






文献分享
来自吕一帆老师小组
ATP驱动的瞬态液-液相分离实现可编程DNA凝聚层
本文选自最新发表在Chem上的文章,题目为“Programmable ATP-Fueled DNA Coacervates by Transient Liquid-Liquid Phase Separation”, 文章通讯作者Andreas Walther来自德国弗莱堡大学施陶丁格高分子研究所,其研究兴趣主要包括微观自组装行为与构建功能性仿生材料。已在Nature,Nature Nanotechnology,Nature Communications,Advanced Materials,Angewandte Chemie等国际顶尖期刊发表学术论文150余篇,引用超过11000次,H-Index 55。
01
引言
无膜生物分子凝聚层在生物细胞中普遍存在,其对于实现亚细胞层面时空有序的生命过程十分重要。这些生物分子凝聚层的作用一般是浓缩蛋白质和核酸,同时提供多种生物学功能,例如RNA代谢,核糖体生物发生和信号转导。许多凝聚层具有类似液体的特性,例如快速凝结和溶解,融合和浸润,比如核仁中的核仁,卡哈尔体和核斑点会随时间变化其大小,数量和组成,并且这一变化是可逆的。尽管目前仍然无法确认所有这些动态变化的具体机制,但生物大分子之间的多价相互作用应该是实现动态调节的关键驱动原理。受到这种自然多价驱动的液-液相分离(LLPS)启发,研究人员已尝试将生物分子识别单元编码为成对的工程蛋白,使这些蛋白在混合后表现出相变的能力,进而产生微米级的液滴。
