现,随着MAFLD的发生CD4+T细胞和CD8+T细胞急剧增加,并异常分泌促炎因子如干扰素γ(IFN-γ)和肿瘤坏死因子α(TNFα)。根据另一份报告表明,MAFLD小鼠模型中的免疫稳态被破坏,如CD4+T细胞亚群失衡和致病性细胞因子(特别是IFN-γ和IL-17A)过度分泌。因此,了解参与MAFLD进展的T细胞的复杂性可能为治疗干预提供新的途径。
T细胞特异性免疫反应
免疫系统是介导过敏、肿瘤免疫和自身免疫等免疫应答的细胞和分子的总称,大致可被分为先天免疫和适应性免疫。其中,T细胞从骨髓内的多能干细胞(PSCs)分化而来,并布散到全身,以协调免疫系统的稳态。T细胞受体 (TCRs) 由α和β蛋白 (αβ T细胞) 和T细胞特异性共受体CD3复合物组成,该复合物识别特异性信号以启动细胞生物学反应。幼稚T细胞已被确定为胸腺相关免疫反应的重要组成部分,其在抗原刺激下将退出静止状态,进入克隆扩增和分化状态。此外,T细胞的分化通过TCR识别并与抗原提呈细胞 (APC) 的主要组织相容性复合体 (MHC) 分子结合,以确保通过特异性识别抗原产生免疫应答。此外,T细胞被划分为CD4+T辅助细胞,CD8+T细胞和先天T细胞,包括γδ T细胞、自然杀伤T细胞 (NKT) 细胞和粘膜相关不变T细胞 (MAIT)细胞(图1)。
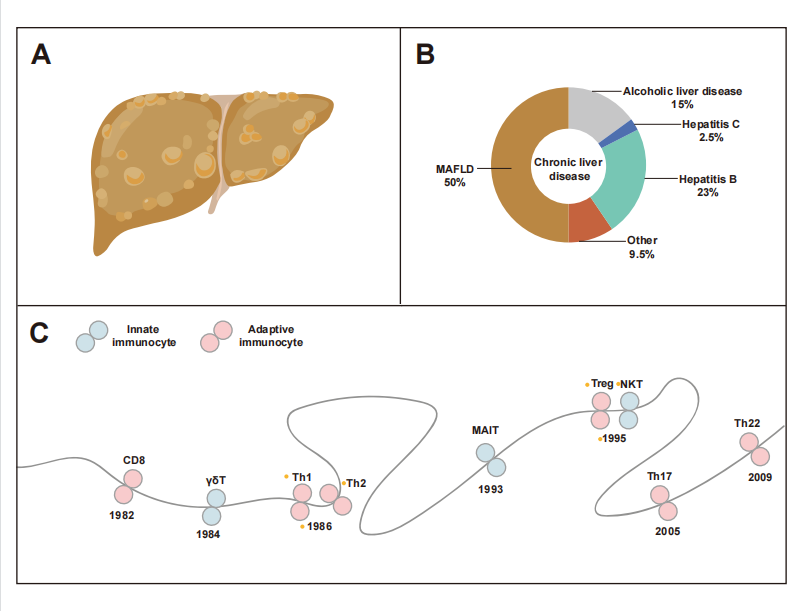
图1. MAFLD和免疫细胞的背景总结
(A)MAFLD的肝脏。(B)常见肝病的流行比例。(C)T细胞发育的时间线。
不同T细胞的生理特性和功能
CD4+T细胞,即T辅助细胞(Th),识别与II类MHC分子结合的多肽,分化为效应细胞的亚群,刺激不同的细胞因子并执行不同的功能。在微生物抗原和/或细胞因子IL-12和IFN-γ的刺激下,CD4+T细胞受到刺激并开始分化为Th1细胞,Th1细胞作为辅助T细胞子集,也能够分泌IL-2、TNF-α和IFN-γ等细胞因子。IFN-γ作为Th1细胞的标志性细胞因子,不仅影响Th1细胞的发育和功能,当真菌、细菌(特别是胞内细菌)和病毒等病原体入侵时,还会导致Th1细胞亚群极化。Th1细胞具有抗纤维化的作用,通过分泌大量的IFN-γ来减少成纤维细胞中的胶原合成。另一方面,Th1细胞和IFN-γ也可能诱导肌成纤维细胞中转化生长因子β(TGF-β)的表达,这提示Th1细胞的免疫调节作用是复杂的,需要进一步研究。Th2细胞的分化受转录因子GATA3或IL-4的控制,这些转录因子主要通过调控转录激活因子5(STAT5)和STAT6,释放多种细胞因子,如IL-4、IL-5和IL-13。值得注意的是,Th2细胞释放的细胞因子如IL-4和IL13可以干扰Th1细胞介导的免疫反应,并替代经典的巨噬细胞激活来对抗细胞内感染。Th2细胞通过抑制Th1相关的IFN-γ表达水平来诱导肝纤维化。此外,Th2细胞分泌的IL-13也可通过激活肝星状细胞(HSC)引起肝纤维化。这些证据指出Th1细胞和Th2细胞与多种疾病过程及其平衡直接相关并影响各种疾病的进展。Th22细胞也属于辅助性T细胞的亚群,受IL-6、TNF-α的调节,特异性分泌IL-22,抑制TGFβ。在肝脏中,Th22细胞通过表达成纤维细胞生长因子(FGF),参与了伤口愈合、组织修复和细胞再生的过程。有趣的是,大量的观察表明,IL-22可以被Th22细胞甚至由其他免疫细胞释放,包括Th1/Th17细胞、CD8+T细胞、NKT细胞以及一些非淋巴细胞。除了上述Th1和Th2细胞的细胞相互调节外,Th17细胞和调节性T(Treg)细胞是分别来源于T淋巴细胞和幼稚T细胞的另一对重要的免疫细胞。Th17细胞作为促炎细胞,可以在关键转录因子维甲酸受体相关孤儿受体γt (RORγt)和STAT3的激活下开始分化,并进一步促进IL-17、IL-22、IL-23和的分泌。在慢性乙型肝炎(CHB)和乙型肝炎病毒(HBV)相关的肝硬化患者中,Th17细胞频率的异常增加可能与TGFβ1/IL-17有关。然而,Treg细胞不同于其他CD4+阳性表型的Th细胞,其主要通过激活叉头盒P3(Foxp3)和产生抑制性细胞因子IL10和TGFβ来发挥免疫控制作用以维持免疫系统平衡。此外,Treg细胞通过多种机制抑制免疫反应和调节自我耐受,但一旦缺乏FOXP3基因,天然Treg细胞的功能障碍会导致宿主自我不耐受,并引发多种免疫失调连锁综合征。此外,研究报道Treg消耗导致CCR2highLy-6Chigh单核/巨噬细胞和前纤维化的Th2细胞高度活化与聚集从而加重肝脏炎症。与对照组相比,高数量的Treg可以对抗自我耐受性、自身免疫性和病毒相关性肝病。此外,Treg细胞参与了1型糖尿病、MAFLD、过敏性疾病等多种炎症性疾病的进展,但这些疾病的发病机制仍有待进一步探索。除了CD4+T细胞之外,CD8+T细胞通过MHC-I和CD4+T或CD8+T细胞衍生的IL-2的共同刺激下,增殖分化为细胞毒性T淋巴细胞(CTL)和CD8+T记忆细胞。CD8+T细胞分泌IFN-γ,激活经典的巨噬细胞免疫反应以杀死细胞外微生物。一方面,CD8+T细胞具有通过释放颗粒酶和穿孔素来启动受感染细胞凋亡的功能;另一方面,它们也与CD4+T细胞合作识别感染细胞或癌细胞的I类MHC相关肽对抗病变。Lan Baudi等报道了特定的CD8+T细胞应答在慢性乙型肝炎小鼠模型中起着清除病毒的核心作用,但保护作用由于慢性抗原刺激或共刺激或细胞因子信号通路被抑制而丧失。
此外,还有一些与CD4/CD8+T细胞不同的细胞,如γδT细胞、NKT细胞和MAIT细胞它们识别非蛋白抗原但不涉及I类或II类MHC分子。这些细胞对感染的反应方式都是适应性免疫的特征,但也具有先天免疫的特征,如快速反应和抗原识别的多样性。γδ T细胞具有T细胞的先天免疫功能,包括分泌细胞因子和清除被感染的细胞。γδ T细胞上的TCR能够识别各种类型的蛋白质和非蛋白抗原,并帮助γδT细胞在抗原特异性T细胞激活之前启动免疫应答来识别微生物。有趣的是,虽然之前的研究主要关注于γδ T细胞在皮肤和肠道中的功能,但γδ T细胞也可以在肝脏中产生IL-17并参与肝脏疾病的进展。其次,NKT细胞表达的αβTCRs具有有限的多样性,并平衡先天性和适应性免疫反应之间的联系。他们不仅可以通过产生许多炎症细胞因子IFN-γ、IL-4/13/17去与其他免疫细胞交互,而且可以抗原识别后通过快速分泌效应分子包括穿孔素和Fas-配体去直接杀死感染细胞,最终参与并影响心血管疾病,自身免疫性疾病和肝硬化的进程。不仅如此,NKT细胞还可以通过快速分泌促炎细胞因子和趋化因子来诱导炎症细胞的聚集,从而加重酒精性肝病、MAFLD和肝细胞癌的肝损伤。此外,MAIT细胞以CD4- CD8-(双阴性)T细胞或CD161+和IL-18Rα+ CD8αβ T细胞为特征,主要归巢于肠道和肝脏,其是遗传上保守的T细胞亚群。大多数MAIT细胞可以表达高水平的多药耐药转运体 (ABCB1)和αβTCR,并产生IFN-γ、颗粒酶-B和IL-17。此外,MAIT细胞通过IFN-γ和TNF-α的大量表达和释放,被快速激活和高度富集以调节肝脏中宿主的防御和炎症。值得注意的是,MAIT细胞促进了肝脏组织的修复,而它的缺失会引发或加重损伤,这表明MAIT细胞是改善肝脏疾病的新兴治疗靶点。
总的来说,越来越多的证据表明,Th细胞亚群,Treg细胞,CD8+T细胞和先天T细胞参与了炎症和脂肪变性的发病机制。T细胞是多种疾病中免疫调节的有吸引力的治疗靶点,包括MAFLD、自身免疫性肝炎(AIH)、肝缺血-再灌注损伤(HIRI)。值得注意的是,明确MAFLD的异质性和发病机制,寻找新的治疗靶点是非常迫切,但免疫与MAFLD之间的关系却被忽视。考虑到药物缺陷等瓶颈问题和免疫调节的重要性,我们全面总结了关于T细胞在MAFLD的关系和矛盾的作用,试图为MAFLD和相关并发症的治疗提供一个新的治疗方向。
T细胞和MAFLD
Th1细胞对MAFLD的影响
作为经典的促炎细胞,Th1细胞要么装备吞噬细胞介导的宿主防御,通过诱导共刺激分子CD40L-CD40的相互作用,或释放IFN-γ,或调节细胞免疫特异性稳态,以摄取或对抗巨噬细胞中存活的病原体。鉴于这些作用,Th1细胞与巨噬细胞介导的炎症反应与MAFLD的发病机制密切相关。在MAFLD肥胖患者的肝脏中,肝脏免疫信号素升高刺激Th1细胞的激活和分化,并且进一步诱导Th1细胞和巨噬细胞分泌促炎细胞因子,引发无组织学异常的低度炎症。Sutti等人进一步研究了Th1细胞在MAFLD病程发展中的作用及潜在机制。他们首先证明了蛋氨酸胆碱缺乏(MCD)饮食促进了小叶炎症和MAFLD损伤,同时伴随着对丙二醛(MDA)衍生抗原的免疫球蛋白G(IgG)的增加。在MCD喂养前用MDA-BSA免疫后,由于Th1细胞的招募和激活,以及随后的M1巨噬细胞,小鼠遭受了更严重的脂肪性肝炎。此外,高脂饮食(HFD)也导致了T细胞数量的升高,特别是Th1细胞的增加。单核细胞趋化蛋白1(MCP1)可触发或升高炎症因子/细胞的表达,随后参与了脂肪变性的过程。值得注意的是,IFN-γ缺乏降低了HFD诱导的炎症细胞和胆固醇瘦素水平的积累,并通过降低小鼠脂肪组织中炎症因子TNF-α和MCP1的表达,提高了胰岛素敏感性。另一项研究发现,MAFLD和多种炎症反应可以通过抑制Th1细胞体,或其关键的细胞因子相关受体,如CXCR3来得到改善。张等人进一步证明了该受体的重要性,并发现CXCR3的缺失通过减弱Th1细胞诱导的免疫反应,减少细胞因子IFN-γ的分泌,抑制NF-κB的激活来改善MCD诱导的小鼠肝脏的脂肪积累、脂肪性肝炎和MAFLD。
除了单独增加的Th1细胞外,越来越多的证据也发现Th1细胞和其他多种免疫细胞可以协同调节炎症反应和MAFLD的进展。肿瘤坏死因子受体OX40(CD134)作为T细胞共刺激分子在活化的T细胞上表达。在HFD喂养的刺激下,敲除OX40通过抑制Th1细胞的分化和增殖,抑制单核细胞迁移和M1巨噬细胞极化,限制促炎细胞因子的表达,如IFN-γ和TNF-α,去显著抑制了脂肪积累、小叶炎症和局灶性坏死。更有趣的是,在MAFLD的早期阶段,B2淋巴细胞通过与激活的Th1细胞相互作用,加速了脂肪性肝炎的进展。在脂多糖(LPS)和antiCD40/IgM的刺激下,肝内B(IHB)细胞通过诱导IL-6、TNF-α和IgG2a的分泌,增强HFD诱导的MAFLD模型中Th1细胞的分化和聚集,加重了肝炎和MAFLD。因此,T细胞与B细胞或其他非实质细胞之间的相互作用复杂地影响着MAFLD的进程和发展。
