






摘要:时间的重要性在我们的世界中一直很普遍,正常的光/暗和睡眠/觉醒周期的中断现在已经成为常态。所有情绪障碍,包括季节性情感障碍(SAD)、重度抑郁症(MDD)和双相情感障碍(BD),都与各种生理过程中的异常睡眠和昼夜节律密切相关。光/暗变化和季节变化都有可能对正常睡眠/觉醒模式产生破坏。此外,针对昼夜节律系统的治疗已被证明在某些情况下是有效的。这篇综述将总结这些疾病如何与特定昼夜节律表型相关联,以及将生物钟与情绪调节联系起来的神经元机制。我们还讨论了从昼夜节律的治疗中学到的东西,以及我们如何利用现有知识开发更多个性化设计的治疗方法。
1. 引言
情绪障碍,包括季节性情感障碍(SAD)、重度抑郁症(MDD)和双相情感障碍(BD),都以睡眠/觉醒模式异常为核心症状。此外,睡眠/觉醒周期的急剧变化会导致情绪发作。有数百项研究发现,晚睡型(即喜欢晚上而不是早上的活动)与患情绪障碍之间存在关联。例如,荷兰一项对近2000人的研究发现,晚睡型与抑郁/焦虑之间存在显著关联,即使在调整了社会人口统计学、身体健康和睡眠相关因素后也是如此,这些因素主要是由抑郁症患者造成的。此外,在健康相关问题的广泛评估中,比较了明确的晚睡型和明确的早睡型,发现心理障碍和晚睡型与其他健康问题的关联最强,但有趣的是,总体而言,时间类型较晚的人健康状况较差,甚至所有死因的死亡风险都有所增加。
我们还知道,以特定方式改变昼夜节律为目标的时间生物学治疗对治疗这些疾病非常有益。然而,由于这些疾病的病因复杂,很难说情绪障碍是否直接由昼夜节律中断引起,或者时间型、昼夜节律、睡眠和情绪障碍之间的关联有多少。
昼夜节律钟的进化是为了计算和记录地球每天自转所经过的时间。如果没有这种发展,生物体就无法预测每天的日出,无法形成可靠的作息时间,也无法有效地控制代谢需求。人类共享着这种古老的计时机制的进化保护,几乎身体的每一个细胞、都适应着这种每日循环。这允许身体在活动期间最大限度地产生能量,并在休息期间保存能量,以及恢复和修复细胞功能。生物钟在定时发送免疫信号以保护生物体免受伤害而不产生内部免疫问题方面也起着至关重要的作用因此,当这种内在的计时机制被抛弃时(这在我们的现代世界经常发生),它可以促成甚至导致过多的生理后果,包括睡眠障碍、心脏病、代谢紊乱、癌症、神经退行性疾病和精神疾病等疾病。
昼夜节律在大脑中由位于下丘脑底部的视交叉上核(SCN)集中控制。SCN核心通过含有光感受器黑视素的细胞接收来自视网膜的直接投射,这使得SCN能够直接对昼夜节律的主要载体光做出反应(图1)。昼夜节律的一个重要方面是,即使在没有光线的情况下,它们也会继续以非常接近24小时的周期运行。光和其他因素的夹带对所有生物过程的正常运作至关重要,因为它确保节奏被定时到太阳光/暗周期的适当阶段以及彼此之间。例如,当一个人旅行到一个新的时区时,一个人通常会经历某种形式的“时差”,这是一种情况,SCN的内部起搏器与环境不同步,需要时间进入新的光明/黑暗周期。时差反应的特点是白天疲劳、情绪变化、头痛、注意力不集中和消化问题向东旅行在节奏上产生一个相位提前,而向西旅行则产生一个相位延迟,改变了节奏高峰和低谷的时间通常,每一小时的旅行就需要一天的时间才能进入新的明暗周期,而且与向西旅行的相位延迟相比,向东旅行对相位提前的反应通常更难调整除了带有或不带有的节奏,一个人的时间类型或生活方式选择可能导致高度不规则的睡眠/觉醒模式,或者与正常的带有节奏相比,阶段提前或延迟,这样他们就会比典型的更早或更晚醒来(图2)。
尽管SCN是指挥大脑和身体节奏的主要起搏器,但核心分子钟几乎存在于每个细胞中。哺乳动物的这一核心机制在于昼夜节律基因及其产物之间的转录-翻译反馈回路(图3)。在初级回路中,有一个次级稳定反馈回路,涉及REV-ERBa和视黄酸相关孤儿受体α (RORa)蛋白,它们调节BMAL1的表达,从而影响初级回路的时间。此外,其他几种蛋白质也参与这些分子节律的时间和速度,从而导致数千种其他时钟控制基因的基因表达的昼夜节律,这些基因的身份可能因细胞类型而大不相同。
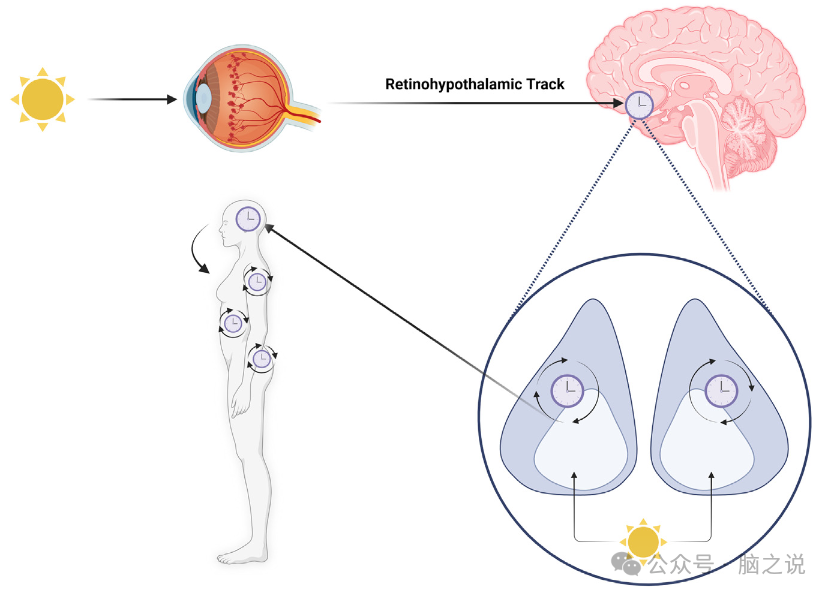
图1 视网膜下丘脑回路对来自环境的光作出反应。
