





大脑区域之间的连接允许病理性扰动从单一源区域扩散到多个区域。多种疾病中的神经退行性变模式,包括前额颞叶痴呆的行为变异型(bvFTD),类似于大尺度功能系统,但bvFTD相关的萎缩模式如何与结构性网络组织相关仍然未知。本研究探讨了散发性和遗传性bvFTD中的神经退行性变模式是否受连接组结构的制约。我们在遗传性bvFTD(75名患者,247名对照)和散发性bvFTD(70名患者,123名对照)中估算了区域萎缩模式。首先,我们识别了bvFTD中的分布式萎缩模式,主要针对与边缘内在网络和岛叶细胞构筑类相关的区域。区域萎缩与结构和功能相连的邻近区域萎缩显著相关,表明网络结构塑造了萎缩模式。通过数据驱动和基于模拟的方法,前岛叶被识别为大脑萎缩的主要群体中心,额叶腹内侧和前内侧颞叶区域为次要中心。我们发现FTD相关基因,即C9orf72和TARDBP,赋予疾病局部转录组易感性,调节病理通过连接组的传播。综上所述,我们的结果表明,散发性和遗传性bvFTD的萎缩模式共同由全局连接组结构和局部转录组易感性塑造,为异质性病理实体如何导致相同的临床综合征提供了解释。本文发表在Brain杂志。
关键词: 连接组、前额颞叶痴呆、疾病中心、基因表达、网络传播
主题:基因表达、前额颞叶痴呆、萎缩、大脑、遗传学
引言
前额颞叶痴呆(FTD)是最常见的早发性痴呆之一。FTD的行为变异型(bvFTD)以各种行为(冷漠、冲动、强迫和刻板行为)、人格(同理心和同情心下降、个人偏好改变)和认知(执行功能障碍和社会认知缺陷)变化的组合为特征,是FTD最常见的临床变异型。尽管其临床表现独特,但bvFTD在病理上具有异质性,最常见的亚型与Tau或TDP-43的过度磷酸化聚集物的积累有关。这组导致FTD的病理性蛋白病被归类为前额颞叶变性(FTLD)。大多数病例是散发性的;然而,约20%是由常染色体显性遗传突变引起的,包括C9orf72、GRN和MAPT附近的六核苷酸重复扩增,这些是最常见的致病基因。
FTLD(前额颞叶变性)病理通过其在额叶和前颞叶脑区的主要定位引起临床bvFTD症状。在临床上,这反映为进行性皮质萎缩,这是诊断的关键生物标志物。虽然散发性和遗传性bvFTD的萎缩模式存在主要重叠,但每种遗传亚型都有独特的特征,包括MAPT中的前内侧萎缩、GRN中的后额叶和顶叶受累以及C9orf72中的丘脑/小脑体积损失。近年来,人们对理解异质性病理变化如何导致相似的临床和萎缩模式产生了兴趣。
早期基于功能性MRI的工作假设,神经退行性疾病中的萎缩主要沿功能神经网络进展,在bvFTD中主要影响突显网络。在突显网络内,前岛叶被确定为最可能的疾病中心,这一发现得到了在该区域特有的叉细胞和冯·埃科诺莫神经元中tau或TDP-43聚集物的病理积累的进一步支持。虽然前岛叶在疾病中明显扮演重要角色,但其他使用数据驱动方法研究结构性萎缩模式的研究揭示了不同的形态学亚型,包括两个以突显网络为主的亚组(额叶/颞叶亚型和额叶亚型)、一个以语义评估网络为主的组和一个以皮层下为主的组。这开启了可能性,即并非所有bvFTD病例的起源都有一个单一的独特中心。
新兴理论强调连接组结构塑造多种神经退行性疾病的进程和表现。内源性蛋白的错误折叠及其随后的跨神经元传播已在FTLD(前额颞叶变性)、阿尔茨海默病、帕金森病、亨廷顿病和肌萎缩侧索硬化症中得到证实。尽管每种疾病的起源和涉及的蛋白质不同,但病理的传播似乎反映了大尺度水平的大脑网络组织。具体来说,解剖连接被认为支持有毒蛋白聚集物的传播,使局部病理可以在连接的神经元群体之间传播并渗透到大脑的分布式网络中。
关于bvFTD中病理传播的两个关键问题仍未得到解答。首先,病理的传播很可能通过物理白质连接发生,但结构连接对萎缩进展的贡献在bvFTD中研究较少。FTLD病理跨神经元传播的证据主要基于功能成像的推断,一些使用动物模型、尸检数据和萎缩模式预测的研究提供了支持。虽然功能连接反映了潜在的结构连接模式,有时在没有这种数据的情况下被用作结构连接的代理,但这两种模态捕捉了大脑网络组织的根本不同特征,它们之间只有中等程度的相关性。其次,局部易感性的作用还不清楚。区域分子和细胞构成的差异可能使某些节点更易受或不易受影响。特别是,最近在其他神经退行性疾病中的报告表明,基因表达的区域差异可能赋予易感性,有效地引导病理过程通过网络。总的来说,我们假设大脑网络结构,结合特定基因表达赋予的局部易感性,塑造了包括FTD在内的大脑疾病中萎缩模式的空间分布。
在本研究中,我们在散发性和遗传性变异中测试了一个基于结构网络的bvFTD萎缩传播模型。具体而言,我们检验了bvFTD中的萎缩模式反映潜在网络组织和局部转录组易感性的假设。首先,我们估计了bvFTD患者皮质萎缩模式,即组织变形的区域变化。然后,我们使用从健康个体独立样本中得到的结构和功能连接网络,研究相互连接的区域是否显示相似的萎缩模式。最后,我们使用数据驱动方法和模拟大脑网络萎缩传播的基于模拟的方法识别了潜在的疾病中心。我们进一步探讨了FTD相关基因对萎缩传播的潜在贡献。
参与者和方法
参与者