此外,虽然MAFLD和HCC是完全不同的表型疾病,但代谢相关的脂肪性肝炎已成为肝恶性肿瘤迅速增加的病因。这一观点得到了临床观察的支持,表明随着时间的推移,西方饮食诱导的小鼠依次发生MAFLD、进行性纤维化和自发性HCC。在这一过程中,Th1的炎症模式占主导地位,而不是依赖于Th17细胞的炎症模式去造成全身炎症,同时促进HCC的发展。因此,Th1细胞在MAFLD的整个过程以及后期的恶性肝病中起着重要的作用。
Th2细胞对MAFLD的影响
Th2细胞通过分泌IL-4、IL-5和IL-13,可有效改善内脏和皮下脂肪组织的炎症水平。肥胖Alms1突变体(foz/foz)C57BL6/J小鼠在喂食HFD后,发生严重MAFLD和肝纤维化,并伴随着IL-4和IFN-γ比例增加,Th2占据主导,血小板源性生长因子(PDGF)-α和结缔组织生长因子(CTGF)表达上调。后来,研究人员继续研究这一现象,并归因于高度极化的Th2免疫反应。积聚的Th2细胞可以分泌大量的IL-4和IL13,促进巨噬细胞释放生长因子如PDGF-α和CTGF,并直接激活成纤维细胞,最终引发损伤肝组织的纤维化。除了经典的th2相关细胞因子外,值得注意的是,IL-33是另一种新兴的免疫调节剂,在慢性炎症疾病中释放信号,提示免疫细胞存在细胞或组织损伤。MAFLD患者和HFD/MCD-fed小鼠表明,IL-33高表达并显著激活Th2细胞和肝脏M2巨噬细胞,招募Th2细胞因子像IL-4,IL-5和IL-13并下调IFN-γ,最终以剂量依赖的方式去改善MAFLD症状。然而,高表达的IL-33通过响应致瘤性2受体(ST2)/IL-13的纤维化信号导致HFD喂养小鼠慢性组织损伤甚至增强肝纤维化,这可以通过敲除半乳糖凝集素-3(Gal-3)来改善。总的来说,免疫微环境和MAFLD之间的联系需要进行一个综合的研究。
Th1细胞-Th2细胞相互作用对MAFLD的影响
考虑到Th1细胞和Th2细胞对免疫应答调节的拮抗作用,越来越多的研究人员试图通过调节Th1细胞-Th2细胞平衡作为突破点来研究MAFLD的发病机制(图2)。临床资料显示,Th1细胞-Th2细胞稳态失调引起肥胖MAFLD患者的外周血和肝静脉(HV)出现严重的炎症症状。在轻度、中度、重度MAFLD患者的肝脏中,Th1全身促炎状态增强,同时刺激抗炎Th2通路来平衡Th1细胞的上调。在大鼠中,HFD显著提高肝脏中TG、TC、低密度脂蛋白(LDL),丙二醛(MDA)的水平和Th1细胞的数量并降低高密度脂蛋白(HDL)的水平、烟酰胺腺嘌呤二核苷酸(NAD)和Th2细胞的数量,这表明Th1细胞和Th2细胞之间存在相互制约关系。Koumine是一种中药提取物可以逆转上述情况,也通过抑制Th1细胞的分化和细胞因子的分泌IFN-γ,TNF-α,并增加Th2细胞的产生和抗炎细胞因子IL-10来改善脂肪性肝炎。Toll样受体9(TLR9)是一个模式识别分子家族和典型的炎症受体,也有助于MAFLD的炎症浸润和促炎过程的激活。有证据证明,敲除TLR9可抑制包括M1型巨噬细胞和Th1细胞等炎症细胞的募集,打破了Th1细胞和Th2细胞的平衡,通过减少Th1细胞释放的促炎症细胞因子(包括MCP-1和TNF-α)和增加Th2细胞释放的抗炎症细胞因子来促进动脉粥样硬化小鼠的M2极化,从而抑制小叶炎症和MAFLD进展。众所周知,T-bet和GATA-3等转录因子可调控T细胞分化,调控细胞增殖。刀豆蛋白(Con A)刺激后,肝炎的加重主要归因于胆碱缺乏饮食(CD)诱导小鼠中Th1反应和增加Th1细胞释放细胞因子如TNF-α,IL-12和IFN-γ,通过上调Th1细胞相关转录因子的表达,同时抑制Th2细胞相关转录因子GATA-3水平。α-GalCer作为NKT细胞的激活因子,可以被不变的自然杀伤T细胞(iNKT)特异性识别。腹腔注射α-GalCer后可以通过增加GATA-3而降低T-bet的表达来显著上调iNKT2的数量,从而逆转HFD诱导的MAFLD。
通过药物治疗恢复免疫稳态正成为一个研究热点。江等人阐述了TFCH药物治疗不仅显著抑制Th1/Th2细胞相关炎症细胞因子也抑制HFD-diet小鼠中NF-κB/MAPK炎症信号转导,减少肝内炎症并改善MAFLD。随着研究的进一步发展,肠道免疫紊乱已成为导致MAFLD的主要原因之一。益生菌丁酸梭菌B1(CB)是一种产生丁酸盐的益生菌,通过促进Th2细胞的产生和抑制Th1细胞的分化,调节T细胞的增殖和分化,减弱肠肝免疫失调和脂肪性肝炎。另一个证据表明,肠道微生物群与肠道和肠系膜淋巴结(MLNs)中的T细胞的分化和增殖有关,MLNs是肠道相关淋巴组织(GALT)的一个组成部分。有趣的是,在肝脏中,从HFD喂养的小鼠模型中分离出的MLNs中Th1/Th2细胞和Th17/Treg细胞的占比时不平衡的。这些MLN CD4+T淋巴细胞表现出向肝脏迁移的倾向和加速炎症反应和脂质积累的趋势;而应用抗生素(新霉素和多粘菌素B)或益生菌(乳酸菌)通过改善肠道细菌和恢复肠道CD4+T MLNs中的T淋巴细胞平衡显著缓解MAFLD。总的来说,调节Th1-Th2细胞平衡以改善MAFLD的策略可能是通过保证肠道微生物群的稳态。
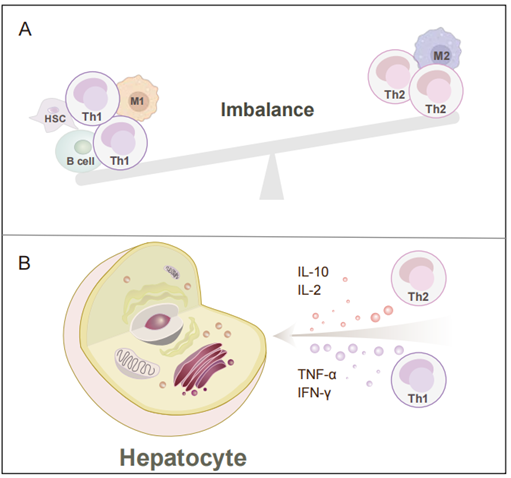
图2. Th1-Th2细胞的稳态在MAFLD的进展中起着决定性的作用
(A)Th1和Th2细胞的不平衡伴随着多细胞的串扰。(B)肝细胞受到MAFLD中Th1和Th2细胞紊乱比例及其关键炎症因子的影响。Th1细胞 上调了炎症细胞因子和受体的水平,加重了肝炎和脂质沉积,甚至参与了HCC的发生过程。此外,Th2细胞通过抗炎细胞因子拮抗Th1细胞的反应,抑制炎症反应。
Th22对MAFLD的影响
Th22细胞,来源于外周血T细胞和CD4+T细胞,可以激活上皮细胞的先天免疫,清除位于体表面的细菌感染,并特异性分泌IL-22。MCD诱导的MAFLD小鼠显示Th1细胞、Th17细胞和Th22的广泛浸润以及通过c-Junn-末端激酶(JNK)诱导引起的肝脏脂肪积累。既往研究报道,磷酸肌醇3-激酶(PI3K)/蛋白激酶B(Akt)可以抑制JNK的激活,但受到10号染色体(PTEN)信号上缺失的磷酸酶和紧张蛋白同源物的负调控。腺苷A2a受体 (A2aR) 是一种重要的免疫脂肪毒性调节剂,它通过调节CD4+Th细胞的应答来识别并结合腺苷去激活免疫抑制环境。同时,A2aR激动剂CGS21680通过降低CXCL10和CCL2来抑制Th22细胞和Th17细胞的募集和极化,通过刺激PI3K/Akt信号来抑制JNK依赖的脂肪毒性,并最终抑制MCD饮食小鼠中MAFLD的发展。随后的研究进一补充了如何通过上调MAFLD中的IL-22和Th22细胞来改善脂肪毒性的机制。在被棕榈酸处理的肝细胞中,IL-22抑制了IL-17/JNK通路相关肝细胞脂肪毒性的增加趋势;但值得注意的是,仅在IL-17-/-小鼠中,TH22细胞降低了JNK活化和PTEN表达,增加了Akt磷酸化,并通过运输IL-22到肝脏来减轻炎症和抑制脂肪毒性。
Th17对MAFLD的影响
不仅是Th22,大量观察表明,Th17细胞也能够分泌IL-22并产生其他细胞因子,如IL17和IL-23,通过招募中性粒细胞和巨噬细胞,促进肝脏炎症,增强适应性免疫反应。值得注意的是,与健康对照组相比,浸润的Th17细胞和上调的Th17细胞相关基因如RORγt、IL-17、IL-21和IL-23在MAFLD患者的肝脏中显著升高。值得注意的是,在MAFLD和脂肪性肝炎的进展中,IL-17轴包括IL-17A、IL-17F和IL-17A受体(IL17RA)积极参与甚至加重了肝脏T细胞和巨噬细胞的浸润。肝脂肪酶(HL)水解TG和磷脂脂蛋白还通过增加循环炎症细胞,包括胰岛细胞、CD4+T细胞和表达IL-17的Th17细胞亚群,以及上调炎症应激p38和JNK信号激酶从而在高脂肪/高胆固醇(HFHC)小鼠中引起肝脏脂肪变性和炎症。此外,也有人指出,氧化应激元素包括烟酰胺腺嘌呤二核苷酸磷酸氧化酶2(NOX2)和活性氧(ROS)与脂肪变性和MAFLD的发病机制有关。此外,敲除NOX2可显著改善肝内炎症环境,并明显降低HFD饮食小鼠的CD4+T细胞水平特别是Th17细胞和促炎细胞因子(IL17A、IFN-γ和TNF-α)。考虑到IL17的重要功能,研究人员也试图通过抑制IL-17来逆转MAFLD。与预期的一样,在HFD和HFD/LPS诱导的小鼠模型中,抗IL17抗体中和IL17通过抑制IL-17/胰岛素信号通路改善肝脏炎症趋势,减少炎性Th17细胞的浸润和脂肪变性。同样,在MCD诱导的MAFLD小鼠中,传统中草药丹参的主要成分隐丹参酮(CTS)抑制Th17增殖和IL17A的产生,特异性抑制含有NOD样受体家族pyrin结构域3(NLRP3)的炎症反应。基于网络药理学的方法,罗等人对中药(TCM)复方附子理中汤(FLD)的活性成分进行预测,发现这些成分可直接调控Th17细胞分化,抑制IL-17信号通路,进一步抑制MAFLD进展。有趣的是,过继细胞转移(ACT)是一种生物治疗方法,它被动地将免疫细胞转移到未免疫的个体中,以发挥治疗效果。CD4+T双阴性细胞通过抑制TNF-α和IL-17的产生,增加M1巨噬细胞和CD4+T细胞包括Th17细胞的凋亡来缓解HFD诱导的MAFLD。
越来越多的研究已经开始探索转录因子、关键受体或其他转录调控因子是否也参与了Th17细胞的分化和招募。OX40作为二级共刺激免疫检查点分子,在HFD诱导的MAFLD小鼠模型中通过上调转录因子RORγt来促进Th17细胞的增殖和分化。同时,OX40-/-抑制了CD4+T细胞向Th17细胞的分化激活,减轻肝脏炎症和MAFLD进展。另一方面,新出现的证据显示,炎症趋化因子CXCR3不仅通过调节炎症性Th1细胞相关的免疫反应,还通过控制Th17细胞的募集和细胞因子IL-17的产生来影响MAFLD的进展。根据最新的发现,炎症性肝脏Th17(ihTh17)细胞是一种特殊的Th17亚群,也能在脂肪变性的肝脏中高表达CXCR3。Moreno-Fernandez, M.E等人指出HFD刺激导致肝ihTh17细胞的积累和各种促炎细胞因子的产生包括IL17A,IFN-γ和TNF-α,从而通过激活CXCR3/CXCL9/CXCL10轴加剧了肝脏脂肪变性,但可以通过糖酵解抑制剂2-脱氧-d-葡萄糖(2DG)或细胞特异性删除基因编码丙酮酸激酶M2(PKM2)等方式改善肝炎。除上述转录因子或受体外,microRNA还可以直接调控相关基因的转录,进而影响IL-17/Th17细胞相关的免疫应答。既往研究表明,mastiha是一种含有活性植物化学物质的天然补充剂,可通过调节甾醇调节元件结合蛋白1c(SREBP1c)通路,调节循环中的microRNA155,抑制肝脏X受体(LXR)/脂肪生成基因的表达,并影响MAFLD患者中Th17细胞的分化。
