





社交焦虑障碍(SAD)的特征是害怕负面评价、消极的自我认知以及极度回避社交场合。这些反复出现的症状被认为会维持社交和认知思维的严重程度和实质性损害。SAD与情绪调节、知觉刺激功能和情绪处理相关的神经网络中断有关,这表明存在一个网络系统来描绘SAD的大脑皮层电活动内表型。本文旨在对静息、预期、刺激处理和恢复状态下社交焦虑中最常研究的脑电图(EEG)谱耦合、事件相关电位(ERP)、视觉诱发电位(VEP)以及其他连接性估计器进行全面综述。在Web of Science上的搜索提供了97项记录SAD个体大脑皮层电活动生物标志物和相关结构的研究。本研究旨在识别SAD神经生物标志物,并基于EEG、ERPs、VEP和大脑连接网络,深入了解SAD患者和健康对照(HC)之间这些生物标志物的差异。此外,我们提出了改进描绘SAD大脑皮层电活动内表型方法的建议,例如,将EEG与功能性磁共振成像(fMRI)和脑磁图(MEG)等其他模态融合,以实现比单独使用EEG更好的效果,最终发展治疗选择过程,并回顾使用大脑皮层电活动测量进行SAD早期诊断和内表型检查的可能性。本文发表在Frontiers in Psychology杂志。
引言
社交焦虑障碍(SAD或社交恐惧症)的诊断首次出现在1980年出版的《精神障碍诊断与统计手册》第三版(DSM-III)中(Nathan, 2019)。SAD被定义为对一个或多个社交情境的明显和持续的恐惧,在这些情境中,一个人暴露于陌生人或可能受到他人审视(Radtke et al., 2020)。SAD患者因社交表现不佳而可能遭受尴尬或羞辱而感到焦虑(Langer et al., 2019)。因此,他们经常回避社交场合,当无法避免时,他们会经历严重的焦虑和压力。SAD与发展共病症的风险增加有关,特别是重度抑郁、急性悲伤和物质滥用障碍(Rapee and Spence, 2004)。基本上,SAD在生命早期就表现出来,预示着显著的社交功能损害、精神病共病以及持续的情感、认知和行为障碍(Grant et al., 2005)。SAD会严重损害患者的生活。当SAD症状通过达到回避社交场合的程度而扰乱个人的日常生活活动时(Givon-Benjio and Okon-Singer, 2020),这些个体通常符合SAD的诊断标准(Welander-Vatn et al., 2019)。例如,一个人可能会拒绝一份需要经常与新人互动的工作机会,或者可能会避免外出。SAD会干扰日常生活的重要方面,如学术和职业环境、家庭关系和社交活动(Frandsen et al., 2019)。如果不加治疗,社交焦虑通常在整个生命过程中保持高度稳定和持续(Yonkers et al., 2001)。
SAD影响约1500万美国成年人(占人口的6.8%),是仅次于特定恐惧症(8.7%)的第二常见精神病状况(Kessler et al., 2005)。目前的调查估计,西方国家社交恐惧症的终生患病率约为7-13%。然而,患病率百分比差异很大,容易出现误差,这是由于一些方法因素,如诊断标准、诊断阈值和评估方法(Furmark, 2002)。一些国家已调查了一般人群和大学生中儿童和青少年SAD的人口比例。