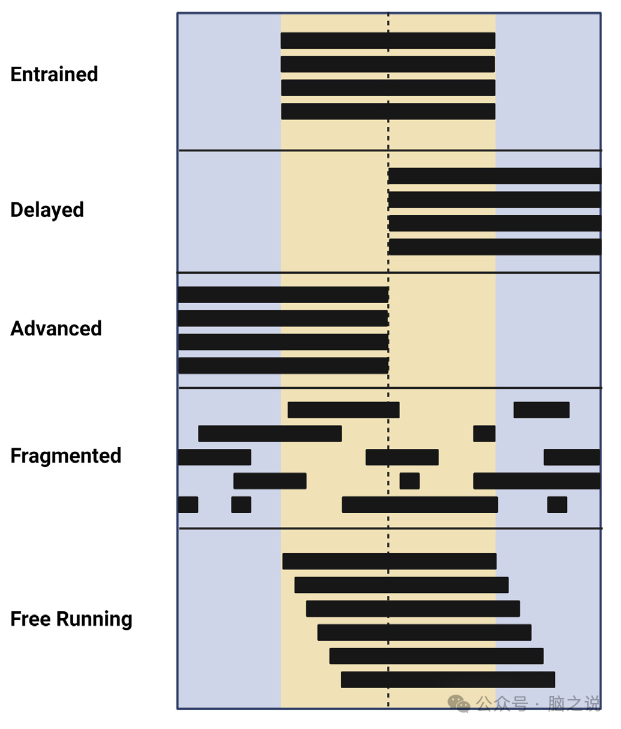
图2 通过活动模式描述的昼夜节律扰动
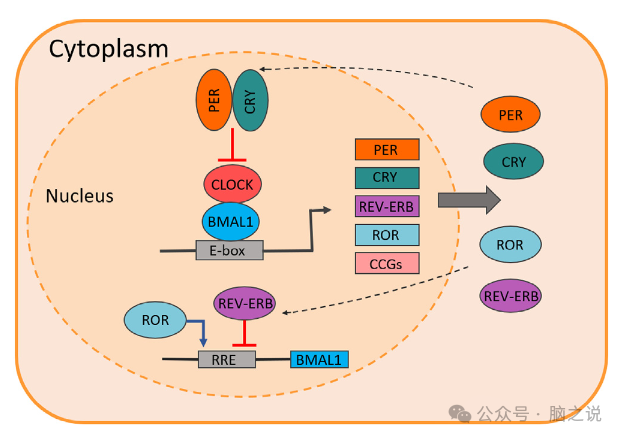
图3 生物钟的分子机制。
2. 季节性情感障碍
根据美国精神病学协会的数据,美国大约有5%的成年人患有SAD。大约25%的双相障碍患者也有季节性的情绪发作模式。SAD的诊断标准可能类似于重度抑郁症,并且与非典型抑郁症的症状更接近重叠,如嗜食,嗜睡和疲劳/嗜睡,而不是忧郁症或典型抑郁症的症状。尽管SAD在很大程度上得到了精神病学社区的认可,但Traffanstedt等人的一项研究未能在美国成年人的横断面调查中发现患者健康问卷抑郁量表的差异,该问卷随纬度或季节而变化,这可能表明季节性抑郁症作为一种疾病可能并不那么明确。抑郁症状持续时间约为一年的40%,而症状的发作和消退时间与白昼时间的长短相对应,表明季节性变化。非典型抑郁症也有昼夜特征,症状有时在晚上加重。然而,那些患有SAD的人通常会在早上看到症状恶化,这表明,尽管这些疾病是相似的,但它们确实是不同的疾病。通常,SAD发生在秋季和冬季之间的过渡月份,此时日光从长光周期到短光周期的变化速率最高。然而,SAD并不局限于冬季,大约10%的SAD患者在夏季也会出现,夏季SAD的症状通常包括失眠和食欲不振症状发作的长度和严重程度是个体的,可以使用季节性模式评估问卷(SPAQ)通过查看全球季节性评分(GSS)来测量。时间类型、性别和年龄也在SAD的发展或表现症状中起作用,对女性有强烈的性别偏见,女性患这种疾病的可能性要高4倍。在Höller等人的一项研究中,发现60岁以下的人和晚上作息类型的人在SAD上的季节性得分更高。有趣的是,纬度(生活在北方)与SAD的关联较弱。然而,离赤道越远,SAD的发病率一般越高。Murray和Hay的一项研究未能发现SAD症状的光周期特异性。然而,这项研究虽然有来自澳大利亚双胞胎登记处的526名女性参与者,但只研究了一个地区(澳大利亚),而不是广泛纬度的地区,这对做出这一评估很重要。
基因也决定了一个人的睡眠类型,这会增加他们患SAD的易感性。一项调查了1539名青少年的研究发现,当用“早睡型”和“晚睡型”问卷进行评估时,“晚睡型”的人在“晚睡型”上的得分明显高于“中睡型”和“早睡型”的人。此外,一项对244人进行的为期3年的研究发现,情绪低落与夜晚状态呈正相关。这可能是由于这样一个事实,即与其他人群相比,晚睡型的人通常在一天中醒来得更晚,这将减少暴露在白天的光线下,特别是在冬季,个人更容易患上SAD。然而,睡眠类型并不能完全预测SAD,而且由于夏季的变化,白天日照减少并不能与所有SAD病例相吻合。另一种可能性是,夜猫子在生理上已经与环境不相适应,他们的节奏是固定的,因此很难适应季节变化。
3. 昼夜节律和悲伤的潜在机制
尽管SAD非常普遍,但其病因尚不完全清楚。动物研究表明,下丘脑和中脑多巴胺(DA)神经元群在SAD中起着重要作用。在一项具有里程碑意义的研究中,暴露于短日光周期的成年大鼠在下丘脑中经历了“神经递质转换”,其中含有生长抑素的gaba能中间神经元变成了DA神经元,阻断或消融这些新的DA神经元会增加焦虑和抑郁样。这些结果很有趣,因为它们表明,大脑中的单胺能系统可以通过改变光周期从根本上改变,这可能导致情绪相关行为的改变。有趣的是,Jameson等人最近的一项研究也发现,光周期长度可以调节伏隔核(NAc)核心的DA信号,这是性别特异性的。与短光周期相比,长光周期培养的雌性小鼠NAc中DA的释放和摄取均增加;然而,在雄性小鼠中没有差异这是特别有趣的,因为与男性相比,女性的SAD患病率更高。
由于老鼠是夜行动物,所以它们通常厌恶光线,这使得一些结果,比如上面提到的,很难与人类进行比较;因此,研究者转而使用昼行草鼠作为具有更高面孔效度的模型。草鼠被安置在8:16的模拟冬季的光周期会增加类似抑郁的行为。这些研究还发现,神经肽食欲素有助于调节这些动物的情绪和焦虑的光诱导变化。与小鼠相反,草大鼠下丘脑DA神经元的数量在短光周期内减少,并且DA神经元的数量可以通过食欲素受体拮抗剂处理而改变,这表明食欲素参与了DA神经元的这些变化。Costello等人的另一项研究发现,在与强光疗法(BLT)类似的方案下,对白天活动的草地大鼠进行光照,会导致更高的觉醒程度,提高夜间睡眠质量,改善节律,这与在人类身上观察到的情况相似。综上所述,有强有力的证据表明,光周期的变化直接影响DA信号,可能通过食欲素,这有助于季节性情绪变化。然而,值得注意的是,光周期诱导的DA信号变化在白天和夜间的物种中是不同的,并且是大脑区域和性别特异性的。