最近的研究发现,与肥胖相关的肠道调节失调也通过增加肠道菌群产物和激活促炎信号通路导致MAFLD的进展。内毒素是肠道微生物的产物,可显著增加肠道通透性,降低免疫力,并在血清中异常升高,与MAFLD患者中Th17细胞和IL17的升高呈正相关。出乎意料的是,IL-17−/−仍导致MAFLD加重和肠道菌群变异,包括致病性微生物(Staphylococcaceae, Enterococcaceae, and Enterobacteriaceae)的增加和有益微生物的减少。而野生型(WT)小鼠或抗生素(Abx)处理小鼠的粪便微生物群移植(FMT)可以改善肠道通透性,重建肠道屏障,以改善MCD引起MAFLD。将IL-17−/−小鼠来源的CD4+T细胞移植到重组激活1缺失(Rag1−/−)小鼠,加重了受体小鼠肠屏障功能障碍和肝脏脂肪变性,说明CD4+Th17细胞来源的IL-17在MCD饮食喂养的小鼠模型中具有维持肠道屏障的完整性功能。因此,肠道微生物群和MAFLD之间的联系表明,肠道菌种失调可能是导致MAFLD的一个危险因素。与一般人群相比,HIV阳性个体对MAFLD更易感,血浆结果表明IL-17A下调,sCD14和IL-22上调。而sCD14和IL-22可能是肠道损伤和内毒素暴露的标志物,因此推测HIV患者高发MAFLD的一个关键因素可能是细胞免疫和肠道菌群的共同失调。
目前,脂肪组织被认为已被视为一个重要的内分泌系统,它影响胰岛素敏感性并导致炎症和脂肪变性,而不仅仅只是一个能量储存器官。临床研究表明,胰岛素抵抗性肥胖患者表现为脂肪组织中Th17细胞和Th22细胞的显著增殖,并且由于循环中的IL-22和IL-6以及脂肪组织释放的细胞因子的增加而导致IL-17和IL22受体在肝脏中的高表达。体外实验表明IL-17和IL-22通过抑制Akt磷酸化,抑制大鼠比目鱼肌葡萄糖摄取,降低人肝细胞胰岛素敏感性,进而导致肝脏和肌肉代谢功能障碍。高脂高果糖饮食(HFHFD)喂养的小鼠的内脏脂肪组织上调了CD8+T细胞和Th17细胞的比例,但通过富集细胞毒性T淋巴细胞相关蛋白4(CTLA4)信号通路和增加RORγt的表达来下调Treg细胞的数量。抗CD8a的抗体通过抑制CD8+T细胞在内脏脂肪组织和肝脏中的增殖来减轻脂肪变性和纤维化,而不改变Th1细胞、Th17细胞或Treg细胞的水平。另一方面,与抗cd8a抗体治疗不同,抗IL-17a抗体只改善了肝内炎症,而没有逆转MAFLD的其他症状。最近的一项研究表明,PsTag600-FGF21(重组多肽PsTag和生长因子FGF21)药物治疗可以减弱胆碱缺乏高脂饮食(CD-HFD)诱导的脂肪积累、炎症巨噬细胞浸润以及通过上调脂联素的产生而间接抑制Th17细胞的增殖和IL-17A表达。
事实上,MAFLD并不是简单的慢性肝脏疾病,它会进一步恶化为肝纤维化,这是发展为肝硬化和HCC的关键步骤。MAFLD及其相关的恶性肝病是由特殊情况驱动的,包括但不限于遗传/表观遗传学的变化、免疫细胞或信号通路的改变。其中,表观遗传学是指通过非基因序列的改变来改变基因表达水平,如DNA甲基化或组蛋白修饰,这一过程需要DNA甲基转移酶(DNMT)和组蛋白去乙酰化酶(HDAC)的参与。有研究证实,MAFLD和肝硬化患者在肝中出现Th17细胞数量增加,HDAC2和DNMT1的异常激活,血管基因IGFBP7和ADAMTS1(标记物不适应上皮细胞)的特异性表达。同时,西方饮食和四氯化碳诱导也会导致Th17细胞脂质积累和迁移募集,HDAC2抑制剂(HDAC2i)和DNMT1抑制剂(DNMT1i)联合治疗后,这种情况得到缓解。正如预期的那样,HDAC2iΔEC经DNMT1i治疗后,小鼠肝脏Th17数量和ADAMTS1表达下降;同样,IGFBP7−/−小鼠由于IGFBP7的缺失,也显示出Th17炎症反应和肝纤维化反应的减少。这一结果表明,HDAC2/DNMT1和IGFBP7/ADAMTS1轴共同影响Th17细胞的增殖和分化,使MAFLD恶化。在HFD、CD-HFD和MCD-HFD营养过剩的刺激下,肝脏非常规前折叠素RPB5相互作用子(URI)增加,但杂合子(URI(+/Δ)hep)小鼠即使有饮食干预,仍然显示有炎症细胞浸润和MAFLD下降。敲除人类URI的小鼠(即突变体),作为多步HCC依赖的基因毒性应激的自发性肝癌模型,通过激活URI依赖的IL-23/IL17A轴可以增加WAT中的中性粒细胞和Th17细胞。同时,地高辛或IL-17A抗体降低了中性粒细胞和Th17细胞的水平,阻断了IL-17A的信号传导,并通过抑制URI来抑制发育不良灶(HCC前兆),以改善伴有MAFLD的HCC。
Treg对MAFLD的影响
在胸腺或外周组织中发育的Treg细胞,抑制过度的效应T细胞反应和随后的组织炎症损伤。免疫细胞比例的紊乱,特别是Treg细胞,参与了饮食诱导的糖脂代谢失调和MAFLD的发展。Lian等人表明口服抗TNF融合蛋白(PRX106)可以恢复免疫细胞的分布,比如肝脏CD4+CD25+FoxP3+Treg的减少,肝脏CD8+CD25+FoxP3+treg和CD3+NKT细胞的增加,从而逆转了HFD诱导的肝炎。哺乳动物雷帕霉素复合物1(mTORC1)是调节SREBP1c的靶点,SREBP1c是脂质生物合成的关键元素,调节Treg细胞分化。随后,研究者深入探讨了Treg在HFD诱导的MAFLD小鼠模型中的作用,发现Foxp3的抑制导致了CD4+Foxp3+Treg细胞和CD8+CD122+T细胞的下调和并通过上调SREBP-1c途径相关蛋白而导致过度脂质生物合成。值得注意的是,从苹果树和其他植物中提取的phloridzin,可以通过抑制HFD小鼠和aP2-SREBF1c转基因小鼠的mTORC1/SREBP1c通路,恢复Treg细胞的功能,改善脂质代谢异常。
随着研究技术的发展,越来越多的基因敲除动物(基因缺失)被用于验证特定基因与MAFLD发病机制之间的关系。共刺激分子B7.1和B7.2主要在APCs上表达,并通过与T细胞上的CD28相互作用激活Treg。在HFD的刺激下,B7.1/B7.2双缺陷小鼠出现因Treg细胞数量减少和M1型巨噬细胞数量增加所导致的肝脏炎症、脂肪变性和代谢失调。有趣的是,过继转移Treg细胞并没有改善缺失B7小鼠的MAFLD。TLR7作为一种内源性受体,可以激活IFN和TNF-α信号通路,从而抑制Treg细胞的活性。与野生型小鼠相比,TLR7或IFN-α/β受体1敲除小鼠对肝细胞死亡和脂肪性肝炎的耐受性更强,TLR7拮抗剂也逆转了Treg细胞的凋亡,改善了MAFLD症状。值得注意的是,Rag1−/−小鼠被认为是包括T/B细胞在内的消融性适应性免疫反应的模型。Rag1−/−高脂高碳水化合物(HFHC)饮食喂养的小鼠表现出更严重的炎症和免疫微环境紊乱,这表现在M1巨噬细胞和NK细胞显著上调,抗炎的M2巨噬细胞显著下调。Janine Dywicki等人进一步表明,尽管过继转移Tregs治疗可以增加Treg细胞的数量,但同时加剧了炎症,甚至导致喂食HFHC的Rag1−/−小鼠的结肠炎。同样,另一项研究也表明,虽然过继转移Tregs增加了皮下脂肪组织中的Treg细胞,减少了肝脏中的Th1细胞,但它在HFHFD诱导的小鼠中引起了更多的免疫破坏并加剧了肝脏脂肪变性。这些发现表明,个体中Treg的数量和功能可能决定其在MAFLD治疗方面的作用。
除上述因素外,衰老也是影响MAFLD发展的关键因素之一。据报道,衰老小鼠表现出脾内Treg细胞、I型监管(Tr1)样细胞和IL-10的显著增加以及炎症介质(CCL2,CCL4IL -6和TNF)水平的减少,从而缓解HFD喂养引起的肥胖相关的代谢紊乱。众所周知,衰老也与细胞癌变密切相关。值得注意的是,Treg紊乱会导致MAFLD向HCC的发展(MAFLD-HCC)。临床资料显示,中度MAFLD-HCC患者具有较高水平的Treg细胞,可抑制宿主的抗肿瘤免疫,并显著促进癌症的发生和进展。丁酸盐是MAFLD-HCC患者肠道微生物群的代谢产物,使Treg细胞扩增,同时通过增强Foxp3的表达和抑制促炎症细胞因子来减少CD8+T细胞和B细胞的数量,以加强非MAFLD个体的外周血单核细胞(PBMC)的免疫抑制反应。另一方面,在MAFLD到HCC的过程中,中性粒细胞胞外陷阱(NETs)相关的代谢重编程通过上调Treg分化和功能相关基因来分化影响了CD4+T细胞分化。令人惊讶的是,消耗Treg显著抑制了MAFLD-HCC的发生和发展。同样,Treg细胞在MAFLD的进展中似乎发挥着比我们想象的更双向和复杂的作用。
Th17-Treg相互作用对MAFLD的影响
Th17细胞和Treg细胞来自相同的前体细胞:前者通常被认为能促进自身免疫和组织损伤,而后者则能拮抗过度免疫反应和肝损伤。Victoria Cairoli等人阐明了Th17细胞和Treg细胞的比例增加会加重炎症,甚至促进中度和重度MAFLD成人的肝脏纤维化。有趣的是,在减肥手术后,MAFLD患者的Th17/rTreg细胞的比例显著改变,这可以从Th17细胞和IL17细胞的产量减少以及外周血中rTreg细胞的产生增加中发现。由于这些临床数据直接指出,Th17/Treg细胞的失衡与MAFLD疾病的发展密切相关,各种MAFLD动物模型被构建以彻底研究Th17/Treg细胞失衡对MAFLD发病机制和治疗药物开发的影响。亚硝酸钠诱导的慢性间歇性缺氧(CIH)触发了缺氧诱导因子1 alpha(HIF1 α)的积累和mTOR信号通路的激活,导致HFD-饮食小鼠中Treg/Th17细胞比例降低,MAFLD加重。与之前关于Th17细胞的研究一致,microRNAs也通过靶向其下游基因参与Treg细胞的稳态。CD40作为T细胞的表面抗原,是miR195的下游调控元件。在HFD诱导的MAFLD大鼠中,miR195过表达和沉默CD40分别上调Treg细胞的表达和下调Th17细胞的增殖维持Th17/Treg细胞平衡以及抑制促炎因子的表达,随后改善炎症和MAFLD的发展。FOS基因作为microRNA-29c的靶标,是一种基因抑制因子,与IL17信号通路的调控密切相关。MCD诱导的MAFLD小鼠显示出FOS的高表达引起Th17细胞增加和保护性Treg细胞减少。相反,过表达microRNA-29c可以通过抑制FOS/IL17信号通路,逆转TH17/Treg细胞的比例,改善肝脏脂肪沉积。不仅局限于microRNA、多肽和炎症因子相关受体也与MAFLD的进展有关。肌肽和α-硫辛酸(ALA)发挥了减少氧化损伤和改善脂质代谢的作用,并通过抑制高热量胆碱缺乏饮食(HCCDD)雄性大鼠模型的细胞凋亡来保护肝细胞,这归因于Treg/Th17细胞和IL-10/IL-17A比率的升高。李等人进一步探索了CXCR3在T细胞上的作用,并发现IFN-γ(MIG)/CXCL9诱导的单因子通过以剂量依赖的方式促进CXCR3在Th17细胞上的表达,并进一步加剧了MCD-饮食小鼠的MAFLD,这一点通过沉默MIG/CXCL9得到了逆转。