SAD的发病率估计在13%到16%之间(Furmark, 2002; Baptista et al., 2012)。与成人研究一样,几种方法被用来调查SAD患病率估计的广泛变化。一项大型新西兰统计研究宣称,11.1%的18岁青少年符合SAD的标准(Baptista et al., 2012)。尽管如此,一项大型英国流行病学统计(Ford et al., 2003)报告称,只有0.32%的5-15岁儿童患有这种疾病,这高于其他精神疾病的发病率,如创伤后应激障碍(PTSD)和强迫症(OCD),但低于惊恐障碍、特定恐惧症和广泛性焦虑。这项研究发现,SAD的诊断率随年龄增长而略有增加,男性多于女性。一项大型美国儿童和青少年精神病学评估研究报告称,9-13岁儿童中有4.1%被诊断为SAD(Costello et al., 2003),而一项德国研究估计14-17岁儿童中SAD的发病率为4%(Iffland et al., 2014)。最严重的SAD形式可以在情感和身体上使患者残疾,许多人难以应对其他心理挑战,如内部感觉新奇寻求。例如,一项研究发现,约有一半的SAD患者有共病精神疾病、药物成瘾或酒精问题(Robichaud et al., 2019)。然而,大多数关于SAD流行病学的可用数据来自西方高收入国家。欧洲的流行病学数据与美国数据高度相关,证实了SAD的高患病率、共病率和发病率(Perna et al., 2020)。
此外,患有SAD的医疗病患者有更高的慢性病程或不完全康复的风险。因此,SAD患者需要特殊的医疗护理,例如缓解抑郁和焦虑症状的抗抑郁药物(Hansen et al., 2008; Potter, 2019)。认知行为疗法(CBT)和其他心理疗法被认为可以改善积极情绪并创造平衡的神经正念。一般来说,CBT在维持人际感觉方面发挥着关键作用。临床干预的证据表明,CBT在管理SAD方面发挥着基本作用(Morrison et al., 2019)。
根据本研究的研究目标,关键词是"社交焦虑障碍"。然后我们指定关键词("社交焦虑障碍")的替代词:社交恐惧症、delta-beta相关、额叶alpha不对称、功能连接、事件相关电位、视觉事件电位和有效连接。在指定搜索词后,将它们组装成一个在研究过程中应用的搜索系列。在本综述中,使用AND运算符将不同的搜索词连接成一个搜索链。以下在线数据库作为搜索的资源:Frontiers、IEEE Xplore、SpringerLink、BioMed Central、Wiley Interscience、ACM和ScienceDirect。我们还手动检查了相应主要研究的参考文献列表,以确保我们的综述完整。文章的选择是通过应用一组纳入和排除标准进行的。排除标准是:不包含任何列出关键词的文章、非英语文章、不完整文章、短文、重复文章以及所有与查询无关的研究。纳入标准基于以下内容设定:包含列出关键词的文章、英语研究和全文论文。根据纳入和排除声明,我们从253项主要研究中排除了93项研究。我们从主要和次要数据库搜索中收集了最终160项研究,以在本研究中报告。
