










大家好,这里是专注表观组学十余年,领跑多组学科研服务的易基因。
水稻是全球重要的农作物,也是单子叶植物模型。在水稻中,N6-甲基腺苷(m6A)mRNA修饰对植物的发育和胁迫响应至关重要。OsFIP37作为m6A甲基化复合体的核心组分,其缺乏会导致雄性不育,强调了m6A在雄性生育中的重要性。m6A是一种可逆的修饰,可以被m6A去甲基化酶去除,但m6A去甲基化酶是否调控水稻的雄性育性尚未明确。
北京大学贾桂芳与中国农业科学院万建民院士团队合作研究鉴定出水稻RNA m6A去甲基酶(OsALKBH9),并表明其参与水稻雄性育性调控。相关研究成果于2024年4月17日以“OsALKBH9-mediated m6A demethylation regulates tapetal PCD and pollen exine accumulation in rice”为题发表在《Plant Biotechnology Journal》期刊。

标题:OsALKBH9-mediated m6A demethylation regulates tapetal PCD and pollen exine accumulation in rice(OsALKBH9介导的m6A去甲基化调控水稻绒毡层细胞死亡和花粉外壁积累)
时间:2024.4.17
期刊:Plant Biotechnology Journal
影响因子:IF 13.8 / 1区
实验方法:细胞学分析、转录组分析、m6A-seq分析等
研究摘要:
本研究鉴定出mRNA m6A去甲基化酶OsALKBH9,并发现其在参与雄性育性中的调控。OsALKBH9敲除导致水稻雄性不育,且这种雄性不育依赖于其m6A去甲基化活性。细胞学分析揭示了Osalkbh9-1突变体中调控花药绒毡层细胞死亡(Tapetal Programmed Cell Death, PCD)的缺失和花粉外壁(exine)的过度积累。花药的转录组分析显示,Osalkbh9-1突变体中与绒毡层发育、花粉壁合成和转运通路相关的基因表达上调。此外,研究人员验证了OsALKBH9可以直接去甲基化TDR和GAMYB转录本中的m6A修饰,影响了这些mRNA稳定性,并最终导致花粉外壁的过度积累。本研究结果强调了mRNA m6A修饰的精确调控,并揭示了OsALKBH9介导的m6A去甲基化在水稻绒毡层PCD和花粉外壁积累中发挥的关键作用。OsALKBH9的发现为杂交育种提供了有价值的雄性不育材料,为理解m6A在植物育性中的作用提供了新视角。
研究方法:
- 利用CRISPR/Cas9基因编辑技术在水稻品种Nipponbare中敲除OsALKBH9基因。
- 通过细胞学分析、转录组分析、m6A-seq分析等方法研究OsALKBH9的功能和作用机制。
- 利用体外去甲基化实验和m6A-IP-qPCR等技术验证OsALKBH9的去甲基化活性。
结果图形
(1)敲除OsALKBH9导致水稻雄性不育,且OsALKBH9在雄性育性中的调控依赖于其m6A去甲基化酶活性
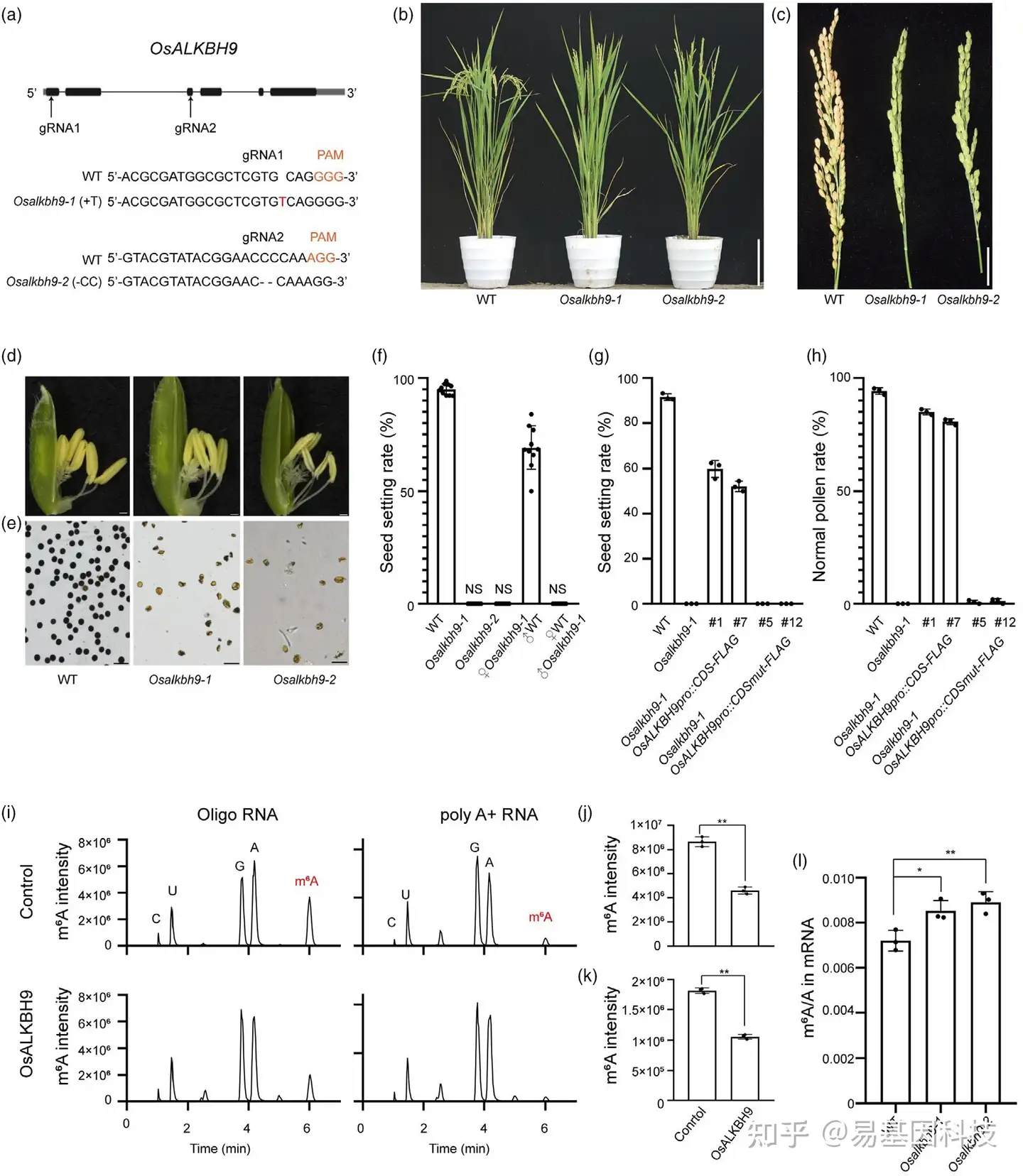
图1:OsALKBH9在调控水稻雄性育性中的作用及其m6A去甲基化活性的重要性
- 利用CRISPR/Cas9技术对OsALKBH9基因进行目标位点突变。
