






摘要:数十年的神经影像学研究显示,抑郁症患者的大脑结构和连接方面存在轻微差异,这阻碍了机制洞察或识别疾病发病风险因素。此外,抑郁症是周期性的,很少有纵向神经影像学研究,限制了对推动情绪状态转变的机制的理解。新兴领域的精密功能映射已经利用密集取样的纵向神经影像数据显示健康个体之间和内部脑网络拓扑和连接的行为上有意义的差异,但这种方法尚未应用于抑郁症。在这里,使用精密功能映射和几个深度取样个体的样本,我们发现大多数抑郁症患者的额-纹状体显著网络在皮质中扩张了近两倍。这种效应在几个样本中均可复制,并主要由网络边界转移引起,不同个体中出现三种不同的侵入模式。显著网络的扩张在时间上是稳定的,不受情绪状态的影响,在青少年抑郁症发生前的儿童中可以检测到。在1.5年内对扫描了多达62次的个体进行纵向分析,发现额-纹状体回路中的连接变化与特定症状波动相关,并预测未来的无欲症状。综上所述,这些发现确定了一种类似特质的脑网络拓扑结构,可能增加了抑郁症的风险,并且额-纹状体回路中的情绪状态相关连接变化可预测随时间推移抑郁症症状的出现和减轻。
1. 引言
抑郁症是一种异质性和周期性的神经精神综合征,与突触丢失和前纹状体网络的连接改变有关,并且是全球残疾的主要原因。导致特定抑郁症状领域或随时间变化情绪改变的神经生物学机制尚不完全了解,尤其是在神经系统水平。到目前为止,大多数功能性磁共振成像(fMRI)研究在交叉组间比较中对抑郁症患者和健康、从未患抑郁症的对照组之间的功能连接差异进行测试,使用群体平均(大小合适的)分割来定义功能性脑区和网络。最近,系统神经科学的开创性工作催生了精密功能映射领域,指的是一套新方法来完全描绘个体的功能网络。精确映射研究表明,个体的功能区域和网络的拓扑结构(大小、形状和空间位置)与群体平均描述明显不同,并且个体之间网络拓扑的差异是稳定的,可遗传的,并且与认知能力和行为相关。除了最近有一个涉及到双侧围产期中风受害个体的病例研究外,这些工具尚未被广泛应用于包括抑郁症在内的临床人群。
抑郁症从根本上来说是一种偶发性神经精神疾病,其定义是在情绪正常时期之间插入不连续的情绪低落时期,但我们对随着时间的推移调节情绪转变的机制的理解是有限的。部分原因在于,到目前为止,大多数研究都是横断面的,涉及在单个时间点获取的数据,或者在某些情况下,在干预前后获取两次或三次扫描——这种方法 不是为个人层面的有意义的统计推断而设计的。了解介导抑郁情绪状态转变的神经生物学机制可能需要对个别患者进行数月的研究。事实上,涉及颅内脑电图记录和其他评估的密集采样 n-of-1 研究已经开始显示调节接受深部脑刺激治疗抑郁症的个体患者情绪状态转变的机制,但这些方法尚未在 fMRI 研究中的量表。如果没有这样的数据集,我们尚不清楚大脑网络连接的变化是否可以预测其他抑郁症状领域中快感缺乏、焦虑和功能障碍的出现,或者从发作中恢复后这些症状随后的缓解。同样,目前还不清楚非典型网络拓扑测量是否会随着抑郁症患者的情绪状态而波动,还是随着时间的推移保持稳定——这是理解临床神经影像学中因果关系、定义神经调节干预或治疗中潜在治疗目标的关键问题。识别高危人群。直到最近,技术限制对在临床样本(包括抑郁症)中进行精确的功能映射和纵向神经成像构成了重大障碍。单个受试者水平的传统 fMRI 测量通常存在噪声且可靠性有限,部分原因是它们对各种成像伪影敏感30。然而,最近的研究已经采取了重大步骤,通过获取每个受试者的大量数据或使用多回波功能磁共振成像来开发这些问题的解决方案。这些方法共同可以在个体受试者水平上生成高度可靠的功能连接测量和网络图,这是开发和部署用于临床转化目的的功能磁共振成像的重要一步。