除了生物分子识别以外,许多其他策略,例如电荷-电荷,偶极-偶极,阳离子-π和π-π相互作用等,也已经被用于制备“部分全合成”的凝聚层或复合凝聚层。近年来,这种凝聚层又重新引起了人们的兴趣,因为作为一种人造的无膜细胞器,凝聚层可以像开放式反应器一样方便地进行物质交换。尽管凝聚层在模拟无膜细胞器和原型细胞等研究方面已经取得了很大的进步,但大多数凝聚层的制备都依赖于聚电解质相互作用,利用高度可编程的分子识别制备LPPS却没有太多报道。而实际上,这类高度可编程的分子相互作用对于模型系统的研究大有裨益,包括可以定量地理解LLPS的动力学和热力学,并且为调节动态和选择性摄取提供手段;如果使用正交交互作用,还可以制备空间自分类的动态人工无膜细胞器。此外,结合由化学反应网络驱动的非平衡化学燃料系统这一新兴概念,又可以产生更多的新特性,例如瞬态和可编程的动态行为。
由DNA制成的液滴可以具有出色的可编程性和分子识别性。本课题组之前报道了通过控制反应通路可以形成结构清晰的无膜全DNA凝聚层,具体是通过温度诱导富含polyA的单链DNA发生的LLPS,并通过降温形成液滴。但是这种需要经历退火的制备方法和涉及生物体系的应用场景,例如酶或其他温度敏感成分的包封,并不兼容。因此,利用独特的分子识别技术来构建基于多组分DNA的凝聚层仍是一个尚需探索的领域,利用DNA在动态调节(如可逆的浓缩和解离以及瞬态凝聚)以及等温LLPS方面的潜力,以实现温度敏感性成分的释放或封装(如酶或蛋白质)的可能性也尚未得到充分发掘。
02
成果简介
在本篇文章中,作者报道了一种ATP驱动的无膜全DNA凝聚层,本质是一种基于多价作用驱动LLPS形成的可编程功能化核酸聚合物(SfNAP),同时可以受ATP供能的酶促反应网络(ERN,包括基于T4 DNA连接酶的连接作用和基于BsaI限制性内切酶的酶切作用)调控。