- WT、Osalkbh9-1和Osalkbh9-2突变植物抽穗后的表型。
- 成熟阶段WT、Osalkbh9-1和Osalkbh9-2的稻穗比较。
- WT、Osalkbh9-1和Osalkbh9-2植物的小穗。
- 用I2/KI溶液染色的WT、Osalkbh9-1和Osalkbh9-2的花粉粒。
- 突变体与WT植物杂交的种子设置率(n=10株植物)。
(g-h) 基因互补植物和去甲基化酶活性缺失互补植物的种子设置率(g)和I2/KI溶液中的花粉粒(h)(n = 3株植物)。
(i-k) 重组OsALKBH9蛋白在体外去甲基化含m6A的单链RNA(ssRNA)和聚腺苷酸+ RNA(poly A+ RNA),n = 3个生物学重复。(i) 体外去甲基化活性反应消化底物的LC-MS/MS色谱图。(j, k) 使用ssRNA (j) 和poly A+ RNA (k) 作为底物,在体外去甲基化活性反应后的m6A变化。
(l) 通过LC-MS/MS在14天龄的WT、Osalkbh9-1和Osalkbh9-2茎中纯化的mRNA中确定的m6A相对于腺苷的百分比(m6A/A比率)。n = 3个生物学重复。
(2)OsALKBH9是绒毡层PCD和花粉外壁积累所必需
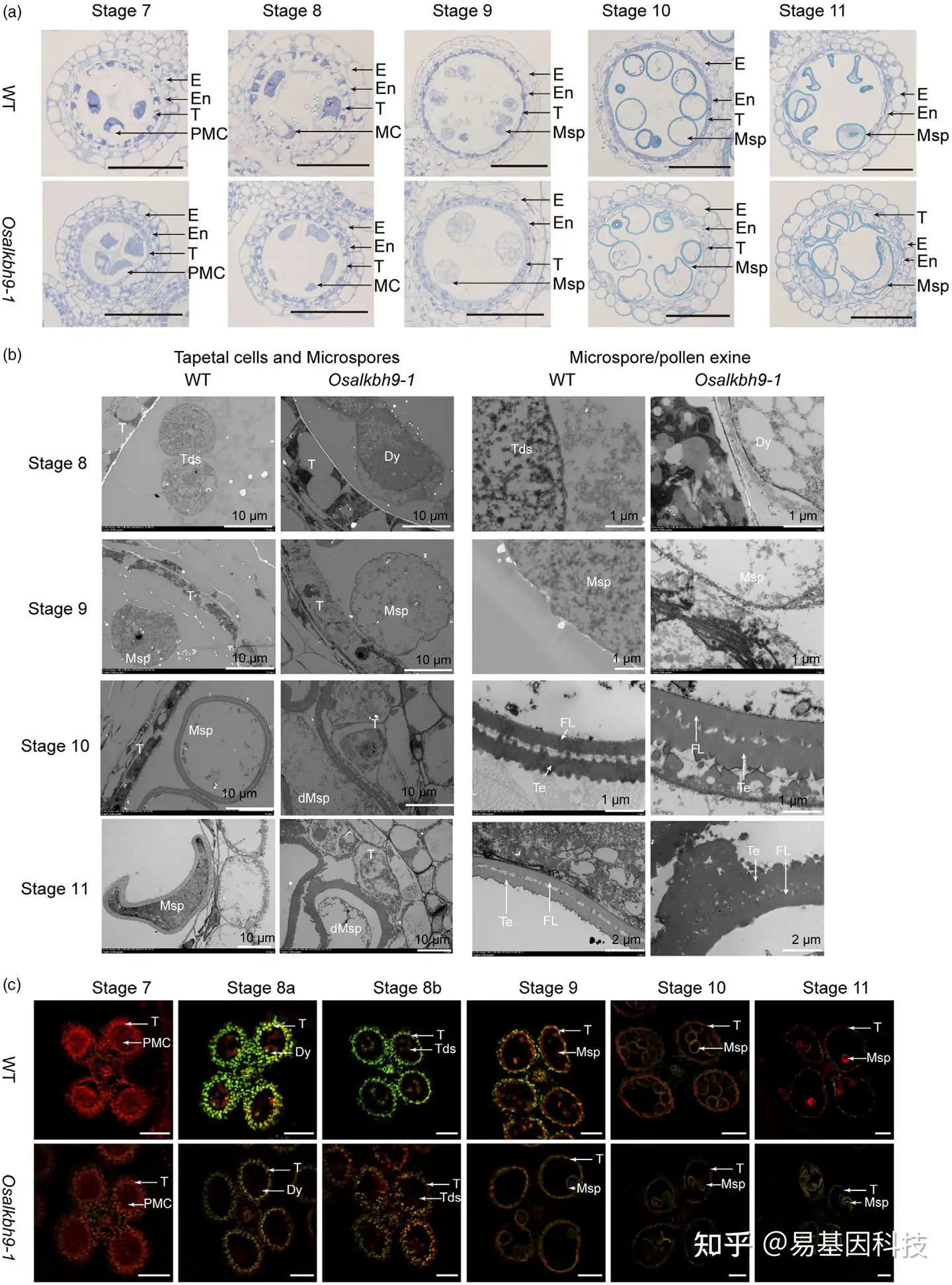
图2:OsALKBH9敲除导致绒毡层细胞降解延迟和花粉壁形态异常。
(a) 野生型和Osalkbh9-1突变体在花药发育阶段的7~11期的半薄切片。
(b) 野生型和Osalkbh9-1花药从第8~11阶段的透射电子显微照片。