在确定了不同的调控机制后,许多研究已经开始发现针对上述相关环节的特定药物。有趣的是,A2aR激动剂的治疗通过抑制棕榈酸(PA)培养的肝细胞中负向调节作用的PTEN表达,改善了Th17/Treg细胞的比例异常和脂肪毒性。同样,用多烯磷脂酰胆碱胶囊(PPC)治疗能够调整Th17/ Treg细胞的失衡,增加FoxP3的mRNA水平、减少Th17细胞相关细胞因子(IL-6,IL17和IL-23)和肝脏RORγt和STAT3的mRNA水平,改善HFD诱导MAFLD小鼠模型的肝脏炎症和纤维化。越来越多的证据表明,中药及其提取物具有改善免疫细胞失衡的多组分和多靶点特性,在临床和基础研究中被广泛应用于MAFLD的治疗。研究进一步研究表明,伤寒论中的小柴胡汤通过影响AKT1、IL-6、JUN、MAPK8和STAT3等多种信号通路,有效介导IL-17功能,改善MAFLD。此外,刘云等人证实十字花科蔬菜中3,3’-二烷基甲烷(DIM)通过激活芳基烃受体(AhR)信号,改变Treg/Th17细胞的失衡,改善肝内淋巴细胞浸润,这个过程会被AhR拮抗剂CH223191减弱,而被抗TLR4中和抗体增强。另一项研究报道,HFD显著增加了促炎脂肪因子如趋化素、趋化素样受体1(CMKLR1)和促炎C-C趋化因子受体2(CCR2)的表达,以调节大鼠Treg/Th17细胞的肝内比例。同时,中药黄连的主要成分小檗碱(BBR)通过修复Treg/Th17比值的平衡,下调趋化素/CMKLR1通路,显著改善了大鼠游离脂肪酸(FFA)的积累、脂质沉积和脂肪性肝炎。同样,Koumine可以通过减少Th17细胞及其细胞因子的分泌,增加Treg细胞和抗炎细胞因子IL-10的水平,抑制炎症细胞浸润,改善HFD诱导的MAFLD大鼠肝细胞的脂肪样病变。同样,中草药配方也是临床预防MAFLD的非常有效的手段。据报道,大黄蛰虫丸(DHZCPs)作为中药经典复方,维持Th17/Treg细胞平衡,调节炎症细胞因子IL17、IL-10和TNF-α水平,改善MAFLD患者的肝功能。肠道微生物群紊乱也是Th17和Treg细胞相关代谢综合征的关键因素。肠道微生物群作为一种MAFLD调节剂,为MAFLD的治疗开辟了一个新的方向。肠道益生菌CB通过显著促进HFD诱导的小鼠模型的肝脏和回肠中CD4+T细胞向Treg分化,增加Foxp3、IL-4和IL-22的水平抑制肠道肝免疫紊乱,并降低CD4+T细胞向Th17分化以及IFN-γ和IL17的表达,保护肝脏免受损害。因此,修复Th17/Treg细胞的失衡,而不是简单地增加这些免疫T细胞的数量,是MAFLD的一个潜在治疗方向 (图3)。
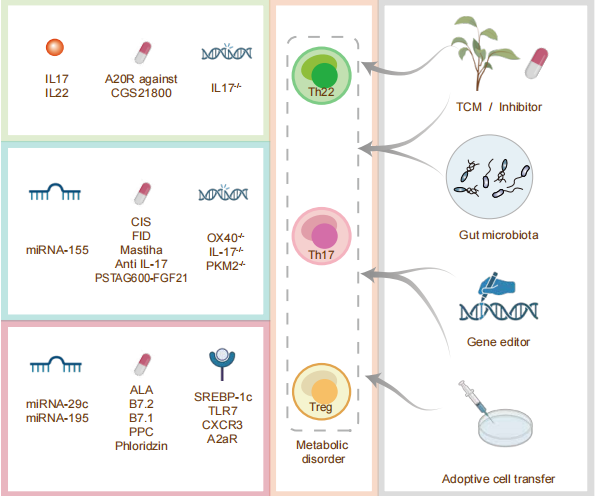
图3. 改变Th22-Th17-Treg细胞的平衡是导致或改善MAFLD病情恶化的关键途径之一
Th22-Th17-Treg细胞稳态的失衡加剧了炎症和纤维化,甚至促炎相关因素导致的肠道细菌失调,但通过microRNA、改变菌群、基因编辑或过继细胞转移 (ACT)以及药物治疗可以逆转MAFLD,其中特别要强调得是中药、益生菌的作用。
CD8+T细胞对MAFLD的影响
最近有研究发现,基于T淋巴细胞簇分化的CD8+T细胞与CD4+T细胞来源的Th细胞相似,表现为在MAFLD患者的肝脏中浸润,同时伴有更高程度的脂肪变性。另一项临床研究显示,与没有代谢综合征的儿童相比,肥胖儿童血清中IL-7RαlowCD8+T cells和IL-7RαlowCX3CR1+CD8+T cells细胞水平较高,但CD4+T细胞水平没有明显改变。此外,magar ghazarian等人报道了在HFD诱导的小鼠模型中,这些CD8+T细胞亚群通过IFN-I反应进行积累并激活,促进胰岛素抵抗并引起代谢紊乱。IFNαR1抑制剂或IFNαR1缺乏均可改善了这些小鼠的代谢参数。除HFD模型外,在铁补充饲料诱导的MAFLD小鼠外周血中CD4+T细胞和CD8+T细胞均有所增加。此外,抗原肽转运体1(TAP1)可以将抗原转运到I-MHC,并促进CD8+T细胞的生成。虽然伴随着肝脏CD4+T细胞的增加,TAP1-/-仍然表现出改善HFD饮食的小鼠 (无CD8+T细胞)肝脏脂肪变性的作用。综上所述,上述一些病例显示MAFLD的发病率可能与CD8+T细胞的激活密切相关(图4)。因此,许多研究者已经开始探索通过抑制CD8+T细胞激活来治疗MAFLD的药物。Lunasin (一种抗炎肽) 和cardioaid (一种植物甾醇) 都能有效地降低经HFD处理的小鼠血清TG水平,并通过抑制CD4+/CD8+T细胞来调节免疫稳态。此外,虾青素类胡萝卜素可通过降低肝细胞内TG含量,抑制肝纤维化相关基因的表达,并抑制肝脏CD4+T细胞和CD8+T细胞聚集和枯否细胞的浸润,有效改善在HFD诱导的MAFLD小鼠中的纤维化。程序性细胞死亡1(PD1)及其配体PD-L1是自侵袭性组织损伤的关键检查点。研究发现,PD1/PD-L1轴升高抑制了CD8+T细胞活性,并减轻了CD8+T细胞介导的免疫损伤。Wabitsch等人还发现,与二甲双胍联合使用可通过损害CD8+T细胞的运动和代谢功能来改善抗PD1对HCC脂肪性肝炎的治疗效果。
除了药物干预外,研究人员想知道CD8+T细胞上表达的关键受体是否可以通过抑制其功能在MAFLD中从而发挥积极作用。在单纯性脂肪变性患者中,TLR9表达的降低可以通过增加低水平的CD8+T细胞、IFN-γ的分泌和枯否细胞的活化来预防MAFLD和肝损伤。相比之下,脂肪性肝炎患者不能自动启动TRL9相关的保护来调节肝细胞损伤。值得注意的是,趋化因子受体CCR2和白细胞粘附分子CD44在MAFLD患者中均增加,并在脂质介导的肝炎中起作用。在喂食致脂性饲料后,CCR2-/-小鼠完全可以预防肝炎,但CD44-/-老鼠仅部分恢复。在没有CCR2的情况下,CD44介导的透明质酸 (HA)没有增加对CD8+T细胞的结合。其他研究人员也注意到通过CXCL12-CXCR4轴内皮细胞和CD8+T细胞之间的免疫串扰。CXCR4拮抗剂或CXCL12中和抗体都可以逆转CXCL10、CXCL12和人白细胞抗原(HLA)I的高表达,减少CD8+T细胞的跨内皮迁移,进而减轻MAFLD患者的内皮损伤。出乎意料的是,CD8+T细胞似乎有抗炎或抗纤维化的作用,而不仅仅是加重炎症。推测是CD4+T细胞和CD8+T细胞的免疫缺陷有助于触发MAFLD。口服含有麦角甾醇(维生素D2的前体) 的提取物可增加肝脏CD4+、CD8+T细胞的水平,并降低促炎细胞因子和循环脂质含量水平,改善HFD引起的肝细胞球囊化。最近,Breuer等人报道了CD8+T细胞在中度MAFLD患者和体外肥胖模型中可以通过分泌IL-10抑制肝脏炎症和星状细胞激活高脂血症诱导的脂肪性肝炎。其他研究还表明,在HFD小鼠模型中,IL-15激活组织驻留记忆CD8+T细胞(Trm)能够改善脂肪性肝炎,以CCR5依赖的方式吸引HSCs,通过诱导FasL-Fas通路诱导HSCs的凋亡。
此外,在许多情况下CD8+T细胞也通过影响其他免疫细胞来调节MAFLD的进展,而不是单独发挥作用。在临床病例中,循环中的中性粒细胞或多形核中性粒细胞 (PMNs) 与肝脂肪变性和肝细胞球囊化呈正相关。有趣的是,在中重度脂肪性肝炎患者的PMNs和PBMCs细胞共培养中,CD4+T和CD8+T细胞的激活和增殖被显著抑制。饮食诱导的肥胖和溴二氯甲烷 (BDCM) 的使用引起了代谢性氧化应激,通过cyp2E1-瘦素轴增加CD3+CD8+T细胞中CD57的表达来加速小鼠脂肪性肝炎,进一步介导肝中促炎细胞因子的释放,如IL1β、IFN-γ和IL-2。此外,脂肪细胞来源的瘦素作为免疫系统的关键元素,促进了HFD诱导的MAFLD小鼠肝脏中CD8+T细胞释放IFN-γ和颗粒酶B,从而促进肝细胞或巨噬细胞焦亡。已经发现巨噬细胞和CD8+T细胞也能协同影响MAFLD的免疫微环境。在早期脂肪性肝炎中,主要发生巨噬细胞的浸润,并伴随着TNF和IL-1β表达的增加,而在进展性脂肪性肝炎中,患者门静脉中CD8+T细胞的炎症浸润明显增强,并伴有IL-6和IL-8水平的升高。LuPing等人观察到LepΔI14/ΔI14自发性大鼠特异性脂肪性肝炎模型中巨噬细胞和CD8+T细胞的浸润增加以及M2巨噬细胞向M1巨噬细胞的表型转化。有趣的是,穿孔素的转移可以调节CD8+T细胞的激活和存活,抑制肝脏中促炎性M1巨噬细胞向促炎表型转化和CD8+T细胞的浸润,从而改善MCD和HFD诱导的性脂肪性肝炎。同样,番茄红素通过抑制NADPH氧化酶的表达有效改善肝脏氧化应激,并通过减少CD3+和CD8+T细胞的聚集和M1巨噬细胞的水平,减轻肝纤维化和肝脏炎症。另一方面,通过静脉注射黄芩苷处理的M2巨噬细胞,可降低外周血CD4+CD25-T和CD8+CD25-T细胞表达,减少肝T细胞浸润,减轻Ath饮食诱导的代谢紊乱小鼠肝损伤。
值得注意的是,肠道免疫功能受损和肠道通透性增加可能会干扰肠道与肝脏之间的串扰,由于肠道微生物群的改变进而加重MAFLD。临床研究表明,MAFLD患者CD4+T和CD8+T淋巴细胞水平下降,同时伴随肠道菌群失调。在MAFLD相关肠道免疫的研究中,CCL5诱导的MLN细胞迁移促进了CD4+T细胞和CD8+T细胞的激活并降低了肝内记忆CD4+T细胞和CD8+T细胞的比例,加重炎症性肝损伤。肠内CD8+T细胞的激活可以优先进入肝脏,然后影响局部免疫环境。临床试验也表明,益生菌sachet (MCP® BCMC®)保护肝脏免受肠道绒毛CD8+T细胞的减少的影响,改善轻度、中度、重度MAFLD患者的肝脂肪变性。