通过使用正交聚合的SfNAPs系统(由于在各种结构单元的主链中具有独特的分子识别序列),作者进一步证明了LLPS在同一系统中对多组分凝聚层的自分类作用。为展示该凝聚层的多功能化,作者还展示了DNA凝聚层中微米级胶体粒子和DNA聚合物的瞬态捕获与释放。此外,利用链霉亲和素与生物素之间的稳定结合作用,链霉亲和素标记的葡萄糖氧化酶(GOx)和辣根过氧化物酶(HPR)可以与生物素标记的SfNAP结合并被封装在凝聚层中,从而改善了GOx / HPR酶促级联反应体系的催化活性。
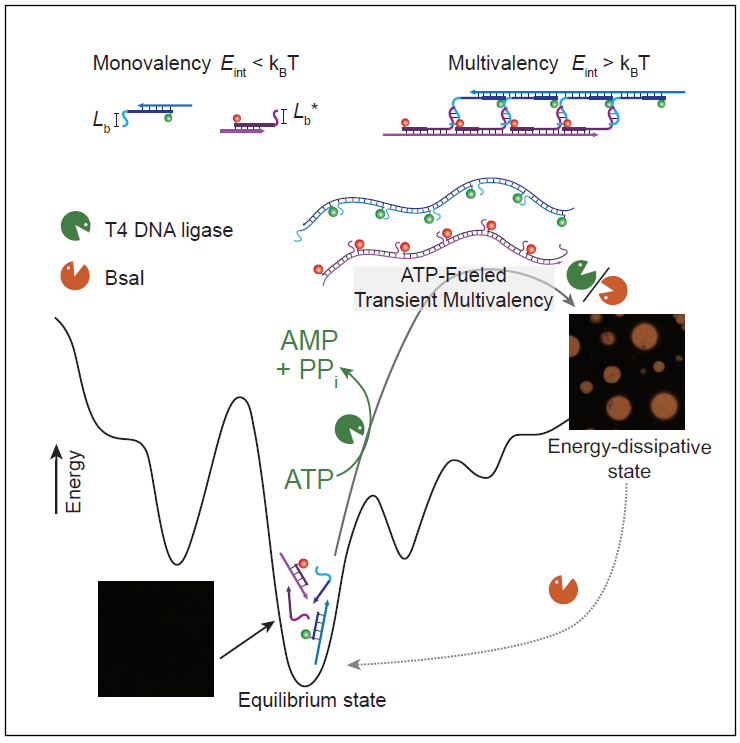
TOC
03
图文解析

图1. ATP驱动的液液相分离:具有互补侧链的多价SfNAP的可编程相互作用。(A)经典凝聚过程中能量分布示意图。(B)ATP驱动的瞬态SfNAP以实现全DNA凝聚层的制备和自分选系统,以及对第二成分的瞬态捕获。
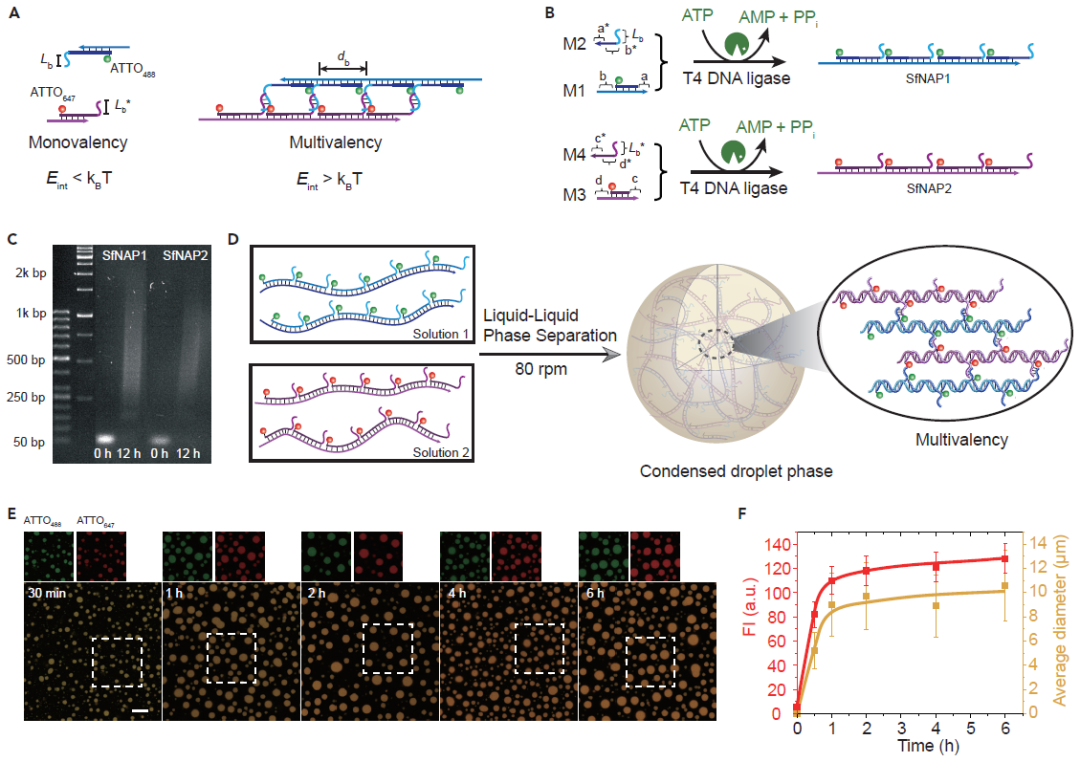
图2. 多价驱动的LLPS用于无膜全DNA凝聚层形成。(A)基于可编程相互作用强度和距离的单价与多价结合。(B)ATP驱动的多个SfNAP示意图。(C)聚合反应起始和结束时的凝胶电泳结果(12h;2wt%,90V,2h)。(D)两个含有互补侧链的SfNAP发生多价LLPS并形成全DNA凝聚层的示意图。(E)DNA凝聚层形成过程的时间依赖荧光成像。标尺:20μm。(F)ATTO647荧光强度以及凝聚层平均直径随时间的变化情况。
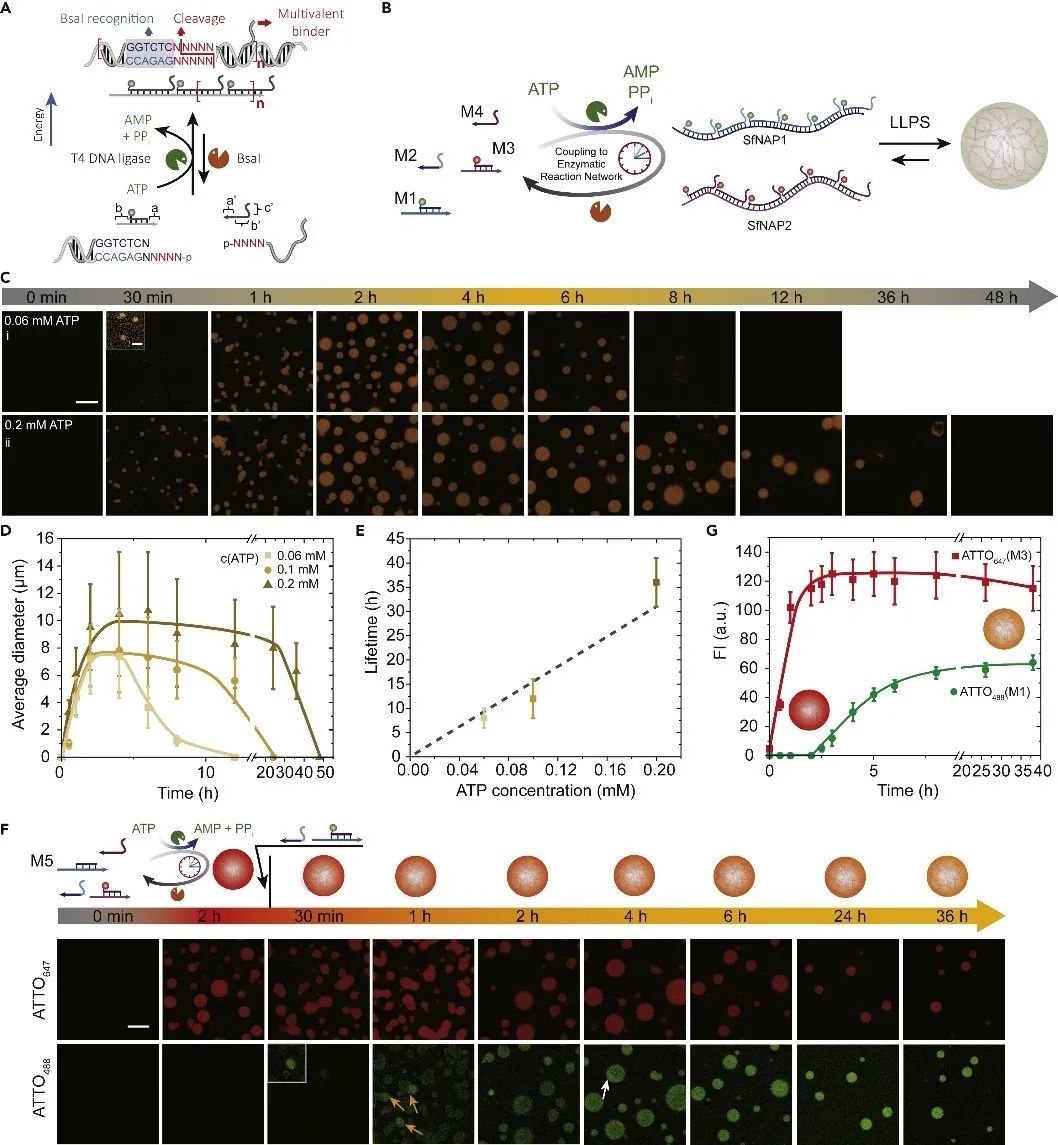