(c) WT和Osalkbh9-1花药7~11期的TUNEL分析。
E:表皮;En:内皮层;T:绒毡层;PMC:花粉母细胞;MC:减数分裂细胞;Msp:小孢子;dMsp:败育的小孢子;Tds:四分体;Dy:二元细胞;FL:基座层;Te:外壁。
(3)OsALKBH9在花药中高表达,主要定位于细胞质中
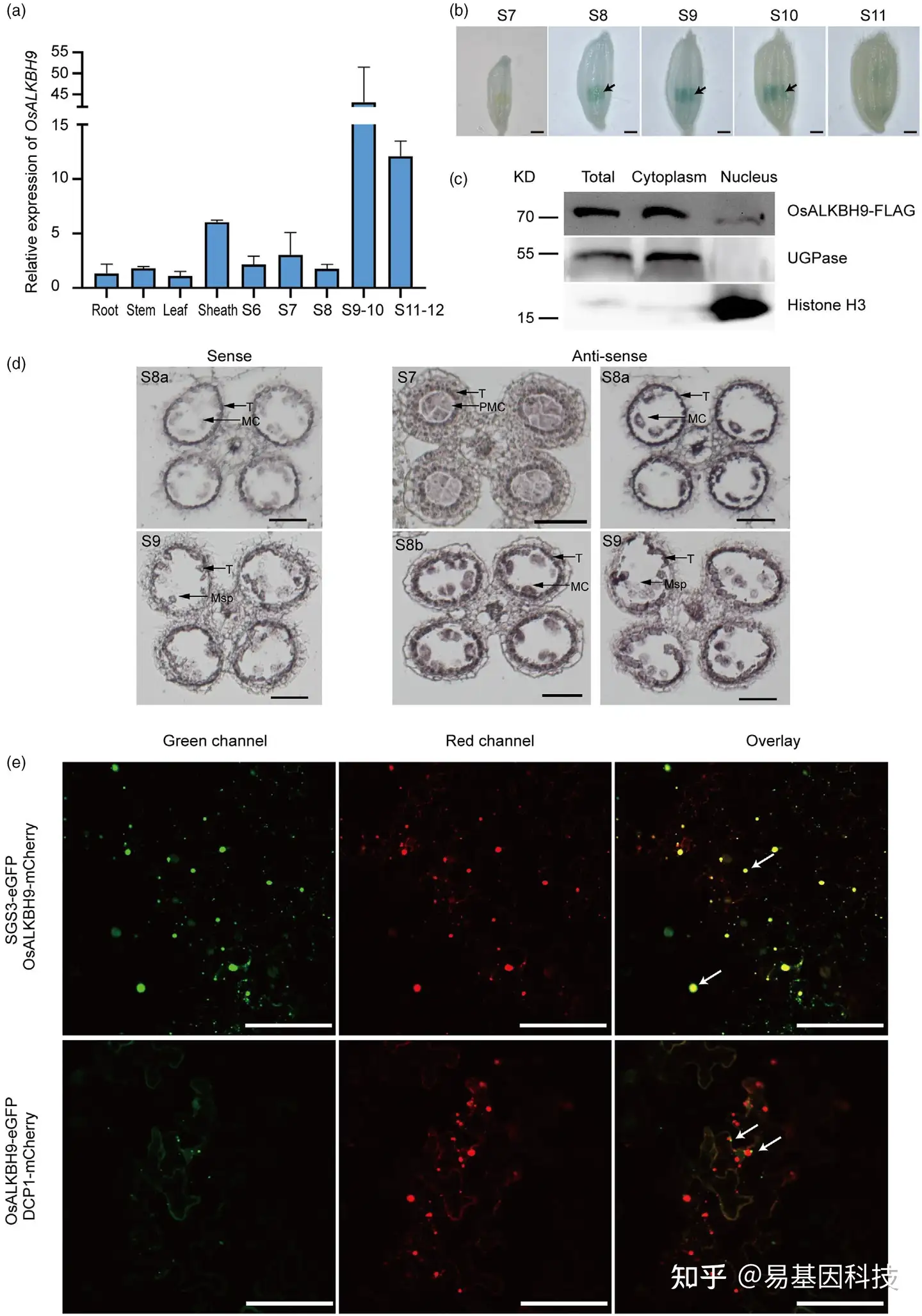
图3:OsALKBH9在花药中高表达,且主要定位于细胞质中。
(a) 通过qRT-PCR分析OsALKBH9在不同组织中的表达。在发育阶段6~12收集花药。其他组织在开花阶段的植物中收获。
(b) 转化了OsALKBH9启动子驱动的β-葡萄糖醛酸酶(GUS)报告基因的转基因花药进行GUS染色,以可视化OsALKBH9表达。
(c) 亚细胞组分和免疫印迹分析。对Osalkbh9-1 OsALKBH9pro::OsALKBH9CDS-FLAG转化体的总蛋白、细胞质和细胞核富集级分的SDS-PAGE。UGPase(细胞质标记)和组蛋白H3(细胞核标记)分别进行western blot。
(d) 野生型花药OsALKBH9转录本的原位杂交。
(e) OsALKBH9在本氏N.benthamiana叶表皮细胞中的亚细胞定位。
(4)OsALKBH9缺失影响与雄性育性和花粉外壁(exine)积累相关基因表达
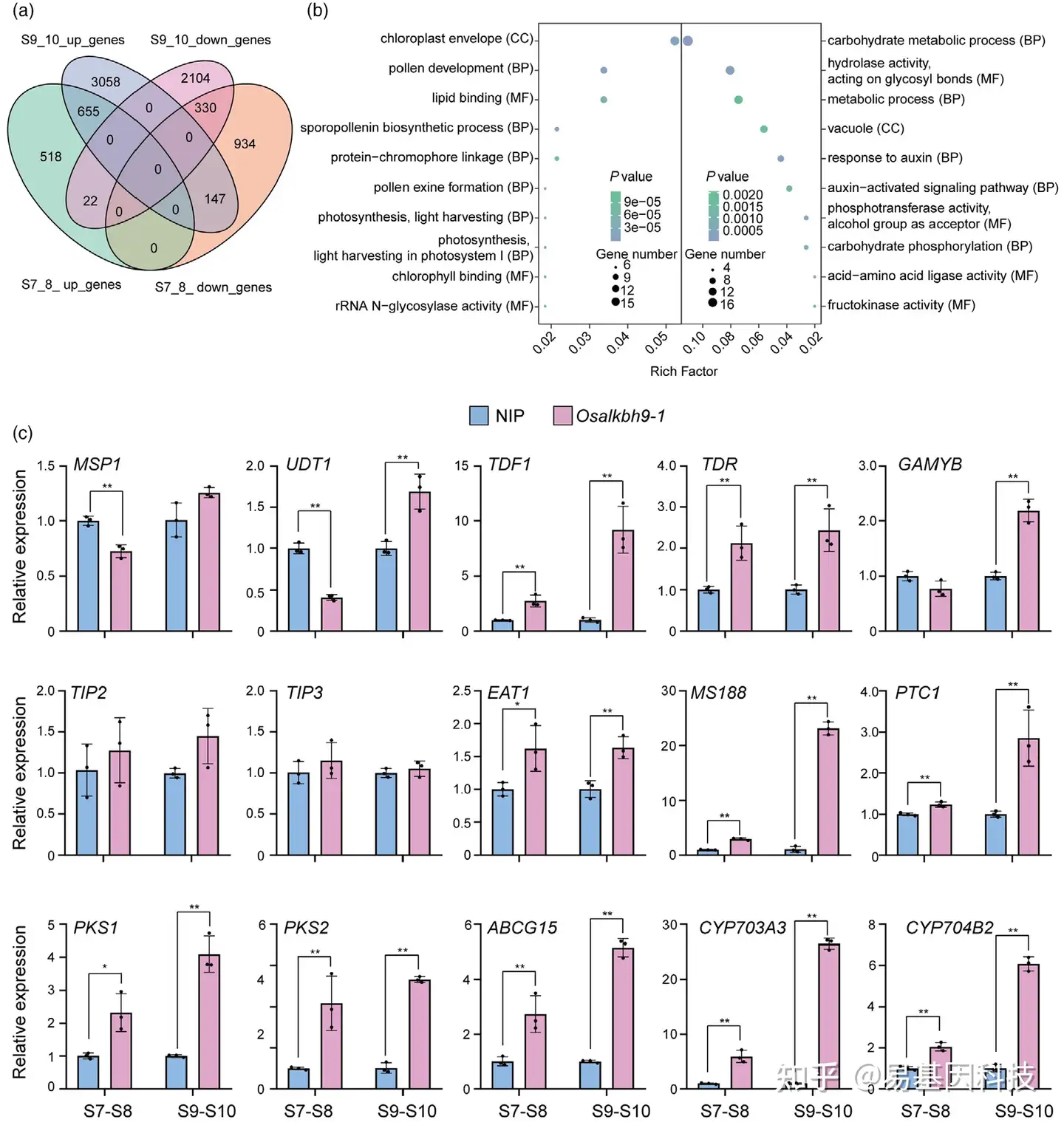
图4:OsALKBH9对雄性育性和花粉外壁积累相关基因表达。