在这里,我们使用最先进的精确功能映射工具来描绘抑郁症患者的功能性大脑网络的拓扑结构,并利用深度采样个体的多个静息态功能磁共振成像数据集。我们发现,在大多数抑郁症患者中,额纹状体显着性网络扩大了近两倍——我们使用重复抽样的抑郁症患者的独立样本和大规模组平均数据,三种不同类型的侵犯在个体之间发生,取代了邻近的功能系统。随着时间的推移,显着性网络的扩张是稳定的,并且不受情绪状态变化的影响。在青春期后期出现抑郁症状之前接受扫描的儿童中也存在这种情况。对密集采样个体的纵向分析显示,纹状体与显着网络的前扣带回和前岛叶节点的连接发生了情绪状态依赖性变化,分别跟踪快感缺乏和焦虑的波动,并预测在未来的研究访问中随后出现的快感缺乏症状。
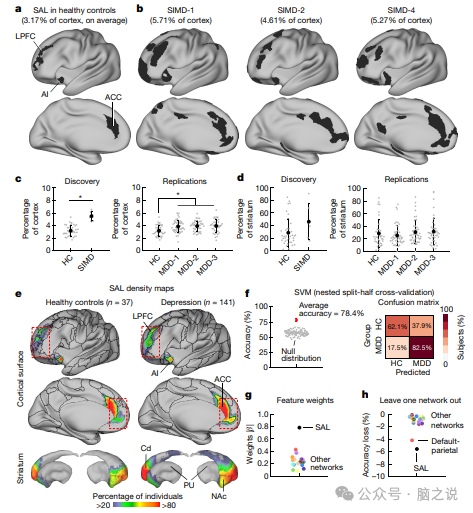
图1 在高度采样的抑郁症患者的皮层中,额纹状体显着网络扩大了近两倍
2. 抑郁症中的显着网络扩展
涉及大量抑郁症患者的大量神经影像学研究已经确定了功能连接和大脑结构的差异,通常涉及前扣带皮层、眶额皮层、岛叶皮层和膝下扣带皮层。大规模功能性大脑网络的拓扑特征(形状、空间位置和大小)是否在抑郁症中发生改变尚不清楚。
我们使用精确的功能映射来描绘 6 名高度采样的单相重度抑郁症个体的功能性大脑网络拓扑,这些个体在 22 个疗程中平均接受了 621.5 分钟的多回波 fMRI 扫描。我们将该数据集称为重度抑郁症串行成像 (SIMD) 数据集。为了了解这些个体抑郁症状的严重程度,17-item39 汉密尔顿抑郁评定量表 (HDRS17) 平均得分为15.7 ± 3.7,表示从轻微到严重的一系列严重程度。同样的精确绘图程序被应用于37名高度采样的健康对照,每个受试者在 12 个疗程中平均获得 327.49 分钟的 fMRI 数据。
通过目视检查可以立即明显看出,这些抑郁症患者的显着性网络明显更大,该显着性网络涉及奖励处理以及自主反馈和反应与内部目标和环境需求的有意识整合(图1a, b)。在这 6 个人中,有 4 个人的显着性网络扩大了两倍以上,超出了所有 37 名健康对照中观察到的范围(图 1c)。平均而言,相对于健康对照组的平均值,显着性网络占据的皮质表面多出 73%,从而产生了一大群 级效应。使用替代网络分区算法并且不使用全局信号回归复制了这种效应,表明它对方法学变化具有鲁棒性,并且不能用大脑解剖或结构的群体差异来解释或头部运动。
为了进一步验证这一发现,我们在三个抑郁症患者样本中重复了这一过程。效果重复三次(图 1c),同样具有中等到大的效果大小,在控制样本中的性别比例失衡时,无论是否进行潜在校正,该结果仍具有统计学显着性 现场或扫描仪引起的偏差。我们还评估了纹状体中显着网络的表示是否同样增加,这被认为与解剖学上明确定义的互连环有关,其中皮层投射到纹状体,纹状体通过丘脑间接投射回皮层,但发现组均值差异无统计学意义(图1d)。