众所周知,皮质醇节律与外界的光/暗周期同步,在傍晚达到最低点,然后逐渐增加,直到在醒来前达到峰值,同时血清素也会出现峰值,然后再次逐渐减少SAD患者在夏季有皮质醇觉醒反应(CAR),与健康人群没有区别,但在冬季,当SAD患者报告压力、抑郁和焦虑的感觉增加时,与健康人群相比,CAR减弱。然而,这一发现并不一定表明SAD是一种低皮质醇状况。在对13篇论文的系统回顾中,那些使用异质方法和样本的论文无法一致地复制SAD患者的CAR反应。因此,需要进一步的研究来了解是否可能存在一个亚组的受试者,其中冬季CAR反应减弱。
季节性神经活动和行为的控制似乎也在一定程度上局限于结节部(PT),这是一种包裹在脑垂体周围的管状鞘,可能与SCN协同作用,产生季节性的计时和夹带信号。SCN的可塑性是随着白昼长度的变化而发生的。在仓鼠SCN切片中,与短日相比,夏季的长日光周期导致种群水平上神经元活动周期的持续时间增加除了种群活动外,特定类型的SCN神经元之间的相位分布对季节性干扰也很重要较短的白天导致这些SCN亚群之间的放电模式相对同步,而较长的白天导致这些亚群在昼夜循环的不同阶段被激活,有可能SAD患者的SCN不能正确适应这些不同的季节性光线。也有可能SAD患者对光的敏感度不同。光和光周期夹带可以使用照明后瞳孔响应(PIPR)来测量。在Roecklein等人的一项研究中,SAD患者或对照组在秋季或冬季暴露在红光或蓝光下他们发现,与对照组相比,SAD组的PIPR降低,总体变化也更低,尤其是在蓝光下。他们提出,PIPR的减少可能与一种特定的黑视素基因OPN4变异有关,这表明这种昼夜节律光感受器对光输入不太敏感。
重要的是,尽管如上所述有证据表明,情绪对光线的相关反应是通过从眼睛到SCN,然后是其他大脑网络的途径来调节的,但Samar Hattar的研究小组发现,眼睛对habenular周围区域有直接的投射,老鼠对光/暗周期变化的抑郁相关反应是通过这条途径发生的,这条途径完全独立于SCN。缰环区和外侧缰环之间的通信可能调节多巴胺能活性,以响应上述光周期变化,导致多巴胺能传递减少。因此,目前尚不清楚在夜间或较短的光周期中,有多少与情绪相关的反应是由SCN调节的,而不是由眼睛投射到habenular周围区域。人体内的MEL在晚上由松果体释放,以促进睡眠的开始。然而,在许多哺乳动物物种中,MEL在季节性过程中也发挥着更大的作用,包括繁殖模式、攻击性和冬眠SAD的症状包括疲劳加剧、睡眠过多和对碳水化合物的渴望,而白天长度减少导致的MEL的季节性变化可能是这些干扰的促成、原因或结果(图4)。事实上,MEL在冬季过量生产在一些SAD患者中是表型的。SAD患者的MEL分泌时间也有明显的延迟,这种情况在冬季加剧,导致白天更疲劳。因此,MEL可能在SAD的许多症状中发挥关键作用。然而,越来越多的证据表明SAD是一种涉及多种神经递质和调节剂的全系统失调问题。
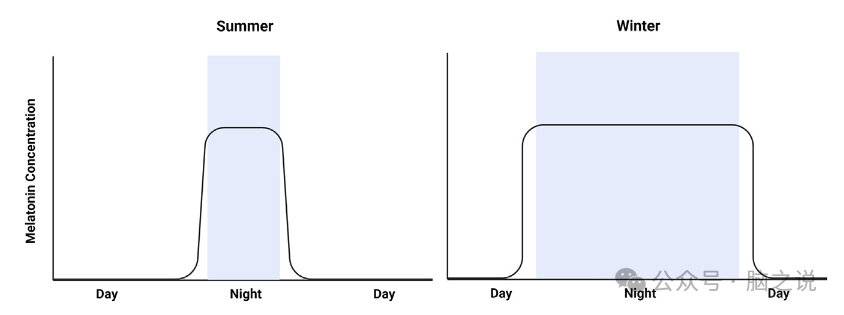
图4 褪黑素分泌持续时间随季节夜间长度的变化而变化
4. SAD的治疗方法
BLT是SAD最广泛使用的治疗方法。当暴露在明亮的广谱光下30分钟时,大多数SAD患者报告在专门使用和参与治疗后症状改善或停止。通常,BLT是在清晨开的,因为研究表明,这是最有效的时间窗口,因为它可以将昼夜节律提前,使其更好地与环境保持一致,而且大多数SAD患者往往是相位延迟的。BLT时间和时间型的差异尚未被观察到。然而,个体的BLT时间存在差异,优化规定的时间和光的“剂量”是重要的。Lewy等人的一项重要研究通过在早上或下午/晚上给药低剂量MEL来诱导期提前或延迟,验证了SAD治疗的相移假说。在这项研究进行的4年期间,他们的给药方案有所不同,每天的MEL总量为0.225毫克(第2年)或0.3毫克(第1、3和4年)。他们发现,根据轻度MEL发作和睡眠中点之间的时间,对于阶段调整患者存在最佳的“治疗窗口”。适当时间的MEL治疗(通常在夜间)与症状改善显着相关,而不适当时间的MEL治疗与症状改善无关。