图4. CD8+T细胞发挥不同的调节作用,影响代谢失调
CD8+T细胞通过与促炎趋化因子受体轴结合或影响其他免疫细胞(巨噬细胞、中性粒细胞)反应迁移到肝脏,促进饮食诱导的MAFLD,但这种炎症损伤可以通过几种抗炎药物治疗改善。然而,在特殊情况下,CD8+T细胞通过恢复肠道微生物群的平衡或与药物联合治疗来抑制纤维化,甚至改善MAFLD。
γδ T细胞对MAFLD的影响
γδT细胞是一种双阴性T细胞,只占T淋巴细胞总数的一小部分,但在免疫系统中具有不同的促炎和抗炎功能。更重要的是,有研究发现,与外周血相比,肝脏中γδT细胞的比例非常丰富,在肝脏免疫中起着至关重要的作用。例如,产生IFN-γ的γδT细胞亚群 (γδT1) 细胞通过增强细胞毒性来消除激活的HSC。此外,最新研究表明,在花生四烯酸刺激下,TCR γδT细胞亚群通过NF-κB信号通路促进IL17A的分泌,加重MAFLD中肝脏的炎症反应。CCR2/5可以将γδT细胞招募到肝脏,并通过加速CD4+T细胞的扩增和分化,导致MCD诱导的小鼠模型中肝脏脂肪变性和白细胞浸润。另一方面,在HFD诱导的小鼠中,肝脏γδT-17细胞通过分泌高水平的IL-17A,并与CD1d呈递的微生物脂质抗原发生反应,加重MAFLD,而采用用Abx治疗或CD1d敲除均能改善这种情况。Zhou等人进一步发现,喂养标准饲料的小鼠FMT通过上调Foxp3、IL-4和IL-22,有效降低HFD喂养小鼠的IL17水平,调节肝内免疫。近年来的研究表明,基于肠-肝轴,中药对MAFLD有显著的治疗作用。中国传统中药(YJT)有效改善了HFHC喂养的小鼠肝脏脂质代谢物和肠道微生物群的组成,改善了MAFLD损伤,这种改善作用与γδT细胞的减少高度相关 。综上所述,γδT细胞可能成为MAFLD的潜在靶点,但仍需进一步探索。
NKT细胞对MAFLD的影响
NKT细胞存在于上皮和淋巴器官中,通过调节星状细胞活化和炎症细胞浸润参与代谢紊乱的免疫调节。既往研究指出,肝NKT细胞分泌hedgehog(Hh)配体,以旁分泌的方式直接诱导骨桥蛋白(OPN)的表达,促进肌成纤维细胞活化。随后,一项口服β-葡萄糖神经酰胺(GC)的双盲临床试验通过降低MAFLD中度患者中的胰岛素抵抗水平、TG水平和肝脏CD4+T细胞和NKT细胞数量来改善脂肪性肝炎。这些研究表明,NKT细胞的调控可能在MAFLD的进展中起决定性作用(图5)。此外,多项研究普遍显示,NKT细胞在MAFLD中的功能可能与不同的细胞因子和趋化因子受体有关。促炎细胞因子LIGHT (TNFSF14) 通过增加维持NKT细胞功能的关键转录因子Zinc finger and BTB domain containing 16 (ZBTB16)的表达,加重了HFHC饮食的小鼠肝脏炎症。促炎趋化因子受体(CXCR16)也促进了肝脏NKT细胞的积累,使巨噬细胞浸润和炎症因子(TNF和MCP1)增加,导致肝脂肪变性的恶化。此外,我们发现CXCR6能促进iNKT在肝脏中的迁移,并促进iNKT细胞释放促炎因子IFN-γ和IL-4,从而进一步导致MCD饮食喂养的小鼠肝脏中巨噬细胞浸润和肝脏炎症。
如今,基因敲除动物模型已经逐渐成为研究不同免疫细胞功能的常用方法之一,NKT细胞也不例外。有趣的是,CXCR6的缺失改善了MAFLD,并通过减少HFD处理的小鼠中肝脏NKT细胞的募集和炎症细胞因子IFNγ的产生而使脂肪肝免受缺血-再灌注损伤(IRI)。同样,PTP1B的缺失也显著改善了MCD诱导的MAFLD,同时减少了NKT细胞的数量,增加了M2型巨噬细胞的数量。另一方面,敲除典型的炎症因子,如NF-κB1,也会通过早期过量产生IL-15而触发NKT细胞的募集,并增加MCD喂养的小鼠中IFN-γ和OPN的水平。出乎意料的是,虽然IL15对NKT细胞的产生和维持很重要,但肝细胞和巨噬细胞中IL-15的特异性缺失并没有显著影响肝脏脂质积累。因此,可能存在许多因素影响NKT细胞的增殖和分化,而不仅仅是IL-15。Invariant alpha-chain Jalpha 18是iNKT细胞TCRs的主要成分,因此Jα18敲除小鼠也被用作iNKT细胞缺陷动物模型。在Jα18基因敲除(Jα18-/-)小鼠喂养胆碱缺乏-氨基酸(CDAA)饮食后,肝脏炎症因子如TNF-α、IFN-γ、IL-6、IL-17A和CXCR3、NLRP3等炎症小体激活相关基因表达的下降,但在WT小鼠没有此类表现。
众所周知,NKT细胞的发育依赖于胸腺细胞上表达的抗原呈递分子CD1d。HFHC饮食促进了肝脏中NKT细胞的浸润,而NKT缺乏(Cd1d-/-小鼠)降低肝脂质含量,纤维化相关基因如Col1a1、Col1a2和α-SMA的肝脏表达,以及CD11阳性巨噬细胞浸润。同样,MAFLD引起的脂质代谢紊乱与肝脏NKT细胞的增加有关,但在CD1d-/-小鼠中能得到改善 ,这表明NKT缺乏在预防MAFLD中起着关键作用。有时,研究人员即使使用了相同的敲除小鼠模型,仍然得到了NKT细胞对MAFLD影响的相互矛盾的结果。最近的研究表明,HFD喂养的CD1d-/-老鼠的肝脏中存在严重的脂肪积累、代谢改变和炎症介质,如IL-6、MCP1的增加。与上述结果一致,KK-A(y)小鼠作为代谢缺陷模型,在喂食HFD饮食后,肝NKT细胞比例减少。值得注意的是,据报道,Jα18-/-可以抑制MCD喂养的小鼠的NKT细胞分化,导致脂质代谢紊乱、脂肪性肝炎和肝纤维化,而过继转移iNKT细胞到Jα18-/-小鼠可以显著改善MAFLD。据报道,除了基因编辑外,腹腔注射α-GalCer (NKT激活剂)可以降低HFD诱导的MAFLD中肝脏促炎CD4/CD8+T细胞和细胞因子 (IL-2、IL-6、IL-17A、TNF-α和IFN-γ)的水平 ,同时增加iNKT细胞和抗炎细胞因子(IL-4、IL10),特别是iNKT2细胞水平,以维持肝稳态,改善肝脂肪变性。T细胞Ig和黏蛋白结构域(Tim)-3及其配体半乳糖凝集素-9 (GAL-9)相互作用,可以调节NKT细胞等免疫细胞,维持肝细胞稳态。GAL-9不仅可以使Tim-3(+)NKT细胞凋亡,诱导NKT细胞消耗,还可以通过与HFD喂养的小鼠中表达Tim-3的枯否细胞相互作用,随后分泌IL-15,促进NKT细胞增殖,改善肝脂肪变性。有趣的是,巨噬细胞迁移抑制因子 (MIF) 的特异性缺失通过降低促纤维化基因的表达,抑制NKT细胞的极化和激活,改善了代谢紊乱和肝纤维化。此外,另一篇文章指出,NK和iNKT细胞通过NKG2D信号直接杀死造血干细胞,防止肝脂肪变性和纤维化。研究还发现,与HFHC饮食相关的原位和自发性MAFLD-HCC与胆固醇升高引起的NKT细胞功能障碍有关。瑞舒伐他汀,作为一种抗胆固醇药物,可以通过抑制mTORC1/srebp2诱导的过度肝脏胆固醇合成来恢复NKT的扩增和细胞毒性。值得注意的是,菌群在MAFLD的治疗中也起着重要的作用。益生菌(双歧杆菌、乳酸菌和嗜热链球菌)通过增加NKT细胞,进而改善小鼠肝脏脂质积累和胰岛素抵抗。益生菌表面的脂质抗原促进了肝NKT细胞的增殖,说明调控肠道微生物群是一种治疗MAFLD的潜在方法。

