







第二章
本章重点:
总结EEG,诱发电位(EP)和事件相关电位(ERP)是怎么产生并被获取的.
根据刺激呈现模态的不同,简要介绍了不同感觉模态下的经典EP 和ERP 成分及其在临床和神经科学研究中的应用。
讨论了脑电技术的优势和局限性。
第一节 自发性脑电活动
大脑的自发性电活动,又被称为自发性脑电活动,表现为在广泛频谱上占主导地位且具有某些特征的波形。
这些 EEG 活动通常被应用于癫痫、昏迷和脑死亡等临床诊断。
自发性 EEG 活动可以分为以下五个频段:
δ \delta δ(<4Hz)、 θ \theta θ(4~8Hz)、 α \alpha α(8~13Hz)、 β \beta β(13~30Hz)和 γ \gamma γ(>30Hz)。
每个频段的节律活动都具有特定的头皮分布和生物学意义。
人处于清醒状态且闭眼, α \alpha α波通常会在其 大脑枕叶区域测得,并且会随着睁眼或心理活动的增加而减弱。
随着频率的增 加,脑电节律倾向于表现为更小的振幅

第二节 诱发电位和事件相关电位
本能在前,感觉在后
概念解释
当人接收到与特定感觉、认知或运动事件相关的刺激时,自发性 EEG 活动会受到干扰。
由事件诱发的神经响应会淹没在自发性 EEG 活动 中,但可以借由简单的平均叠加技术或更复杂的单试次分析和时频分析等技术,将这些响应从自发性 EEG 活动中提取出来。
这些经过平均叠加所获得的脑电响应被称为事件相关电位,表示它们是与特定事件相关的电位。
传统观点认为大脑响应的早期成分是由刺激本身所诱发的,反映的是对刺激进行基本感觉处理的大脑活动。
研究者使用**“诱发电位”**这一术语来指代对外部刺激诱发大脑响应的测量。
诱发电位是指神经系统(从感受器到大脑皮层)对特定的刺激(如光、声、触等)做出的反应,这些反应可以在脑的相应部位被检测出来,并且与刺激有相对固定的时间间隔和特定位相。
当神经活动与心理事件具有锁时相关性时,某些大脑响应(尤其是那些潜伏期较晚的响应)反映的更可能是大脑皮层对心理活动的处理,而不是简单的感觉处理,因此 这些被记录到的大脑响应也被称为“事件相关电位”
不只是本能的反应,还有心理活动反应
事件相关电位则是在特定的心理事件或物理事件发生时,在大脑中产生的电位变化,这些电位变化与事件有直接的时间关联,并且可以通过滤波和信号叠加技术从脑电信号中被提取出来。
表明该响应不仅反映了感觉运动过程所诱发的活动,还显示了与可定义事件之间稳定的时间关系.
目前,ERP 技术已被生理学、心理学和临床医学等多个领域的研究者广泛 应用。
成分特点
诱发电位的成分通常受刺激物理特性的影响,例如视觉诱发电位中的P1、N1、P2等成分,它们的波形和潜伏期与刺激的亮度、颜色等物理特性有关。
P1
P1波是视觉ERP中的一个早期成分,它出现在刺激后大约100ms,主要起源于背侧外纹状皮层(枕中回)的早期部分,晚期部分的起源则主要在腹侧梭状回。P1波的振幅会受到觉醒水平的影响,在高觉醒条件下,P1波的振幅较高。
N1
N1波紧随P1波之后出现,它包含了多个子成分,这些子成分的峰值在刺激后100-150ms出现,位于前部头皮电极位置。N1波对注意力敏感,且具有高度的不应性,即在短时间内连续出现两个刺激时,第二个刺激诱发的响应会减小很多。
P2
P2波跟随N1波形之后出现,位于头前部与中央头皮区。P2波的振幅在刺激包含目标特征时会增大,而且在目标出现概率较低时,其反应也会增强。
参考https://blog.csdn.net/zyb228/article/details/106734977

事件相关电位的成分则更多地反映了心理活动的某些方面,例如P300(P3)是ERPs中最受关注的内源性成分,它与被试的精神状态和注意力有关,不受刺激物理特性的影响。
第三节 EP和ERP的概述
脑电是一种低空间分辨率的大脑活动测量方法,具 有较低的信噪比。其原始形式不能直接用于测量大多数与特定事件或任务相关的神经活动。
通过简单的平均技术可以从连续的 EEG 数据中提取与刺激有关的锁时的事件相关响应(Mouraux & Iannetti, 2008;Pfurtscheller & Lopes da Silva,1999)。
锁相性:
时间发生的时序间隔的固定性。
相对于刺激呈现,锁时的事件相关响应可以是锁相的,也可以是非锁相的。当事件相关的响应与刺激开始时刻相位锁定时,研究者通过平均多个试次后可以在时域中很容易地观测到它们,即 ERP。
当事件相关响应与刺激开始时刻相位不锁定时,多个试次的平均叠加会让它们隐匿于时域中而难以观测。这些非锁相响应被称为事件相关振荡(event-related oscillation,ERO)。
表示正在进行的 EEG 活动的特定频率的变化。ERO 可能表现为与潜在神经元集群同步性的瞬时增加,即事件相关同步化(event-related synchronization, ERS)
或瞬时减少,即事件相关去同步化(event-related desynchronization, ERD)
解释ERP的产生
ERP是怎么产生的?