(a) Osalkbh9-1突变体与野生型(WT)在花药发育7~8阶段和9~10阶段的基因表达差异。通过维恩图展示两个阶段共有和特有的差异表达基因。
(b) 对第7~8阶段和第9~10阶段的共上调和共下调基因进行GO分析。左部分表示共上调基因,右半部分表示共下调基因。CC代表细胞组分(cell component),BP代表生物过程(biological process),MF代表分子功能(molecular function)。
(c) qRT-PCR检测绒毡层发育和花粉外壁积累所需的基因。
(5)OsALKBH9基因功能丧失导致水稻转录组范围内m6A修饰高甲基化
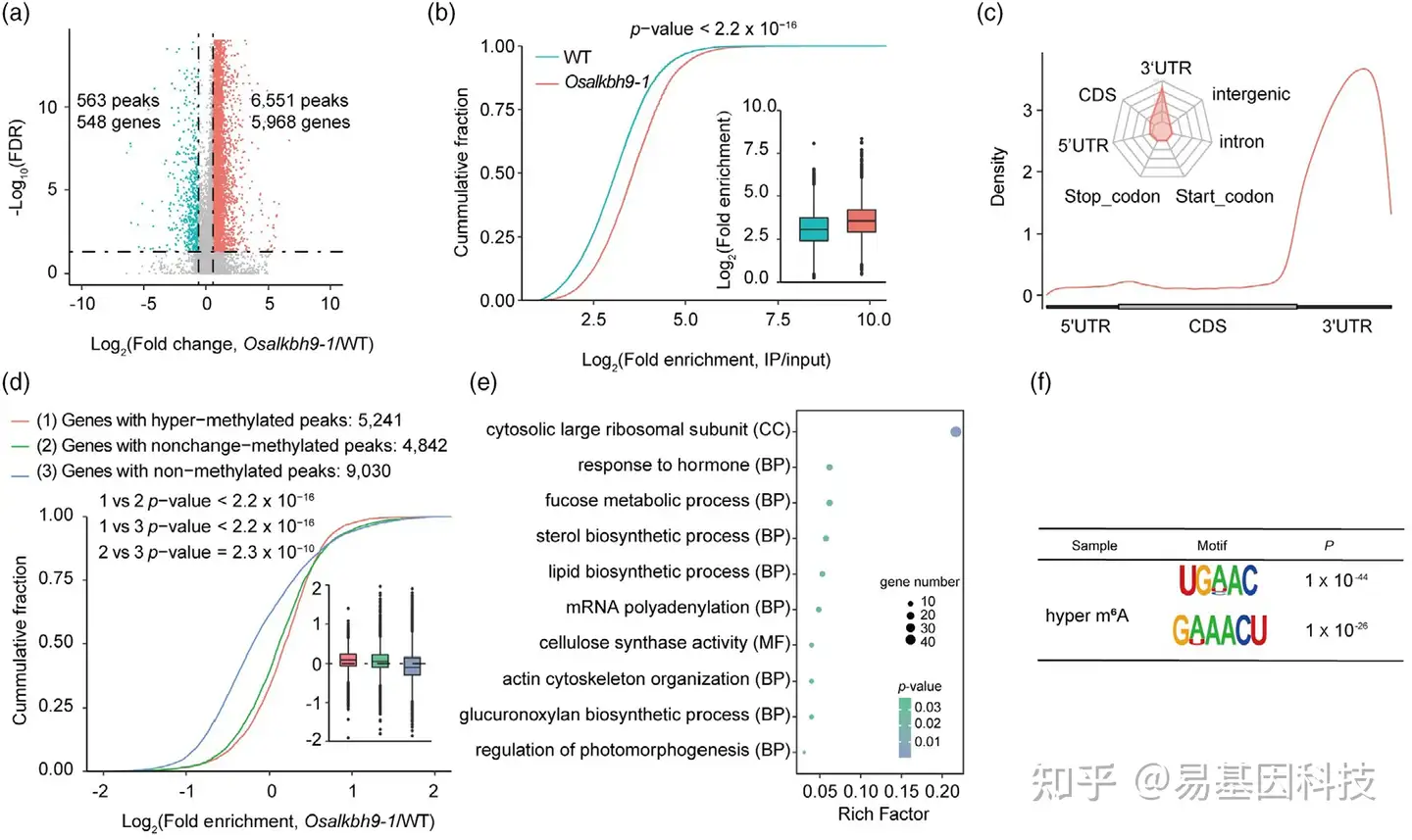
图5:OsALKBH9基因功能丧失导致转录组范围的m6A高甲基化。
- 火山图结果显示了Osalkbh9-1与野生型(WT)之间m6A修饰变化。
- 累积分数图和箱线图显示Osalkbh9-1和WT中m6A峰值的log2(富集倍数)。
- meta图和雷达图显示Osalkbh9-1中鉴定出的m6A高甲基化峰值在指定mRNA片段的分布。
- 累积分数图和箱线图显示Osalkbh9-1/WT中高甲基化、未变化甲基化和低甲基化峰值的基因表达模式。
- 对具有高甲基化峰值的基因进行GO分析。
- 基于前1000个峰值,使用HOMER软件鉴定了依赖OsALKBH9的motif。
(6)OsALKBH9功能丧失导致TDR和GAMYB转录本中的m6A高甲基化
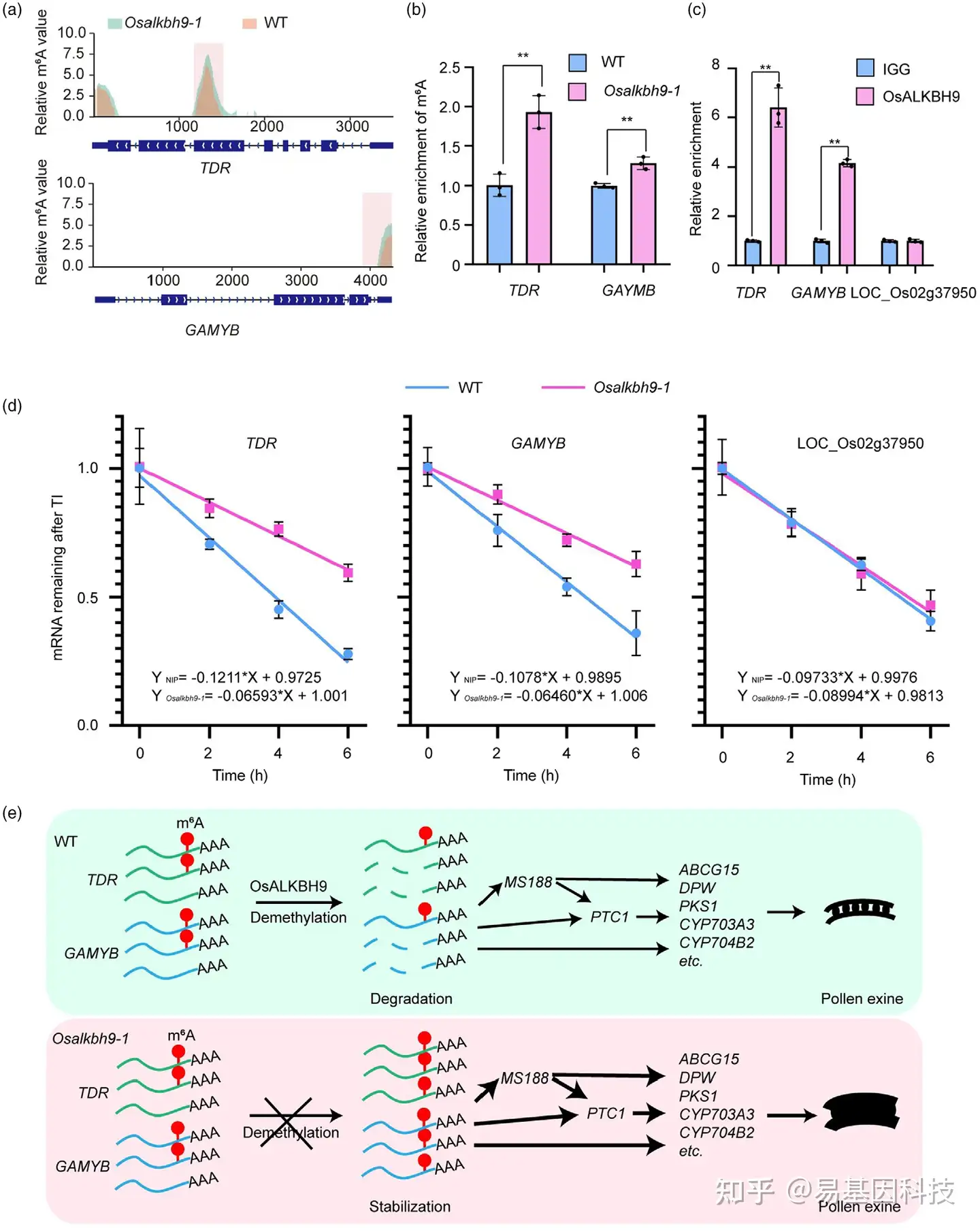
图6:OsALKBH9依赖的m6A去甲基化调控花粉外壁积累。