图5. NKT细胞对MAFLD均有正、负两种作用
(A)桑基图显示了网络群落的组成在盐度胁迫增加过程中发生显著更迭。(B)各网络群落中丰富和稀有物种各占的比例。(C)不同网络之间重叠节点中丰富和稀有物种各占的比例。(D)丰富和稀有物种的生态位宽度差异(***P < 0.001;Wilcoxon秩和检验)。
NKT细胞具有更复杂的免疫特性,是治疗MAFLD的一把双刃剑。即使在相同的MAFLD基因敲除动物模型中,NKT细胞也表现出不同的实验结果,即促炎免疫细胞和因子促进脂质积累和炎症浸润,但另一方面,抗炎因子抑制MAFLD。基因编辑、药物和益生菌是通过调节NKT细胞来改善代谢损伤的有效途径。插图的一些材料引用自生物渲染器。
MAIT细胞对MAFLD的影响
MAIT细胞特异性识别微生物的抗原,仅占血液T细胞的5%,但占人类肝脏中所有T细胞的50%。因此,研究者将注意力转向了MAIT细胞和MAFLD之间的关系。非经典的I-MHC分子MR1可以捕获抗原并将其呈递给MAIT细胞,并使MAIT细胞激活。值得注意的是,MAFLD患者外周血中MAIT细胞的水平也下降,而枯否细胞中MR1的表达增加。在MCD的诱导下,MAIT细胞被招募到肝脏并以MR1依赖的方式激活,诱导M2巨噬细胞极化,并在MAFLD小鼠中改善肝脏脂肪变性。但Toubal等人发现MAIT细胞通过诱导MR1依赖的M1巨噬细胞极化,损害脂肪组织的脂质代谢,并通过诱导肠道菌群失调和增加通透性,导致肠道代谢功能障碍。其他证据还表明,MAIT细胞表现出炎症特征,改变HFD喂养小鼠内脏脂肪组织和回肠的炎症以及免疫细胞稳态。新的证据表明,与健康组相比,从MAFLD患者的PBMC中分离出的MAIT细胞显著减少。此外,人MAIT细胞和人肝肌成纤维细胞 (HMFs) 共培养实验发现,活化的MAIT细胞分泌的TNF可以增加HMFs中IL-6和IL-8的产生,增强肌成纤维细胞的活性,加重MAFLD。本研究揭示了不同的细胞因子可诱导MAIT细胞对MAFLD的相反免疫作用,为疾病治疗提供了新的方向。
讨 论
MAFLD的主要临床表现和致病机制是血脂异常和肝脏脂肪异常增加。因此,以往和目前的研究主要集中在糖脂代谢的调节作用上,而不是T细胞免疫失调在MAFLD发生中的作用。随着时间的推移,在临床和动物研究中,研究人员逐渐观察到Th1细胞、M1巨噬细胞、Th17细胞、CD8+T细胞和γδ T细胞等炎症免疫细胞在肝脏和脂肪组织中积累,而Th2细胞、Treg细胞和NKT细胞等抗炎细胞的数量和活力下降。这些观察结果表明,介导炎症细胞因子和趋化因子相关的信号通路,能够破坏免疫稳态,进而引起肝炎症反应、肝纤维化或肝癌的进一步发生。
有趣的是,各种研究表明由于细胞复杂的调节特性,相同的免疫细胞可能在不同甚至相同的动物模型中发挥相反的免疫作用。例如,在Ath喂养的TLR9缺陷小鼠中,Th2细胞的浸润诱导BM巨噬细胞抵抗M1激活,抑制了小叶炎症和MAFLD进展,而在HFD诱导的肥胖Alms1突变小鼠中引起了严重的MAFLD和肝纤维化。造成这种差异的原因可能是由于目前的模型不能完全模拟人的MAFLD。在IL-17缺陷小鼠模型中,Th22细胞浸润也加重了MCD刺激下的肝细胞脂肪毒性,但改善了肝脏炎症。这可能与Th1细胞与Th2细胞或Treg细胞与Th17细胞之间的拮抗关系、Th1细胞与CD8+T细胞或Th1细胞与Th17细胞之间的协同关系直接相关。此外,T细胞的几个亚群可以分泌多种细胞因子,同时这些细胞因子还会反过来作用于自身,进一步导致更复杂的免疫反应。尽管CD8+T细胞的激活通常会引起肝脏局部炎症,但在特殊情况下也能抑制肝纤维化和脂肪变性。据推测,某些细胞因子可能特异性地激活了CD8+T细胞的一个亚群发挥其肝保护作用。同样地,NKT细胞像一把双刃剑,作为先天免疫和获得性免疫之间的桥梁,在不同的体内动物模型中发挥恶化脂肪性肝炎或改善MAFLD的作用。此外,MAIT细胞被证明可以改善脂肪毒性和肝炎,但也会导致局部组织炎症,甚至肝纤维化。假设NKT细胞或MAIT细胞在MAFLD发生过程中受到特定因子的刺激后分化为不同的功能亚群。当然,也不能排除由免疫细胞的多重复杂性所造成的不同后果。特别值得注意的是,在以前的研究中也发现了单个免疫细胞在不同的肝病中发挥的作用是相互矛盾的。例如,Treg细胞在MAFLD中发挥保护作用,但在MAFLD-HCC过程中增强免疫抑制以加重HCC,这可能是由于Treg细胞的重编程效应引起的。
结合这些结果,我们需要关注整个免疫微环境的调节作用,而不是单个细胞或炎症因子在MAFLD中发挥的作用。此外,免疫串扰引起肝肠循环和脂肪组织的一系列变化,导致更严重的肝脏炎症。临床病例显示,饮食诱导的脂肪积累破坏了肝内T细胞的稳态(包括Th17细胞,Treg细胞,NKT细胞和CD8+T细胞),并进一步加重了MAFLD。其他研究也发现,高热量与异常的T细胞糖酵解和增殖有关。此外,有报道称MAIT细胞通过抑制双歧杆菌科(放线菌门)和乳酸菌科(厚壁菌门)的功能破坏肠道完整性,通过诱导肥胖小鼠M1巨噬细胞极化加重炎症。此外,饮食干预或有氧运动干预的患者均显示出MAFLD参数的改善,这与肝内MAIT细胞数量的显著减少相关。这些结果提示,遗传和环境以及免疫细胞代谢因素可能是影响MAFLD进展中T细胞功能的主要因素。此外,异常的T细胞会诱发免疫相关疾病,这种情况下更易感MAFLD。除了上述的免疫T细胞外,滤泡辅助性CD4+T细胞(TFH细胞)和及其分泌的分子帮助B细胞在体液免疫中发挥重要作用。然而,BMI>35和BMI<35的MAFLD患者中pTFH细胞反应没有差异。因此,TFH细胞在MAFLD中的功能仍有待探索。此外,AIH有与MAFLD相似的炎症反应,但没有明显的病理标志物,一些抗MAFLD药物还会加重这些炎症反应。因此,需要阐明免疫调节的复杂性,以探索治疗MAFLD的新有效药物和治疗方法。
不仅如此,研究人员当前面临的问题还有MAFLD在早期阶段很难被检测到,而且在临床中缺乏一线治疗。T细胞在MAFLD过程中发挥的复杂免疫调控越来越受到关注。此外,我们认为,MAFLD的治疗策略应该更多地关注如何恢复健康的免疫模式,而不是仅仅简单地诱导或抑制某些特定免疫细胞的功能或数量。另外,中药因其多靶点和多组分的特性,被认为是改善MAFLD的潜在免疫治疗调节剂。如小柴胡汤、大黄中臣丸均能调节Th17/Treg细胞平衡,抑制炎症细胞因子水平,有效减轻脂质积累和MAFLD发展。值得注意的是,传统中药也能改善肠道菌群的平衡。例如,来自植物的菊粉通过增加西式饮食小鼠中的抗菌肽和减少回肠腔内细菌来减轻饮食诱导的屏障功能障碍。天黄配方主要由三七和黄连组成,能重塑肠道菌群,增加乳酸菌及其代谢物,改善HFD诱导的MAFLD。肠道微生物群除了受到中药的影响外,还与免疫微环境密切相关。因此,肠道微生物群也有可能成为一种治疗MAFLD的有效方法。微生物群靶向干预 (可溶性膳食纤维和丁酸梭菌) 是增加MAFLD菌群多样性和改善MAFLD慢性炎症的有效方案。有证据表明,健康的肠道细菌参与缓解肠道疾病和调节免疫力。因此,在不久的将来,FMT有可能成为治疗MAFLD的特效方法。
结 论
总之,我们首先综述了MAFLD的发病机制和不同T细胞的功能。其次,我们深入总结和分析了不同的T细胞如何相互作用并参与MAFLD的发展。本研究不仅指出了T细胞的免疫调节功能在MAFLD发生过程中的重要性,而且为进一步的药物试验和机制研究的发现和发展提供了新的思路和突破。
数据可用性:
国家中医药现代化重点研发计划项目(NO.2022YFC3502100, 李晓骄阳);北京中医药大学:校级研究项目 (2022-syjs-03, 刘佳) ;国家高水平人才专项支持计划;国家自然科学基金(资助号:82274186和拨款号:82004045, 李晓骄阳);中国中医药协会青年人才促进项目(资助号:2020 -QNRC2-01至XL);北京市科学技术委员会(拨款号:7212174, 李晓骄阳)。
本文没有使用新的数据和脚本。补充材料 (图表、表格、脚本、图形摘要、幻灯片、视频、中文翻译版本和更新材料) 可以在在线DOI或iMeta科学http://www.imeta.science/中找到。
引文格式:
Jia Liu, Mingning Ding,Jinzhao Bai, Ranyi Luo, Runping Liu, Jiaorong Qu, Xiaojiaoyang Li. 2022. Decoding the role of immune T cells: A new territory for improvement of metabolic-associated fatty liver disease. iMeta. https://doi.org/10.1002/imt2.76
作者简介