图3. 全DNA凝聚层的可编程寿命。(A)酶促反应网络驱动的SfNAP示意图。(B)正交SfNAP通过瞬态多价效应实现LLPS并形成全DNA凝聚层的过程示意图。(C)受不同浓度的ATP(0.06和0.2 mM)驱动的具有可编程寿命的瞬态全DNA凝聚层:时间依赖性荧光成像。(D)瞬态凝聚层的尺寸与可编程寿命随时间的变化情况。(E)ATP浓度控制凝聚层寿命。(F)运行中的瞬态凝聚系统里tiles的集成和动态化。(G)时间依赖的荧光强度。
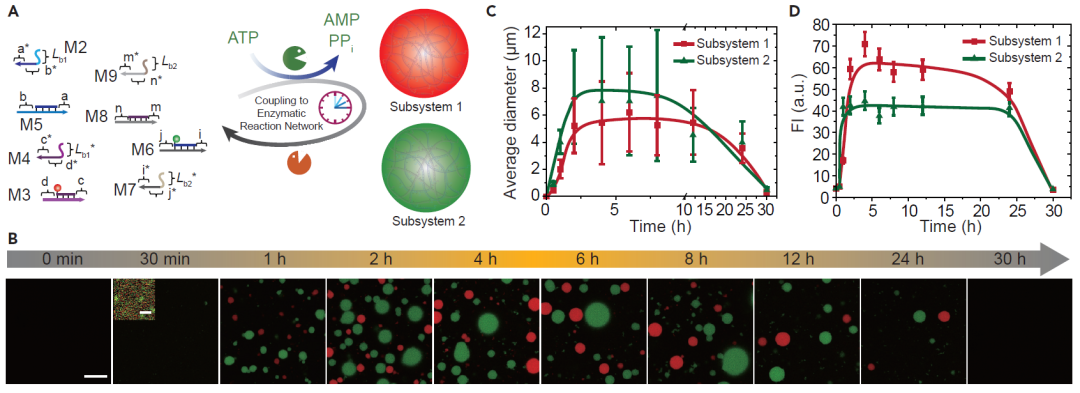
图4.瞬态多组分无膜凝聚层的自分类效应。(A)ATP驱动的多重凝聚系统示意图。子系统1由M2-M5组成,子系统2由M6-M9组成。(B)多个瞬态凝聚层的时间依赖性荧光成像。(C)两个子系统中瞬态凝聚层平均直径随时间的变化。(D)两个系统中瞬态凝聚层荧光强度的随时间的变化。
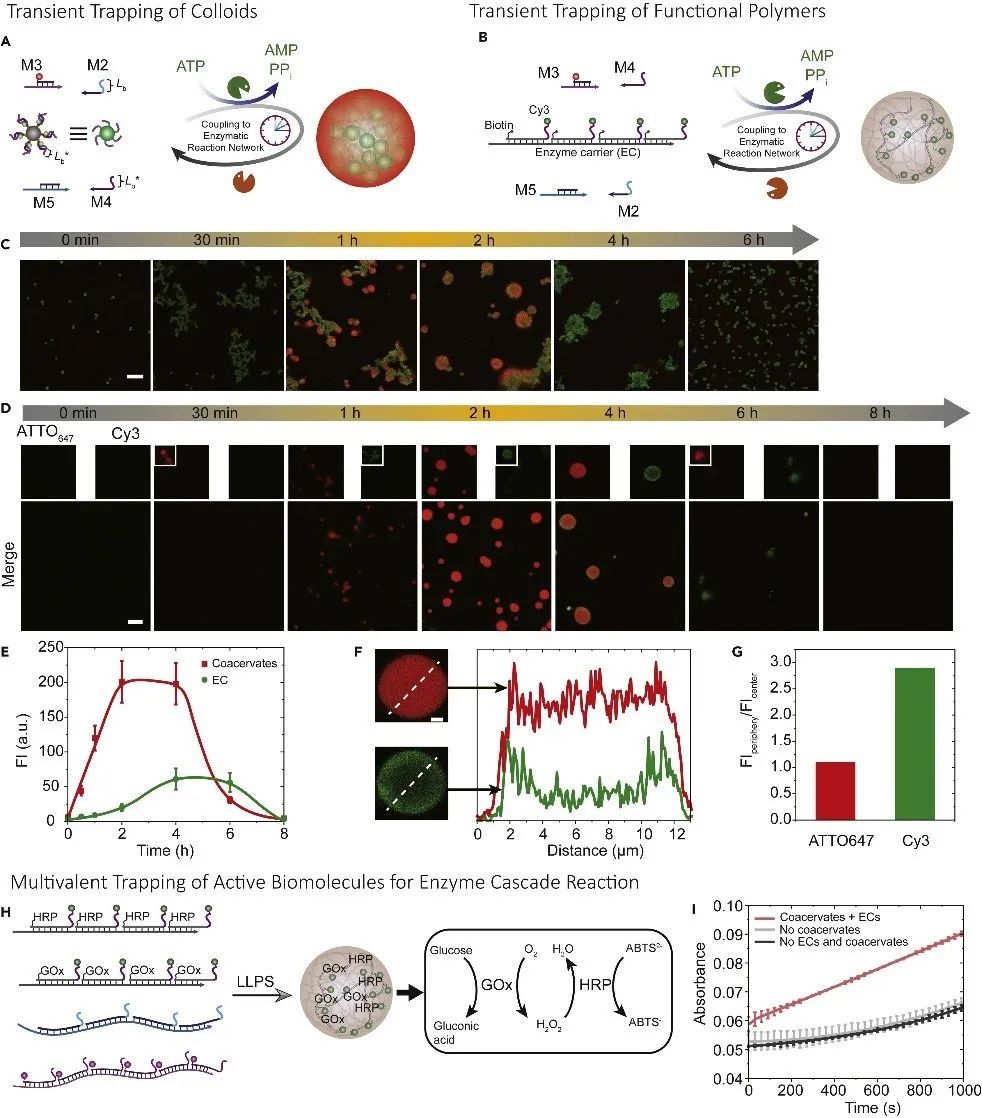
图5.多价驱动的超结构和催化活性凝聚层。(A)可捕获微米级胶体的瞬态凝聚层示意图。(B)功能性DNA聚合物通过多价效应包封进DNA瞬态凝聚层中的示意图。(C)瞬态胶体组装体和瞬态凝聚层的时间依赖性荧光成像。(D)通过瞬态多价效应捕获和释放功能性DNA聚合物的瞬态凝聚层的时间依赖性荧光成像。(E)瞬态凝聚层和包封的荧光DNA聚合物荧光强度随时间的变化。(F)反应4小时后凝聚层和包封的功能性DNA聚合物的荧光强度截面图。(G)凝聚层外围与中心的定量荧光强度比值。(H)GOx和HRP封装进凝聚层中的示意图。(I)封装在凝聚层内部的多酶复合物的催化活性。
总结