(a) TDR和GAMYB的相对m6A值。
(b) m6A-IP-qPCR结果显示WT和Osalkbh9-1的第9~10阶段花药中的相对m6A水平。
(c) Osalkbh9-1 OsALKBH9pro::OsALKBH9CDS FLAG小穗(第7~12阶段花药)中的RIP-qPCR显示Osalkbh9直接与TDR和GAMYB转录本结合。
(d) WT和Osalkbh9-1小穗(花药第7~12阶段)中TDR和GAMYB的mRNA半衰期。
(e) OsALKBH9如何调控花粉外壁积累的工作模型。在野生型中,OsALKBH9去除TDR和GAMYB转录本上的m6A,降低其mRNA稳定性,确保花粉外壁适当积累。在Osalkbh9-1突变体中,TDR和GAMYB转录本中m6A高甲基化,促进其mRNA稳定性,激活参与花粉外壁积累的基因表达,导致花粉外壁过度积累和异常的外壁形态。
关于易基因RNA m6A甲基化测序(MeRIP-seq)技术
易基因MeRIP-seq技术利用m6A特异性抗体富集发生m6A修饰的RNA片段(包括mRNA、lncRNA等rRNA去除所有RNA),结合高通量测序,可以对RNA上的m6A修饰进行定位与定量,总RNA起始量可降低至10μg,最低仅需1μg总RNA。广泛应用于组织发育、干细胞自我更新和分化、热休克或DNA损伤应答、癌症发生与发展、药物应答等研究领域;可应用于动物、植物、细胞及组织的m6A检测。
大样本量m6A-QTL性状关联分析,传统MeRIP单个样品价格高,通常难以承担。易基因开发建立MeRIP-seq2技术,显著提成IP平行性,实现不同样本间相对定量,降低检测成本。