我们从前额颞叶变性神经影像学计划(FTLDNI)数据库中获取了bvFTD患者和认知正常对照(CNCs)的数据,该数据库包含与每次临床访视相匹配的T1加权MRI扫描(http://4rtni-ftldni.ini.usc.edu/)。bvFTD患者的纳入标准是根据FTD联盟标准3诊断为可能或可能的bvFTD,最终纳入70名bvFTD患者(主要是散发性)和123名CNCs(正常对照)进行分析。一些患者有多次扫描;因此,bvFTD组共有156次扫描,CNC组有326次。我们还获取了遗传性前额颞叶痴呆计划2(GENFI2—http://genfi.org.uk/)第三次数据冻结(2017年12月)的数据,该计划包括英国、欧洲和加拿大的23个中心。GENFI2参与者包括已知的C9orf72、GRN或MAPT致病性突变的有症状携带者及其一级亲属,这些亲属有携带突变的风险,但不显示任何症状(即风险受试者)。经确认不携带突变的健康一级亲属被视为CNCs。由于本研究的目的是研究bvFTD临床表型中萎缩的网络传播,因此排除了无症状携带者和临床诊断非bvFTD的有症状携带者。这个GENFI2队列包括75名bvFTD患者和247名CNCs。表1描述了这两个队列的人口统计学和临床特征。使用双样本t检验来检验基线时的人口统计学和临床变量。分类变量使用χ2分析进行分析。结果以平均值±标准差(SD)和中位数(四分位数范围)表示,视情况而定。
表1 FTLDNI和GENFI2样本的人口统计学和临床特征
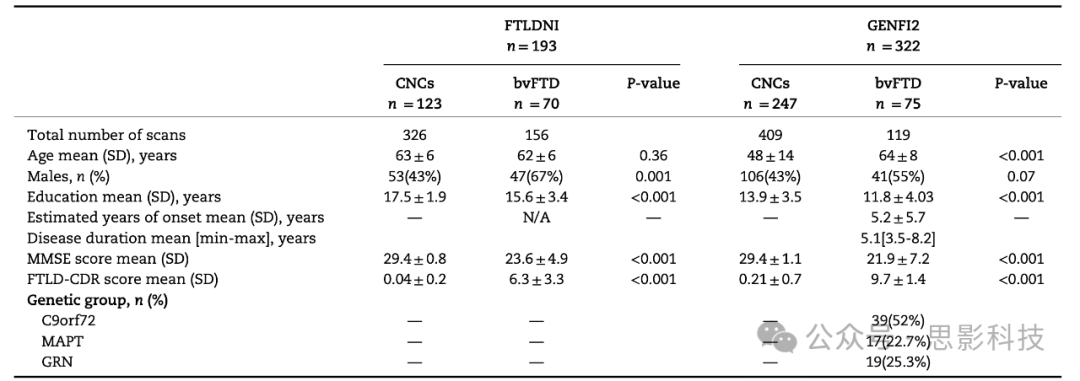
GENFI2队列中的CNCs对应于有记录的FTD相关遗传突变家庭成员的非携带者一级亲属。GENFI2队列中CNCs的遗传组别指这些非携带者受试者家族中存在的突变。数值以平均值±标准差、中位数[四分位数范围]表示。每个临床变量的可用数据以n指定,而N/A表示原始数据库中没有可用数据。
MRI获取和处理
对于FTLDNI队列,在三个站点获取了3.0 T MRI[T1加权磁化准备快速梯度回波成像(MPRAGE),重复时间(TR) = 2 ms,回波时间 = 3 ms,反转时间 = 900 ms,翻转角9°,矩阵256 × 240,切片厚度1 mm,体素大小1 mm3]。对于GENFI2样本,在多个中心使用GENFI成像协议在西门子Trio 3 T、西门子Skyra 3T、西门子1.5T、飞利浦3 T、通用电气(GE) 1.5T或GE 3T扫描仪上获得体积T1加权MPRAGE MRI。扫描协议在研究开始时设计,以确保扫描仪之间的充分匹配和图像质量控制。
所有T1加权扫描都通过我们的纵向管道进行预处理,包括图像去噪、强度非均匀性校正和使用直方图匹配将图像强度归一化到范围(0-100)。本研究中使用的图像处理工具旨在处理来自多站点研究的数据,以处理由多站点扫描引起的偏差,并已成功应用于多个多站点项目。每个时间点的每个原生T1加权体积首先线性配准到受试者特定模板,然后配准到ICBM152模板。然后,所有图像使用ANTs微分同胚配准管道非线性配准到ICBM152模板。两名有经验的评分者(M.D.和A.L.M.)对图像进行视觉评估,以排除具有显著成像伪影(如运动、不完整的视野)或不准确的线性/非线性配准的案例。这种视觉质量控制在不知道诊断的情况下完成。在1724次扫描中,只有43次(2.5%,GENFI2中36次,FTLDNI中7次)被拒绝。这导致共有515名受试者被纳入进行横断面形态测量分析。
基于变形的形态测量分析
使用蒙特利尔神经病学研究所(MNI) MINC工具进行基于变形的形态测量(DBM)分析。从将MNI-ICBM152-2009c模板映射到受试者MRI的非线性变换中获得的局部变形编码了MNI平均模板和受试者大脑之间的局部组织体积差异。在每个体素测量变形场雅可比行列式的行列式。行列式值>1.0表示受试者的局部体积大于平均模板(例如,在FTD情况下的脑室或沟扩大)。行列式值<1.0表示受试者的局部体积小于模板。后者通常被解释为组织萎缩,尽管仅使用横断面数据。DBM用于评估体素级横断面组相关的体积差异。为了获得反映bvFTD和CNCs之间差异模式的体素级图,对每个数据集分别在体素基础上应用以下混合效应模型:
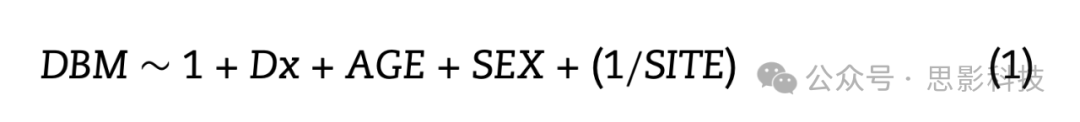
混合效应模型包括"年龄"作为连续固定变量,"诊断(Dx)"和"性别"作为固定分类变量。"站点"作为分类随机变量包括在内。感兴趣的变量是"诊断",反映了bvFTD和CNCs之间显著不同的大脑区域,同时控制年龄和性别。从模型中提取统计t图,并用于本文其余的分析。最后,t统计量乘以-1,使得较高的正值对应于较高的萎缩,负值对应于患者的体积扩张。
解剖学分区
通过DBM分析和混合效应模型获得的统计t图使用Cammoun图谱(解剖学Desikan-Killiany图谱的多分辨率扩展)被分为219和1000个大小近似相等的皮质区域或分区。我们将219和1000分区分辨率分别称为低分辨率和高分辨率分区。分区wise的t统计量(即萎缩)被估计为根据图谱分配给该分区的所有体素的平均t统计量。我们在两种分区分辨率下重复了所有分析,以确保结果在多个空间尺度上可重复。
结构和功能网络重建