诱发模型:
一些研究者认为,ERP 与正在进行的自发性 EEG 活动无关,是一种额外的神经元放电活动(Jervis et al.,1983; Niedermeyer & Lopes da Silva,2005;Schroeder et al.,1995)
相位重置模型:
ERP 是由正在进行的自发 EEG 信号的相位重组而产生的.
综合两种模型的解释更合理
Min 等 (2007)观察到部分相位重置和部分附加功率对 ERP 的产生都有贡献, 并且刺激前的大脑状态显著影响事件相关的大脑响应,提示综合两种模型来解释 ERP 的发生可能更加合理且全面.
获得ERP并定位其神经源
在大多数情况下,背景 EEG 活动 (大约几十微伏)的幅度比 ERP的幅度(微伏级)大几倍(Hu et al.,2010; Rugg & Coles,1995)
ERP 的识别依赖于增强信噪比的信号处理 方法。
被广泛使用的信噪比增强方法(平均化技术)是时域上的跨试次平均叠加.
(Dawson,1951,1954)。
平均后的 ERP 成为具有极性、潜伏期、振幅和头皮分布等多个特征的单相偏转.
(Callaway et al.,1978;Mouraux & Iannetti,2008)。
这几个特征的说明:
- 极性(Polarity):ERP的极性指的是电位的正负方向。通常,正电位表示为向上的波峰,而负电位表示为向下的波谷。
- 潜伏期(Latency):潜伏期是指从刺激发生到ERP成分达到峰值所需的时间。潜伏期可以反映神经信号在大脑中的传导速度和处理时间。
- 振幅(Amplitude):振幅是指ERP波形的高度,通常以微伏(μV)为单位。振幅的大小可以反映神经活动的强度或参与的神经元数量。
- 头皮分布(Scalp Distribution):ERP的头皮分布指的是电位在头皮上的地理分布模式。不同的ERP成分可能在头皮的不同区域有最大的响应,这可以提供有关大脑激活区域的信息。
- 单相偏转(Monophasic Deflection):这通常指的是ERP波形中单一方向的偏转,可以是正向(向上)或负向(向下)的。单相偏转的ERP成分相对简单,易于识别和测量。
信噪比增强的方法:
信号处理技术
预处理滤波器
使用数字滤波器如均值滤波器、中值滤波器和高斯滤波器等来去除噪声,这些滤波器可以利用信号的统计特性来减小噪声的影响。
自适应滤波器
该算法使用一个模型来估计信号和噪声的特性,并根据估计结果对信号进行滤波。自适应滤波器能够根据实时环境的变化自动调整滤波参数,从而提高滤波效果。
子空间方法
这些方法利用信号和噪声在不同子空间的分布特性来分离它们。常见的子空间方法包括主成分分析(PCA)和独立成分分析(ICA)等
去噪算法:去噪算法通过对信号进行降噪处理来提升信噪比。常见的去噪算法包括小波去噪、基于稀疏表示的去噪和低秩矩阵恢复等。
物理修正算法
空间滤波技术:通过对多个传感器的信号进行组合,得到增强信号,从而有效地消除噪声。
时域滤波技术:通过对信号的波形进行调整,以消除干扰信号,提升信噪比。
平均化技术
时间平均化:通过多次采集同一信号,在时间轴上进行平均,使得信号的噪声得到消除。
空间平均化:通过多个传感器对同一信号进行采集,在空间上进行平均,以提升信号质量。