BLT可能对SAD有益的另一种方式是它减少了MEL的产生量。当暴露在光线下,特别是蓝光下,MEL的产生停止,并迅速从体内排出对于那些白天MEL产量增加或MEL释放相移的个体,在白天它仍然存在,BLT将降低MEL水平。选择性5 -羟色胺再摄取抑制剂(SSRIs)也被用于治疗SAD,并取得了令人鼓舞的结果;然而,应该指出的是,仍然需要对副作用“成本效益”和结果的复制进行进一步的研究,并且在许多研究中都指出了这一点。
5. 重度抑郁症
重度抑郁症是一种常见的使人衰弱的疾病,影响着全世界3亿多人在过去的几十年里,重度抑郁症的患病率一直在稳步上升重度抑郁症的特征是情绪低落,对愉快的活动失去兴趣,以及生理和认知症状。失眠或睡过头、疲劳和精力下降以及食欲变化在重度抑郁症患者中很常见。失眠或睡过头是重度抑郁症的主要诊断标准之一,超过80%的重度抑郁症患者报告有睡眠障碍健康人的情绪在24小时内波动;然而,重度抑郁症患者的情绪昼夜变化是改变的。与健康对照者相比,抑郁症患者在积极情绪和消极情绪方面表现出明显的昼夜干扰患者通常在清晨表现出最严重的症状,一小部分患者在下午也会出现萎靡不振睡眠类型也可能影响情绪的昼夜变化,晚上睡眠类型的重度抑郁症患者更有可能在早上经历症状恶化,整体上增加反刍。此外,夜间睡眠类型是重度抑郁症的一个重要危险因素,与更频繁、更严重的抑郁发作、更高的自杀风险和SSRI治疗效果降低有关世界范围内抑郁症发病率的上升可能与社会的现代化有关,人们越来越多地接触人造光、轮班工作和跨越多个时区的航空旅行。生活在夜间室外光照水平最高的地区的人出现抑郁症状和自杀行为的几率更高。
倒班工作会扰乱昼夜节律,导致许多负面健康后果,包括睡眠障碍、疲劳、认知障碍,以及心血管和代谢紊乱以及某些类型癌症的风险增加。李等人的荟萃分析表明,夜班工作也与患抑郁症的风险增加有关此外,与一般人群相比,空乘人员有更高的睡眠障碍、疲劳、抑郁和焦虑的患病率在啮齿类动物中,暴露于时差模式会使抑郁样行为恶化,这在蔗糖摄入量减少、强迫游泳测试中不活动时间增加和开阔场地测试中进入中心区域的次数减少中得到了证明也有证据表明,长途飞行可以引发有精神病史的人的抑郁发作。
一项对人类死后大脑中昼夜节律基因表达的研究发现,在观察一群在一天中不同时间死亡的受试者时,重度抑郁症患者的基因表达节律模式要弱得多(即振幅较低),在个体基因之间的同步时间和中断阶段关系中尤其如此。动物研究也表明,昼夜节律在重度抑郁症中起着重要作用。在慢性社会失败压力下易患抑郁样综合征的小鼠也会在核心体温中丧失昼夜节律性,而在压力后能适应抑郁样表型的小鼠则不会最近,利用CRISPRCas9编辑猴胚胎培育了一只Bmal1基因敲除的食蟹猴,这些猴子有更多的夜间运动和减少睡眠,当它们被关在恒定的光线下时,这种情况会加剧。此外,这些猴子表现出与焦虑和抑郁一致的表型,包括血液皮质醇升高。血液转录组分析显示,与炎症和应激反应相关的基因发生了多种变化,该基因也存在于重度抑郁症患者和睡眠剥夺患者的血液转录组中。
6. 昼夜节律和重度抑郁症的潜在机制
下丘脑-垂体-肾上腺(HPA)轴的失调是重度抑郁症患者的常见发现。HPA控制糖皮质激素的产生和分泌,糖皮质激素是所有主要生理系统的有效调节剂。糖皮质激素的表达具有强烈的昼夜节律。在健康受试者中,皮质醇分泌最大值发生在早晨醒来前的几个小时,然后在白天逐渐减少然而,在抑郁的个体中,研究报告显示,皮质醇在早上水平较低,而在晚上水平较高此外,与对照组相比,重度抑郁症患者早晨皮质醇的阶段提前也有报道此外,皮质醇水平失调可能使个体易患随后的抑郁发作许多研究表明,HPA和免疫系统之间的相互作用介导了对压力的行为反应,这是抑郁症发展的常见诱因。然而,失调的免疫系统会导致慢性炎症,并有助于精神疾病的病理生理,包括抑郁症。昼夜节律紊乱导致促炎细胞因子水平升高,包括肿瘤坏死因子- α (TNF-a)反过来,TNF-a已被证明在调节昼夜节律中发挥重要作用,主要是通过干扰e -box介导的转录,从而抑制时钟基因Per1和Per2的表达,在体外和体内都是如此。由于昼夜节律紊乱而导致的慢性睡眠剥夺,可导致小鼠炎症特征增加,小胶质细胞激活和星形细胞吞噬增加另一方面,小胶质细胞功能障碍也可能是睡眠障碍病理生理学的一个促成因素,可能导致睡眠和情绪紊乱的恶性循环。