本综述的主要目的是对休息、预期、视觉刺激任务和从社交任务中恢复期间最常研究的EEG、ERP和大脑连接性测量提供广泛概述。然而,我们发现有必要包括功能性磁共振成像(fMRI)和磁共振成像(MRI)研究,以验证EEG研究的发现。此外,我们排除了基于正电子发射断层扫描(PET)的研究,因为PET扫描使用患者通过注射接受的放射性示踪剂。然而,PET不是确定神经元位置的准确模型(Lefevre et al., 2019)。
社交焦虑障碍的认知模型
当代SAD认知模型(Clark and Wells, 1995; Hofmann, 2007; Rapee and Heimberg, 1997)被广泛用于扩展理论框架,以理解SAD中涉及的关键情绪维持过程(Arditte Hall et al., 2019)。这些框架明确指出,负面信念、对社交线索的偏见性反刍以及对焦虑刺激的回避都导致了SAD的扩展和维持。先前报告的模型(Arditte Hall et al., 2019)在理论上是一致的,主要关注SAD患者经历的维持属性。此外,Hofmann (2007)的研究强调了感知到的高社交标准在社交活动前触发预期焦虑的主要作用。
基于先前研究(Clark and McManus, 2002)提出的认知模型,SAD患者更有可能将社交信息视为威胁并理解为威胁,这是因为他们对自己、他人和周围环境存在认知偏见。例如,SAD患者通常为社交生活设定极高的标准,这自然会引发失败感和焦虑感(Clark and Wells, 1995)。如图1所示,Clarke的模型假设存在许多不同的过程参与维持SAD。面对社交情境的SAD个体更有可能回忆起具有破坏性的社交威胁假设,导致注意力转向自身并进行详尽的自我监控。不善交际和自我关注的行为进一步提高了焦虑的神经生理和身体属性的激活,如颤抖、出汗和对负面信念的反刍。SAD患者然后利用这些身体信号和破坏性假设来创造自己的负面心理表征(因为他们认为会受到他人的批评和负面评判)。此外,SAD患者习惯于采取安全行为(Clark and McManus, 2002; Kim, 2012),如回避或内部心理思考(例如,排练下一步该说什么或做什么;Reyes et al., 2020)。
该模型假设社交焦虑与不切实际的社交标准以及选择可实现的社交目标的不足有关。当经历和面对具有挑战性的社交事件时,SAD患者将注意力转向自身焦虑,将自己视为负面的社交对象,高估社交对抗的负面后果,想象自己无法控制情绪反应,并认为自己的社交技能不足以有效应对社交情境(Hofmann, 2007)。认知模型表明,当面临社交威胁时,SAD个体将注意力集中在内部,并进行详细的自我观察过程(Hofmann, 2000; Hirsch et al., 2003)。图1所示的模型取决于这样一种观点:各种重复的行为和思维模式与社交焦虑的躯体和情绪指标相关。SAD的主要认知模型(Clark and Wells, 1995; Hofmann, 2007; Reyes et al., 2020)在理论上是结构化的,强调社交事件后的负面信念作为SAD循环中的调节因素的重要性。