使用偶极子源模型(Valeriani et al.,2001)或分布式源模型(Michel et al.,2001)可以从头皮分布推断 ERP 的神经源。
缺陷:同一个给定的头皮地形图可以推导出不同的源位置,因此从头皮 位置确定源位置是典型的逆问题(Grech et al.,2008)。这种情况需要额外的假设(例如,偶极子源的数量或来自功能磁共振成像的空间约束) 以获得唯一解。由于这个原因,所获源位置可能存在严重的假设偏差, 导致其在实际应用方面受到极大限制(Mouraux & Iannetti,2008)
无法进行有效的约束求解。
第四节 常见的EP和ERP成分
说明:N,P加响应时间,这个N和P代表的是电极的正负
一、听觉诱发电位
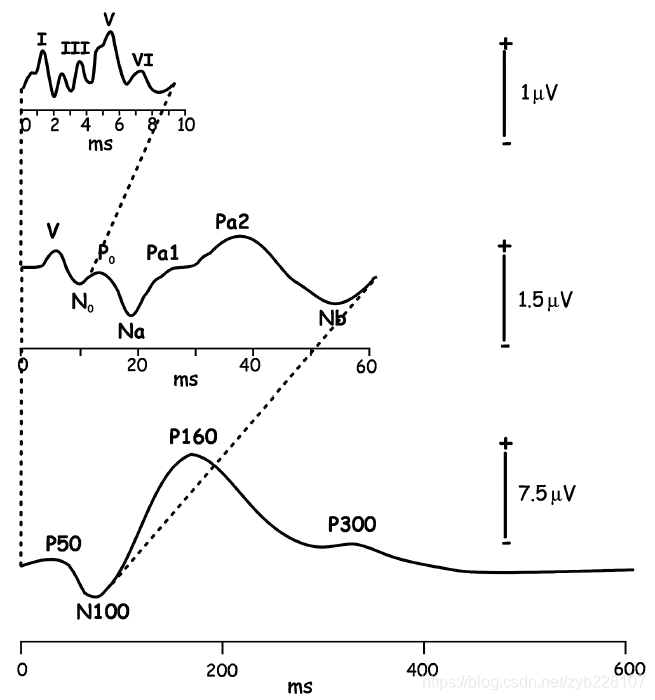
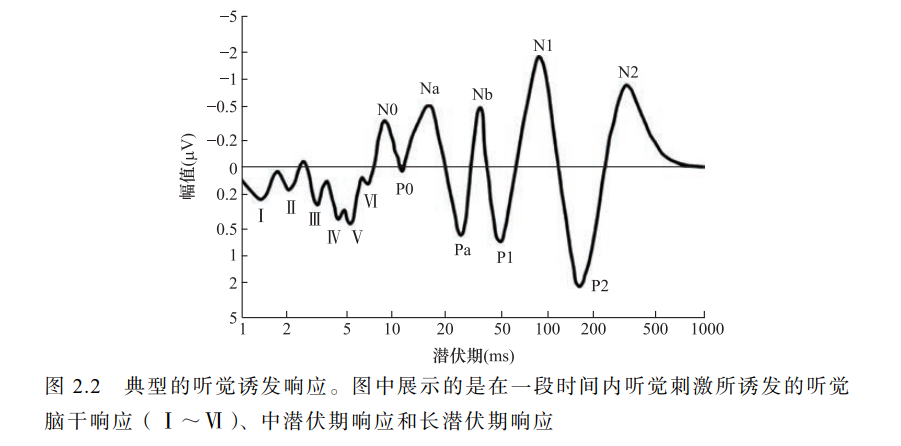
听觉刺激的呈现可以在耳蜗诱发电位。在突然出现一个声音刺激后, 第一组听觉响应在声音呈现后的几毫秒内发生,反映了声音信号从耳蜗通过脑干传入丘脑(图 2.2)。
BAEP
这些信号的源在大脑深处且通常情况下振幅较小(约 0.5μV),因此需要平均数千个试次才能获得清晰的脑干听觉 诱发电位(brainstem auditory evoked potential,BAEP)。
这些听觉脑干响 应通常用罗马数字按顺序标注,分别代表听觉神经(Ⅰ)、耳蜗核(Ⅱ)、 上橄榄(Ⅲ)、侧脑室束和核(Ⅳ)及下丘(Ⅴ)的激活。
BAEP 是高度自动化的,并且较少受睡眠、麻醉、昏迷的影响(Goldie et al.,1981)。
因此,BAEP 可用于评估听觉通路的完整性,尤其是对新生儿和婴儿的听觉评估,是早期识别听力损伤或丧失的重要工具。
潜伏响应:
在 BAEP 之后是中潜伏期响应和长潜伏期响应。
中潜伏期响应是指 刺激呈现后 10~50ms 的神经响应,至少有一部分来自内侧膝状核和初级听觉皮层;
长潜伏期响应通常从 P50(也称为 P1)、N100(也称为 N1) 和 P160(也称为 P2)开始(Luck,2005)
高级认知成分
“长潜伏期响应”这一 术语在某种程度上具有混淆性,因为这些响应的潜伏期相对于中潜伏期响应更长,但与高级认知成分(例如,P300 和 N400)相比,则潜伏期较短。
因此,一些研究者在提及“中潜伏期响应”时一般是指发生在 50~ 200ms 的诱发电位成分(Boutros et al.,2006;Roth et al.,1980)。
听觉 N1
峰值潜伏期为 50~150ms 的听觉 N1 是听觉诱发电位中最突出的成分之一。
N1 有多个子成分,它们叠加在一起形成了 N1 的峰值。
其中包括
-
在颞上平面记录到的与刺激的物理和时间属性以及被试的一般状态相 关的成分,
-
在颞叶和顶叶皮层侧面的联合皮层中产生的成分,
-
以及在运动和前运动皮层中产生的成分。
这些子成分的诱发不仅取决于听觉刺激本身,也取决于刺激发生的条件(Näätänen & Picton,1987)。听觉 N1 对注意力敏感(Woldorff et al.,1993),研究者可以通过改变事件的任务相关性或通过调节被试对事件本身的预期来操纵 N1(Lange,2013).