抑郁症中的显着性网络扩张在密度图中也很明显(图1e),它传达了在每个皮质顶点或纹状体体素处具有显着性网络表示的个体的百分比。这些图证实了两组中皮质和皮质下表征的相似总体模式,与之前报告中的描述一致,但也表明抑郁个体的每个皮质区域的显着网络的边界经常从质心进一步向外延伸。例如,在前扣带皮层中,网络边界更向前移动到前皮层,而在外侧前额叶皮层中,网络边界更向前移动到额极(图1e)。因此,SIMD 样本中皮层显着性网络的扩张伴随着邻近功能系统的收缩。然而,特定的收缩模式并未在所有数据集中复制——我们将在下一节中回顾这一发现。否则,网络大小的一致且可重复的组差异特定于显着性网络。
为了更好地了解这种效应是否也可以在涉及传统单回波 fMRI 数据的大型、先前发布的样本中检测到,我们从涉及 n = 812和 n 的两个大型数据集中的组平均功能连接数据中确定了显着性网络 分别有 120 名健康对照者,以及与神经调节干预研究相关的扫描涉及 n = 299 名难治性抑郁症患者的第三个数据集48。与两个健康对照样本相比,299 名抑郁症受试者样本中显着性网络的皮质表征要大 70% 以上。此外,在每个 SIMD 数据集的分半分析中产生了高度相似的显着网络拓扑和功能连接模式,表明显着网络扩展是这些高度采样个体的大脑的稳健且可重复的特征。
考虑到图 1c 中报告的影响程度,我们继续测试是否可以仅使用每个功能网络的大小作为预测特征,通过算法将抑郁症患者与健康对照者区分开来。因此,我们训练了一个线性支持向量机分类器,根据所有 20 个功能网络的大小区分抑郁症患者和健康对照个体,汇集从 5 个不同扫描仪获取的 n = 37 名健康对照者和 n = 141 名患有抑郁症的个体的数据。从两个不同制造商的两个不同扫描仪获取的抑郁症。总体而言,支持向量机分类器以 78.4% 的准确率(图 1f)正确区分抑郁症病例和健康对照,正确识别了 82.5% 的抑郁症病例,阳性预测值为 89.5%。通过检查线性预测系数并计算排除后精度的变化来评估特征重要性(图1g,h)。正如预期的那样,显着网络大小是最显着的特征。总之,这些分析表明,在大多数抑郁症患者中,显着性网络显着扩展,效应量大,在涉及不同数据采集和分析程序的多个样本中可重现,并且其量级足以支持高准确率的个体分类。机器分类器根据所有20个功能网络的大小区分抑郁症患者和健康对照个体,汇集从五个不同扫描仪获取的n = 37名健康对照者和从两个不同扫描仪获取的 n = 141名抑郁症个体的数据 不同制造商。总体而言,支持向量机分类器以78.4%的准确率(图 1f)正确区分抑郁症病例和健康对照,正确识别了 82.5% 的抑郁症病例,阳性预测值为 89.5%。通过检查线性预测系数并计算排除后精度的变化来评估特征重要性(图1g,h)。正如预期的那样,显着网络大小是最显着的特征。总之,这些分析表明,在大多数抑郁症患者中,显着性网络显着扩展,效应量大,在涉及不同数据采集和分析程序的多个样本中可重现,并且其量级足以支持高准确率的个体分类。
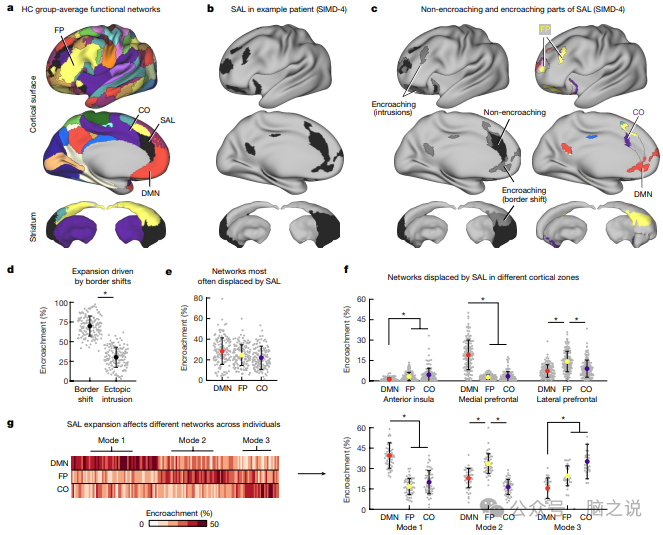
图2 抑郁症中显着网络扩展的三种模式