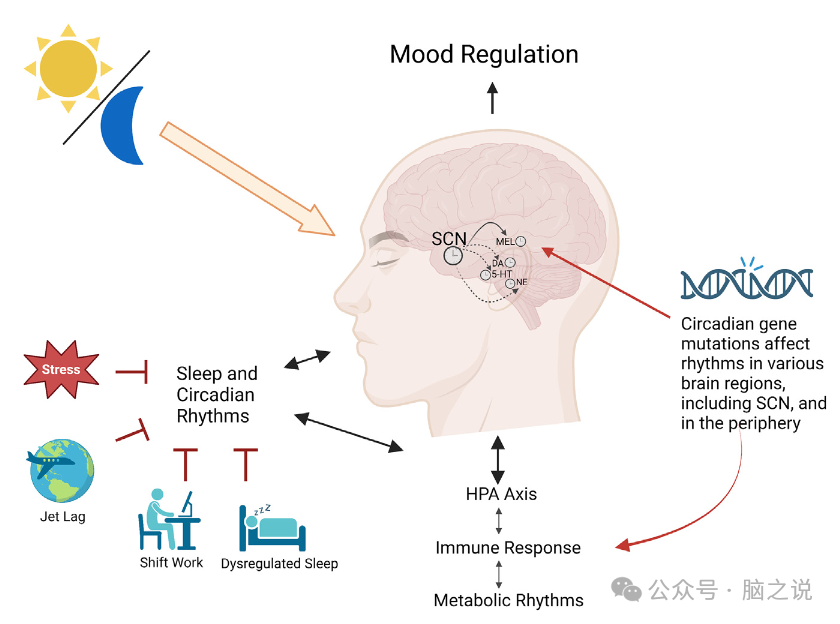
图5 昼夜节律和环境对情绪的调节
与季节性抑郁症类似,重度抑郁症也可能涉及单胺信号的中断。大鼠在持续的黑暗中保持6周,这样它们就不能进入光中,发展出抑郁样表型,在强迫游泳测试中,静止时间增加,这与去肾上腺素能蓝斑系统的损伤有关。此外,这些自由运行条件导致去甲肾上腺素能、多巴胺能和血清素能神经元的凋亡增加,而去西帕明治疗可部分挽救这些神经元。有趣的是,这些对单胺能细胞的昼夜节律影响与压力的增加无关,这表明这种影响与一般的压力反应无关。大脑中富含单胺的区域内的昼夜节律基因也在情绪调节中发挥作用,无论是否有慢性压力。慢性社会失败压力显著降低了NAc中Per1和Per2的表达,与此一致的是,在NAc中特异性地降低Per1和Per2会导致焦虑样行为的增加。氟西汀治疗使压力后的Per基因表达正常化,并逆转社会互动缺陷,表明Per1和Per2基因参与抑郁样和焦虑样行为。有趣的是,在习得性无助范式中,敲除NAc中的Bmal1导致对无助行为的易感性降低。此外,Bmal1的敲低,特别是NAc的星形胶质细胞中的Bmal1的敲低,会导致探索性驱动的增加。因此,需要做更多的工作来了解个体昼夜节律基因在这些大脑区域和特定细胞类型中的复杂作用,以及所有这些不同因素如何促进MDD的发展(图5)。
7. 重度抑郁症的治疗
昼夜节律和抑郁症之间联系的进一步证据来自这样一个事实,即直接针对昼夜节律系统的治疗(如急性睡眠剥夺、BLT和睡眠阶段提前)可以成功治疗抑郁症。抑郁症患者通常表现为睡眠阶段延迟,睡眠开始时间在凌晨2点到6点之间。这种明显的阶段延迟可能是由于生物或自我施加的内部生物钟与睡眠/觉醒周期之间的失调,针对昼夜节律系统和/或认知行为疗法(CBT)的治疗可能有助于阶段提前和调整昼夜节律。用于快速治疗抑郁症的最有效的时间疗法之一是急性睡眠剥夺,这已被证明可改善约40%至60%患者的抑郁症状,而且反复进行急性睡眠剥夺治疗的效果越来越好。然而,急性治疗的效果通常不会持续很长时间,而且通常在一夜睡眠后症状又会复发。睡眠剥夺可能通过重置昼夜节律基因表达异常来发挥其抗抑郁作用。有趣的是,睡眠剥夺和氯胺酮(另一种速效治疗)都会导致前扣带皮层的转录变化,其中许多基因下调,包括Per2和Npas4.125。有趣的是,重度抑郁症患者对氯胺酮的急性和持久反应都可以根据他们的日常活动谱来预测。在氯胺酮治疗之前,较早阶段的活动模式和较低的中神经(节奏调整的平均值)区分了随后的应答者和无应答者。此外,在第1天,氯胺酮无反应者相对于基线有钝化的24小时振幅,而氯胺酮治疗后的第3天更高的振幅与持续的临床反应相关。这些数据表明,氯胺酮对那些在基线时具有特定昼夜节律特征的人最有效,对那些服用氯胺酮的人有持续的抗抑郁反应。
如前所述,与SAD类似,许多MDD患者是期延迟的,一些MDD患者的MEL水平较低,或者时间不合适,这一点在保持适当的睡眠/觉醒节奏中非常重要。在临床环境中,8周的晨间BLT单独或联合氟西汀对减轻重度抑郁症患者的抑郁症状具有良好的耐受性和有效性,蒙哥马利-阿斯伯格抑郁评定量表的评分变化证明了这一点。最近Huang及其同事提出了BLT治疗重度抑郁症的抗抑郁作用的潜在机制。他们发现一个专门的视网膜-腹侧外侧膝状核和膝间小叶(vLGN/IGL)外侧缰状通路调节抑郁样行为。具体来说,通过蔗糖偏好、强迫游泳和穿梭箱试验,激活投射到vLGN/IGL的内在光敏视网膜神经节细胞(ipRGCs),激活投射到LHb的vLGN/IGL神经元,或抑制突触后LHb神经元,足以减少啮齿动物的抑郁样行为。
