







涌现主义/突现主义 (Emergentism)
在哲学、系统论、科学和艺术中,当一个实体被观察到具有其所有组成部分本身没有的属性时,涌现 (emergence) 就出现了。这些属性或行为只有当各个部分在一个更广泛的整体中相互作用时才会涌现。
换言之,整体是和部分具有一样重要的作用,在一个整体中每个部分的相互作用、联系和每个单独的部分一样重要。例如,当一辆自行车和骑手互动时,平稳的向前运动就出现了,但是两个部分都不能独自产生此行为。
本来是哲学上的观点,涌现主义用在这里形容大脑连接也很恰当,本文的中心思想也就是各部分的大脑连接(connections) 结合在一起也会产生和整体一样重要的作用。
目录
一、大脑连接整合
越来越多的人认为大脑功能是大脑连接所引起的一种属性(而不是传统的各个/多个脑区单独的作用),因此研究大脑活动需要整合(原文中为integrative一词,本文中都译为整合)这些大脑不同区域之间的连接。白质连接将很多大脑区域串联在一起,根据不同的功能使用不同的尺寸以及髓鞘量身定制网络回路。所以可以通过研究(脑叶内与脑叶间、半球内与半球间)白质连接来帮助整合大脑不同区域之间的联系(图一)。

图一:不同的大脑模型决定了大脑状态(功能)的灵活性。(A)在模块化系统(Modular system) 中,一个区域在没有串扰的情况下执行一个功能,并且大脑状态的数量随着区域数量线性增加。(B) 在层次系统 (Hierarchical system) 中,功能的激活与区域的顺序有关。因此,例如重复读单词的任务将依赖于从听觉到运动过程中颞叶—顶叶—额叶的传播。与(A)和(B)相比,(C) 整合模型 (Integrative model)提供了最高的计算灵活性,考虑到我们所知道的语言过程的高度复杂性和灵活性(例如听写、默写这样很接近的任务也可能有不同的大脑状态)。每个模型都可以在大脑区域(X, Y, Z)的组合下转换成不同的大脑状态模式 (D中白色表格),即ABC三个区域可以任意激活,不像模块化系统只能每一个区域单独的激活,以及分层系统有顺序的激活。(E) 大脑区域数目(横轴)与不同大脑模型所能表达的的大脑状态数目(纵轴)的关系。说明在相同的大脑区域数量下,大脑整合模型能表达出更多数量的大脑状态,为拟合更加精细/灵活的功能打下基础。
二、测量活体大脑中的连接
如今人们使用非侵入神经影像来测量大脑连接。结构上,测量沿轴突方向的水扩散可以推导出诸如白质通路(即纤维束)密度、直径和分散度等属性。功能上,测量不同区域活动之间的一致性来推断大脑区域之间的活动。再将数学模型应用到功能磁共振fMRI结果上,可以估计大脑区域之间交互的强度(即功能连通性)或方向性(即有效连通性)。