听觉N2
N2 是另一个常见的听觉成分。
偶尔出现在重复刺激序列(标准刺激) 中的变异刺激往往会诱发出一个波幅较大的 N2。
N2 通常包含三个子成分:N2a、N2b 和 N2c。
N2a 也被称为听觉失匹配负波(mismatch negativity, MMN),反映的是无须意识参与的变异探测过程。MMN 指的是标准刺激和变异刺激所诱发的神经响应之间的差异波,通常在变异刺激呈现后 150~250ms 出现。MMN 的分布主要集中在头皮的前中部,其主要发生源为听觉皮层和前额叶(Näätänen et al.,2007)。此外,在违反语法规则或声音缺失时,MMN 也会被诱发(Yabe et al.,1997)。尽管 MMN 被认为是一种独立于注意力的过程,但有新证据表明可以通过调节注意力来对 MMN 进行调节(Sussman,2007)。总的来说,MMN 因其相对较高的自动性,已被广泛用于对说话困难或行为不便人群的听觉变异探测能力的评估。
N2b 反映的是注意条件下的变异探测过程,其潜伏期要晚于 MMN,在变异刺激呈现后 200~300ms 达到峰值。此外,N2b 的头皮分 布也与 MMN 不同,主要集中在中央顶叶,且极性在乳突位置不反转, 提示其发生源可能在听觉皮层之外。
N2c 与 MMN 和 N2b 都不同,被认为反映的是刺激归类过程而非变异探测.
二、视觉诱发电位
C1,P1,N1,P2,N2,P3
第一个典型的视觉 ERP 成分是 P1(图 2.3),
但有时 P1 之前会出现起始潜伏期为 40~70ms、峰值潜伏期为 60~100ms 的 C1 成分。
C1 对空间频率和对比度等基本视觉刺激属性高度敏感。
有证据表明,C1 源于纹状皮层,会对上下视野中的视觉刺激出现极性反转(Jeffreys & Axford, 1972)。
这种反转对应于纹状皮层的视网膜组织,其中下部和上部的视觉信息分别被映射在矩状裂的上部和下部。
上下视野刺激会引起C1的电极变化
当 C1 波为正时,它与 P1 成分相加产生单个正向波。
因此,除非通过上视野刺激产生负向 C1 波以区分正向 P1 波,否则我们通常难以观察到明显的 C1 波(Luck,2005)。
不容易观察,因为容易和P1进行叠加
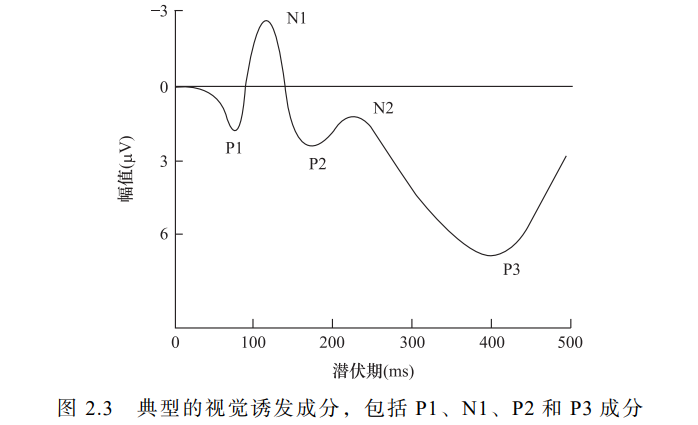
视觉 P1
视觉 P1 在两侧枕区的电极位置处最大,并且在刺激呈现后 60~90ms 出现,峰值在 100~130ms 达到最大。
与 C1 成分不同,P1 成分对于上视野刺激和下视野刺激不会出现极性反转。尽管学术界目前尚未就其来源 的确切位置达成共识,但研究者普遍认为 P1 成分产生于外部视觉区域 (Di Russo et al.,2002)。另外,P1 波与 C1 波一样对刺激属性的变化敏感, 受到选择性注意(Hillyard et al.,1998)和个人唤醒度的调节(Vogel & Luck,2000)。
视觉 N1
视觉 N1 出现在 P1 之后,易出现不应期。如果一个视觉刺激出现在某一位置,而另一个视觉刺激在短暂延迟后出现在同一位置,那么第二 个刺激所诱发的 N1 响应会大大减弱。这是由于产生 N1 的神经元在响应后进入不应期,需要一段时间才能逐渐恢复。
与听觉 N1 一样,视觉 N1也包括多个子成分。
例如,最早的 N1 子成分在刺激呈现后 100~150ms 在额叶电极达到峰值,
而在刺激呈现后 150~200ms 至少有两个 N1子成分在顶叶皮层和枕叶皮层达到峰值,而且这些 N1 子成分都会受到空间 注意力的影响(Hillyard et al.,1998)。
此外,当被试执行辨别任务时, 两侧枕区的 N1 子成分的响应似乎比执行检测任务时更大,提示这个 N1 子成分可能反映了某种辨别过程(Vogel & Luck,2000)。
三、痛觉诱发电位
红外激光刺激器产生的短暂热脉冲辐射会选择性地激活位于皮肤表 层的 Aδ 和 C 纤维(Bromm & Treede,1984;Carmon et al.,1976),在 EEG 上表现为多个响应(Carmon et al.,1976)。
激光诱发电位(laser evoked potential,LEP)与Ⅱ型 Aδ 机械热伤害感受器(Treede et al.,1998)和 位于脊髓前角的脊髓神经元的激活有关(Treede et al.,2003)。
LEP 包括 一系列的锁时 ERP 成分——N1、N2 和 P2 波。
这些成分反映的是来自初级和次级躯体感觉皮层、脑岛和前扣带皮层的皮层活动组合(Cruccu et al.,2008;Garcia-Larrea et al.,2003)。
如图 2.4 所示,LEP 中最大的成分是负正顶点电位,即 N2-P2 复合波,通常在激光刺激施加于**手背后 200~350ms **达到峰值(Bromm & Treede,1984)。