健康个体的连接模式用于表示bvFTD患者观察到的分布式萎缩模式的大脑网络结构。从一个公开可用的数据集52获得了70名健康个体(平均年龄28.8±9.1岁)的结构和功能连接数据。有关数据获取参数和预处理分析的详细信息可在Griffa等人的文章中找到。简而言之,参与者在使用32通道头部线圈的3 T MRI扫描仪(Trio, Siemens Medical)中进行扫描。扫描协议包括:(i)对白质/灰质对比敏感的MPRAGE序列(1毫米平面分辨率,1.2毫米切片厚度);(ii)扩散谱成像(DSI)序列(128个扩散加权体积和一个b0体积,最大b值8000 s/mm2,2.2×2.2×3.0毫米体素大小);以及(iii)对血氧水平依赖(BOLD)对比敏感的梯度回波平面成像序列(3.3毫米平面分辨率和切片厚度,0.3毫米间隙,TR 1920毫秒,每位参与者产生280个图像)。DSI数据和确定性流线追踪法用于构建每个健康个体的结构连接网络。每对结构连接由纤维密度的对数变换加权。使用前述Cammoun图谱将个体结构连接网络分为低分辨率和高分辨率。在同一健康个体(睁眼状态)中收集的静息态功能MRI数据用于构建功能连接网络。预处理的静息态功能MRI时间序列也使用Cammoun图谱的低分辨率和高分辨率版本进行分区,并使用皮尔逊相关系数估计脑区对之间的功能连接。最后,构建了一个保留个体网络中边长分布的一致性群体平均结构连接,并将群体平均功能连接估计为个体间平均配对连接。
网络萎缩
使用群体平均结构和功能连接网络来估计每个大脑区域邻近区域的平均萎缩值。简而言之,对于结构定义和功能定义的邻近区域,给定大脑区域的邻近区域被定义为与之通过结构连接相连的区域。然后,每个大脑区域的结构连接邻近萎缩值被估计为该区域所有邻近区域的加权平均萎缩值:
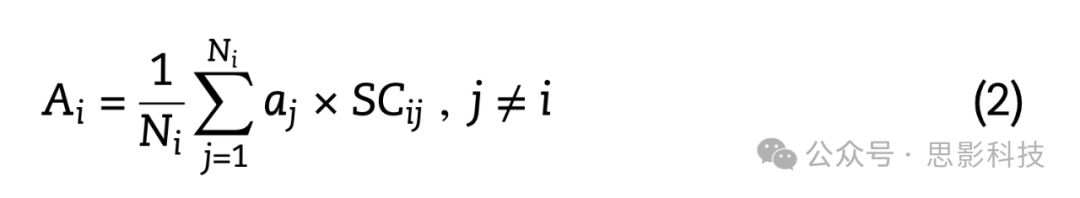
其中Ai是大脑区域或节点i的平均邻近萎缩值,aj是节点i的第j个邻近节点的萎缩值,SCij是节点i和j之间结构连接的强度,Ni是与节点i通过结构连接相连的邻近节点总数(即节点度)。通过Ni项进行归一化确保估计的邻近萎缩值独立于节点度。邻近萎缩估计排除了自连接(j ≠ i)。功能连接的邻近萎缩值使用与上述相同的方程估计,但区域萎缩值由节点i和j之间的功能连接强度(FCij)加权:
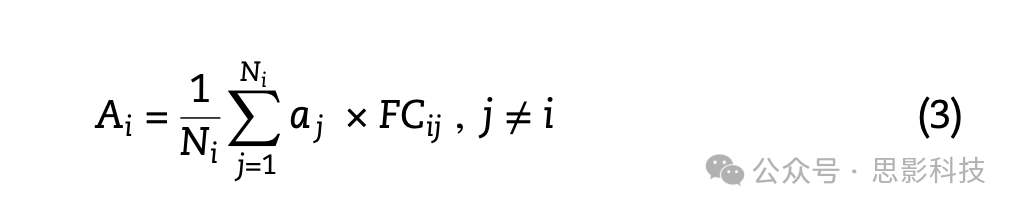
对于结构定义和功能定义的邻近萎缩估计,邻近节点被定义为与考虑中的节点结构连接的节点。总的来说,为每个区域估计了一个单一的邻近萎缩值。我们使用皮尔逊相关系数分别评估节点萎缩与结构定义和功能定义邻近节点的平均萎缩之间的关系(图2A)。
数据驱动的疾病中心分析
为了识别潜在的疾病中心,我们假设疾病中心是一个高度萎缩且连接到高度萎缩邻近区域的节点,相比于一个高萎缩但邻近区域健康的节点,或一个健康但邻近区域萎缩的节点。使用数据驱动的方法,我们首先根据估计的区域萎缩值对节点进行排序。我们还在一个单独的列表中根据邻近区域的平均萎缩值对节点进行排序。然后,我们计算每个节点在两个列表中的平均排名,并将在两个列表中都高度排名的节点(即局部和邻域平均排名都高的节点)识别为潜在的疾病中心(图2B)。
基于代理的传播模型
基于模拟的疾病中心分析
为了研究跨神经元传播假说,我们使用易感-感染-移除(SIR)基于代理的模型在低分辨率加权一致结构连接网络(111个区域)的左半球上模拟病理的传播。简而言之,该模型包括模拟皮质中正常蛋白的错误折叠及其通过脑区之间的结构连接进行跨神经元传播。病理的积累作为致病因子,然后导致受影响区域的萎缩。重要的是,该模型包含了合成和清除率,这些率可以在脑区之间异质性地变化。有关模型主要方程的更多细节可以在补充材料中找到。为了探索大脑区域作为这一传播过程中心的可能性,我们首先对所有区域使用基线清除和合成率。我们通过逐一使用每个单独的脑区作为过程的种子来模拟病理的传播和由此产生的萎缩。对于每个种子区域,在每个时间点,我们然后计算模拟和经验萎缩模式之间的皮尔逊相关性。
基因表达
为了研究基因表达在塑造模拟萎缩模式中的潜在作用,我们访问了Allen人脑图谱,该图谱提供了来自六个尸检大脑(一名女性,年龄24-57岁,42.5±13.38岁)的区域微阵列表达数据。我们为左半球的111个区域分区生成了存储基因表达得分的向量。这些向量随后被纳入SIR(易感-感染-移除)模型,以调节每个区域的合成和清除率,使得更高的表达得分意味着更高的合成和清除率。更具体地说,我们的分析集中在四个与先前已经与bvFTD相关的基因相关的基因表达向量上,即MAPT、GRN、C9orf72和TARDBP。鉴于受试者是基于其临床表型(bvFTD)而不是特定的病理亚型或基因突变选择的,我们探讨了所有四种基因表达对合成和清除的潜在作用。我们的目标是识别潜在的新机制过程,这些过程可能潜在地解释萎缩的传播,特别是在散发性bvFTD中,我们对与各种蛋白病相关的基因的贡献知之甚少。在补充实验中,我们还使用了完整"基因×脑区"基因表达矩阵的第一主成分。该成分捕捉了人类皮质转录变异的主要轴。
