







1. 摘要
三维空间中的所有意志运动都需要多感觉整合,特别是视觉和前庭信号。人类的大脑哪个位置以及如何处理和整合自我运动信号仍然是个谜。在这里,我们应用视觉和前庭自我运动刺激,使用快速和精准的全脑神经成像,在一个相对较大的数据集(n=131)中描述和表征整个皮层和皮层下自我运动网络。我们的研究结果确定了一个核心的自我运动网络,包括扣带沟(CSv、PcM/pCi)、小脑(小脑小舌)和颞顶叶皮层,包括VPS区和缘上回的一个未命名区域。基于其大脑连接模式和解剖定位,我们认为该区域代表了猕猴7A区的人类同源脑区。全脑连接和梯度分析表明,扣带沟和小脑小舌之间的连接在自旋运动感知中起着重要作用。这可能是通过涉及更新视觉空间和前庭信息的反馈回路。PcM/pCi独特的功能连接模式暗示了它在多感觉整合中的中心作用,以感知自我参照的空间意识。所有皮层自我运动中枢都表现出与其他视觉、前庭、体感和更高阶运动区域的模块化功能连接,强调了它们在一般感觉运动整合中的相互功能。
2. 引言
视觉和前庭线索是感知自我运动中必不可少的。这种本质上的多感觉过程需要从视觉到前庭、体感和运动系统的各种模式的有效整合。
早期神经影像学研究的重点主要是利用视觉光流研究参与处理视觉自我运动信息的皮层区域。几个皮质层域发现了反应,包括扣带沟视觉区(CSv)、颞中视觉区(hMT)、内侧颞上回(MST)、腹侧顶叶内区(VIP)、后丘视觉区(VPS)、外侧顶叶内沟(IPS),V6复合体、楔前运动区(PcM)或后扣带区(PCi)和假定区2v。然而,这些区域是否也对前庭刺激有反应,并形成多模态自我运动网络的一部分,目前尚不清楚。一项使用前庭电刺激(GVS)的研究(n=9)报道了CSv、MST、STSms的前庭反应和VIP的“弱”反应,但只关注这些特定区域。
此外,脑干和小脑中枢在人类自我运动感知中的参与尚未被研究。在猕猴中,小脑小舌和结节被认为是自我运动加工的关键区域。它们在前庭加工中的关键作用是通过来自耳石器官和半规管的初级前庭传入神经和来自前庭核和下橄榄的次级传入神经到达小舌和结节。此外,在人类中,可以显示出一种眼球运动的功能。此外,小舌和结节的病变损害了在噪声背景下感知视觉运动方向的能力,导致头部运动的感知阈值升高。
在这里,我们系统地研究了在一个大数据集(n=131)中使用视觉和前庭刺激研究全脑高分辨率神经成像的人类自我运动网络。我们试图通过在三种不同条件下对视觉自我运动兼容刺激的反应来描绘这个网络的多模态中心——睁开眼睛和注视一个点,在黑暗中睁开眼睛和闭上眼睛。在一种独特的多模态方法中,我们进一步通过功能连接、功能连接映射和与核成像衍生估计的相关性来表征该网络。
3. 方法
3.1 被试
共招募了131名健康被试(70名女性,61名男性,平均年龄26.4岁,年龄范围23-35岁)。被试被分为三个组。46名被试(23名女性,23名男性)参加睁开眼睛的视觉刺激实验和前庭电刺激(GVS)实验(组1),42名被试(21女性,21男性)在GVS实验时在黑暗中睁着眼睛(组2)和43名被试(22女性,21男性)在GVS实验闭着眼睛(组3;实验的详细描述见3.3节)。所有被试均给予了书面的知情同意书。我们使用爱丁堡测试的14项清单确定了利手性。所有被试均为右撇子,视力正常,既往无神经学或眼部运动障碍史,也无常规药物治疗。该研究根据《赫尔辛基宣言》(2013年)进行,并得到了当地伦理委员会的批准。被试获得了参与活动的报酬。遵循fMRI研究的指南和原则。