三、大脑连接的整合功能
大脑连接不仅仅可以传递大脑区域之间的信号,还可以放大或减少信号,因此决定大脑皮层的结构和功能。人们休息时人们大脑区域之间的通信和活动存在相似性,根据白质的皮层投射可以预测某个功能在大脑中的何处出现(图二A),该种预测证明用于识别视觉词形区域,甚至在获得识字能力之前。
通常,大脑连接的主要趋势可以捕获大脑功能的空间组织。如果一个大脑区域失去了连接,它剩下的树突和突触就会被修剪,神经元就会枯萎或死亡。因此,该区域的网络在功能上(dysconnection)和结构上(disconnection)都出现了故障,不再能发挥某种功能。这种现象,也被称为一种失联(diaschisis),证明了连接在维持较远大脑区域的完整性及其功能方面的重要性。
最后,通过手术交换感觉皮层之间连接的研究进一步证明了白质连接在大脑功能组织中的主导性和决定性作用。具体来说,将视觉输入强加于听觉皮层会使听觉皮层获得很多视觉行为的细胞和功能性质。这表明大脑区域之间的相互连接和作用可以影响细胞性质。这样的功能是大脑整合机制的新特性。因此这些证据支持大脑连接决定大脑功能和认知这一潜在机制。
从仅仅考虑单独的脑区到功能网络的转变,需要考虑到白质连接对功能网络的影响。因此高分辨率的纤维追踪结果可以系统地揭示大脑激活网络,例如最近的一个功能白质图谱的成果整合了连接的功能磁共振(fMRI)的统计数据和扩散磁共振(dMRI)的结构数据(图2B)。这个图谱的结果也反映了左右半球功能已知的颗粒度(granularity) (指细节水平)之间的不对称性。这是一种人们认知的不平衡,所以我们需要对一直以来认为的非优势右半球做更多的研究。
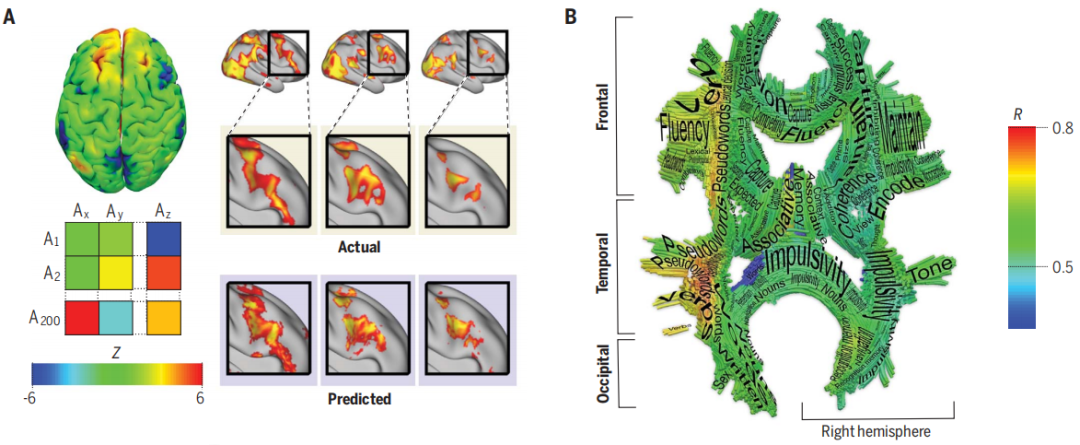
图二A-B:(A) 功能连接(左上)可以总结为每个个体特定的功能连接矩阵(左下) A代表区域(Area)。(右)功能连接的主要趋势可以预测任务相关激活的个体模式。(B) 可以将fMRI的功能信息投射到大脑的结构连接(白质)上,从而使结构连接可以预测相关任务(例如图上语言流利度、语音语调)的皮层激活模式。
半球偏侧化是由半球间的连接引起的。一些皮质区域在认知功能例如语言、感知、行动、情感和决策中的半球间的不对称性增加,这些皮质区域通过胼胝体与对侧半球的连接较少(图2C)。扩散加权成像(DWI)揭示了胼胝体结构受大脑尺寸以及物种的影响,轴突传导特性因此受到改变,例如轴突直径与半球间传导速度相关(图2D)。此外,不同物种间的半球内、半球间的连接数量不同。综上所述,随着物种进化,大脑尺寸的增加可能导致偏侧化的形成,旨在避免形成一个不成比例的胼胝体(连接数量增多,口径不变,胼胝体大小增加),或半球间的传导太过延迟(胼胝体大小不变,连接数量增多,口径变小)。