AHBA数据使用abagen工具箱进行预处理并映射到分区的脑区。在预处理过程中,我们首先将组织样本的MNI坐标更新为通过非线性对齐到ICBM152模板解剖结构生成的坐标。我们还使用Arnatkevic̆iūtė等人提供的数据重新注释了所有基因的微阵列探针信息。然后,我们通过仅保留信噪比大于0.5的探针来过滤探针。当多个探针索引同一基因的表达时,我们选择了在供体间区域变异模式最一致的探针。然后将样本分配到Cammoun图谱中的单个区域。如果在分区内直接找不到样本,则选择最近的样本,距离最多2毫米。如果在分区2毫米范围内没有找到样本,我们使用所有供体中最接近空分区质心的样本。为了减少错误分配的可能性,样本到区域的匹配受到半球和粗结构分区的限制(即皮质、皮质下/脑干和小脑,例如左皮质中的样本只能分配给左皮质中的图谱分区)。所有未分配到提供的图谱中脑区的组织样本都被丢弃。然后使用缩放的鲁棒S形函数对组织样本表达得分进行跨基因归一化,并重新缩放到单位区间。表达得分也使用相同的程序在组织样本间进行归一化。然后,我们通过计算每个供体个别分区的样本间平均表达来聚合属于同一区域的微阵列样本。最后,区域表达谱在供体间取平均。
空模型
为了评估节点-邻近关系和疾病中心分析的统计显著性,我们使用了一个保持空间自相关的空模型(即"旋转测试")。我们首先使用Connectome Mapper工具包在Freesurfer fsaverage表面上生成Cammoun图谱(低分辨率和高分辨率)的基于表面的表示。然后,我们通过选择生成的fsaverage表面的球面投影上最接近分区质心的顶点来定义每个分区的空间坐标。最后,我们使用得到的分区空间坐标通过随机旋转地图并用最近分区的值重新分配节点值来生成大脑地图的空模型(如萎缩地图、疾病中心排名)。旋转首先应用于一个半球,然后镜像旋转用于另一个半球。这个程序重复10 000次,以生成具有保持空间自相关的大脑地图的空分布。
为了确保我们转录组结果的特异性,我们依赖于第二个保持空间自相关的空模型。这个模型由Burt等人提出,可以使用brainSMASH python工具箱实现。首先,经验大脑地图被随机置换。然后,这个置换的大脑地图被空间平滑并重新缩放,以重新引入经验大脑地图的空间自相关(SA)。平滑过程通过以下转换实现:

其中y是替代地图,x是置换数据,z是随机高斯噪声向量。α和β参数通过原始数据和置换数据的变异图之间的最小二乘优化来估计。通过最大化原始数据和置换数据变异图之间的拟合,我们确保替代地图的SA(空间自相)与经验地图的SA相匹配。
为了确保基于代理的模型中经验萎缩地图和模拟萎缩地图之间观察到的相关性是由大脑区域之间结构连接的拓扑组织解释的,而不仅仅是由大脑区域的空间嵌入解释的,我们生成了保持结构连接组几何形状的替代网络。一致网络的边首先根据区域间欧几里德距离进行分组。在每个长度组内,然后随机选择边对并交换。这个程序重复500次,生成一组重新连接的结构网络,这些网络保持原始网络的度序列,并大致保持经验网络的边长分布(即布线成本)。
结果
人口统计学
表1比较了两个研究数据库中bvFTD患者和认知正常对照(CNCs)之间的人口统计学和临床变量。在GENFI2队列中,bvFTD受试者平均年龄大于CNCs,但在FTLDNI中没有这种差异。如预期,与健康对照相比,有症状的受试者的平均MMSE得分显著较低,而FTLD-CDR得分较高。
萎缩分布、静息态网络和细胞构筑学分类
我们使用线性混合效应模型获得了群体水平的、与bvFTD相关的萎缩图,控制了年龄、性别和采集站点的影响。体素水平和分区萎缩图在补充图1A和B中描绘。为了评估分布式萎缩模式是否在特定脑系统中更为明显,我们使用了两种脑系统定义(图1):(i) Yeo等人定义的内在功能网络;(ii) 基于经典von Economo图谱的人类皮层细胞构筑学分类。首先根据Yeo网络和von Economo分类对节点进行分层。然后,我们分别计算了FTLDNI(图1A)和GENFI(图1B)数据集中每个内在网络(图1,左)和细胞构筑学类别(图1,右)的平均萎缩值。为了评估网络萎缩值的统计显著性,我们将经验值与从一组保持空间自相关的空模型(即"旋转测试"63,64;有关空模型的更多细节,请参见"材料和方法"部分)计算出的均值分布进行了比较。具体而言,网络标签被随机旋转,同时保持空间自相关,并为每次旋转计算平均网络萎缩值(10 000次重复;双尾检验)。
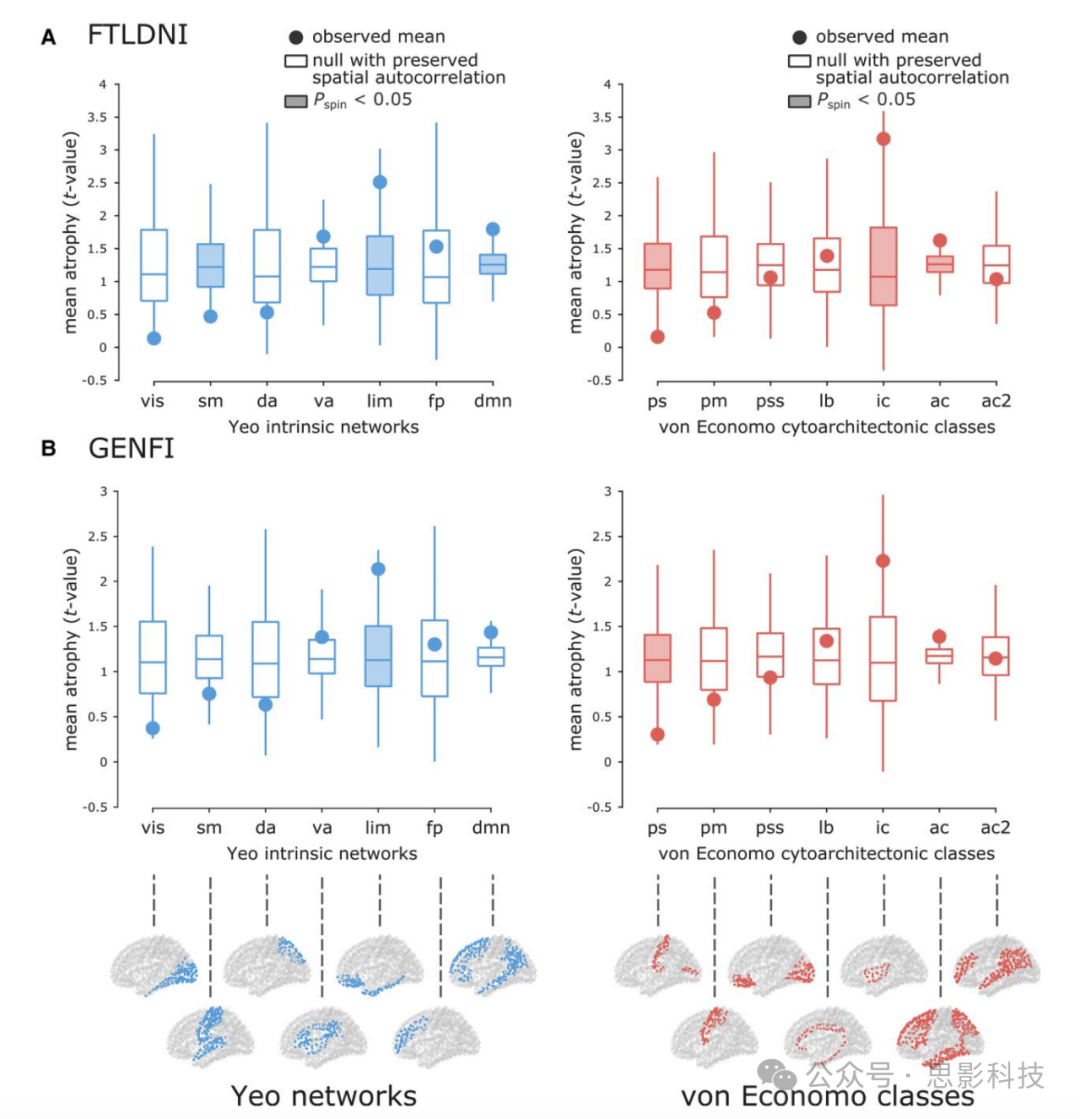
图1. 内在网络和细胞构筑学类别中的萎缩模式。
为Yeo内在功能网络(左)和von Economo细胞构筑学类别(右)计算了平均网络萎缩(即t值)。较高的t值对应于更大的萎缩。观察到的平均萎缩值用实心圆表示,分别对应每个内在网络和细胞构筑学类别。然后使用旋转测试的10 000次旋转随机排列网络标签,保持数据中的空间自相关。来自旋转测试的均值空分布使用箱线图描绘,分别针对(A) FTLDNI和(B) GENFI数据集的内在网络和细胞构筑学类别(10 000次重复;双尾检验)。底行显示了内在网络(左)和细胞构筑学类别(右)在皮层上的位置。
Yeo网络列表:视觉(vis)、躯体运动(sm)、背侧注意(da)、腹侧注意(va)、边缘(lim)、额顶(fp)、默认模式(dmn)。