3. 三种显着网络扩展模式
功能性大脑组织的个体差异以两种形式出现:异位入侵,即在非典型位置观察到功能网络的孤立部分,以及边界转移,即网络的边界扩张(或收缩)并侵犯其邻居。边界转移是可遗传的,并且与已知的由遗传程序控制的皮质扩张机制相关,这些遗传程序在发育过程中以及经验或对环境影响的反应中细化功能区域之间的边界。此外,人类和非人类灵长类动物的宏观网络都是按照与基因表达和功能特性的皮质梯度相关的层次结构组织的,其中单模态感觉运动区域位于底部,异模态关联区域例如位于顶点的默认模式网络。因此,作为了解抑郁症中显着性网络皮质扩张的机制的一步,我们测试了它是否主要由边界转移或异位入侵驱动,以及它是否倾向于影响较低水平的单峰感觉运动网络或异模态网络。关联区域在此层次结构中的位置较高。为此,我们首先为 37 名健康对照生成了一个集中趋势功能网络图(图 2a)。其次,我们识别了 141 名抑郁症患者中每个人的显着网络部分,这些部分与健康对照组的组平均图中的显着网络不重叠,并将它们分类为异位入侵或边界转移(图 2b,c)。接下来,我们通过量化对每个其他功能网络的侵占程度来计算每个主题的侵占概况,定义为每个功能网络对显着网络侵占部分总表面积的相对贡献(图2c) 。
这项分析证实,显着性网络扩张不是随机分布的,相反,这主要是由于边界变化影响了三个相邻的高阶功能系统,在不同的个体中发生了三种不同的侵蚀情况。尽管显着性网络扩张涉及异位侵入和边界转移,但后者更为常见(图2d),并且两者都倾向于导致对默认的额顶叶或扣带-盖网络(图2e)的侵犯,而不是单峰感觉运动网络。与神经图谱工具箱中73 个独立的分子、微观结构、电生理、发育和功能脑图的比较表明,显着性网络扩张经常发生在皮质内髓磷脂较少的大脑区域,因此具有更大的突触可塑性。与 FC 重测可靠性图和时间信噪比图的进一步比较证实,这些大脑区域并不比机会更容易受到噪音的影响。同样明显的是,显着网络倾向于侵入不同皮质区域的特定功能网络(图2f)。例如,在外侧前额叶皮层中,显着网络向喙部扩展,并倾向于取代额顶叶网络。相比之下,在前扣带皮层和前岛叶皮质中,默认模式和扣带盖网络分别受到不成比例的影响。通过侵犯概况对个体进行聚类显示出三种不同的模式(图2g),主要涉及默认模式网络、额顶网络或额顶网络和扣带盖网络的组合。这种异质性可能部分解释了我们的观察结果,即显着性网络在所有三个数据集中一致扩展,但其他功能网络的相应收缩变化更大。
上述结果表明,显着性网络扩张主要是由额顶叶网络、扣带盖骨网络和默认模式网络的侵犯所驱动的,并表明网络之间边界的皮质空间可能被分配给抑郁症个体的不同功能系统。为了测试这一点,并进一步验证我们的发现,我们比较了显着网络的侵入节点(图2c左侧的深灰色顶点)和健康对照中通常占据该空间的功能网络之间的功能连接强度。该分析是使用每个人的静息态 fMRI 数据集的两半进行的,以评估与相对于亚军分配。正如预期的那样,侵入的显着性网络节点与其余显着性网络的功能连通性明显强于移位网络,这与连通性减弱的情况一致 入侵节点与通常在健康对照中占据该空间的功能网络之间的关系。总之,这些结果表明,额纹状体显着性网络扩张主要是由网络边界变化驱动的,这些变化影响三个特定的高阶功能系统并不影响其他系统,在三个亚组患者中发生不同的侵犯模式。