N2-P2 复合波之前是波幅较小的 N1 波,它在刺激对侧的中央或颞叶区域的分布最大。N1 被认为反映的是躯体感觉信息的输入(Lee et al.,2009;Mouraux & Iannetti,2009)
N1 波会在时间和空间上与后续的 N2 波重叠(Cruccu et al.,2008;Kunde & Treede,1993;Treede et al.,2003)。

N1波的研究有更大的价值
N1 波所代表的感觉处理的早期阶段与伤害性信息的上传直接相关
为了将 N1 波与 N2 波分离, 研究者通常在刺激施加的**对侧中心电极(C3 或 C4)**处检测 N1b 波,该方法也被推荐用于临床试验中 LEP 的记录(Hu et al.,2010;Valentini et al.,2012)。
N2-P2 复合波的特点是信噪比高(峰峰间振幅在平均叠加 20~30 个 试次后即可达几十微伏)(Carmon et al.,1980;Iannetti et al.,2005), 但 N1 波的信噪比较低,也因此更难以探测。
信噪比低的原因
这种困难的形成不仅是因为产生 N1 波的神经活动小于产生 N2 和 P2 波的神经活动(Cruccu et al., 2008;Treede et al.,2003),也是因为 N1 和 N2 波在时间和空间上具有相同极性且容易重叠,而且颞叶电极经常被颞肌活动相关的伪迹所污染 (Kunde & Treede,1993)。
因此,在过去几十年中,绝大多数的生理学 (Iannetti et al.,2003)和临床(Treede et al.,2003)LEP 研究,都单独依赖于对 N2 和 P2 波的测量。然而,近年来越来越多的研究开始探索并通过实验调控 N1 波的潜伏期和振幅,以描述其功能意义(Ellrich et al., 2007;Iannetti et al.,2008;Lee et al.,2009;Legrain et al.,2002;Mouraux & Iannetti,2009;Schmahl et al.,2004)。
事实上,有实验证据表明,N1 波所代表的感觉处理的早期阶段与伤害性信息的上传直接相关(Lee et al.,2009),而 N2 和 P2 波所反映的神经活动在很大程度上不具备刺激模态的特异性(Mouraux & Iannetti,2009)。
基于这些原因,研究者建议 对 N1波进行更系统的检查,以促进 LEP 在临床中的应用(Cruccu et al., 2008;Treede et al.,2003)
晚期成分P4
LEP 的晚期成分可能对躯体感觉特异(Liang et al.,2010;Mouraux & Iannetti,2009)。
虽然多年来一直被忽视,但这一成分已经被成功地识别并从 LEP 波形中提取,标记为 P4(Hu et al.,2014)。
P4 波的特征为信噪比较低、波幅较小且潜伏期较长。具体而言,在手部施加激光刺激后 390~410ms,可以在施加刺激侧对侧的中央顶点电极处观察到较大的 P4 波。
P4和N1的紧密联系
相比较而言,当刺激施加于足部时,诱发的 P4 波是呈中央分布的, 在 Cz 和 Pz 电极之间具有最大振幅,潜伏期在 430~450ms(Hu et al., 2014)。值得注意的是,P4 和 N1 波的神经活动之间存在紧密的生理联系,表现为 P4 波的峰值潜伏期和振幅在很大程度上依赖于 N1 波的峰值潜伏期和振幅。
同时,大量证据表明,P4 波与 N1 波可能来自相同的神经发生源(Hu et al.,2010;Hu et al.,2014;Valentini et al.,2012)。可见,除了 LEP 波形中的前三个主要成分之外,由初级躯体感觉皮层(至 少部分地)产生的 P4 波可以被认为是第四个独立的 LEP 成分,反映了对伤害性信息处理的独特功能。
临床应用
在生理学和临床研究中,LEP 被认为是评估伤害性刺激传导通路功能的最佳工具(Bromm & Treede,1991;Cruccu et al.,2008;Iannetti et al.,2001)。
如果某些损伤影响了脊髓-丘脑系统中的任何位置,只要将激光刺激作用于相应的皮肤区域,其所诱发的 LEP 就可以反映伤害性刺激传导通路中的异常(Cruccu et al.,2008)。
在外周水平上,LEP 可用于 评估“小纤维疾病”,其特征是对温度和疼痛敏感性的降低以及自主周围神经功能的丧失,在 LEP 上表现为振幅的衰减或潜伏期的延迟,甚至是没有响应(Agostino et al.,2000;Kakigi et al.,1992;Kakigi et al.,1991b)。 例如,研究者观察到 LEP 的潜伏期和振幅的变化与腓肠神经活检中 Aδ 纤维的丧失相关,并且 LEP 的完全消失可能表明 Aδ 纤维和 C 纤维均受 损(Kakigi et al.,1991b;Treede et al.,2003)。
