







文献介绍

文献题目: Cell2location maps fine-grained cell types in spatial transcriptomics
研究团队: Omer Ali Bayraktar(英国威康桑格研究所)、Oliver Stegle(英国威康桑格研究所)
发表时间: 2022-01-13
发表期刊: Nature Biotechnology
影响因子: 68.1(2022年)
DOI: 10.1038/s41587-021-01139-4
摘要
空间转录组学技术有望解决健康和疾病中组织的细胞线路图,但原位细胞类型的全面定位仍然是一个挑战。在这里,作者提出了 Cell2location,一个贝叶斯模型,它可以解决空间转录组数据中的细粒度细胞类型,并创建不同组织的全面细胞图。Cell2location 解释了变异的技术来源,并借用了不同位置的统计强度,从而使单细胞和空间转录组学的整合具有比现有工具更高的灵敏度和分辨率。作者在三种不同组织中评估了 Cell2location,并显示了细粒度细胞类型的改进映射。在小鼠大脑中,作者发现了横跨丘脑和下丘脑的精细区域星形胶质细胞亚型。在人类淋巴结中,作者用空间图谱绘制了一个罕见的 pre-germinal center B cell 类群。在人类肠道中,作者解决了淋巴滤泡中的精细免疫细胞群。总的来说,作者的研究结果将 Cell2location 作为一种通用的分析工具,以全面的方式绘制组织结构。
前言
组织的细胞结构(其中不同的细胞类型在空间中组织)是细胞间通讯、器官功能和病理学的基础。新兴的空间转录组学技术为以可扩展的方式原位绘制驻留细胞类型和细胞信号传导提供了关键机会。尽管这些技术已有概念验证应用,但定义空间转录组学工作流程以全面绘制组织中的常驻细胞类型仍然是一个挑战。原因之一是各个器官之间存在巨大的细胞多样性,包括无数细粒度细胞类型,例如免疫、基质和神经元亚群。这些精细细胞类型以微妙的转录差异为标志,通常在组织中特异化,并产生巨大的发现潜力。另一个原因是空间组织结构的多样性,从具有不同细胞类型的离散解剖区域的大脑到具有细粒度细胞类型的动态修改微环境的免疫器官。为了利用空间转录组学创建组织的全面细胞接线图,有必要建立足够灵敏的实验和计算方法,以解决不同组织中细胞类型的细粒度差异。
生成耦合单细胞和空间分辨转录组学的策略提供了一种可扩展的方法来应对这些挑战。关键原则是首先根据分离组织的单细胞 RNA 测序 (scRNA-seq) 识别驻留细胞类型,然后根据空间转录组图谱将识别出的细胞类型原位映射到其组织位置。在这里,空间 RNA 测序技术,例如 Visium、HDST 和 Slide-seq,其中使用微阵列或磁珠阵列网格从薄组织切片中定位捕获 mRNA,以高通量提供转录组范围的数据,并依赖于简单的组织学和分子生物学方案。
现有基于网格的分析方法的一个关键限制是缺乏单细胞分辨率,因为空间 RNA-seq 测量结合了多个细胞(Visium 和 Tomo-Seq)或多个细胞的一部分(Slide-seq 和 HDST)在每个组织位置。这一挑战可以通过空间 RNA-seq 与从耦合的 scRNA-seq 表达谱文件中获得的细胞类型的参考转录组特征的计算整合来克服。尽管最近针对此任务提出了几种计算方法,但尚不清楚这些方法是否提供足够的灵敏度来解析复杂组织中的细粒度细胞类型,这对于充分利用空间转录组数据的发现潜力至关重要。此外,需要考虑不同的混杂变异来源,这使得由多个实验组成的数据整合任务变得更加复杂,包括跨组织位置的可变细胞数量以及跨细胞类型和单个细胞的可变总 mRNA 含量。
为了解决上述挑战,作者提出了 cell2location,这是一种贝叶斯模型,旨在解析空间转录组数据中的细粒度细胞类型并创建不同组织的细胞图谱。Cell2location 可以处理复杂的实验设置,对多个 scRNA-seq 和空间转录组数据集进行联合分析。该模型的独特之处在于采用分层设计,借用跨位置的统计强度,从而提高分辨率和灵敏度,特别是在解析复杂组织中的细粒度细胞类型时。在这里,作者评估了三种不同组织中的细胞定位,在这些组织中,始终显示出绘制细粒度细胞类型的灵敏度有所提高。此外,作者绘制了人类淋巴结中罕见的 B 细胞状态图,并识别了小鼠大脑中的区域星形胶质细胞亚型。
研究结果
1. Cell2location:一个用于细胞类型空间映射的贝叶斯模型
Cell2location 通过整合单细胞或单核 RNA-seq 和给定组织的空间转录组数据来绘制细胞类型的空间分布(Fig. 1a)。cell2location 工作流程的第一步是估计来自单细胞和单核 RNA-seq 谱的参考细胞类型特征,这些特征对应于使用传统聚类或其他方法识别的一组用户提供的细胞类型和亚群的基因表达谱。默认情况下,cell2location 在该步骤中使用负二项式回归,这允许跨技术和批次的数据进行稳健组合(Methods and Supplementary Fig. 1)。第二步,cell2location 使用这些参考特征和一个或多个空间转录组数据集作为输入,将各个空间位置的 mRNA 计数分解为参考细胞类型(Fig. 1a and Supplementary Fig. 1)。
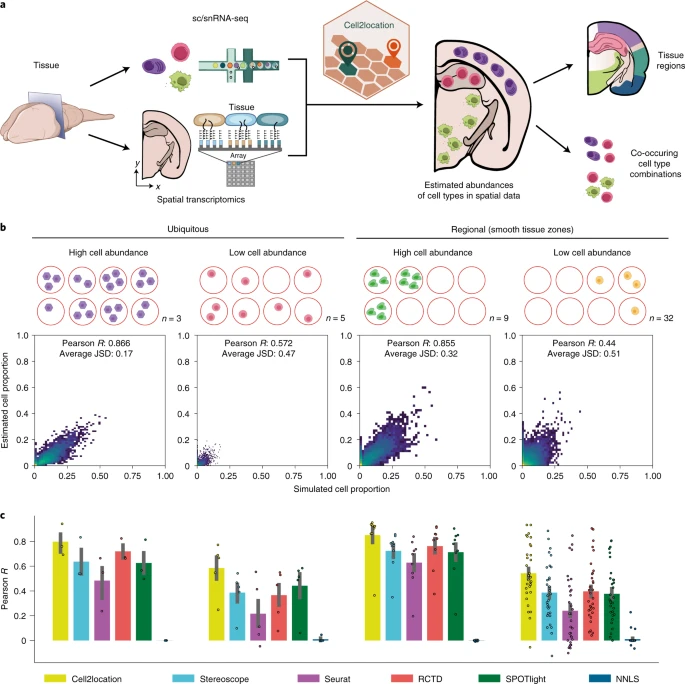
a. cell2location 的空间映射方法和分析工作流程概述。从左到右:scRNA-seq 和 spa
 最低0.47元/天 解锁文章
最低0.47元/天 解锁文章




















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言