当SAD患者过度评估过去的社交表现时,就会出现负面反刍的做法(Brozovich and Heimberg, 2008)。研究人员发现,SAD个体在社交评估事件前后会进行过度和持续的负面反思(Rachman et al., 2000; Vassilopoulos et al., 2014)。严重社交焦虑的个体在社交事件(如即兴演讲)后会有更大程度的负面反刍(Edwards et al., 2003),或任何需要与他人互动的社交活动(Mellings and Alden, 2000)。先前的研究(Vassilopoulos et al., 2014)进行了首次实证阐述,阐明了特质焦虑与较高水平负面预期之间的关系,其中SAD个体报告称,他们对未来社交事件的反复负面想法具有侵入性、不可控性,并干扰了他们在社交事件中集中注意力的能力。虽然高社交焦虑(HSA)和低社交焦虑(LSA)的个体都会预测未来社交事件的代价和后果,但HSA(高社交焦虑)参与者的这些预测显著更为负面。当要求LSA和HSA的本科生参与引起社交焦虑的任务并在短暂演讲前进行事前反刍时,HSA参与者报告的焦虑程度更高,自我形象更为负面(Brown and Stopa, 2007; Blöte et al., 2019)。
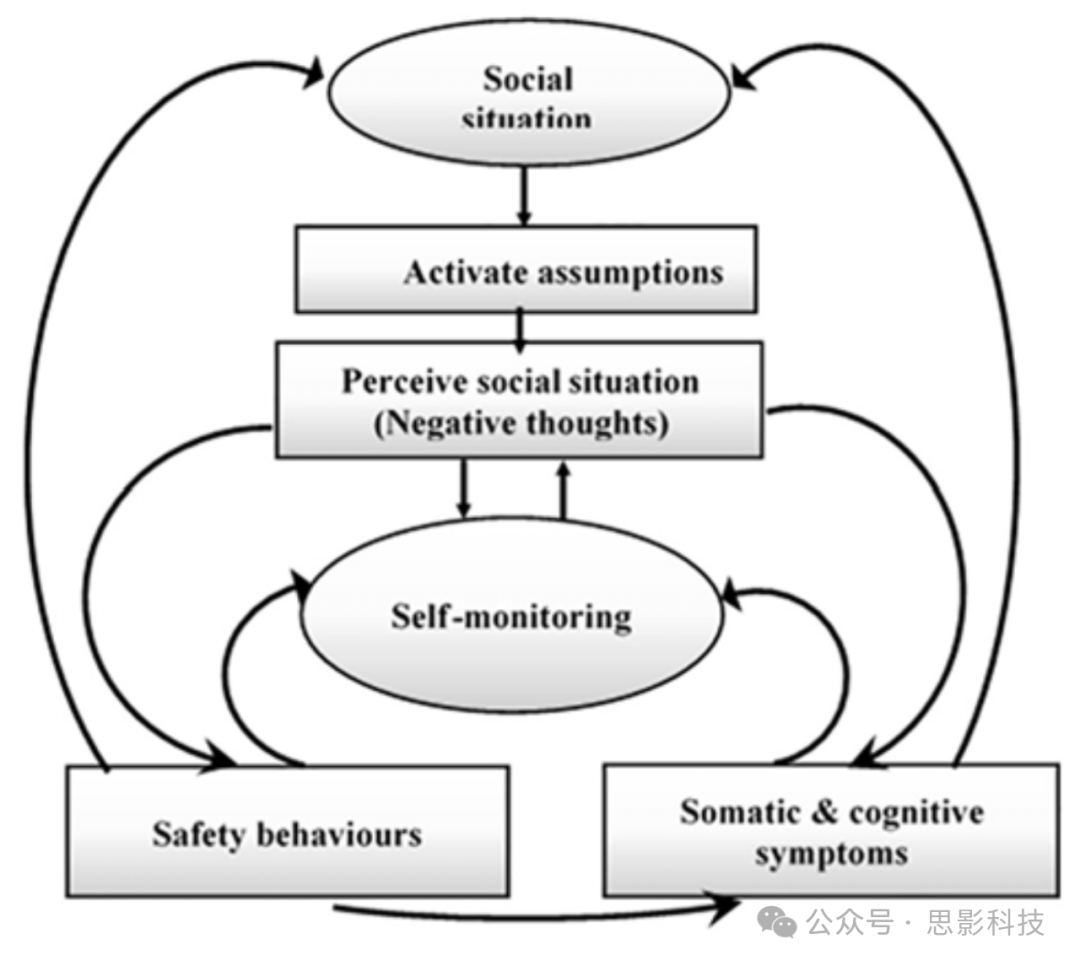
图1. SAD的认知模型(Clark and Wells, 1995)
一项先前的研究(Coles et al., 2002)通过评估参与演讲和进行社交互动的SAD患者的记忆视角来调查事后处理。正如预测的那样,严重SAD的参与者在几天后比低焦虑的参与者更频繁地回忆社交情境。许多研究者(Edwards et al., 2003; Abbott and Rapee, 2004; Perini et al., 2006)支持他们的假设,即社交焦虑水平较高的个体在治疗期间回忆起更多负面反应而非正面反应,而健康对照(HC)在记忆中不表现出任何偏见。关于事后处理,HSA(高社交焦虑)个体在1周间隔后对即兴演讲的负面因素和负面反馈表示不满,而HC则没有表现出这种倾向。SAD个体在事后反刍条件下经历更多负面想法,并创造更多反事实推理和信念。因此,严重SAD患者在阅读温和的社交情境后产生了更多负面的"假设性"陈述(Mellings and Alden, 2000; Rachman et al., 2000)。事后治疗已被应用于调查对事件进行负面评价的SAD个体和HC的记忆重激活。结果显示,无论事后治疗的类型如何,SAD患者回忆起的负面和尴尬记忆显著多于HC(Field et al., 2004)。总的来说,这些发现证实SAD患者以负面方式评估自己对社交事件的反应,并沉溺于持续的负面思考。这些特征表明,这些认知偏见的损害可以得到缓解,这是令人鼓舞的迹象。
社交焦虑障碍的生物标志物
各种研究对SAD信息处理的实证证据进行了批判性讨论,并利用几种社交认知任务来研究电生理效应和与这些任务相关的注意力(Davidson et al., 2000; Miskovic et al., 2010, 2011a,b)。这些研究旨在揭示与SAD相关的电生理和跨诊断症状维度之间的关联。所呈现的结果(MacNamara et al., 2013)显示,神经反应性维度与通过执行工作记忆任务建立的超越传统诊断界限的焦虑症状维度之间存在显著相关。基于这些结果,最近的一项研究(Yeung and Fernandes, 2019)调查了SAD是否与威胁记忆偏差相关,或者记忆偏差是否由威胁性或中性干扰物创造。对于威胁和中性目标,研究组之间在记忆方面没有差异。然而,严重SAD患者比轻度SAD患者对社交威胁的意识更显著增强和认识。