在脊髓水平上,LEP 可 以提供脊柱病变的功能评估。研究表明,LEP 异常与脊髓空洞症、动静 脉畸形和炎症性脊髓病患者的脊柱病变相关(Bromm et al.,1991;Kakigi et al.,1991a;Treede et al.,1991;Treede et al.,2003)。
在脑干水平, 下侧脑干的损伤也可能导致 LEP 异常,表现为振幅减弱、潜伏期延迟或 没有响应(Treede et al.,2003)。
在皮层水平上,额顶岛盖(次级躯体感 觉皮层或脑岛)的病变与疼痛敏感性的降低相关(Greenspan et al.,1999), 表现为 LEP 的异常。因此,LEP(特别是 N1)经常被用于评估皮层病变 (Treede et al.,2003)。
四、触觉诱发电位
体感诱发电位(somatosensory evoked potential,SEP)是指大髓鞘纤维 Aβ 接受刺激后在皮层和皮层下所诱发的响应(Cruccu et al.,2008; Devlin et al.,2006)。
Aβ 纤维检测到诱发触觉,由同侧脊髓传至脑干, 然后将这种触觉信息直接投射到对侧丘脑,并终止于腹侧后外侧核(初级躯体感觉皮层)(Kandel et al.,2000)和其他与躯体感觉相关的区域,这 个过程可以实现对机械刺激的感知,如触觉识别和振动检测(Treede, 2007)。
例如,当对胫神经施加电刺激时(通常使用双极经皮电极,其中阴极位于跟腱内侧边缘和内踝后缘之间的位置,阳极位于远端约 3cm 处),所诱发的胫神经 SEP 可在不同的水平被记录到,包括外周(N8)、 腰椎(N22)、皮层下(P30)和皮层上(P39)(Cruccu et al.,2008)。
P39
在皮层水平,SEP 通常在 Cz 电极处记录。如图 2.5 所示,虽然皮层水平的胫神经 SEP 包含多个锁时成分**(P39、N50、P60)**,但只有 P39 具有明确 的临床价值(Cruccu et al.,2008;Mauguiere,1999)

检测应用
SEP 的潜伏期和波幅都很明确(Minahan,2002),所以 SEP 常用于 检测从外周感觉神经到大脑感觉区域的躯体感觉通路的功能的完整性 (Cruccu et al.,2008;Kraft et al.,1998)。
SEP 响应的异常为躯体感觉系统的损害提供了可靠的证据(Cruccu et al.,2008),因此 SEP 已被广泛用于临床诊断(Aminoff et al.,1988;Yiannikas & Vucic,2008;Zeman & Yiannikas,1989)和术中神经生理学监测(Deletis & Shils,2002;Devlin et al.,2006;Hu et al.,2003;Luk et al.,2001;Minahan,2002;Nuwer, 1998)。
例如,与 LEP 类似,SEP 的异常通常表现为峰峰之间的间隔显著延长或在严重情况下缺乏必要的响应(Kraft et al.,1998)。在外周水平 上,SEP 可用于诊断发生严重病变的周围神经,包括炎症性多发性神经 病变(Parry & Aminoff,1987)、感染性疾病(Moglia et al.,1991)和中 毒性神经病变(Krarup-Hansen et al.,1993)。
在脊髓水平上,当疾病影 响脊髓的上行通路时,触觉信息的传导也会受到影响,因此可以使用 SEP 来评估通路的功能完整性,例如,利用 SEP 评估由脊髓肿瘤引起的躯体 感觉通路损伤(Livshits et al.,1992)。此外,SEP 还可以帮助确定生理 上未受影响的神经组织的边界,如脊髓空洞症(Anderson et al.,1986), 从而辅助医生进行病灶的手术切除。
在脑干水平上,当脑干病变涉及丘索通路时,SEP 也会表现异常(Cruccu et al.,2008)。
在皮层水平上,躯体感觉通路的损害也会导致 SEP 的异常。例如,多发性硬化患者常伴有胫神经 SEP 异常(Davis et al.,1985;De Pablos & Agirre,2006;Loncarevic et al.,2008)。
在术中监测方面,SEP 可以用于防止神经损伤、跟踪生理 变化以及定位中央沟(Cruccu et al.,2008;Deletis & Shils,2002)。此外,实时记录的 SEP 有助于监测脊髓手术期间因脊髓血液供应不足或机械压迫导致的暂时性功能障碍,从而预防不可逆的脊髓损伤(Rossi et al., 2007;Wiedemayer et al.,2002)
五、稳态诱发电位
诱发电位不仅可以由以相对缓慢的速率所呈现的独立、离散的刺激诱发产生(即瞬态响应),还可以由以快速固定速率呈现的系列刺激诱发产生。
是否可以理解为身体对某种连续的刺激采取的高效应激反应.