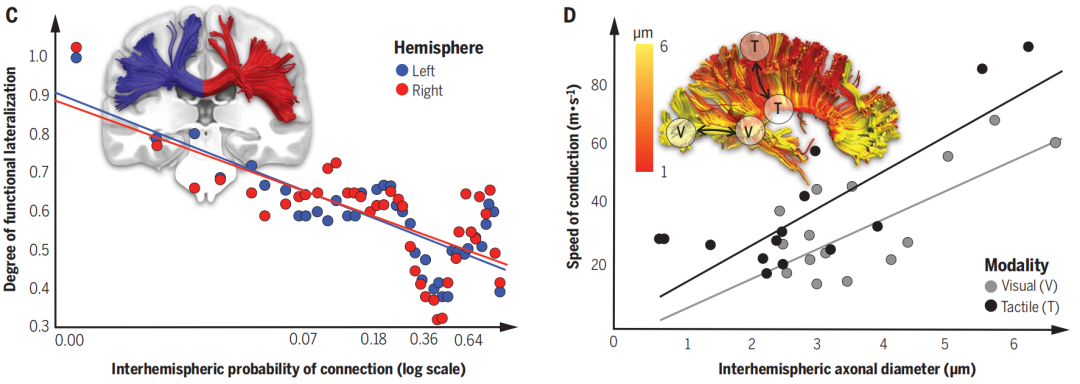
图二C-D:(C) 大脑半球功能偏侧和半球间连接的影响(纵轴是偏侧化程度,横轴是连接的数量,即连接越少的地方偏侧化程度越高)(D) 功能的传导速率(纵轴)与轴突直径(横轴)的关系,即轴突口径越大功能的传播速率越快(图中是以视觉功能(灰)和触觉功能(黑)为例)。
四、通过大脑连接认识进化
由于结构连通性可以用来破译大脑的功能组织,多项研究比较了不同的灵长类物种,以了解人类的独特性,并阐明其进化过程中的机制。例如,人类的语言能力与左半球弓状束的显著的扩张相关(图3A)。白质束(如弓状束或胼胝体)的解剖学描述允许提取跨物种的大脑连接特征, 因此可以计算物种大脑之间的变形场(deformation fields)(图3B)。(变形场就是一种矩阵映射,这里的变形场定义了物种之间的相似性和差异性)。
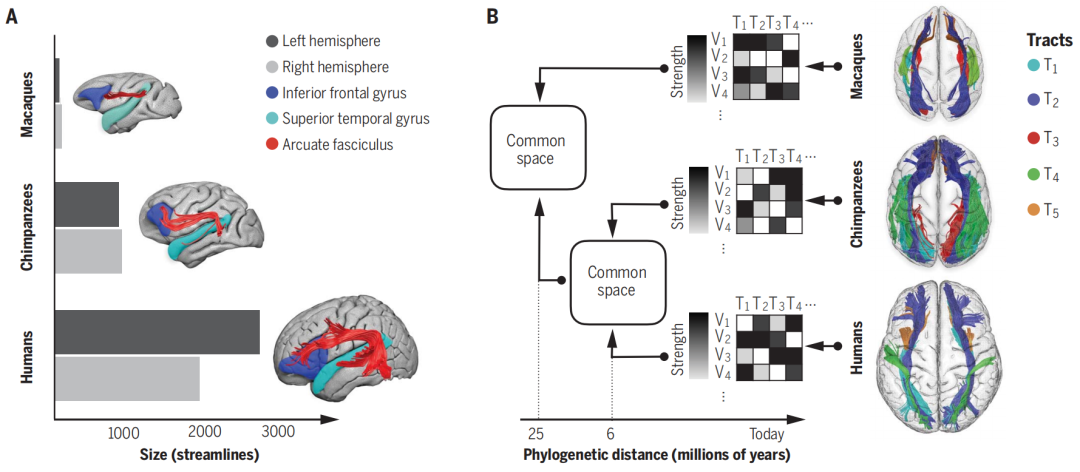
图三A-B:(A)对比人类、猩猩、猕猴在额叶和颞叶区域的连接揭示了弓状束的扩张(横轴上代表三种物种的纤维数量不同)和偏侧化程度(深灰-左脑、浅灰-右脑)。(B)提取跨物种的可靠连接性分布可以计算公共空间(common space)(通过文中所述变形场计算),从而找到我们祖先大脑的相似性。
比较研究(comparative studies)认为,物种之间的相似性可以追溯到一个共同的祖先,并解释了在进化过程中保留特定功能的原因。最近的比较工作揭示了第一个有关大脑区域的系统发育组织图谱。这种技术进步——神经影像之间的比较将更有针对性的去匹配人脑机制与其发育机制。这些进展也可能有助于发现和模拟动物的神经保护机制,可能会转化为改善人类疾病模型和治疗方法。例如,额顶叶断连是持续性视觉忽视障碍(persistent disorders of visual neglect)非常敏感和特异的生物标志物。大多数患有这种视觉缺失的人都无法从视觉忽视中恢复过来,而患有同样的视觉缺失的猴子则会在几天内恢复过来。因此,猴子有一种独特的机制,可以促进大脑的恢复。然而,这一机制尚未被确定,其对人类的可推广性也还需要进一步探索。