使用潜在类别增长曲线分析,证明了在不同时间评估中,唾液皮质醇水平较高且稳定的儿童唾液皮质醇水平更强。下丘脑-垂体-肾上腺(HPA)轴是下丘脑、垂体和肾上腺三个元素之间直接影响和反馈连接的网络组合。这三个元素及其通信构成了HPA轴,这是调节应激反应和调节身体多个过程(包括情绪)的神经内分泌通路的重要机制(Malenk et al., 2009)。另一种解释是,SAD是对紧张相关情况的反应,人类所见的核心内分泌反应是对激活HPA轴的应激反应,导致皮质醇增加(Condren et al., 2002)。HPA轴与皮质醇产生有关,较高水平的皮质醇与焦虑、恐惧和回避有关(Poole and Schmidt, 2019)。急性应激源(如焦虑)可以改变许多生物功能,如HPA轴(Foley and Kirschbaum, 2010)、身体防御系统(Steptoe et al., 2007)、自主神经系统的调节过程(Xhyheri et al., 2012)、HPA轴活动(Juruena et al., 2020)和肠神经系统(Ziegler, 2012)。心理上,急性焦虑是一种主观的负面体验,可能对认知产生积极和消极的影响(Starcke and Brand, 2012)。
与这些发现一致,研究者在以焦虑为应激源的基线条件下检查了SAD患者和健康对照的心率变异性(HRV)。SAD的小效应量支持SAD个体HRV的适度降低(Pittig et al., 2013)。在静息状态和执行社交认知任务时,SAD参与者的HRV没有显著差异(Harrewijn et al., 2018a)。相比之下,SAD与情绪识别任务期间的HRV呈负相关,参与者表现出HRV的显著降低(Madison, 2019)。与健康对照相比,SAD女性的HRV降低,服用精神药物的患者与未服药患者相比也出现HRV降低(Alvares et al., 2013)。
我们基于Rinck et al. (2013)和Harrewijn et al. (2016)的先前实验设计,设计了一项新颖的社交表现任务,如图2所示,其中所有受试者都被要求在观众面前完成自我展示任务,并对演讲进行录像,这与先前的研究(Miskovic et al., 2010)不同。EEG信号在六个阶段记录:静息状态、预期、演讲、视觉诱发电位(VEPs)和从社交任务中恢复。在社交表现任务的三个阶段(图2),要求参与者以0(否)到10(是)的评分表明他们想参与实验下一阶段的程度,以测量回避。我们是首次使用VEPs测试和通过整个社交任务的有效连接性分析来研究SAD严重程度的研究。

图2. 社交认知任务示例
该任务包括自我展示前后的视觉诱发电位(VEPs)测试;与更常用的设计相比,这种设计是新颖的。红色表示EEG记录阶段,黄色表示自我评估。修改自《认知、情感和行为神经科学杂志》16卷,Harrewijn A.等人,"社交焦虑的假设EEG测量:比较额叶alpha不对称和delta-beta跨频率相关"。