因为其对这种周期性刺激的响应随时间变化的幅度和相位非常稳定,所以这些响应也被称为稳态诱发电位(steady-state evoked potential, SSEP),表明神经系统处于一种稳定的状态(Regan,1966)。
定义:
稳态诱发电位(Steady-State Evoked Potentials, SSEPs)是一种特殊类型的脑电活动,它由持续的周期性刺激(如声音、光线或触觉)诱发,并在脑电图(EEG)中表现为与刺激频率同步的波动。这种电位反应与刺激的频率具有稳定的关系,因此得名“稳态”。SSEPs在临床医学中有着广泛的应用,特别是在听力学和神经生理学领域。
SSEP 是神经元以刺激呈现频率对刺激的共振响应,可以被认为是由感觉刺激的快速重复而引起的独立瞬态响应的线性叠加。
不同的 SSEP 频率可用于标记不同感觉模态下的刺激输入,从而分离出与每个感觉输入特定相关的神经活动(Nozaradan et al., 2012)。
此外,由于刺激呈现速度快,可以在短时间内进行数百个试次, 因此 SSEP 常被用于对感觉障碍的诊断。
分类:
视觉稳态诱发电位(SSVEP)、听觉稳态诱发电位(ASSR)和触觉稳态诱发电位等
稳态听觉诱发电位
(steady-state auditory evoked potential, SSAEP)
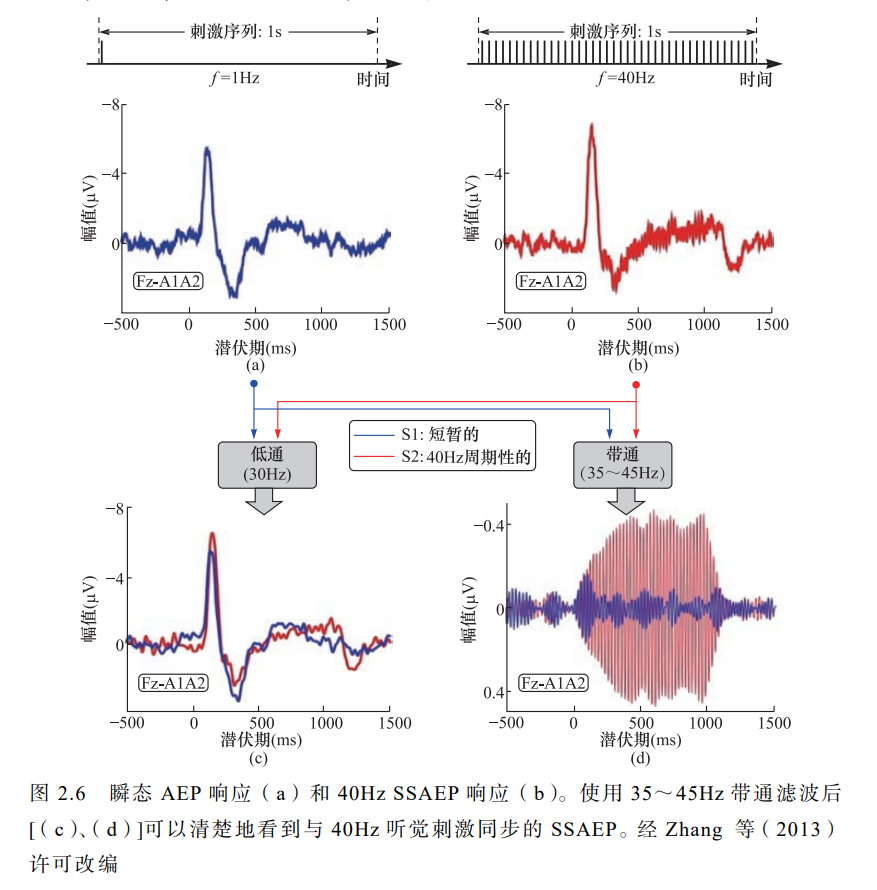
ASSR是由多个频率持续的声音刺激信号诱发而产生的,它可以一次性地对多个频率的听力状况做出判断,并且可以将客观检查的结果转换成纯音听力图。
 经常在 40Hz 出现清晰的频谱峰值( Bohórquez & Ozdamar,2008;Zhang et al.,2013)。由 40Hz 刺激引起的 SSAEP 在很 大程度上可以由瞬时听觉诱发电位(auditory evoked potential,AEP)
经常在 40Hz 出现清晰的频谱峰值( Bohórquez & Ozdamar,2008;Zhang et al.,2013)。由 40Hz 刺激引起的 SSAEP 在很 大程度上可以由瞬时听觉诱发电位(auditory evoked potential,AEP)
例 如,听觉脑干响应和中潜伏期响应的线性总和来解释。
与缺乏频率特异性的 AEP 相比,SSAEP 可以通过调幅和/或调频来诱发。
由于其频率特 异性, SSAEP 在临床上常用于预测所有年龄段患者的听力敏感性 (O’Donnell et al.,2013)。
听力学应用
听力评估:ASSR可以用来评估个体的听力水平,特别是对于婴幼儿和听障者,他们可能无法完成传统的主观测听。通过ASSR,可以得到不同频率处的听阈,从而评估听力状况。
助听器验配:ASSR可以帮助验配助听器及其评估助听效果。由于其频率特异性好,可以分别得到左、右耳500Hz、1000Hz、2000Hz和4000Hz八个频率处的听阈,有助于调整助听器的设置。
残余听力测定:对于已经遭受听力损伤的个体,ASSR可以用来测定残余听力,这对于制定听力恢复计划和干预措施至关重要
诊断听力障碍:ASSR可以帮助诊断各种类型的听力障碍,包括传导性聋、感音神经性聋等,因为它能够反映听觉系统的功能状态。
手术前后评估:在进行耳部手术之前和之后,ASSR可以用来评估听力的变化,帮助医生做出最佳的治疗决策。
稳态视觉诱发电位
对于视觉模态,周期性视觉刺激(例如,闪烁、 反转棋盘格和移动光栅)可以诱发稳态视觉诱发电位(steady-state visual evoked potential,SSVEP),这些响应通常在视觉皮层的 V1 中产生.