比较研究的一个常见局限性是每个物种使用的大脑数量很少(通常少于10个),这无法完全捕捉到个体间的差异。但大脑连接变化的数量(即大小)与大脑大小成正比,其变化模式在人类和其他灵长类动物之间是相似的。因此,最近进化的大脑区域在个体之间的差异越来越大,而进化较老的大脑区域往往更加稳定。因此,大脑连接的种内变异性可能是我们理解进化机制的一个新维度(图3C)。
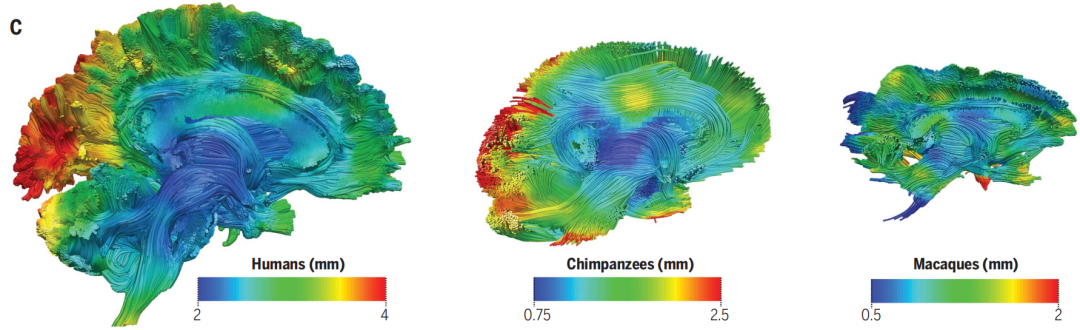
图三C:连接可变性(即不同物种匹配一个共同物种特定模板所需的白质平均变形程度)表明,使我们个体彼此不同的(连接)可变性也是我们与祖先和最接近的亲属物种之间差异的根源。
五、大脑连接揭示个体之间的差异
这种连接可变性,或称“神经可变性”,对我们的个体至关重要。我们所知道的,我们是谁,以及我们如何与他人交流,都归功于大脑的整合机制。因此,为了理解我们身份的起源,我们需要在个人层面上破译大脑区域之间的连接是如何协调我们的大脑功能的。从功能连接中获得的大脑区域之间的通信强度可以预测任务期间大脑激活的个体差异(图2A)。已经有初步工作在健康志愿者中创建了与行为和认知相关的大脑功能连接模式的“指纹” [即基于连接组的预测模型]。这些大脑连接的“指纹”对每个人来说都是特定的,并且能够非常准确地预测流体智力和创造力。这些与认知的相关性也与大脑连接结构的差异有关 (图4A)。例如,较强的左弓状神经束似乎有助于学习新单词。这些纤维束的差异影响了大脑功能,以及神经发育、精神和神经疾病症状的严重程度。发育中的大脑连接模式(即白质纤维连接的发育)有助于诊断神经发育的学习缺陷。
在神经学上,更强的弓状束有助于中风后的恢复,而其退化与症状严重程度的增加有关。在失语症(即语言障碍)被描述为一种分离综合征(disconnection syndrome)的150年后,这些观点引起了新的解剖—认知模型。
由于环境和学习诱导的可塑性机制,大脑连接可以在整个生命周期内发生变化,导致越来越不同的分子和电路水平的变化,这些变化在数周、数月、数年甚至数十年内发展。一个很好的例子就是即使是在晚年获得的读写能力,也可以改变大脑视觉和听觉系统之间的联系。但是可塑性机制会限制基于大脑连接“指纹”的长期认知预测。重要的是,虽然识别预测性生物标志物具有改变个人和人群健康的巨大潜力,但它也面临根本的伦理风险和道德挑战——例如,对于预测的大脑疾病,真正的治疗方案可能根本不会在大脑恢复/重塑的预测中表现出来。但总的来说,这些最近的研究提出了证据,证明新的行为模式和认知功能可以通过大脑连接的相互作用的微小变化而产生。
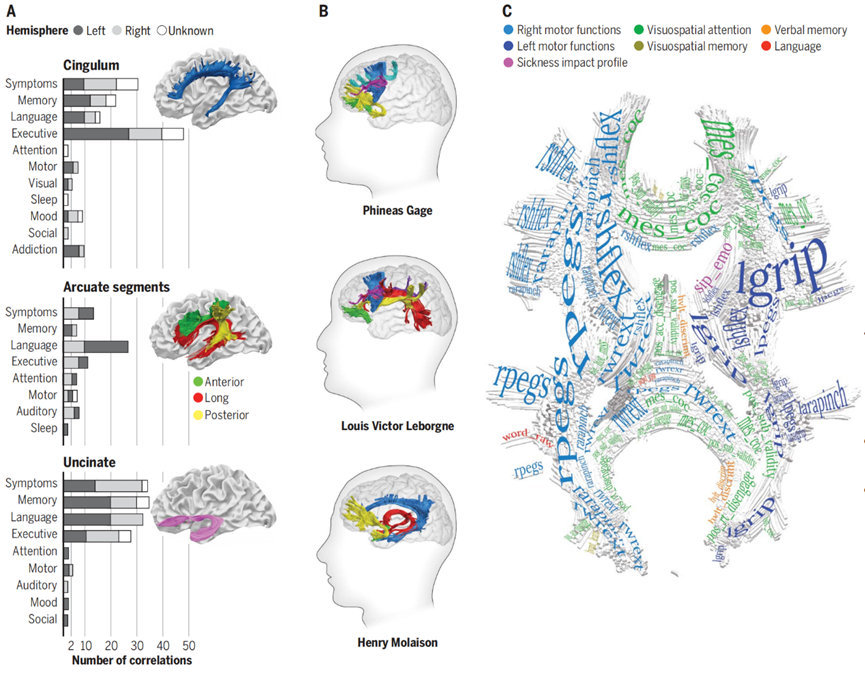