von Economo类别列表:初级感觉皮层(ps)、初级运动皮层(pm)、初级/次级感觉皮层(pss)、边缘(lb)、岛叶皮层(ic)、联合皮层(ac, ac2)。
图1中展示了每个内在网络和细胞构筑学类别观察到的平均网络萎缩以及相应的空分布均值。图1底部展示了内在网络和细胞构筑学类别的解剖分布。注意内在网络和细胞构筑学类别中"边缘"系统定义的差异。内在边缘网络主要包括颞极和眶额皮层,而细胞构筑学边缘类别主要包括扣带回。就内在网络而言,边缘和默认模式内在网络受影响最大(即高于预期的萎缩),而躯体运动和视觉内在网络相对保留(即低于预期的萎缩)。就细胞构筑学类别而言,岛叶和联合细胞构筑学类别与空模型相比显示出更大的萎缩,而初级感觉细胞构筑学类别萎缩较低。虽然研究结果的统计显著性有轻微变化,但两个数据集中网络萎缩模式的总体趋势是一致的。
萎缩图与连接性的关系
接下来,我们研究了bvFTD中的萎缩模式是否受网络组织的制约,使得相连的区域显示相似的萎缩模式。具体而言,我们通过研究节点和邻近节点萎缩值之间的关系来评估节点的连接性特征是否可以预测其邻近节点的萎缩(图2A)。结构和功能连接(SC和FC)网络(补充图1C)来自70名健康参与者的独立样本52,用于估计每个区域的平均邻近萎缩值。然后通过将平均邻近萎缩与节点萎缩相关来检验节点和邻近萎缩之间的关系(图2C和D)。在两个数据集中,区域萎缩与其连接邻近节点的平均萎缩显著相关。图2C(左面板)显示了FTLDNI数据集的结果(高分辨率分区:r = 0.69, Pspin = 0.0001和r = 0.65, Pspin = 0.0001,分别对应SC和FC定义的邻近节点),图2D(左面板)显示了GENFI数据集的结果(高分辨率分区:r = 0.61, Pspin = 0.001和r = 0.54, Pspin = 0.0006,分别对应SC和FC定义的邻近节点)。这些相关性在考虑连接与非连接邻近节点时,在数据集和分辨率之间显著更大(补充表1)。
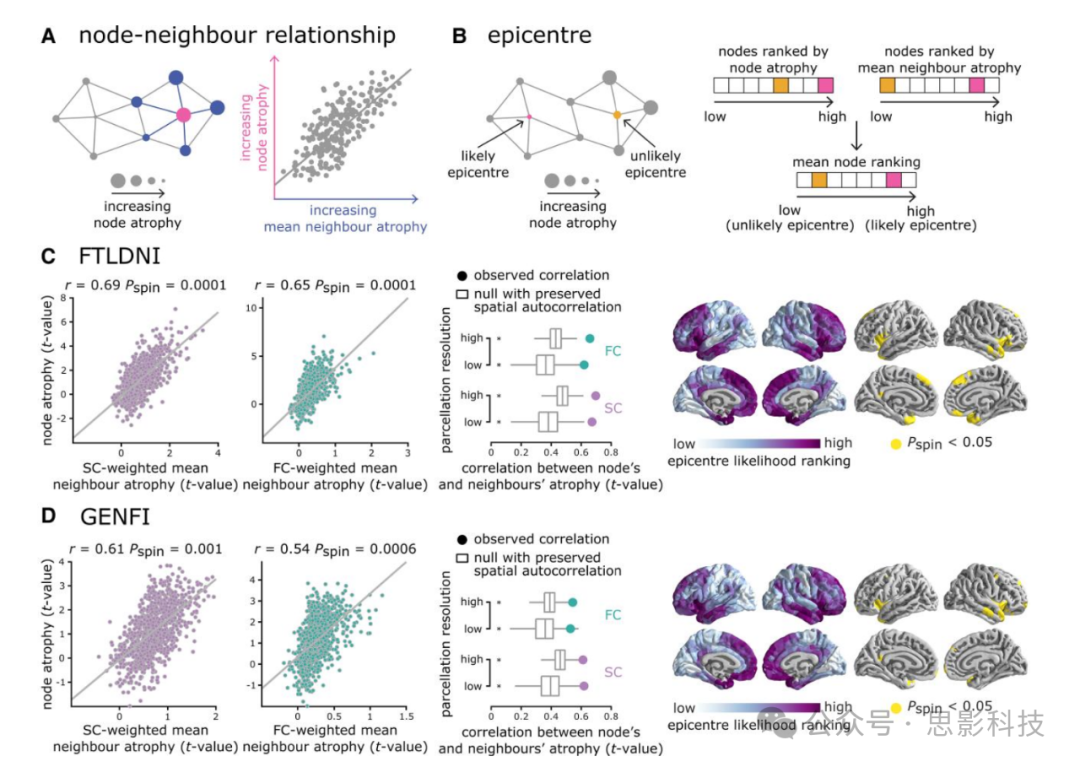
图2. 网络依赖的萎缩
(A)通过t值估计的节点萎缩与其连接邻近节点的平均萎缩相关,以检验bvFTD中分布式萎缩模式是否反映了潜在的网络组织。
(B)如果节点的萎缩与其连接邻近节点的萎缩相关(A),那么一个高度萎缩且其邻近节点也高度萎缩的节点更可能成为潜在的疾病中心,相比于一个高度萎缩但邻近节点健康的节点。为了量化皮层中的中心可能性,首先根据节点的萎缩值和其邻近节点的萎缩值对节点进行排序。然后将每个节点的中心可能性排名定义为其在两个列表中的平均排名。
(C和D)左:节点萎缩值与其结构和功能加权邻近节点(SC和FC)的平均萎缩值相关,分别针对FTLDNI(C)和GENFI(D)数据集。散点图显示了高分辨率分区的相关性。中:观察到的相关值(用实心圆表示)与从10000次旋转测试获得的一组相关值(用箱线图表示)进行了比较。星号表示统计显著性(Pspin < 0.05,双尾)。节点和邻近萎缩之间的关联在分辨率之间保持一致,并且在两个数据集中,经验网络与空网络相比显著更大。右:中心可能性排名在皮层上描绘。最可能的中心具有高显著排名,主要位于双侧前岛叶皮层和颞叶(10000次旋转测试)。
为了评估节点和邻近萎缩之间的关系是否特别由网络拓扑驱动而非空间自相关,我们使用了一个保持空间自相关的空模型来构建节点-邻近相关的空分布。图2C和D(中间面板)显示了观察到的节点和邻近萎缩之间的相关性,以及两个数据集相应的相关性空分布。我们还在较低的分区分辨率下重复了所有分析,以确保发现对网络节点的定义方式具有稳健性。节点和邻近萎缩之间的关系在分辨率之间保持一致,并且在两个数据集中,经验网络与空网络相比显著更大(图2C和D; Pspin < 0.05,双尾检验)。当使用二值化结构连接网络定义SC定义的邻近节点时,结果保持一致(补充图2)。
数据驱动的中心分析
鉴于萎缩模式的分布反映了结构和功能网络组织,我们接下来研究是否存在可能作为bvFTD潜在中心的脑区。我们将中心定义为一个高度萎缩且连接到高度萎缩邻近节点的节点(图2B)。根据节点的萎缩和其邻近节点的平均萎缩值对节点进行排序。然后将中心可能性排名估计为两个列表中节点排名的平均值。图2C和D(右)显示了FTLDNI(图2C)和GENFI(图2D)数据集在皮层上的中心可能性排名,其中高排名区域与岛叶皮层、腹内侧皮层和前内侧颞叶区域相关。然后将经验中心可能性排名与从保持空间自相关的空模型(10,000次旋转测试)估计的排名进行比较。几个区域被确定为潜在中心,包括双侧前岛叶皮层,以及前颞极区域,此外还有腹内侧和背内侧区域。当使用二值化结构连接网络定义SC定义的邻近节点时,结果保持一致(补充图2)。
动态传播模型