需要强调的是,虽然早晨的光照有助于缓解抑郁症状,但夜间的光照会对健康产生负面影响,并增加患抑郁症的几率。An等人最近的一项研究有助于解释为什么光照在白天和晚上对情绪的影响不同。研究人员发现了一条昼夜节律门控的神经通路,从固有的光敏视网膜神经节细胞ipRGCs到背侧habenular周围核(dpHb)再到NAc,后者在夜间优先传递光信号。特别是,他们发现投射到NAc的dpHb神经元亚群在夜间更容易兴奋,并且优先对夜间的光线而不是日光做出反应。在夜间给药时,这些dpHb神经元的光遗传学或化学遗传学兴奋导致蔗糖偏好降低,而在白天则没有。
在人类和啮齿动物身上进行的研究表明,抗抑郁药物治疗也会导致昼夜节律的阶段性提前。氟西汀,一种SSRI,可以在SCN中提前神经元放电。用西酞普兰(另一种SSRI类药物)治疗增加了重度抑郁症患者昼夜节律系统对光线的敏感性,这可能解释了为什么BLT与SSRI类药物联合使用时更有效。阿戈美拉汀是MEL受体MT1和MT2的高亲和力激动剂和5-HT2C(5-羟色胺受体)拮抗剂,已被证明可以减轻MDD患者的抑郁症状,并已被证明可以改变体温和激素释放的昼夜节律相移,纠正成年大鼠产前约束应激引起的昼夜节律中断,这表明其作为抗抑郁药的作用机制涉及调节昼夜节律系统。
失眠是重度抑郁症的常见症状之一,其中一种可以改善重度抑郁症和失眠合并症的非药物治疗失眠的方法是CBT- I。有趣的是,CBT-I已被证明在降低抑郁发作的风险和治疗抑郁症方面是有效的。有趣的是,一项评估CBT-I对合并抑郁和失眠的个体有效性的随机对照试验发现,CBT-I对偏爱晚上的个体尤其有效。研究人员建议,如果考虑到患者的昼夜节律偏好,临床护理应予以考虑。
重度抑郁症是一种异质性很强的疾病,未来临床研究的关键将是测量每个人的昼夜节律和睡眠模式以及生理节律,以便选择合适的时间治疗方法,产生最佳反应。对慢性压力和其他因素如何与昼夜节律系统相互作用的更好的机制理解也将导致更有针对性的治疗的发展。
8. 双相情感障碍
双相障碍是一种复杂的精神障碍,以躁狂和抑郁状态以及两种状态的混合状态为特征。在两种状态之间,个体可能会经历一段相对稳定的时期,称为心境愉悦。据估计,美国有2.8%的人被诊断为双相障碍,男性和女性患双相障碍的比例相当,只是在症状表现上存在一些临床差异。多年来,人们已经很清楚,除了与这些状态相关的情绪和情绪变化外,躁狂和抑郁发作的主要特征还包括能量和活动的变化在躁狂期间,个体通常精力充沛,特点是承担多项任务,表现出语速快,思维敏捷,尽管睡眠很少。在抑郁症发作期间,人们通常会描述自己缺乏精力,日常工作使他们筋疲力尽,而行为、动作、语言和思想都可能变慢,他们通常会睡过头。多项研究发现双相障碍患者的睡眠和昼夜节律紊乱,表明双相障碍患者的睡眠/觉醒模式高度紊乱,在躁狂或抑郁发作期间更加不同步。多达三分之一的双相障碍患者有可诊断的昼夜节律障碍,最常见的是睡眠阶段延迟障碍。事实上,与对照人群相比,大多数双相障碍患者的夜间睡眠类型与重度抑郁症和重度抑郁症患者相似。此外,在5年的随访中,较晚的时间型预示着双相障碍中更多的抑郁症状和更少的躁狂发作。许多研究已经确定了调节昼夜节律的基因中的SNP,并发现了核心昼夜节律钟的各种元件与BD之间的表面上显著的关联。最常见的是clock、PER3和ARNTL中的SNP。ARNTL之所以特别有趣,是因为它在BD的大型GWASs中缺乏全基因组意义。此外,当昼夜节律系统被视为一个完整的网络而不是单个基因时,与BD有显著的联系。
15%-25%的双相障碍患者有季节性发作模式。在这些患者中,躁狂发作倾向于在春季发生,而抑郁发作倾向于在秋季/冬季发生。有趣的是,PER3基因的一种变异与抑郁症的季节模式有关。随着季节变化,正常睡眠模式的改变是躁狂或抑郁发作的最常见诱因之一。不规律和紊乱的睡眠/觉醒节律与双相障碍的首次发病有关,并可能预示着这一点,尤其是在高危人群中。旅行引起的时差反应可能是一个重要的诱因,有趣的是,一些研究表明旅行方向很重要,向东旅行会增加躁狂发作次数,向西旅行会增加抑郁发作次数。这与一些研究一致,这些研究发现,控制昼夜节律的基因在经历躁狂发作的外围样本中往往运行得更快(即少于24小时的节奏),而这些基因在经历抑郁发作的样本中运行得更慢(大于24小时),在心境平静期间恢复到更常规的24小时周期。有趣的是,研究还发现,患有双相障碍的人,平均而言,对夜间光线过敏,因为夜间的光线会导致MEL的更大抑制和睡眠延迟,一些人认为这可能是双相障碍的特征标志。过量的咖啡因、酒精和滥用药物也会引发发作,暴露在压力生活事件中也是如此。由于这些环境因素会导致睡眠时间不恰当、效率低下和睡眠时间缩短,因此睡眠/觉醒周期的中断可能是导致发作的共同线索。这些观察结果形成了双相障碍的社会授时因子理论,该理论最初由Ehlers、Frank和Kupfer于1988年提出,该理论假设“生活事件干扰社会授时因子,从而扰乱生物节律,导致易感个体出现情感症状。