3.2 功能磁共振成像实验
实验是在3T核磁共振扫描仪(Skyra,西门子,德国)中进行,使用回声平面成像(EPI)T2∗加权梯度回波序列(TR= 700 ms,54个连续轴向切片覆盖整个大脑,2.5mm各向同性体素,视场为210mm2,TE=33 ms)。在功能磁共振成像任务之前,使用相同的多波段EPI序列(600卷)获得了静息态数据,被试在相同的设置中注视着圆点。获得了矢状面的高分辨率T1加权MPrage序列(TR = 2060 ms , TE = 2.17 ms,翻转角度为12°视场为 240 mm2,切片厚度为 0.8 mm,A-P相位编码,0.8 mm各向同性体素)。
对于视觉刺激的投影,使用了一个位于被试头部后面的监视器和一个可调节的镜盒,反映了头部线圈在观看距离为16cm的图案。用红外VOG单元(MRI兼容相机)记录眼球运动。扫描房间和扫描仪隧道都进行调暗处理。被试戴着耳塞和一个可充气的头垫。
3.3 任务和刺激
fMRI实验包括两种视觉刺激条件,自我运动相容(EC)和自我运动不相容(EIC),以及三个GVS阶段(睁开眼睛,在黑暗中睁开眼睛,闭上眼睛)。在视觉刺激和睁眼GVS中,采用了随机区块设计,被试注视屏幕中央的一个点。
EC刺激是一个高对比度的,由360个光点组成的单光流补丁(图1)。这些点围绕着一个中心连贯地移动,一个连续存在的注视点以顺时针或逆时针方向旋转。对于EIC刺激,显示了9个大小相同且分布于视野的光流补丁,每个都包含与EC刺激相同的运动轮廓。两种刺激均以随机顺序交替显示,视觉休息条件的区块长度不同(视觉刺激的区块长度为10 TR(7s),每种条件重复20次,视觉休息的区块长度为10-19 TR(7-13.3s),1005卷(11.73 min))。所有条件下都包含一个位于屏幕中间的注视点。使用功能定位器来描述MST区域的反应。
在耳后区域局部麻醉(区块长度为10 TR(7s),休息条件为13-13TR(1.1-16.1s),1013卷(11.82分钟))后,通过双窦电极进行GVS。在三个不同的GVS阶段(睁开眼睛,黑暗中睁开眼睛,睁开眼睛)中刺激是相同的。在同时接受视觉和前庭刺激的组中,刺激以随机顺序呈现,两个任务之间的最小时间间隔至少为30分钟。所有被试都使用标准化的问卷对自我运动强度进行评分,其中自我运动强度必须以从0(根本没有自我运动)到10(强烈的自我运动感知)的量表进行评分。
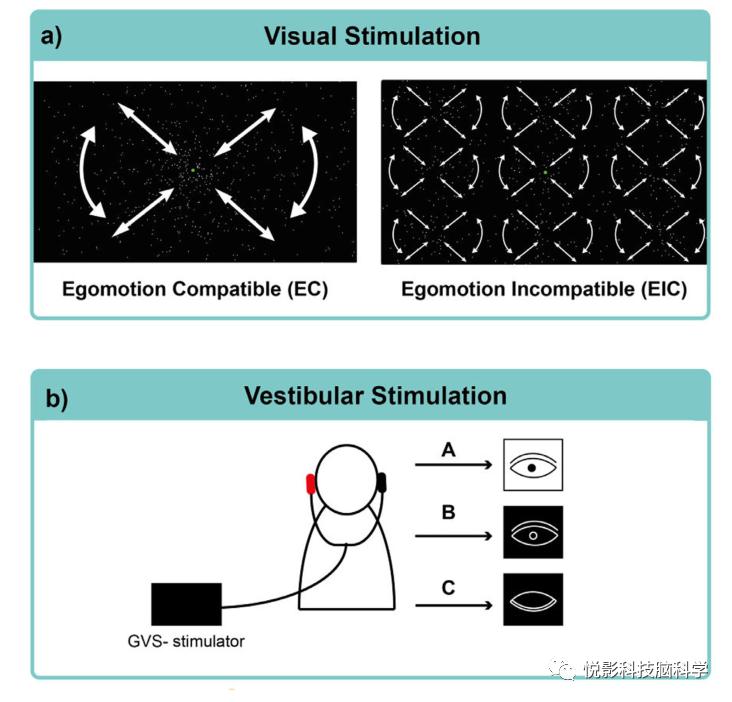
图1. 实验设计。(a)左:视觉自我运动兼容刺激(EC,左),一个由360个白点组成,或顺时针/逆时针旋转。右:运动不相容刺激(EIC,右),9个光流补丁大小相等,分布在整个视野,每个都包含与运动相容刺激相同的运动轮廓。(b)前庭电刺激(GVS)通过电极在每个乳突上应用,使用0.875Hz的交流电刺激模式(3 mA)于三种条件,包括睁开眼睛,注视着一个点(A,组1),在黑暗中睁开眼睛(B,组2),或闭上眼睛(C,组3)。
3.4 基于任务的功能磁共振成像的数据分析