接下来,我们使用SIR(易感-感染-移除)模型探讨大脑的结构连接如何塑造FTLD变化的渐进传播。这个模型之前被用于研究帕金森病相关的萎缩,它通过模拟皮层中正常蛋白的错误折叠及其通过脑区之间的结构连接进行跨神经元传播来工作。病理的积累作为致病因子,导致受影响区域的萎缩(图3A)。中心被定义为引入错误折叠蛋白的那些区域。我们通过运行模型并在每个区域启动传播来测试哪个是观察到的经验萎缩模式最可能的中心。随着错误折叠的因子通过网络传播,我们测量了模拟和经验(FTLDNI)萎缩模式之间的皮尔逊相关性(图3B,左)。然后我们将一个区域的rmax(最大相关分数)定义为当它被用作传播过程的中心时,在所有t值中观察到的最大相关值。具有大rmax分数的区域是最可能的中心。具有最大rmax的三个节点位于岛叶、上额叶和外侧眶额皮层。对于这三个潜在中心,当考虑直接连接的邻近节点时,rmax大于考虑非连接节点时的值(补充图3)。
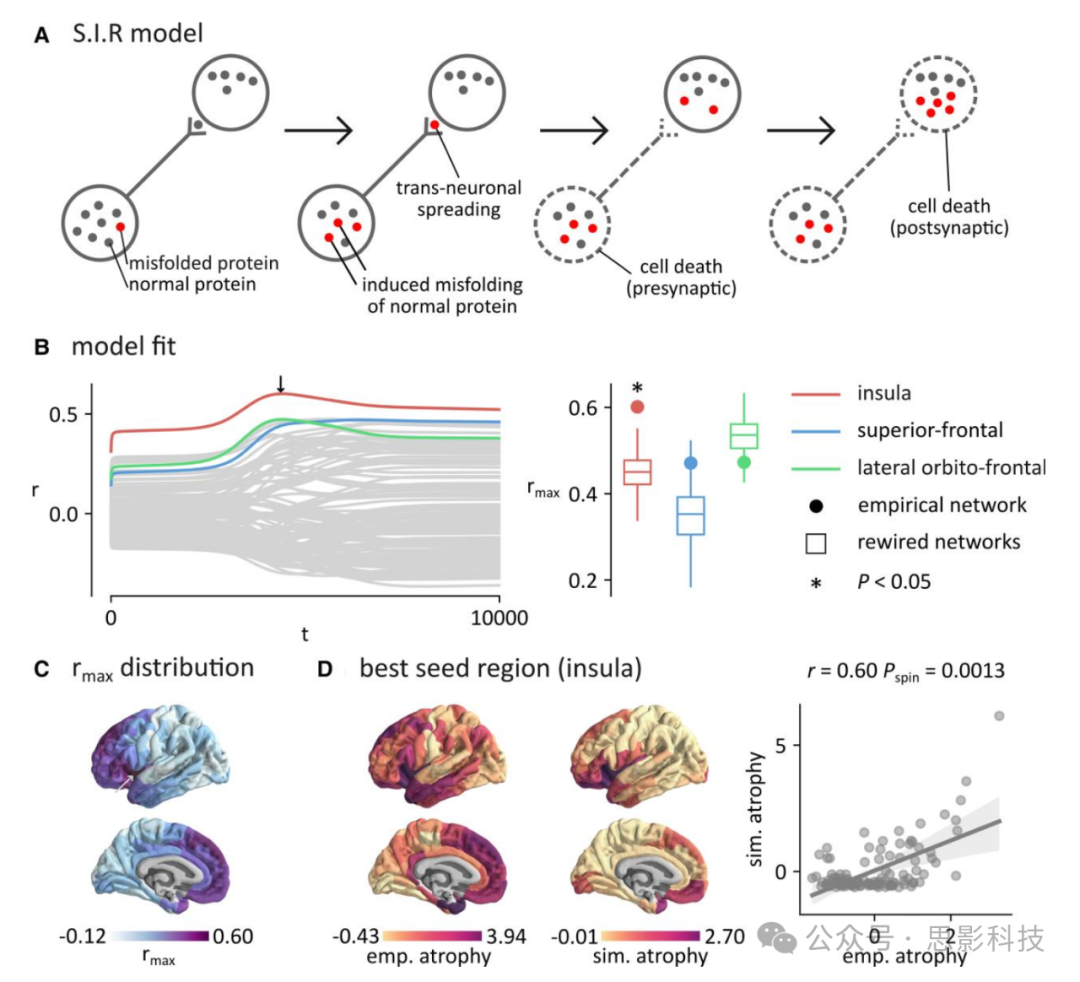
图3. 基于代理的建模
(A)SIR(易感-感染-移除)模型模拟大脑中病理的传播。蛋白质通过脑区之间的结构连接传播,并在突触前和突触后诱导萎缩。
(B)左:在每个脑区启动传播过程,并计算模拟和经验萎缩模式之间的相关性。通过岛叶种子点(rmax (最大相关分数)= 0.601)、上额叶皮层(rmax = 0.473)和外侧眶额皮层(rmax = 0.471)区域获得了三个最大的相关性。右:为了控制脑区空间嵌入的潜在影响,将rmax值与使用保留经验结构网络布线成本的重新连接网络获得的rmax相关性进行比较。星号表示统计显著性(P < 0.05,双尾)。通过经验网络的岛叶种子点计算的rmax(rmax = 0.60)显著大于通过重新连接网络的岛叶种子点计算的rmax(P < 0.002)。
(C)通过每个脑区种子点获得的最大拟合(rmax)显示在大脑表面上。通常以岛叶和前额叶区域为种子点获得较大的rmax(最大相关分数)值。
(D)左:经验萎缩模式(FTLDNI)。中:产生最大拟合的模拟萎缩模式。这种萎缩模式是以岛叶为种子,在t = 4410时获得的(见B中的箭头)。右:标准化经验和模拟萎缩模式之间关系的散点图(r = 0.60, P = 0.0013)。
影响大脑区域被识别为萎缩模式中心的一个重要因素是其在大脑中的空间位置。为了分离结构连接的作用,我们将这些rmax分数与在保留了经验结构网络的密度、度序列和布线成本的重新连接网络中模拟病理传播得到的分数进行了比较(图3B,右)。我们发现,在经验网络的岛叶区域启动传播获得的拟合显著大于在重新连接网络中获得的拟合(r = 0.601, P < 0.002)。我们还发现,它大于用欧几里德距离或几何距离矩阵替换我们模型中的结构连接矩阵所获得的拟合(补充图4A)。换句话说,通过岛叶种子点观察到的拟合显著大于仅从其度和空间位置所预期的,这可以归因于其在网络全局拓扑中的嵌入。这一结果表明,结构连接组的拓扑在塑造与经验萎缩高度对应的模拟萎缩模式中起着重要作用。
更一般地,通过查看rmax分数的地形分布,我们发现显示最大拟合的脑区位于岛叶、内侧前额叶和前颞叶皮层(图3C)。这些结果与我们发现这些区域具有较高的中心可能性排名一致。图3D显示了FTLDNI数据集的经验萎缩模式。这个模式与产生最大拟合的模拟萎缩模式进行了比较。这个最大拟合是通过岛叶种子点获得的,并在t = 4410时测量。我们发现两种分布之间存在显著关系(r = 0.60, P = 0.0013)。结果是针对FTLDNI数据集呈现的,但在GENFI数据集中也发现了类似的结果(补充图5)。到目前为止,我们专注于群体效应,因为基于变形的形态测量主要是用于检测大脑结构的群体水平差异。为了更好地理解患者的异质性,我们考虑了GENFI中的遗传亚型。我们将GENFI队列分层为C9orf72、GRN和MAPT突变携带者,并分别对每个遗传组重复了主要分析。补充图6A-C分别显示了每个遗传组的三个主要发现。我们在每个组中都发现了突显网络传播效应。此外,数据驱动和基于模拟的分析都在C9orf72和MAPT携带者中识别出额叶、颞叶和岛叶中心,MAPT中前内侧颞极的参与更为明显,而GRN携带者中额叶和外侧顶叶区域的参与更为明显,这与先前的研究一致。
基因表达对网络传播的贡献
鉴于遗传变异对bvFTD的贡献,我们接下来评估将基因表达信息纳入SIR(易感-感染-移除)模型是否可以增强拟合。我们使用来自AHBA的区域微阵列表达数据为四个先前与bvFTD相关的基因生成基因表达向量:MAPT、GRN、C9orf72和TARDBP。图4A显示了它们的地形分布。我们使用这些遗传信息来设置模型中蛋白质清除和合成的区域异质性。我们使用岛叶作为传播过程的种子区域,因为它是与经验数据拟合最好的区域。
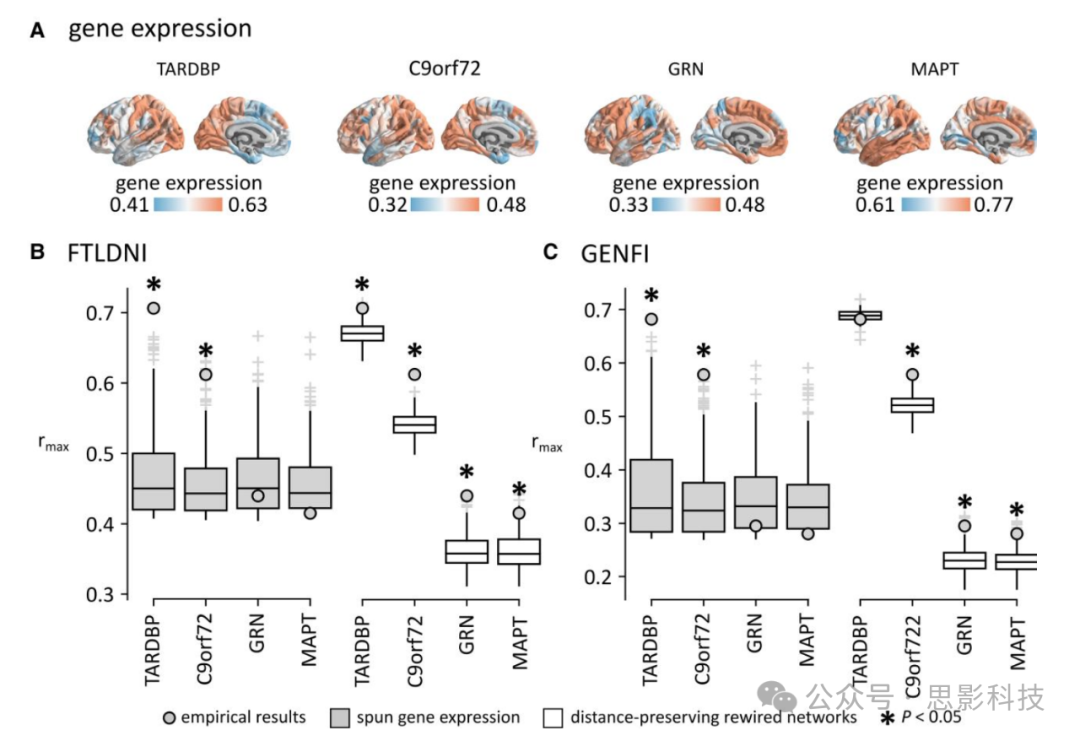
图4. 基因表达的贡献。
(A)为四个与bvFTD相关的基因生成了区域基因表达向量:TARDBP、C9orf72、GRN和MAPT。这些基因表达向量被纳入SIR(易感-感染-移除)模型。然后,以岛叶作为模拟传播过程的种子,计算了经验萎缩和模拟萎缩之间的相关性,分别针对FTLDNI数据集(B)和GENFI数据集(C)。每个基因获得的最大相关分数(rmax)与使用旋转分布的基因表达向量获得的最大相关分数(rmax)进行了比较(左侧箱线图)。星号表示统计显著性(P < 0.05,双尾)。对于两个数据集,我们发现通过纳入C9orf72和TARDBP表达信息获得的rmax分数显著大于使用置换基因表达向量获得的分数。最大相关性还与在保持距离的重新连接网络中获得的最大相关分数进行了比较(右侧箱线图)。