SSVEP 可用于视觉注意和工作记忆等认知评估,以及衰老、抑郁、焦虑和精神分裂症等的临床研究。
SSVEP 也被应用于脑机接口的相关研究中,(Norcia et al.,2015;Vialatte et al.,2010)。
优势:
首先,高频刺激的使用能够缓解基于SSVEP的BCI产生的用户视觉疲劳,提升系统的舒适度和安全性。这对于长时间使用BCI系统的用户来说尤为重要,因为它可以减少因视觉疲劳导致的不适感和错误率。
其次,高频SSVEP系统的研究有助于提高BCI的信息传输速率(ITR)。例如,一项研究展示了一种新型双频SSVEP BCIs系统,它利用主视眼效应引起的双眼视觉资源分布差异有效区分相同频率的目标,并通过全球优化确定编码,构建出多个不同的目标。该系统的平均信息传输速率达到了234.13比特/分钟,这是目前双频SSVEP BCI中性能较高的研究之一。
https://www.sohu.com/a/782073501_121181007/
高频SSVEP系统的研究还涉及到新的刺激编码方法,如亮度和运动同步调制的新型SSVEP BCIs系统。这种系统通过结合亮度和运动的视觉闪烁对刺激目标的频率和相位进行编码,提高了系统性能。
https://blog.csdn.net/weixin_51306846/article/details/135072541
伤害性躯体感觉模态
当在手背或脚背施加快速的周期性热刺激时(如红外激光脉冲序列),这 种皮肤伤害性感受器的选择性激活也可以引起 SSEP,并在头皮顶点处达 到最大且对称分布在两个半球上,表明产生源来自大脑的中线区域(Colon et al.,2012;Mouraux et al.,2011)。
第五节 脑电技术的优势和局限性
简要介绍了自发性 EEG 活动,以及不同感觉模态下的经典 EP和 ERP 成分及其在临床和神经科学研究中的应用。EEG 和 ERP 因其出 色的时间分辨率越来越受欢迎,很好地弥补了高空间分辨率的正电子发射断层扫描技术和功能磁共振成像技术 (一种依赖脑血流和脑血氧的变化的脑功能研究技术)(De Yoe et al., 1994;Villringer & Dirnagl,1995)在时间分辨率上的不足。
首先,因为设备相对便宜且进行实验所需的一次性用品较少,相比于其他研究脑功能的神经成像技术,EEG 成本更低。其次,相比于微电极记录(microelectrode recording,MER)(Hua et al.,2000)和皮层脑电图(Kuruvilla & Flink,2003)等其他电生理学技术,EEG 技术具有无创性优点。最后,EEG 是对神经元活动的直接测量,其采样率可以高达几万赫兹(Inoue et al.,2001;Ozaki et al.,1998),因此具有极高的时间分辨率。
然而,EEG 技术的空间分辨率较差,这主要由三个因素引起:
①介于大脑和记录电极之间的皮肤、颅骨和脑膜层会对神经元电流施加空间低通滤波(Nunez & Srinivasan,2006);
高频分量传播距离短
②即使使用 128 通道的 EEG 数 据采集设备,所记录到的头皮信号也仅具有厘米级的空间分辨率;
③由头皮记录估算 EEG 或 ERP 来源是一个逆问题(Schroeder et al.,1995), 会导致使用 EEG 和 ERP 技术检查人脑的功能变化时存在一定的局限性。
其次,相比于微电极记录(microelectrode recording,MER)(Hua et al.,2000)和皮层脑电图(Kuruvilla & Flink,2003)等其他电生理学技术,EEG 技术具有无创性优点。最后,EEG 是对神经元活动的直接测量,其采样率可以高达几万赫兹(Inoue et al.,2001;Ozaki et al.,1998),因此具有极高的时间分辨率。
然而,EEG 技术的空间分辨率较差,这主要由三个因素引起:
①介于大脑和记录电极之间的皮肤、颅骨和脑膜层会对神经元电流施加空间低通滤波(Nunez & Srinivasan,2006);
高频分量传播距离短
②即使使用 128 通道的 EEG 数 据采集设备,所记录到的头皮信号也仅具有厘米级的空间分辨率;
③由头皮记录估算 EEG 或 ERP 来源是一个逆问题(Schroeder et al.,1995), 会导致使用 EEG 和 ERP 技术检查人脑的功能变化时存在一定的局限性。















 140
140











 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