统计分析使用SPM12,运行在Matlab 2018b。通过DARTEL算法,将使用现有的MNI模板和蒙特利尔神经学研究所进行立体定向归一化到定义的标准解剖空间。图像用各向同性的4 mm高斯核进行平滑处理。将重新调整参数和一个高通滤波器(128 s)集成到设计矩阵中。统计参数图(SPMs)是在体素水平上生成的,并使用了扫描期间刺激期间的血流动力学模型。计算每种刺激条件下与各自休息条件下的单个被试的t值。对条件图像进行二级统计分析。对视觉刺激和GVS实验的被试进行了单向方差分析。使用双样本t检验来比较不同GVS条件(睁开眼睛,黑暗中睁开眼睛,闭上眼睛)下的EC>EIC对比。在三个不同的组中,对比EC>EIC与各自的GVS条件进行了联合分析。在10 000次置换后,p < 0.001处被认为是显著的。
3.5 眼动轨迹数据分析
视觉运动实验的眼球运动视频以未压缩的AVIs形式进行搜集。除了AVI文件之外,还收集了一个包含每个视频帧的时间戳和fMRI运行中每个卷的开始时间的文本文件,以将视频与功能数据同步。采集一个五点校准视频,将眼睛的位置转换到空间。这些视频被切割到fMRI序列的长度,并转换为使用EyeSeeCam软件进行进一步分析所需的文件类型。所得到的眼睛位置数据与每种情况下的帧一起保存,并用Matlab(版本2018b)进行分析。通过将眨眼前50 ms到眨眼后50 ms的时间点设置为nan,从眼球追踪数据中去除眨眼。然后用高斯低通滤波器对数据进行线性插值和低通滤波。检测扫视,并以扫视期间眼睛位置的变化作为注视期间眼睛的稳定性。然后用配对Wilcoxon符号秩检验和𝛼= 0.025的多重比较校正后的alpha来检验两种情况下的扫视率和注视稳定性的差异。
3.6 功能连接数据分析
ROI是使用任务实验中描述的区域(阈值TFCE FDR p<0.001)来定义的,用于功能连接(FC)分析。分析使用静息态fMRI数据和所有扫描的任务数据进行分析,使用CONN工具箱的Matlab(第2019a版)。白质(WM),脑脊液(脑脊液)信号和所使用的运动参数的六个被回归以去除噪声,使用带通滤波器(0.008-0.09 Hz)进行滤波。所有任务区块的起始时间和持续时间与基于spm12的GLM方法相似,分别分析包括剩余任务在内的不同任务的ROI-ROI网络结构,并研究任务与其余条件下的网络结构的差异。经过非参数阈值的5000次置换后,所有结果的阈值均为p<0.05(FWE校正,TFCE)。
3.7 与核成像得出的跨模态相关性估计
空间关联核成像的特定受体系统的分布来源估计,包括不同ROI(CSv,PcM/pCi,VPS,h7a,小舌)的静息态FC和基于任务的FC,空间关联计算使用JuSpace工具箱。
3.8 功能连接映射
使用先前定义ROI,我们使用Congrads工具箱(https://github.com/koenhaak/congrads.git)检查了所有自我运动网络hub的功能连接映射。使用PALM(线性模型置换分析)检验空间模型参数的显著性,在10 000次置换后,阈值为p<0.05(FWE)为显著。
4. 结果
4.1 基于任务的分析
通过对比EC>EIC定义了对自我运动兼容的特异性反应,并在视觉皮层(V1、V2、V6区)和所有自我运动中心(CSv、PcM/pCi、h7a、VPS、小舌)中发现。其他反应区域包括左侧OP3,双侧Id7区,右侧6mc区其包括辅助视野(SEF)。小脑幕下的反应包括小舌、结节状和小叶VI(图2)。