对于FTLDNI(图4B)和GENFI(图4C)两个数据集,我们测量了通过纳入每个基因的区域表达获得的rmax分数。对于MAPT、GRN、C9orf72和TARDBP,在FTLDNI数据集中我们获得了rmax = 0.42、rmax = 0.44、rmax = 0.61和rmax = 0.71的相关分数,在GENFI数据集中获得了rmax = 0.28、rmax = 0.30、rmax = 0.58和rmax = 0.68的相关分数。我们发现,使用C9orf72和TARDBP的表达添加合成和清除的区域异质性增加了模型拟合,而纳入GRN和MAPT的区域表达信息降低了模型拟合。为了研究发现的显著性,我们将基因表达向量旋转10000次,生成空间自相关的rmax分数空分布,并将其与经验结果进行比较。我们发现,使用C9orf72和TARDBP获得的分数显著大于使用置换基因表达向量获得的分数(FTLDNI数据集分别为P = 0.014和P < 0.0001,GENFI数据集两个基因均为P < 0.0001)。使用C9orf72和TARDBP获得的分数也显著大于使用基于变异图方法生成的空间自相关基因表达分布获得的分数(补充图7)。这些结果表明,C9orf72和TARDBP可能在驱动经验萎缩的空间模式中起重要作用。
为了研究基因表达与大脑结构连接之间的关系,我们将拟合与使用保留经验网络布线成本的重新连接网络获得的拟合进行了比较。对于C9orf72,我们发现使用经验网络获得的拟合显著大于使用重新连接空网络获得的拟合,无论是FTLDNI(P < 0.002)还是GENFI(P < 0.002)。使用经验连接组获得的拟合也大于用欧几里德距离或几何距离矩阵替换SIR模型中的结构连接矩阵获得的拟合(补充图4B)。对于TARDBP,我们发现使用经验网络获得的拟合在FTLDNI(P = 0.014)中显著大于使用重新连接空模型获得的拟合,但在GENFI(P = 0.508)中并非如此。类似地,在FTLDNI数据集中,使用经验连接组获得的拟合大于用欧几里德或几何距离矩阵替换它获得的拟合,但在GENFI数据集中并非如此(补充图4B)。总的来说,这些结果表明,结构连接组的拓扑对将TARDBP或C9orf72纳入SIR模型时观察到的模型拟合增加有积极影响,这种影响比节点之间的空间距离更大。对于TARDBP,这种影响在尝试将我们的模型拟合到与散发性和遗传性bvFTD相关的萎缩模式时都观察到,而对于C9orf72,这种影响仅在尝试将我们的模型拟合到与散发性bvFTD相关的萎缩模式时观察到。换句话说,我们的调查表明,C9orf72和TARDBP表达可以影响在结构连接组上展开的致病性传播过程。有趣的是,TARDBP(r = 0.83)和C9orf72(r = 0.61)都与人类皮层转录变异的主轴(基因PC159)强烈相关,后者在用于纳入区域异质性时也增强了SIR模型的拟合(补充图8)。然而,与TARDBP和C9orf72不同,这种增加的拟合并不显著大于使用重新连接网络获得的拟合。因此,这表明使用基因PC1获得的拟合主要是由于其在皮层表面的空间分布,而不一定是由于其对在结构连接组上展开的致病性传播过程的影响。为了完整起见,我们还在GENFI的三个遗传亚组中重复了这些分析。补充图6D显示结果在各亚组中是一致的,TARDBP和C9orf72是给出最大模型拟合的两个基因分布。
讨论
本研究为支持bvFTD(包括散发性和遗传性形式)中基于网络的萎缩理论提供了全面且统计上稳健的模型。我们的发现在两个数据集和遗传/散发性异质性中是一致的。具体来说,对于散发性和遗传性变异,节点变形与由结构和功能连接定义的邻近变形平均值之间存在强相关,支持萎缩通过基于网络的连接进行进展的理论。在小型(219)和大型(1000)皮质分区分辨率下观察到类似的发现。数据驱动的中心映射识别出双侧前岛叶皮层以及腹内侧皮层和前颞叶区域作为潜在的中心。前内侧区域作为中心的参与与先前的研究结果一致,即数据驱动的萎缩亚型包括一个"语义评估网络"为主的群体。遗传性bvFTD队列显示了非常相似的最可能中心的概况,还增加了一些背侧额叶区域。基于代理的传播模型进一步支持了这些区域作为中心的作用。
皮质萎缩的定位在边缘静息态网络中最为显著,在视觉空间网络中较少出现(考虑到其后部定位,这是可以预期的)。在遗传性FTD中,默认模式网络(DMN)存在显著萎缩,在散发性FTD中也有正向趋势。值得注意的是,先前被认为主要涉及bvFTD的突显网络并未显示统计学上显著的萎缩。然而,当查看von Economo细胞构筑学类别时,岛叶皮层受影响最大,而初级感觉神经元相对保留。这表明岛叶皮层在疾病中起着中心作用,但不一定通过整个腹侧注意网络(包括其最后部区域)传播。此外,虽然有一些报告显示bvFTD和阿尔茨海默病中突显网络与DMN的连接模式变化相反,但我们的结果表明bvFTD中DMN区域有显著参与。