9. 昼夜节律与双相障碍的潜在联系机制
分子钟严密地控制着细胞的能量状态、新陈代谢、蛋白质翻译过程、神经传递、这些节律是必不可少的,因为当个体清醒时,大脑中的细胞需要高度代谢活跃,然后有一段时间的恢复,在这段时间内,它们可以清除细胞中错误折叠的蛋白质、自由基和活性氧。核心分子钟蛋白直接与线粒体呼吸产生的代谢物结合,充当“氧化还原传感器”,通知细胞核中的分子钟细胞的代谢状态反过来,时钟蛋白直接调节线粒体功能和抗氧化功能所必需的基因。线粒体的融合和裂变以及新线粒体的产生具有昼夜规律,并依赖于具有功能的分子钟此外,当分子钟在动物模型中被破坏时,这将导致大脑中活性氧的大量增加,广泛的胶质细胞增生,最终导致神经退行性变。许多关于双相障碍的研究表明,线粒体功能的破坏和氧化应激是该疾病的核心。因此,昼夜节律钟对这些过程的直接调节可能在疾病发展中至关重要。
多项研究表明,双相障碍与多巴胺能传递的改变有关。躁狂通常与较高水平的双相情感障碍有关,而双相情感障碍的再摄取可能更多,尽管这些研究需要进一步的复制。特别是,狂躁症包括过度的目标导向活动和由高度的奖励寻求驱动的冲动决策。这些过程是由皮质-边缘多巴胺能系统调节的。ClockΔ19小鼠中脑多巴胺能活性增加,酪氨酸羟化酶(TH)水平增加,DA合成增加,这在它们的非活性期更为明显。ClockΔ19小鼠的躁狂样行为可以通过野生型小鼠腹侧被盖区DA神经元的慢性光遗传刺激重现,但仅在小鼠的非活动期,这表明是不适当的DA增加时间导致躁狂,而不一定是在一天中的任何时间过量的DA。在光期而非暗期阻断TH可逆转躁狂样行为。这与因光照/黑暗周期改变而引起的躁狂症的沉淀是一致的,因为这可能导致一天中预料不到的时间DA激增。因此,昼夜节律系统直接调节关键的细胞过程和多巴胺能传递,这可能有助于双相障碍。
另一个有趣的可能性是“代谢时差”的概念。双相障碍通常与代谢问题有关,包括肥胖。代谢时差指的是“一种转变的状态”。能量平衡的昼夜节律模式,影响神经内分泌、免疫和脂肪组织功能。可能是双相障碍患者不稳定的睡眠和饮食模式导致微生物组功能、胰岛素-d信号和代谢肽(如胃饥饿素和瘦素)表达的节律高度紊乱,所有这些都已知会影响情绪。最近对使用生酮饮食或限时喂养来控制双相障碍发作的兴趣可能至少通过稳定这些系统而获得部分效果,尽管需要做更多的工作来充分了解BD的机制和有效性。
10. BD的治疗
人际和社会节律疗法(IPSRT)是一种基于社会时代体理论的治疗方法。IPSRT是一种手动的、基于证据的疗法,有助于调整和稳定睡眠/觉醒和社会节律,以帮助预防发作和减轻症状。在临床研究中,IPSRT产生的治疗效果比强化的临床管理更快速、更持久。有趣的是,最近一项针对SRT治疗本身的研究,对青少年和年轻双相障碍患者的远程医疗服务进行了改进,支持其在减少双相障碍症状和自杀倾向方面的作用。
双相障碍通常用心境稳定药物治疗,包括锂盐、抗惊厥药物(如VPA)和/或第二代抗精神病药物。早上的BLT,通常用于治疗SAD和某些形式的重度抑郁症,已经被用于治疗双相障碍,然而,它可以触发躁狂症的转变。研究发现,在下午给予BLT,不会导致昼夜节律周期的大阶段提前,可能对双相抑郁症患者有帮助,引发躁狂发作的风险更小。重要的是,所有这些治疗都可能对昼夜节律机制产生强烈影响。例如,锂增加分子节律的振幅,可能是通过抑制糖原合成酶激酶,这是已知的磷酸化分子钟的几个成员,改变他们的稳定性和核进入这可以像IPSRT一样保持睡眠/觉醒模式稳定,防止发作。在386名双相障碍患者的锂离子多中心前瞻性试验中,接受稳定锂离子治疗的个体的昼夜节律干扰最小,而没有锂离子暴露史的个体的昼夜节律干扰最明显。