这两种情况下的眼球运动没有发现差异。2v、3av、MT+、MST和VIP的反应在对比运动兼容刺激中看到,但也出现在运动不兼容刺激期间,因此不是视觉运动感知的特异性。前庭刺激在所有运动中枢、岛叶和顶叶盖(OP1-4)、hIP1/3区、MST前颞中回、副扣带回(包括前SMA、SEF)、前扣带回、壳核、丘脑、丘脑、中脑、双侧前庭核、小结节小脑蚓部、HIV-VI、小腿I部、HVIIIA被激活。
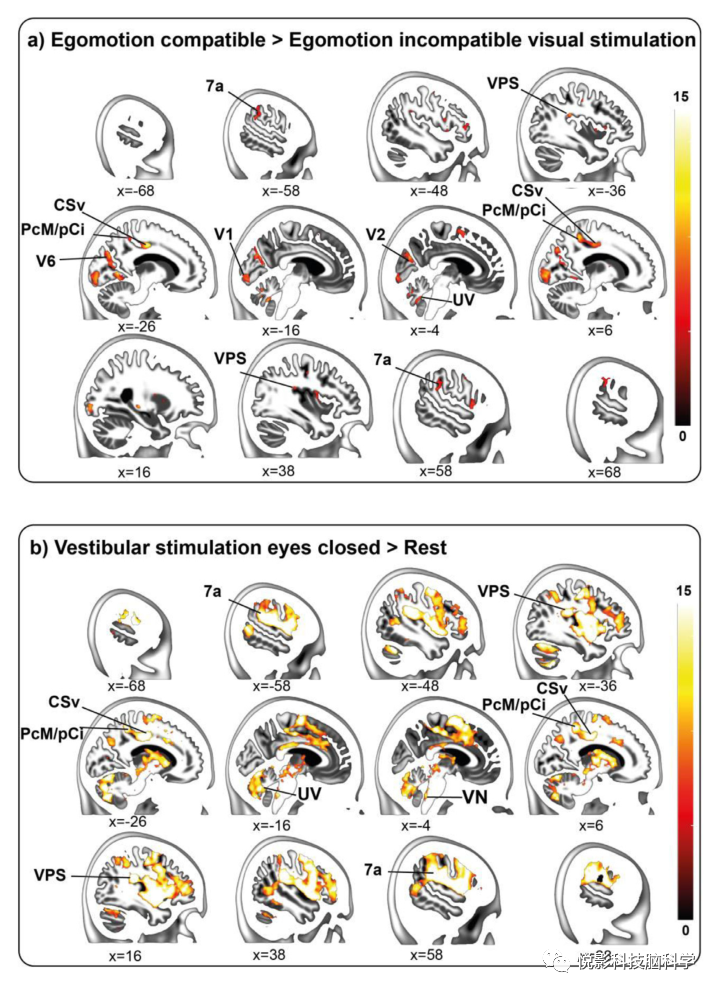
图2.对视觉自我运动刺激和前庭刺激的反应。(a)对自我运动兼容刺激的特异性反应(对比EC >EIC)在h7a、VPS和视觉皮层(V1、V2、V6)以及CSv和PCM/pCi(组1,n=46)中发现了强烈反应。闭上眼睛的(b) GVS导致了整个自我运动网络的反应,包括7a、VPS、CSv、PcM/pCi和小舌(组3,n=43)。
4.2 联合分析
在EC>EIC和GVS与闭眼的联合分析中,常见的反应定位于小舌、CSv、PcM/pCi、h7a和VPS。较小的簇包括右侧SMA(SEF)、右侧中央前回、右侧壳核和双侧44区。小脑的幕下反应包括双侧小舌、小叶VIIb和HVI(图3)。在黑暗中睁开眼睛的EC>EIC和GVS的结果与闭上眼睛的EC>EIC和GVS的结果相似。与GVS相结合,在顶叶-枕沟(V1、V2、V3d、V3、V6)、双侧前岛、丘脑发现了闭眼GVS的额外反应。
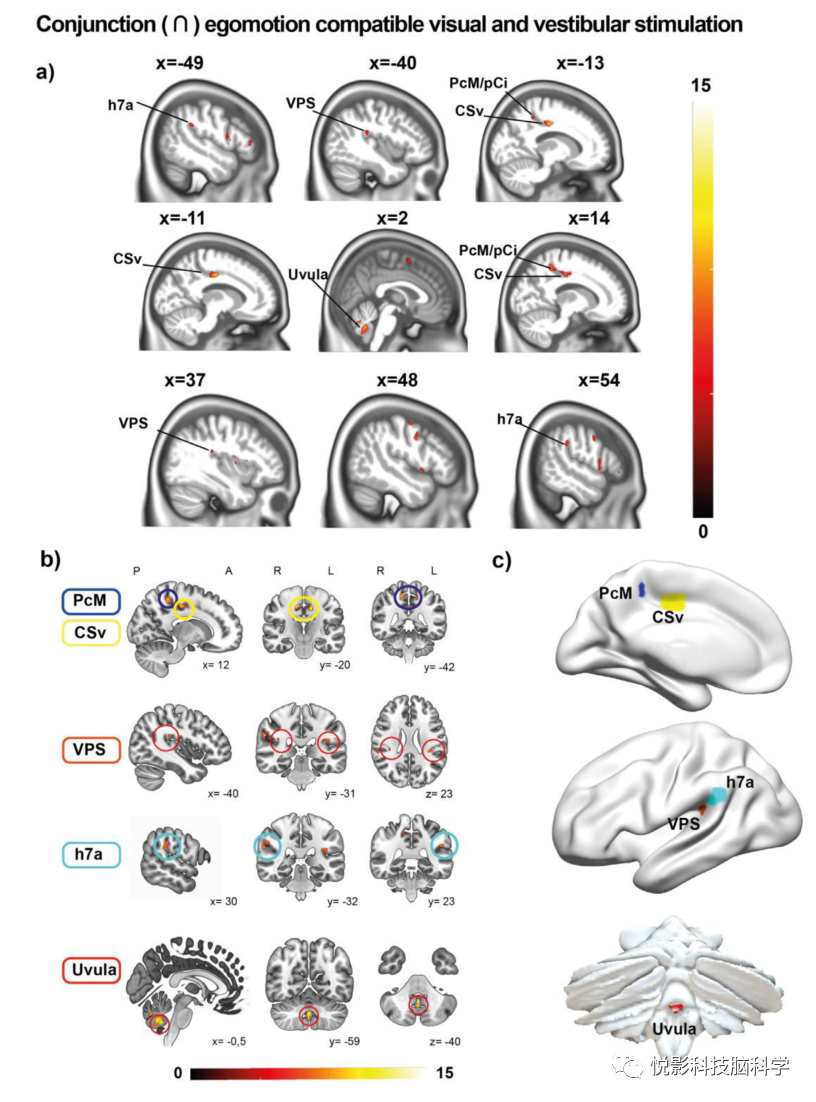
图3.人类自我运动网络。定义人类自我运动网络(EC>EIC和GVS)与中心五个自我运动核心枢纽的连接分析结果概述。(a)与视觉自我运动兼容刺激(EC>EIC)和闭眼前庭刺激的联合分析结果的矢状面切片视图。人脑7a区和VPS的双侧颞顶叶皮层、CSv和PcM/pCI的扣带回和回以及小脑小舌的信号增加。(b)五核心枢纽矢状面、冠状的定位,包括扣带沟的PcM/pCi(蓝色)和CSv(黄色),小脑的小舌(红色)和VPS区(橙色)和颞叶的h7a(浅蓝色)。
4.3 心理物理学结果及与功能磁共振成像数据的相关性分析
所有被试在EC任务中都感知到自我运动。在睁开眼睛的GVS(n=46),被试感知自我运动的平均强度为6.13(SD 1.99;0-10, 0=无运动知觉,10 =强烈运动知觉),在闭眼的GVS(n=43),平均强度为5.84(SD 1.87)。将EC和GVS期间感知的运动强度与基于任务的功能磁共振成像数据相关联,并没有给出显著的结果。
4.4 眼动轨迹的分析结果
在注视点稳定性(通过眼睛位置的变异性测量)(z=0.6376,p=0.5237,Wilcoxon符号秩检验)和扫视率(z = 0.7287,p = 0.4662)的自我运动相容和自我运动不相容的刺激比较中没有发现显著差异。
4.5 功能连接分析
4.5.1 h7a区的静息态和任务态的功能连接
h7a区显示和所有皮层自我运动中心的FC,包括视觉(-运动)区域(V1、V2、V3A、V4、V6、MT/V5、MST部分)和多感觉区VIP和hPEc,腿表征区域hPE和S1leg,双侧运动前皮层(6d2、SEF)和顶叶盖包括前庭核心区域OP2。
在自我运动相容的视觉刺激中,双侧h7a 与CSv、PcM/pCi、同侧额视野(FEF)、SEF显示出FC。在前庭刺激期间,h7a均显示FC随着幕下前庭区(小舌、右侧小脑扁桃体)、CSv区、前庭区、结节区和右侧h7a的反应增加而增加(图4)。