图四:(A)系统的回顾了不同大脑连接与认知功能、临床症状的关系。(B)连接重现了神经科学中最著名的神经科学案例,显示经典神经行为综合征(Neurobehavioral syndromes)中的神经纤维束断连。(C) 展示了断连导致功能损失的白质模板。
六、大脑连接断开导致功能损失
大脑相互作用的连接中断会导致继发性病理,并可诱发持久的功能性症状。例如,视觉和语言网络之间的断连会导致无法补救的失读症(alexia) [即无法阅读]。虽然先进的神经成像可以识别断连,但这些方法还没有系统地应用于临床领域。因此,需要使用更高分辨率数据集的连接先验知识(DWI或fMRI)来估计脑损伤后的断开情况,以可靠地绘制断开与症状之间的关系。这样做,就有可能重新评估脑网络中的经典临床神经解剖现象,并了解连接对功能实现的关键贡献。这些新方法甚至可以证明,神经科学中最著名病例中的临床解剖损伤研究可以扩展到连接断开的范式(disconnection paradigm)。这个新范式通过几个经典的连接断开案例研究了所影响的相关功能:情感和决策(Phineas Gage) (编者附图)、语言产生(Louis Victor Leborgne)和陈述性记忆(Henry Molaison)的网络(图4B)。
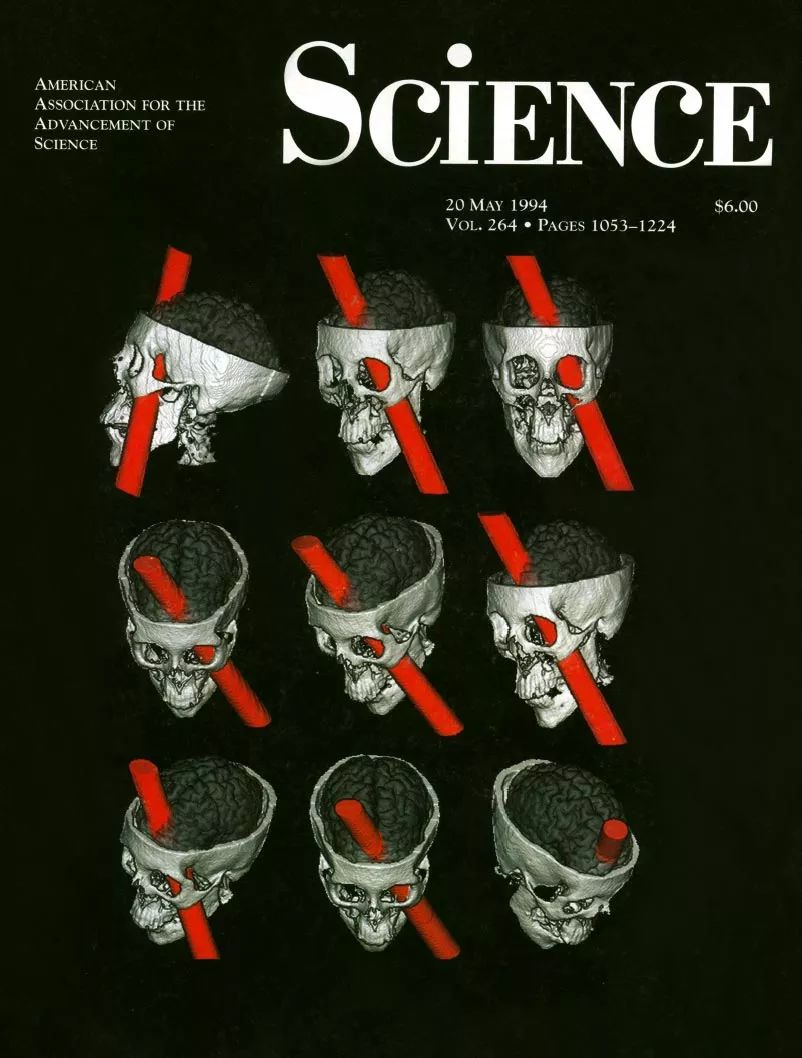
编者附图:1994年5月20日《科学》杂志264期封面。马西奥夫妇通过颅骨的测量和现代神经成像技术重建了事发的意外,确定了可能损伤的位置。他们认为:盖奇的左右脑都受到了创伤,从而导致了盖奇理性决策和情感处理的缺陷,并以此解释他们提出的情绪原理——情绪与理性互助互补。(菲尼斯·盖奇-Phineas Gage:1848年在他工作时炸药意外炸飞了他手中的铁夯,从他的左颧骨下方穿入头部,撕毁了他大脑左侧额叶的组织,从眉骨射出)
因此,考虑大脑连接似乎使健康志愿者的脑损伤研究与功能神经成像相协调,并且比起大脑过程解体(disintegration)的临床表现提供了更全面的生物学解释。此外,大脑连接断开结果通常比单独的病变定位具有更高的解释能力。连接断开的框架最近已扩展到整个大脑,以提供与特定大脑断开相关的症状的第一张临床地图(图4C)。因此,如果测量大脑连接被转化为关注康复的个性化神经科学的操作标准,支持症状恢复的预测,同时为药物治疗提供新的靶点,这将被证明是有益的。所提出的证据表明,理解大脑的关键在于多个区域之间的相互交互,而了解这些相互交互的本质就是大脑的连接基础。
七、未来展望