这项研究证明了基于程序化核酸聚合物的瞬态多价驱动的液-液相分离(LLPS),可制备功能性全DNA凝聚层。凝聚层需要ATP作为燃料来驱动其形成,并且一旦在燃料驱动的过程中消耗了ATP,该凝聚层就会消失,这是一个具有可调寿命的自治凝聚系统。DNA系统的天然可编程性质允许并行运行两个不同物种的自分类LLPS,以用于多组分凝聚层,并可用于诱捕生物活性物种以产生多种功能。
这项研究首次展示了两个正交SfNAP的多价驱动LLPS用于人工无膜细胞器的制备,并作为理解生物系统中多价驱动LLPS的模型系统。通过将形成SfNAP的结构单元与酶促反应网络偶联,可以使瞬态多价驱动的全DNA凝聚层显示出与ATP水平相关的可编程寿命。通过在不同结构单元的粘性末端和侧链中编码不同分子识别序列,研究进一步证明了在同一系统中针对多组分凝聚层LLPS的自我分选。LLPS期间的分子识别允许基于碱基互补配对实现对功能化组分的多价捕获,从而为封装温度敏感的成分(如酶或蛋白质)提供了可能性。
展望
这种瞬态多价驱动的LLPS对于推动类生命材料,系统化学,合成生物学等诸多研究领域,以及更好地理解生命系统中动态结构的某些原理,提供全新的见解。基于DNA分子的灵活设计特性,其他触发机制应当可以很容易的整合进本文所报道的全DNA凝集层中,从而实现更多更复杂的功能。运用本研究中提出的瞬态多价驱动的DNA凝聚层这一概念,将有助于研究凝聚层内部的生物催化反应,实现蛋白质和核酸的动态浓缩以实现可编程的生物功能,编码具有多层动态结构的人工细胞,并在生物医学工程领域中找到应用。

湖南大学MBL公众号
欢迎关注~














 642
642











 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