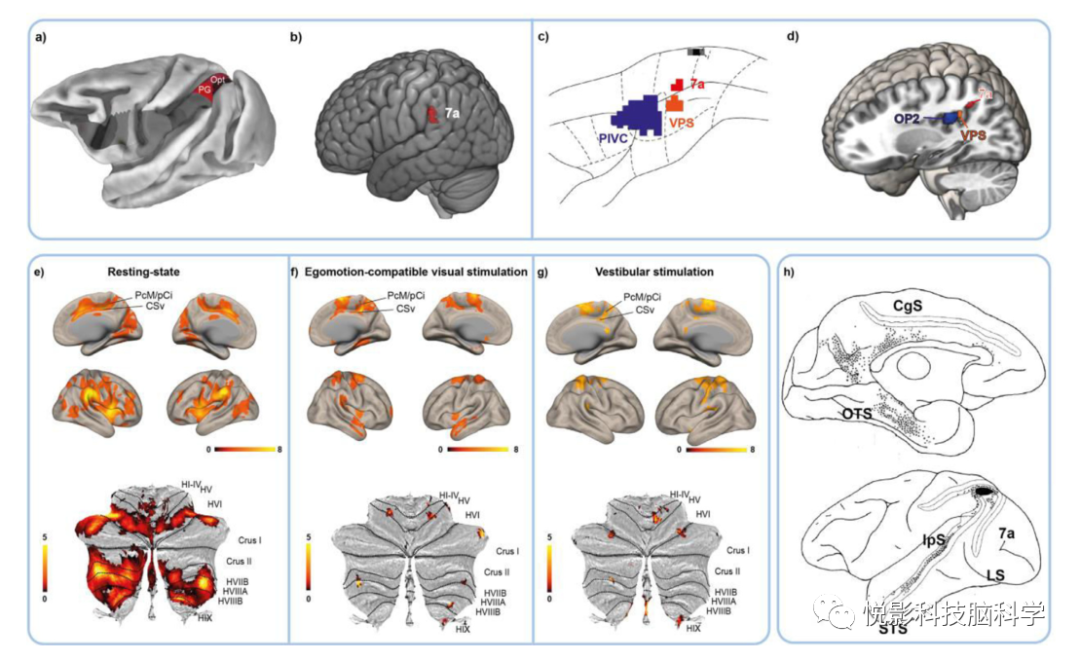
图4.h7a区与猕猴7a区相比的定位和连接。猕猴和人类7a的定位a)-d)和功能连接的比较。(a)猕猴7a、PG和Opt区(红色)可能的细胞结构相关示意图。(b)功能连接分析自我运动兼容的视觉刺激和GVS的边缘上回的反应。(c)皮层前庭区PIVC、VPS和7的定位。(d)在假定的人类VPS和人类7a区与OP2相关的多模态自我运动反应与非人类灵长类动物的结果重叠,如c)所示。(e) H7a显示RS-FC具有所有皮层自我运动中枢,具有视觉(-运动)区域、多感觉区(pVIP#1,hPEc)和腿部表征区hPE和S1leg、双侧运动前皮质和顶盖前,包括前庭核心区OP2。在小脑,底部部分)发现了双侧小叶VI、VIIa、VIII、IX和小腿II的功能连接。(f)在视觉自我运动刺激期间,FC随双侧CSv、PcM/pCi、同侧额视野(FEF)、双侧SEF、前sma和双侧簇增加,局部达到相关反应。右侧h7a使左侧OP2的FC额外增加。在小脑(底部),左侧小腿III、右侧VIIIa、双侧小叶VI和小脑扁桃体FC增加。(g)在前庭刺激过程中,双侧CSv区、右侧PcM/pCi、前庭区OP2和幕下前庭区(小舌、右侧小脑扁桃体)以及双侧小脑小叶VIII、I、HVI小叶FC增加。(h)Cavada等人的示踪剂研究结果显示了猕猴7a区的部分连接。这些结果与h7a(e)-g)的FC图有很好的重叠,包括扣带回(扣带沟下方,CgS)、枕颞沟(OTS)上方、颞上沟(STS)。
4.5.2 小舌的静息态和任务态的功能连接
小舌伴有蚓部和前庭-小脑中枢(小凝体、扁桃体)、前庭核、动眼肌核、左侧臂旁复合体和左侧蓝斑。这些与觉醒/自主功能脑干中枢的联系与动物文献一致,表明小脑蚓部参与心血管控制。在皮层水平上,没有发现任何自我运动中枢的RS-FC,但FC随着右侧OP2的增加,双侧视觉区域(V1、V3v)和左侧杏仁核的增加。
在自我运动兼容的视觉刺激过程中,与CSv、PcM/pCi、视觉区域(V1、2、V3v、V3A、V6)、FEF和前庭小脑区(小叶、结节状)显示出FC。前庭刺激使双侧PcM/pCi和双侧CSv、视觉区域(V3d、V3A、右V6、右背侧hMT+)、杏仁核和小脑蚓部(小叶VIIb、VIIIb、结节状)的FC增加。
4.5.3 CSv的静息态和任务态的功能连接
CSv与小舌的FC最强,其次是前庭小脑中枢(小叶体、小脑扁桃体)和前庭核。CSv显示与所有皮层自运动枢纽显示出FC,包括顶盖(OP1、OP2、OP4、OP8)、扣带视野(CEF)、SEF、FEF、视觉区(V1、V2、V3A、V4、hMT+、V6)以及辅助运动区(SMA)、preSMA和运动整合区(hPEc、hPE)。在自我运动相容性视觉刺激期间,FC随着PcM/pCi、对侧CSv和小舌、所有视野(FEF、CEF、SEF、同侧顶叶视野(PEF))、双侧视野(V1、V1、V3d、V3A、V6)和OP2后侧的簇而增加。在GVS期间,CSv的FC随着PcM/pCi、右侧h7a、小舌、SEF、海马和双侧杏仁核的增加而增加。左侧CSv随右侧VPS、左侧OP1和OP2而增加。
4.6 PcM/pCi的静息态和任务态的功能连接