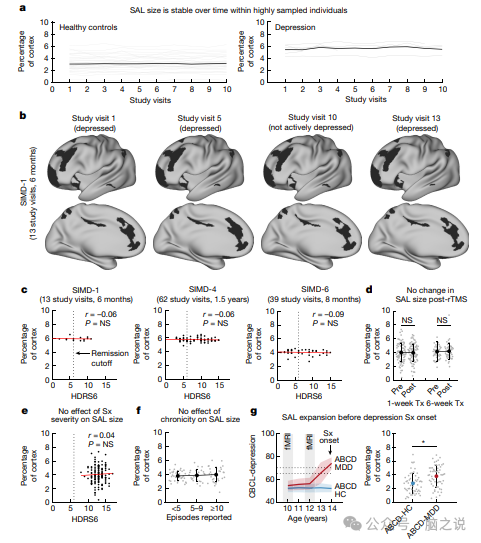
图3 显着性网络扩张随着时间的推移是稳定的,并且在症状出现之前就存在
4. 显着网络拓扑是特征状的
重度抑郁症从根本上来说是一种偶发性病症,其定义是在情绪正常的时期之间插入不连续的情绪低落时期。我们评估了显着网络拓扑的变化是否伴随着情绪状态转变期间发生的抑郁症状总体严重程度的变化——我们的纵向 SIMD 数据集非常适合测试这一假设。然而,与之前的研究一致,将健康成年人的功能网络拓扑描述为非常稳定的特征,几乎不受认知状态或日常变化的影响,我们发现,在患有和没有抑郁症的个体中,显着性网络拓扑随着时间的推移是稳定的(图3a)。此外,受试者内分析显示,在我们的 SIMD 数据集中的任何密集采样个体中,抑郁症状的波动与显着性网络大小随时间的变化之间没有显着相关性(图3b)。为了解决同一问题,我们利用传统的 6 周重复经颅磁刺激疗程或加速的、为期1周的 rTMS 强化课程(n = 45)。两个样本的显着性网络大小前后没有显着变化(图 3d)。此外,当前发作期间症状的严重程度(图3e)和个体报告的一生中经历的抑郁发作总数(图3f)都不能解释显着网络大小的个体差异。总的来说,这些发现表明,显着网络扩张是重度抑郁症患者的一个稳定特征,但不是抑郁发作的标志,并且与症状的严重程度或疾病的慢性程度无关。
这些观察结果使我们提出,显着网络扩张可能是患抑郁症风险的稳定标志,而不是随着时间的推移推动抑郁症状的变化。为了检验这一假设,我们询问显着性网络扩张是否存在于个体生命早期、抑郁症状出现之前。利用青少年大脑认知发展 (ABCD) 研究的数据,我们确定了 57 名儿童,他们在 10 岁和 12 岁时接受扫描时没有明显的抑郁症状,但随后在 13 岁时出现了临床上显着的抑郁症状(图 3g)。ABCD 研究中在任何时间点都没有抑郁症状的同等数量的儿童也被确定为对照样本。精确的功能映射显示,与没有进行 fMRI 扫描的儿童相比,在当前或既往没有抑郁症症状但随后出现了临床上显着的抑郁症症状的儿童中,显着网络平均占据的皮层多出 35.93%。任何研究时间点的抑郁症状(图 3g)。两个样本的基线和 2 年随访之间的显着性网络大小在 2 年内没有显着变化。在患有晚发性抑郁症的成年人中也观察到了类似的效果。总之,这些结果表明,显着网络的皮质扩张是大脑网络组织的一种类似特征的特征,它可以在数周、数月和数年内保持稳定,不受情绪状态的影响,并且在青春期抑郁症状出现之前的儿童中可以检测到。
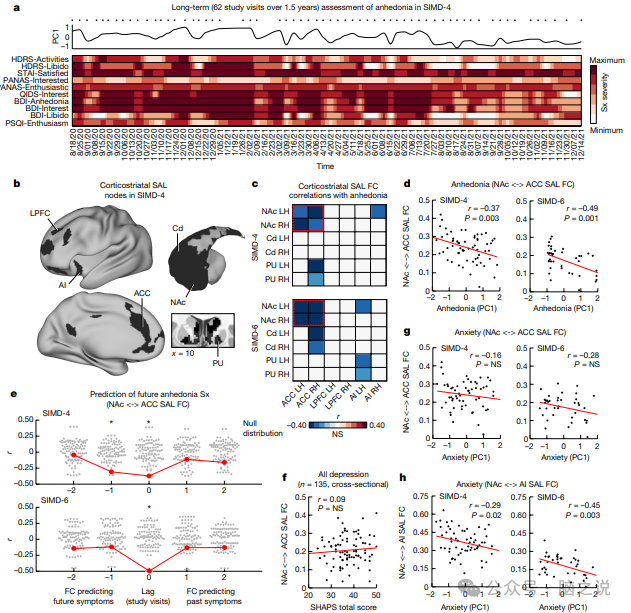
图4 额纹状体显着性网络连接可预测深度采样的抑郁症个体的快感缺失和焦虑症状随时间的波动
5. 连接状态预测快感缺乏