刘佳(第一作者)
● 北京中医药大学生命科学学院,实验师。
● 目前研究方向为中药防治免疫性和炎症型肝病,参与承担多项国家自然科学基金面上及青年项目、北京市自然科学基金面上项目等科研课题,以第一作者发表相关学术成果于iMeta、Animal Models and Experimental Medicine、Electroanalysis等期刊。

丁明宁(第一作者)
● 北京中医药大学生命科学学院中西医结合基础,医学硕士。
● 在校期间获研究生学业一等奖学金。目前研究的主要课题非酒精性脂肪肝病等慢性肝病的中药防治,以第一作者发表相关学术成果于iMeta、Frontiers in Pharmacology、Chinese Herbal Medicines等期刊。

李晓骄阳(通讯作者)
● 北京中医药大学生命科学学院 教授,博士生导师。
● 美国弗吉尼亚大学博士后。入选国家特殊人才支持计划、北京市科技新星,中华中医药学会青年托举工程等,获得AASLD Foundation Fellow Award。兼任中华中医药学会青年委员会委员、中华中医药学会感染病分会委员、中国中西医结合学会科研院所工作委员会青年委员等;担任Frontiers in Pharmacology、CJNM、CHM、中草药等责任编辑及编委。担任国家自然科学基金评审专家、北京市科技专家库科技研发类评审专家、中华中医药学会科技奖励专家等。主要研究方向为中医药防治肝纤维化。主持国家自然科学基金面上及青年项目、北京市自然科学基金面上项目等10项科研课题,参与承担国家重点研发计划、国家自然科学基金国际合作项目等。以通讯或第一作者在iMeta、Hepatology、Theranostics、APSB、APS等期刊发表论文40余篇,总引用次数1600余次。申请专利13项,参编著作1部。两次参与获得省科学技术进步一等奖。
更多推荐
(▼ 点击跳转)
iMeta | 引用7000+,海普洛斯陈实富发布新版fastp,更快更好地处理FASTQ数据
iMeta | 德国国家肿瘤中心顾祖光发表复杂热图(ComplexHeatmap)可视化方法