PcM/pCi与对侧PcM/pCi,双侧CSv和h7a显示出FC。FC随着视觉区域(V1、V2、V3A、V6)、SMA、前SMA、SEF、CEF和视觉运动整合区(hPEc、HPE)、顶盖(OP1、OP2、OP4、OP8)、同侧海马旁回、海马和前庭小脑区(小叶、小脑扁桃体)的增加而增加。
运动兼容的视觉刺激使对侧区域PcM/pCi、CSv、h7a和VPS的FC增加。随着视觉区域(V1- V6)、视野(FEF、SEF、同侧CEF)、SMA/pre-SMA、前庭核心区OP2、OP1、双侧海马、同侧杏仁核和小叶,FC进一步增加。前庭刺激导致双侧CSv、同侧h7a、小舌、双侧前庭区OP2、右侧OP1、前同侧MST和SEF的FC增加。
4.6.1 VPS(aPIC)的静息态功能连接
人类VPS显示出包括所有自我运动枢纽的双侧FC,包括视觉区(V3A、MT/V5)、OP1、3、4区、前庭区OP2、CEF、SMA、preSMA、海马和杏仁核。在自我运动相容性视觉刺激期间,对侧VPS、对侧PcM/pCi和h7a之间以及与视觉区域(双侧V1、双侧V侧V2、V3d、V6)、双侧SMA、SEF和对侧HVIIIa之间的FC增加。前庭刺激并没有导致VPS区FC的增加。
4.6.2 自我运动网络的ROI-ROI的功能连接
对比EC > EIC显示小舌与PcM/pCi、CSv和右侧h7a之间的FC增加(图5)。对比EC >休息显示PcM/pCi与所有roi之间的双侧信号增加,除左侧VPS外,只有右侧PcM/pCi显示FC增加。在GVS期间,FC在PcM/pCi和CSv、小舌膜、右VPS和右侧7a以及PcM/pCi、右侧PcM/pCi和左侧VPS之间的两侧增加。此外,小舌与左侧h7a、左侧VPS和右侧h7a之间的FC增加,但在两个h7a之间下降(图5)。
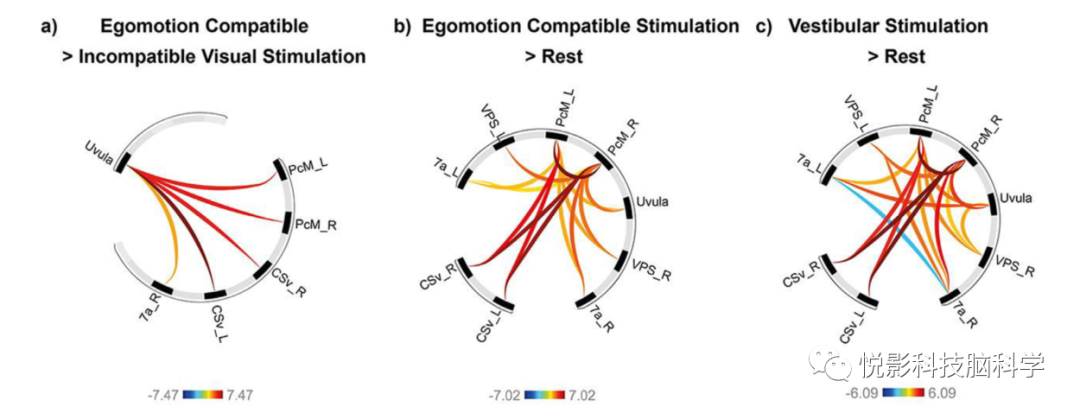
图5. 自我运动网络的ROI到ROI的功能连接图。(a)运动特异性视觉刺激(EC>EIC)导致小舌和PcM/pCi、CSv和右侧7a之间的FC增加,表明在视觉运动感知中,小舌和扣带沟轴之间的皮层-小脑幕下的中心作用。(b)自我运动兼容刺激与休息条件相比,c)前庭刺激导致PcM/pCi双侧和所有其他自我运动枢纽之间的FC增加,表明PcM/pCi作为多模态整合枢纽的独特作用。
4.6.3 与核成像结果的跨模态相关性估计
在静息态中,不同皮层自我运动中心的FC图与5-羟色胺能-(5HT1a、b、2a)GABAa-、阿片-(MU)和去甲肾上腺素(NAT)受体图的表现出显著相关。基于任务的FC图显示,在EC期间,所有枢纽与5-羟色胺能(5HT1b,5Ht2a)和GABAa受体存在显著相关性,而在GVS期间,5-羟色胺能(5HT1b)和NAT受体存在相关性。
4.6.4 功能连接映射
小脑小舌的连接映射分析显示有一个显著的梯度(图6c),主要是前后方向。对两个区域CSv的平均连接点的分析给出了相似的模式(图6d)。CSv在其梯度组成中描绘了一个前后轴和外侧到内侧轴的镜像(图6a)。这些梯度的性质目前还不清楚。对于所有其他的自我运动枢纽,没有发现显著的梯度结构。
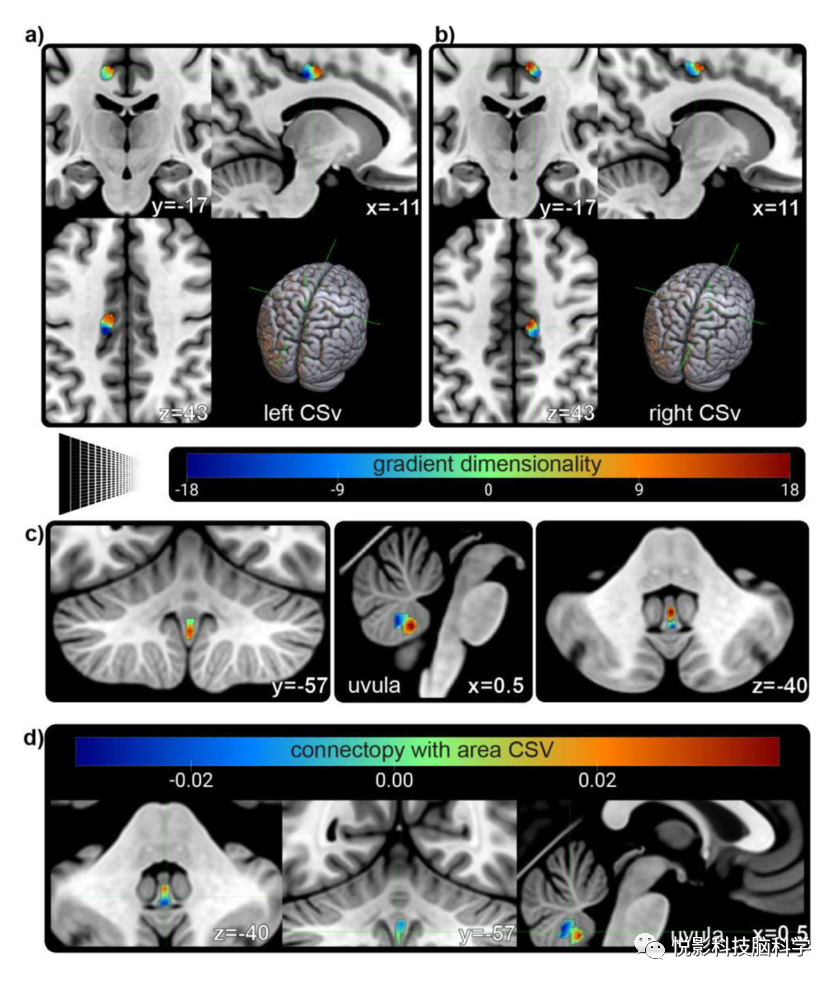