上述结果表明,显着网络的拓扑特征,例如其大小、形状和空间位置随着时间的推移是稳定的,并且不随情绪状态而波动。然而,这一观察结果并不排除特定显着网络节点之间的功能连接强度波动的可能性,并且这种波动有助于抑郁发作及其随后的缓解。为了测试这一点,我们首先询问显着网络节点之间的功能连接强度的变化是否与个体症状严重程度随时间的波动同时发生或预测,首先关注享乐功能,这是抑郁症的一个核心特征,与额纹状体环路,并且与显着网络和伏隔核-前扣带回环路在奖励处理和目标导向的努力行为中的假定作用一致。我们的分析重点关注SIMD 数据集(SIMD-4 和 SIMD-6)中的两名患者,临床医生在 8-18 个月的时间里对这两名患者进行了纵向重复扫描和评估,为该分析提供了足够的数据。这提供了一个机会,首次在单个密集采样个体的水平上提出问题——其数据有效地充当了独立的、强有力的 n-of-1 实验——大脑网络功能连接的变异性与特定的波动之间的关系。
我们从 SIMD-4 开始,因为该个体的研究时间最长并且拥有最多的 fMRI 数据,并保留 SIMD-6 作为复制数据集,我们观察到 10 项快感缺乏相关指标的显着波动(图 4a),这些指标源自 5 个标准化抑郁症状量表,并由三位研究共同作者的一致临床决策确定,范围从轻微或可忽略不计到严重。我们测试了显着网络节点之间功能连接的变化是否与该个体随时间的快感缺失变化相关,如图 4a 中十个快感缺失相关指标的主成分分析所示,并通过第一个成分得分进行总结。我们发现,几个皮质和纹状体显着网络节点之间的功能连接与快感缺失随时间的变化相关(图4c,d),其中伏隔核和前扣带皮层之间的连接影响最强。SIMD-6 中的一项相同分析复制了这种效应(图 4c、d)。当将每次研究访视时的头部运动作为协变量时,这一发现对于两个人来说仍然很重要。
接下来,我们询问显着网络功能连接是否可以预测未来研究访问中的症状严重程度,以及这种影响是否特定于快感缺失或扩展到其他症状领域。值得注意的是,一项交叉相关分析检查了过去、现在和未来研究访问中症状的相关性,表明伏隔核和前扣带回中的显着网络节点之间的功能连接不仅与当前的快感缺失症状相关,而且还预测了未来的出现或 快感缺乏症状在 SIMD-4 的下一次研究访问中得到缓解(图 4e),通常有大约 1 周的滞后时间。使用循环旋转的排列测试来保持时间自相关性证实了这种效应的重要性,表明在一次给定访问中伏隔核-前扣带回的连接预测了大约 1 周后未来的快感缺失,即使在控制了快感缺失测量随时间的相关性之后也是如此。值得注意的是,显着网络连接相关性在 SIMD-6 中针对当前症状进行了复制,但并未针对未来症状进行复制(图 4e),这可能与在此观察环境中抗抑郁治疗的差异有关。
为了确定伏隔核-前扣带回功能连接的变化是否不仅可以预测个体受试者随时间的快感缺失的变化,而且可以解释给定时间点快感缺失的个体差异,我们使用整个 n = 135 进行横断面重复这一分析。使用标准化的快感缺乏自我报告测量的复制受试者队列。然而,该分析并未显示前扣带回和伏隔核之间的功能连接的个体差异与个体之间的快感缺乏之间存在显着相关性(图4f),这强调了受试者内分析的价值。