【技术上的局限】尽管目前在估计大脑连通性方面取得了进展,但新的挑战已经出现,我们需要依靠协同努力来解决。首先,目前还没有一种方法可以直接测量健康人脑中神经连接的激活程度。相反,最近的间接方法都是在统计上将皮层的功能信号投射到白质上。这些间接方法依赖于大脑区域之间连接在群体水平上的先验知识。然而,无论用于构建这些先验数据的数据集质量如何,常规神经成像在体内的最小分辨率为1mm3,死后大脑的最小分辨率为200mm,都不能完全达到轴突约为1至6微米的直径。同样,大脑中的单突触连接区域同步延迟仅仅2-3ms,然而,标准的功能连接依赖于时间分辨率最高只能达到1秒。因此,在估计纤维束方向和大脑区域之间的通信只能是一定程度的近似,这为准确估计连接组(即全脑连接)留下了改进的空间。
【缺乏验证集】同时也迫切需要补充的方法来提供一个实用的“金标准”和一种验证方法,类似于用于细胞结构测量和皮层厚度的BigBrain模型。非神经成像方法可能会让我们更接近这个目标。例如,高级偏光成像或nissl-based的结构张量可以从死后组织中估计轴突方向。然而,这些死后方法仅限于二维平面内重建,需要付出巨大的努力才能产生与BigBrain相当的整个3D人脑。初步研究已经表明,这种尝试在猕猴身上是可行的,并且在人类身上的努力正在进行中。这些新的(细胞学/组织学)数据集可以验证当前都来自神经成像的神经生物学近似值,例如轴突直径、神经突密度、髓鞘形成的程度、信息流或全脑水平的突触复杂性。这一步骤对于确保连通性矩阵可靠地转化为临床领域并导致治疗干预的进展至关重要。
展望未来,我们需要通过开发专用软件来巩固这些新概念:可以在相同的潜在空间中表示高维数据的工具,这些工具可以跨尺度(细胞、体素、电路)、跨物种(猕猴、黑猩猩、人类)和跨成像方式(纤维追踪、偏光成像)寻找解剖信息,同时考虑个体水平上的神经可变性来捕捉大脑连接之间的相互作用。研究人员应该建立专业的网络,整合想法,并公开分享数据。总之,这些努力将推动大脑连接研究整合各种成像模式(如白质功能),制定新的框架(如神经可变性),促进我们对大脑发育和进化的理解。这一共同努力将推动我们当前的研究,并促进更先进的神经成像方法,个性化解剖模型和临床上的影响。因此,需要一个综合的系统(研究联盟)来破译大脑连接的这个复杂系统,探索大脑连接的涌现性。
原文:The emergent properties of the connected brain

全文转自公众号AI4DMR


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