图6. 自我运动网络枢纽的梯度。(a)使用与所有其他大脑区域的连接映射来描述在左侧区域CSv中发现的显著梯度。主轴是前后的梯度和外侧到内侧的梯度。检测空间模式似乎是双极的,有一个小的过渡区(绿色)。(b)右侧区域CSv的空间连接模式在其梯度表征和方向上稳健地反映了其同伦区域。(c)在连接组分析中,发现小脑小舌包含显著的前后梯度。梯度的潜在性质仍有待研究。(d)小舌与两个CSv区域的平均连接模式与c)中检测到的一般梯度相似的空间排列相似。
5. 讨论
我们的结果显示了5个自我运动核心枢纽:CSv区、PcM/pCi、VPS,细胞结构区域PF/PFcm区域,代表猕猴7a区的人类同源脑区,以及小脑小舌(图3)。FC模式突出了这些中心与视觉、前庭、体感和(眼)运动区域的多模态相互连接。我们的结果表明,所有枢纽的共同神经递质基础和显著的梯度只在小舌和CSv。所有皮层自我运动网络枢纽与SMA和前SMA的功能连接强调了它们参与了运动计划。
我们的数据点将CSv作为处理自我运动信息的中心多感觉界面,特别是在有视觉线索存在的情况下。因此,我们基于任务和功能连接的结果补充了先前报道的CSv在视流、眼运动和前庭处理中的功能。与早期的研究相比,我们发现了与主要视觉和视觉运动区域(V1、V3A、V6)的功能连接的明确证据。这些可能代表了将视觉输入传递到CSv区域的路径,然后可能在自我运动网络中提供视觉感觉和动眼肌信息。在GVS期间,CSv的功能连接模式从视觉运动区转向自我运动区、SEF和OP2,支持了CSv作为视觉感知、(眼)运动和前庭/导航相关区域之间的重要中介枢纽的作用。
特别是扣带沟(CSv,PcM/pCi)和小舌之间的功能连接似乎在视觉自我运动知觉中起着重要的作用。与CSv/PcM的连接可能会重新融合皮层-小脑环路,参与多模式自我运动信息的不断更新,以协调视觉-空间信号和更高阶的运动行为。与其他运动枢纽相比,CSv和小舌独特的连接梯度进一步证实了它们在运动网络中的重要作用。
我们的研究结果表明,小舌是小脑自我运动核心枢纽,从非人类灵长类动物扩展到人类和早期关于慢性小脑中线退行性变患者自我运动知觉改变的报道。通过其多模态的功能连接,小舌可以通过反馈回路向皮层枢纽提供无意识的前庭自我运动信息。
CSv附近的信号增加对应于区域PcM/pCi。它对自我运动兼容的光流刺激、自主头部和腿部运动的反应表明它在感觉运动整合和运动指导中发挥作用。我们发现的多模态反应及其独特的与所有自我运动枢纽的功能连接使PcM/pCI成为多模态整合和自我参照空间意识的理想候选者。PcM/pCi位于扣带沟/楔前叶的边界,与颞顶叶连接,可参与其他自我运动枢纽提供的视觉和前庭信息的空间编码。在两种刺激方式中,PcM/pCi是唯一显示与前庭区OP2双侧连接增加的枢纽。这些连接可能向前庭核心区提供与自我运动相关的信号。在视觉自我运动刺激中,FC随着海马体的增加,进一步支持了自我中心记忆加工和空间更新的作用,如先前的假设正。有趣的是,在CSv和h7a区也发现了与海马体的功能连接的增加。研究它们在空间记忆加工中的作用可能是未来研究的目的。
在双侧缘上回中发现了对自我运动刺激的多模态反应。我们认为该区域对应于猕猴7a区域,迄今为止尚未在人类中发现。