最后,为了评估这种效应的特异性,我们询问伏隔核-前扣带回的连通性是否也与焦虑相关,焦虑是一种与抑郁症同时发生但通常与快感缺失无关的症状域。例如,在最近的一项研究中,“烦躁”(悲伤和快感缺失)和“焦虑”(焦虑和躯体)症状是相互分离的,该研究将 rTMS 干预的反应映射到不同的刺激部位。我们没有观察到任何一个人的伏隔核-前扣带回连接与焦虑之间存在显着相关性(图4g),这表明该回路在快感缺失中发挥着更重要的作用。值得注意的是,有几项神经影像学和回路生理学研究表明岛叶与焦虑的表达和厌恶状态的处理有关。受这项工作的启发,我们进行了一项类似的分析,询问每个受试者中纹状体与显着网络前岛叶区域的连接性变化是否与焦虑症状随时间的波动相关。根据我们的预测,我们发现纹状体与前岛叶的连接与 SIMD-4 中的焦虑症状显着相关,并在 SIMD-6 中复制了这种效应(图 4h)。总的来说,这些发现表明,尽管显着性网络稳定 该网络在抑郁症患者中扩展,并且这种扩展似乎发生在生命早期,该网络中的额纹状体连接性也会随着时间的推移而波动,并且纹状体与前扣带回和前岛叶的连接性变化分别跟踪快感缺乏和焦虑症状的出现和缓解 。
6. 解释拓扑的差异
在这项工作中,深度采样的抑郁症个体的精确功能图谱显示,显着性网络显着扩展,该网络在多个样本中是稳健且可重复的,相对于之前报道的抑郁症神经影像异常,具有中等到大的效应大小。这种效应主要是由网络边界变化驱动的,网络边界变化侵入了三个特定的功能系统——额顶叶网络、扣带盖骨网络和默认模式网络——在不同的个体中具有三种不同的侵入模式。这种效应随着时间的推移而稳定,对情绪状态或抑郁发作的标志不敏感,并且在儿童的早期出现,并在青春期后期出现抑郁症状。与此同时,纹状体与显着网络的前扣带回和前岛叶节点的连接性变化分别追踪快感缺乏和焦虑的出现和缓解,并预测一个人的享乐功能未来的变化。值得注意的是,我们的分析受益于精确功能映射与大量高质量、密集采样的多回波 fMRI 数据的结合使用,这对于精确映射网络拓扑中的个体差异可能至关重要和 这可能部分解释了为什么这些发现以前没有在文献中报道过。
尽管需要做更多的工作来阐明抑郁症显着性网络扩张的机制,但本报告和其他研究的主要结果至少指出了两个假设。首先,来自多个来源的证据表明,网络拓扑的个体差异受到活动依赖机制的调节,并且与给定网络的活跃使用程度有关。到目前为止,大多数评估人类或其他动物功能区域或网络大小变异性的研究主要集中在运动和视觉系统上。这些研究表明,不同的身体部位在初级运动皮层(M1)中具有不同的表征,其大小不同,并且皮质表征与相应肢体的灵活性密切相关,使得上肢比下肢占据更多的皮质表面积。以四肢为例,运动训练可以增加 M1 中训练过的肌肉或肢体的表现,而肢体截肢、石膏和先天性肢体缺陷都会减少废弃肢体的表现,并增加其他身体部位的表现。健康年轻人的初级视觉皮层 (V1) 总表面积最多可变化三倍,并且与视觉意识和对比敏感度的个体差异相关。同样,发现额顶叶网络的总皮质表征与儿童的执行功能能力呈正相关。总之,这些报告表明,显着网络扩张——伴随着额顶叶、扣带盖或默认模式网络的相应收缩——可能反映了抑郁症患者皮质区域和信息处理优先级的重新分配,这反过来又可能导致改变 显着性网络功能,例如内感受意识、奖励学习、自主信号处理和努力评估。