易基因提供适用于不同科研需求的MeRIP技术:
- m6A甲基化-常量mRNA 甲基化测序(MeRIP-seq)
- m6A甲基化-常量mRNA +lncRNA甲基化测序(lnc-MeRIP-seq)
- m6A甲基化-微量mRNA +lncRNA甲基化测序(Micro-lnc-MeRIP-seq)
- 高通量m6A甲基化-常量mRNA甲基化测序(MeRIP-seq2)
技术优势:
- 起始量低:样本起始量可降低至10-20μg,最低仅需1μg总RNA;
- 转录组范围内:可以同时检测mRNA和lncRNA;
- 样本要求:可用于动物、植物、细胞及组织的m6A检测;
- 重复性高:IP富集重复性高,最大化降低抗体富集偏差;
- 应用范围广:广泛应用于组织发育、干细胞自我更新和分化、热休克或DNA损伤应答、癌症的发生与发展、药物应答等研究领域。
研究方向:
m6A甲基化目前主要运用在分子机制的理论性研究
- 疾病发生发展:肿瘤、代谢疾病(如肥胖/糖尿病)、神经和精神疾病(如阿尔兹海默症/抑郁症)、炎症…
- 发育和分化:早期胚胎发育、个体/组织/器官生长发育、干细胞分化与命运决定、衰老
- 环境暴露与响应:污染、抗逆、生活方式
关于m6A甲基化研究思路
(1)整体把握m6A甲基化图谱特征:m6A peak数量变化、m6A修饰基因数量变化、单个基因m6A peak数量分析、m6A peak在基因元件上的分布、m6A peak的motif分析、m6A peak修饰基因的功能分析
(2)筛选具体差异m6A peak和基因:差异m6A peak鉴定、非时序数据的分析策略、时序数据的分析策略、差异m6A修饰基因的功能分析、差异m6A修饰基因的PPI分析、候选基因的m6A修饰可视化展示
(3)m6A甲基化组学&转录组学关联分析:Meta genes整体关联、DMG-DEG对应关联、m6A修饰目标基因的筛选策略
(4)进一步验证或后期试验
易基因提供全面的表观基因组学(DNA甲基化、DNA羟甲基化)和表观转录组学(m6A、m5C、m1A、m7G)、染色质结构与功能组学技术方案(ChIP-seq、ATAC-seq),详询易基因:0755-28317900.

参考文献:
Tang J, Lei D, Yang J, Chen S, Wang X, Huang X, Zhang S, Cai Z, Zhu S, Wan J, Jia G. OsALKBH9-mediated mA demethylation regulates tapetal PCD and pollen exine accumulation in rice. Plant Biotechnol J. 2024 Apr 17. doi: 10.1111/pbi.14354. Epub ahead of print. PMID: 38634166.
相关阅读:
RNA m6A甲基转移酶METTL14在儿童肿瘤进展中的作用研究 | 科研速递
项目文章 | MeRIP-seq揭示m6A修饰在肺动脉高压(PAH)发病机制中的潜在作用和新治疗靶点


















 6619
6619

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