锂的另一个众所周知的作用是延长昼夜节律周期,使其增加到24小时以上除了Gsk3β外,锂对肌醇三磷酸(IP3)信号的影响可能是锂的一些延长周期作用的基础。早期的研究发现,锂反应者是那些内源性周期少于24小时的人,这也通常在躁狂状态下发现。从双相障碍患者身上分离出来的成纤维细胞,如果长时间服用,对锂的昼夜节律反应就会减弱。有趣的是,这些患者在收集组织时,医生没有给他们开锂的处方,这表明他们对锂没有反应。此外,锂反应者诱导多能干细胞(iPSCs)分化为神经元前体细胞和谷氨酸能神经元的昼夜节律周期更短,锂延长了这些细胞的昼夜节律周期。IP3信号的遗传变异可能是对锂有反应和无反应的一些差异的基础有趣的是,VPA对细胞培养中分子节律的节律幅度有类似的影响,但它对周期的影响可能与锂相反,产生更短的昼夜节律周期,并且很容易推测测量BD的昼夜节律可能有助于预测锂或VPA的治疗反应。已经进行了多项临床试验,测试MEL或MEL受体激动剂对BD的疗效,结果好坏参半。这些试验的两项荟萃分析得出结论,受体激动剂拉美替恩可能预防双相障碍患者抑郁复发,并且检测到的最大疗效信号是用于躁狂症状。事实上,在两项评估急性躁狂期间躁狂症状的试验中,辅助MEL的治疗效果优于安慰剂,但研究和患者特征之间存在很大的异质性,一些研究发现患者结局没有显著差异。
11. 结论
总之,很明显,昼夜节律和适应光周期变化的能力在情绪调节中起着关键作用。有趣的是,在这些疾病的多项研究中,一个一致发现的风险因素是晚时型。然而,尚不清楚这种与精神疾病的关联是由于导致“夜猫子”的遗传因素,还是由于学校或工作导致的内部节律与早醒时间不一致所致。最近的COVID-19大流行以及随后的学校和办公室关闭提供了一个难得的机会,可以在没有早起环境压力的情况下自行检查睡眠类型。有趣的是,一项研究来自12个不同国家的19000多名成年人发现,夜猫子的睡眠延迟时间大大增加,尤其是在工作日,睡眠时间也增加了。然而,与其他昼夜节律类型的人相比,他们的心理健康、幸福感和生活质量自我报告也更差。在对年龄、性别、社会经济地位或大流行期间禁闭时间进行校正后,这些结果站住脚。这些结果表明,即使没有早起的压力,夜猫子也可能更容易患抑郁症或其他情绪发作,尤其是在面临慢性压力时。然而,其他一些关于推迟上学时间和灵活的工作时间的影响的研究表明,环境的变化对晚睡的人的睡眠和情绪有有益的影响,所以,很可能存在生物脆弱性,而且早起引起的社交时差加剧了这种脆弱性的影响。
各种疾病的另一个共同特征是,不稳定的睡眠/觉醒或不断变化的光/暗周期会加速发作或使发作恶化。这表明,许多患有情绪障碍的人可能有一个不灵活的生物钟,无法适当地适应变化。这可能是由于夹带机制的缺陷,可能对晨光不太敏感,或者SCN不能正确地响应提示。事实上,动物模型的研究表明,与大脑其他部分不同步的SCN会导致比整个SCN损伤更糟糕的行为结果,后者实际上可以产生对抗压力的保护作用。也有可能,特别是在BD的情况下,SCN太容易陷入相位提前或相位延迟的状态,而日常的不稳定性是造成发作脆弱性的原因。在面对睡眠模式或环境的改变时,需要做更多的工作来确定哪些类型的机制驱动这些发作。
也有越来越多的证据表明,稳定和协调昼夜节律对情绪障碍有治疗作用。在SAD的情况下,节奏需要更好地与环境保持一致,这通常可以通过早上的BLT来实现,将节奏提前,但有些人可能会从MEL或其他授时基因在一天中的不同时间的干预中受益。对于重度抑郁症,节律的阶段推进可能对某些人也有治疗作用,但对另一些人来说,通过氯胺酮等药物增加节律振幅也可能有治疗作用。对于双相障碍,日常节律稳定是关键,但也有证据表明,在躁狂期间,节律可能需要延迟,在抑郁期间,节律可能需要提前。此外,还应评估每个人在健康状态下的内部相,以确定适当的时间治疗。更好的个人昼夜节律分析和更彻底的时间治疗方法测试将最终指导适当的个性化治疗方案,这可以增强或取代现有的情绪调节疗法。
参考文献:Circadian rhythms and mood disorders:Time to see the light.


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