猕猴7a位于颞上沟和顶叶内沟之间,称为细胞结构区PG。然而,最近的一项研究将猕猴7a定位在下顶叶的尾侧部分(图4)。通过比较结构观察,提出了角回中PG区域的人类同源脑区。我们的数据表明,人类7a区(h7a)定位于缘上回的前部,覆盖部分细胞结构区PFcm和PF。猕猴7a对视流和前庭刺激有反应,其中前庭在自我运动加工中占优势,只有一小部分神经元参与多感觉加工。这与我们在前庭刺激期间h7a中更大的激活簇和更高的t值一致,并且在EC刺激过程中,FC的缺失随着视觉区域的增加而增加。
最近的一项人类运动平台研究表明,在自我运动过程中,缘上回的一个区域的活动有很强的调节作用,可能代表h7a。这里的反应取决于头部和眼睛的位置或平移运动方向的一致性。有趣的是,在猕猴7a的反应也受眼睛和头部位置的调节,这表明它在构建“以世界为中心”的空间表征中发挥了作用。此外,我们的FC分析结果与7a区动物示踪研究结果强烈重叠(图4),证实了我们的假设。特别是它与前庭核心区域OP2的静息态和基于任务的功能连接信号嵌入前庭网络,并与松鼠和爪哇猴的示踪研究一致,显示了7a到PIVC的强皮层输入。我们的结果进一步表明h7a在协调肢体运动准备和运动计划指导运动行为和运动,基于任务的结果发现前运动区域,视野,前楔前叶区域到视觉运动整合区FC增加。
人类VPS区(前后岛叶皮层,aPIC)是发现对多模态自我刺激有反应的中枢。此前显示的对前庭刺激、自主头部运动和自主腿部运动的反应表明其在多模式感觉整合中发挥了作用。前庭刺激过程中VPS、OP2和h7a的反应与松鼠猴的数据完全一致,其中VPS位于前庭核心区PIVC后方,位于7a区前方。这些发现进一步证实了我们对人类区域7a的定位,以及OP2区域作为PIVC区域的人类同源物的作用。FC模式表明VPS在视觉自我运动感知过程中主要参与,FC随着视觉区域(V3A、MT/V5)、h7a和PcM/pCI的增加而增加(图4)。
在MST和VIP中没有发现视觉自我运动特异性反应,因为这两个区域在自我运动相容和不相容的刺激时都有反应。最近在动物身上的发现提出,前庭信号可能通过增强可分离性来促进MSTd神经元的运动处理。这可能解释了MST中的前庭反应,它可以区分物体运动和自我运动,但对自我运动感知没有本质上的贡献。VIP中缺乏对视觉自我运动加工的特定反应,这与最近的一项研究相一致,即VIP在物体-运动加工中具有主要功能。我们的研究结果支持VIP和MST作为运动检测系统的一部分的观点,但没有对自我运动刺激的唯一反应。
我们发现所有枢纽的在EC刺激任务时FC与血清素能(5HT1b,5Ht2a)和GABAa受体表现出显著相关,这与前人的研究中显示直接影响5HT激动剂在动物视觉处理和GABAa受体结合潜力和人类视觉皮层活动关系的结果相呼应。GVS与5HT1b和NAT受体的相关性与动物数据一致,显示前庭核复合体中去甲肾上腺素能和血清素能反应。我们的数据表明,这些神经递质在前庭诱导的自我运动感知中发挥作用,然而,皮层前庭网络的神经递质结构迄今尚未被研究。
6. 结论
总之,我们的研究结果显示,人类的自我运动网络从扣带沟(Csv、PCi)和颞顶叶皮层(VPS、h7a)到小脑(小舌),具有共同的神经递质基础和多感觉功能连接。视觉自旋运动知觉似乎特别涉及到扣带沟和小舌之间的皮层小脑轴。CSv和小舌在这个网络中的突出作用被其独特的连接梯度进一步证实。我们的数据支持了区域PcM/pCi作为自我参照空间意识感知所必需的多模态整合功能枢纽的假设。


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