社交焦虑障碍的电生理相关性
不同的研究已经进行,以确定社交恐惧症个体诱发的预期焦虑的脑电图和电生理相关性。有许多方式可以捕捉大脑结构和功能的信息。三种常用和频繁使用的方式是MRI、脑磁图MEG和脑电图EEG。在这些方法中,EEG是研究大脑活动最通用和最具成本效益的方法,是研究社交焦虑神经相关物和获得大脑功能大规模连接模型的有效模型。在过去十年中,电生理信号的分析通过使用用于降维的盲源分离(BSS)技术不断普及。BSS还用于将混合信号中的信息分离到多变量记录的数据通道中,为更一般的数据分析准备多变量数据集,并提取不同条件下EEG数据的SAD生物标志物。以下小节概述了社交焦虑最突出的基于EEG的生物标志物。
社交焦虑中的Delta-Beta跨频率相关
EEG活动反映了数百万个空间排列的皮层神经元同步活动的时间聚合。最受关注的波形包括delta(0.5-4 Hz)、theta(4-7 Hz)、alpha(8-12 Hz)、sigma(12-16 Hz)和beta(13-30 Hz)。Delta-beta跨频率相关是研究社交焦虑的广泛使用的测量(表1)。这种测量代表了delta频带和beta频带振荡幅度之间的跨频率相关,被称为delta-beta相关。Delta和beta振荡之间的谱耦合已被证明与社交焦虑有关(Davidson et al., 2000; Miskovic et al., 2010, 2011a,b)。广泛来说,EEG频带与不同的功能和行为相关性相关。例如,慢波(SW)脑振荡如delta与负责动机、情绪和奖励处理的皮层下区域相关(Knyazev, 2012; Newson and Thiagarajan, 2019)。相比之下,快波反映了皮层间连接,在需要注意控制、认知处理和调节时被激活(Engel et al., 2001)。一般来说,当一个人警惕、专注、参与问题解决、判断、决策或心理集中时存在beta波。人们认为,额叶的快波(FW)和SW(慢波)频率之间的相关反映了皮层和皮层下回路之间的相互作用(Knyazev et al., 2019; De Pascalis et al., 2020; Knyazev, 2012)。因此,在与焦虑相关的行为特征中SW和FW活动同步增加被认为反映了两个不同脑区之间的神经通信。
一系列成人研究发现,额叶delta-beta耦合的大小对类固醇激素(包括皮质醇)敏感。最近的一项研究(Poole and Schmidt, 2019)将电生理研究扩展到7-17岁的儿童,发现基础皮质醇水平和SAD相对较高的儿童额叶EEG delta-beta节律强烈。这些发现表明,早期儿童期神经内分泌应激活动和社交焦虑的纵向模式可能与额叶缓慢和快速大脑振荡的EEG功率有关。先前的研究发现,在遗传皮质醇水平较高的年轻成年人中,额叶delta-beta耦合更强(Engel et al., 2001),并且在成年人外源性皮质醇给药后进一步加强(Knyazev, 2007)。这些结果表明,皮质醇水平可能与额叶SW和FW活动的增加有关。