其次,汇聚的数据表明,皮质网络拓扑不仅受到外部调节、活动依赖机制的强烈影响,而且还受到内在遗传程序的强烈影响。许多转录因子调节细胞粘附分子,在发育过程中在整个皮质片上表现出强烈的表达梯度。删除这些模式因素可能会导致功能区域的收缩或扩张。相反,Emx2 表达的增加会增加 V1 的大小并减小躯体运动区域的大小。尽管我们的研究结果没有直接回答这个问题,但至少三个观察结果与内在发育遗传程序的作用一致,而不是完全依赖于活动的机制。首先,无论个人当前的情绪状态如何,显着性网络扩张都是高度稳定的,这表明网络活动中与情绪相关的剧烈变化不会影响网络规模。其次,显着性网络的扩张在生命早期就出现了,这与发育调节机制是一致的。第三,显着网络扩展是由空间组织的边界转移驱动的,已知这种边界转移是可遗传的,并且往往会在特定方向上侵入邻近网络,向前扩展且不成比例地针对高阶异模态关联皮层,尽管不影响单峰感觉运动区域。
6. 抑郁症中的特质与状态效应
我们的研究结果还可能为解决两个基本挑战开辟新途径,利用临床神经影像学研究的见解来重新思考我们诊断和治疗抑郁症的方法。首先,如上所述,跨越二十年的 MRI 研究已经确定了解剖学和功能连接的改变,这些改变在大规模荟萃分析中是稳健且可重复的,但在效应大小适中的受试者之间差异很大,这使得利用这些效应用于临床目的的效应变得复杂。相比之下,在我们样本中的大多数抑郁症患者中观察到显着性网络扩张,通过目视检查很明显,并且与中到大的效应大小相关。这种效应是在没有对位点或扫描仪引起的偏差进行校正的情况下检测到的,这对于多位点神经影像数据来说可能是一个重大的混淆。
多个医学领域的生物标记物有不同的形式,其中一些对当前症状敏感,而另一些则是稳定的疾病特征标记或出现症状的风险标记。我们的研究并不是为了全面验证抑郁症的神经影像生物标志物,未来的工作需要评估我们的研究结果相对于其他形式的精神病理学的特异性或评估其潜在的临床效用。初步分析表明,两个患有双相情感障碍但不是自闭症谱系障碍或强迫症的个体的显着网络也比正常人大,这可能反映了行为领域的常见缺陷,例如奖励处理,这也与显着性网络功能有关。然而,我们的结果确实表明,显着性网络扩展有可能帮助预测对抑郁症状的易感性,并且可能对设计治疗性神经调节干预措施产生重要影响,由于网络拓扑的个体差异,神经调节干预措施可能会产生广泛不同的影响。另一个需要记住的警告是,精确可靠地绘制抑郁症患者的显着性网络和一致地检测显着性网络扩展可能需要每个受试者 1.5-2 小时的高质量 fMRI 数据,这可能 这成为对传统功能磁共振成像数据集进行回顾性分析的障碍,这些数据集未针对个体层面的精确功能映射进行优化。
最后,我们的研究提供了原理验证数据,以支持使用精确功能映射和深度纵向采样来了解抑郁症临床神经影像研究中的因果关系。我们的分析显示,显着性网络拓扑结构存在稳定的、类似特征的差异,这些差异不仅与抑郁症相关,而且在没有抑郁症病史的儿童的早期就会出现,并预测随后在青春期出现抑郁症状。同时,他们展示了特定显着网络节点之间功能连接强度的变化如何随着时间的推移跟踪个体特定症状域功能障碍的出现和缓解,并且在至少一个个体中,至少预测未来快感缺乏症状的出现 发生前 1 周。通过这种方式,他们展示了密集采样和纵向设计将如何为理解因果关系以及设计个性化的预防性治疗开辟新的途径。
参考文献:Frontostriatal salience network expansion in individuals in depression.


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