1卷1期

1卷2期

1卷3期

1卷4期

2卷1期

2卷2期

2卷3期

2卷4期

3卷1期

2卷2期封底

2卷4期封底

3卷2期
期刊简介
“iMeta” 是由威立、肠菌分会和本领域数百位华人科学家合作出版的开放获取期刊,主编由中科院微生物所刘双江研究员和荷兰格罗宁根大学傅静远教授担任。目的是发表原创研究、方法和综述以促进宏基因组学、微生物组和生物信息学发展。目标是发表前10%(IF > 20)的高影响力论文。期刊特色包括视频投稿、可重复分析、图片打磨、青年编委、50万用户的社交媒体宣传等。2022年2月正式创刊发行!目前期刊已经被ESCI、PubMed、Scopus等数据库收录。
联系我们
iMeta主页:
http://www.imeta.science
姊妹刊iMetaOmics主页:
http://www.imeta.science/imetaomics/
出版社iMeta主页:
https://onlinelibrary.wiley.com/journal/2770596x
出版社iMetaOmics主页:
https://onlinelibrary.wiley.com/journal/29969514
iMeta投稿:
https://wiley.atyponrex.com/journal/IMT2
iMetaOmics投稿:
https://wiley.atyponrex.com/journal/IMO2
邮箱:
office@imeta.science
高颜值免费 SCI 在线绘图(点击图片直达)
最全植物基因组数据库IMP (点击图片直达)
往期精品(点击图片直达文字对应教程)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
















































 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