最后,尽管具有探索性,但使用基于模拟的方法和来自AHBA的基因表达数据,我们发现C9orf72和TARDPB基因表达可能在散发性bvFTD的萎缩传播中发挥作用。事实上,考虑这两个与TDP-43相关的基因对清除和合成的影响,改善了传播模型与基于DBM的实际萎缩图之间的拟合。虽然我们不能排除FTLDNI中的一些受试者有未识别的C9orf72突变,但TARDPB的参与很有意思,因为这个基因的突变只构成遗传性FTD的很小一部分。结果表明,这个基因的活动可能在散发性bvFTD中发挥作用,这对未来的治疗方向可能很有意义。有趣的是,我们发现GENFI中三个遗传组(C9orf72、GRN、MAPT)中的萎缩都显示出显著的网络传播效应,整体上对局部基因表达有类似的依赖,但网络中心不同。与先前的研究一致,C9orf72和MAPT以额叶、颞叶和岛叶皮质中的突出中心为特征,类似于散发性病例,而MAPT中前内侧颞叶的参与更为明显。GRN相关的萎缩以更背侧额叶区域和外侧顶叶皮质中更大的中心可能性为特征。群体水平的遗传变异如何塑造导致萎缩的分子和网络级联仍然是未来研究的一个令人兴奋的问题。
重要的是要注意,这些发现主要是相关性的,并不能证明网络结构对萎缩的因果影响。具体来说,无法确定连接性是否驱动灰质萎缩的进展,或者连接组架构本身是否因bvFTD中报告的白质病变(通过白质高信号估计)而在患者中受损。我们使用从年轻健康参与者样本重建的结构和功能网络作为支持病原体传播的潜在架构。然而,网络架构的广泛变化可能会重新路由或限制病理的传播。白质变化也可能破坏正常的跨神经元营养因子运输,导致相连区域的萎缩,而不涉及任何病原体或错误折叠的蛋白质。这些额外因素可以在更复杂的模型中进一步研究,使用FTD患者纵向样本中区域萎缩和白质架构以及结构和功能网络变化的同时测量。具体而言,一个高度采样的、多模态纵向数据集,同时测量bvFTD中的区域病理、白质病变、基因表达、代谢、脑脊液生物标志物、血管和神经影像因素,将允许对疾病进行精确的多因素建模,改善bvFTD患者的个体化诊断、治疗干预和预后。
这些结果如何应用于个体患者?由于基于变形的形态测量是一种旨在检测大脑结构群体水平差异的技术,我们专注于群体效应。我们的发现帮助为什么具有如此不同的遗传和病理变异的患者在实践中可以表现出相似的临床综合征(即因为疾病传播受网络结构的约束)提供了神经生物学解释。考虑到疾病的多因素性质和个体间的显著差异,有必要根据个体患者的需求定制治疗干预。这些网络模型的核心临床前景是它们可以有效地总结可用的复杂多模态测量,产生少量临床相关特征。这些特征可能随后允许识别处于风险中的无症状个体、临床试验入组候选者以及新型疾病修饰疗法的靶点或结果指标。
局限性:
总的来说,我们的结果建立在先前文献的基础上,即神经退行性变化的模式反映了网络结构。与其他神经退行性疾病的报告一致,我们证明bvFTD的萎缩模式与全局连接组结构和局部转录组易感性相关。本研究的发现在遗传性和散发性bvFTD的两个独立样本中得到重复,并通过一系列方法选择进行了验证。我们还确认这些发现独立于潜在的混杂因素,如空间距离和分区分辨率。然而,在解释这些发现时需要考虑几个方法学问题。
首先,目前还没有可用的分子技术来直接测量体内FTLD的变化。为克服这一限制,我们选择使用DBM来估计bvFTD患者的萎缩,因为它是捕捉局部大脑组织体积变化的稳健方法。鉴于目前体内TDP-43和tau PET示踪剂在FTLD中不可靠,使用死后病理学评估(如磷酸化TDP-4381的免疫组织化学)将提供更直接的FTD相关病理测量。
其次,我们使用横断面数据和无向网络识别潜在的疾病中心,这阻碍了病理时间序列的重建。特别是,中心模型无法评估可能在灰质萎缩之前开始并在临床综合征和疾病诊断出现前多年开始的病理级联,包括分子、代谢、血管和功能变化。模拟疾病进展和萎缩在大脑网络中随时间的传播仍然是一个令人兴奋的开放问题,最终可能通过大型FTD数据集(包括GENFI和ALLFTD86,87)中增加纵向采样来解决,这些数据集包括从无症状遗传携带者到疾病晚期有症状患者的多模态数据。
第三,使用DSI和流线追踪法来估计结构连接网络。尽管最近的技术和分析发展提供了强大的方法来体内重建白质纤维,具有生物学上可解释的权重和与组织学良好的对应关系,但它们仍可能产生假阳性和假阴性结果。
第四,本研究包括的两个多站点数据集具有不同的人口统计学特征,可能潜在地影响结果。尽管形态测量程序控制了站点、年龄和性别,并且结果在两个数据集中是一致的,但我们的发现应该在考虑这些潜在混杂变量的情况下进行解释。
结论
总的来说,本研究使用了结构和功能连接网络以及考虑空间自相关和网络嵌入的严格统计分析,证明了bvFTD相关的神经退行性变化受连接组结构的制约,解释了30-40%的萎缩变异以及局部转录组易感性。FTD相关的萎缩似乎针对与前岛叶皮层相关的特定区域,但可能存在多个潜在的中心导致bvFTD临床表型。遗传性和散发性bvFTD形式之间的相似性表明,多种病理变化在萎缩的传播中受到网络结构的约束,这解释了为什么许多不同的病理和遗传实体导致相同的临床综合征。尽管具有探索性,我们的结果表明TARDPB基因表达可能对疾病进展有显著贡献,特别是在散发性bvFTD中。


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