虽然在delta-beta相关性背景下的行为研究有限,但一些研究发现,在学龄前儿童和皮质醇反应性高的婴儿中,delta-beta耦合与社交恐慌的严重程度(Knyazev and Slobodskaya, 2003)和失调的恐惧(Schutter and van Honk, 2005)有更强的关系(van Peer et al., 2008)。一项代际研究发现,与健康父母相比,有社交焦虑父母的孩子在应对社交压力源时表现出更强的静息状态额叶delta-beta耦合,这可能是SAD的一个可能的内表型(Brooker et al., 2016; Harrewijn et al., 2018c)。更大的社交恐惧与delta和beta频带之间更大的正向额叶耦合相关。
表1. 与SAD相关的delta-beta相关研究总结
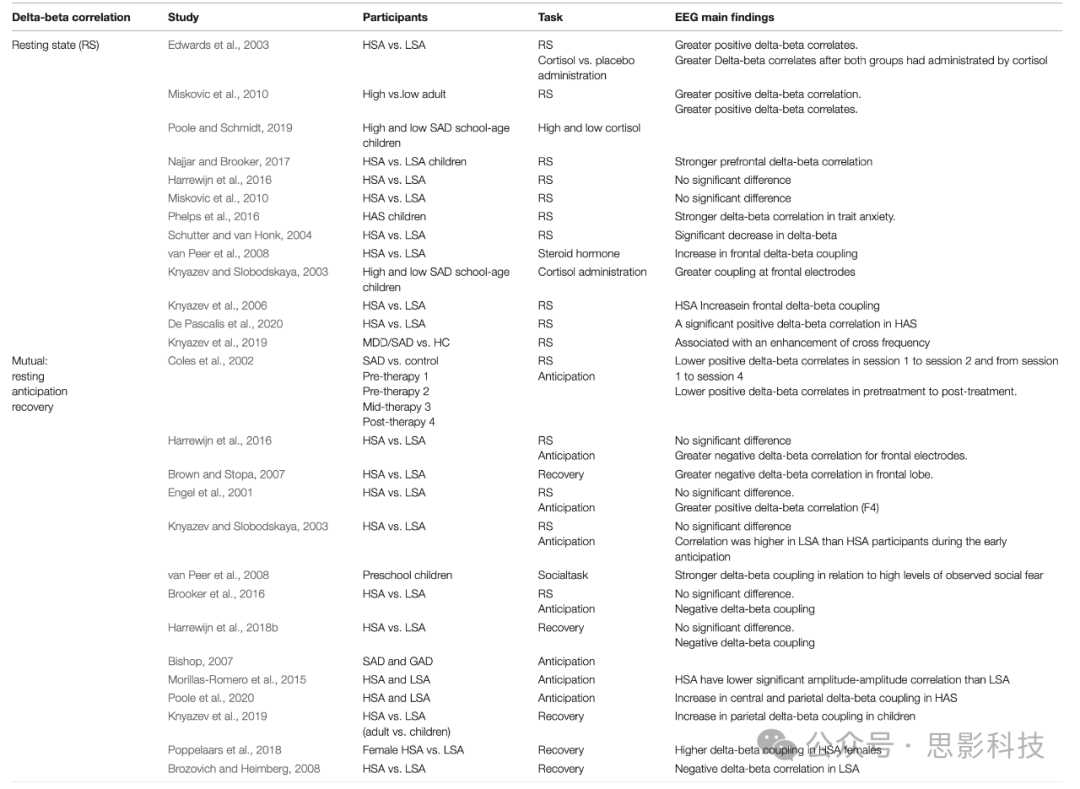
静息状态(基线)下的Delta-Beta相关
在学龄前儿童静息状态下记录了EEG信号,用于测试delta-beta跨频率个体差异是否与同情或残酷的父母行为相关(Knyazev and Slobodskaya, 2003; Poole and Schmidt, 2019)。一些证据表明,父亲表现出高水平严厉父母行为的学龄儿童在额叶电极上的耦合比父亲表现出低水平严厉行为的儿童更大。Delta-beta耦合是皮质神经网络下调皮层下网络情绪反应的实时指标(Najjar and Brooker, 2017)。在低威胁情况下表现出失调恐惧或高回避的儿童在基线状态下显示比在低威胁环境中表现出低水平回避的儿童更高的相关性(Phelps et al., 2016)。尽管对学龄前儿童delta-beta耦合的研究仍处于早期阶段,但初步发现表明,静息状态delta-beta耦合可能指示认知导向神经系统下调情绪反应神经系统的特质水平倾向(Phelps et al., 2016)。
有人试图检查静息区域大脑活动状态下男性delta-beta EEG谱耦合与内源性睾酮水平之间的关系。高睾酮水平的男性显示出非显著的delta-beta耦合(delta-beta解耦),而睾酮水平下降的男性显示出显著的delta-beta耦合。这些关系仅在额叶构建(Miskovic and Schmidt, 2009)。Delta-beta耦合的水平可以通过实验进行影响和操纵。例如,使用合成皮质醇(van Peer et al., 2008)和诱导焦虑(Knyazev et al., 2006)会导致额叶delta-beta耦合增加。相反,皮下给予睾酮导致delta-beta跨频率显著减少,这与类固醇激素的抗焦虑特性一致(Schutter and van Honk, 2004)。
关于基线跨频率耦合的几项发现已经报告,先前的研究(Miskovic et al., 2011b)证明,治疗SAD前的SW-FW(慢波-快波)相关比治疗后更强。尽管如此,SAD患者治疗前的delta-beta相关与健康对照相比没有显著变化(Miskovic et al., 2011b),快慢频带之间的跨频率耦合大于自然抑制较低的组(van Peer et al., 2008)。相反,两项研究没有发现特质和状态社交焦虑之间的任何差异(Miskovic et al., 2010; Harrewijn et al., 2016)。有假设认为,自发EEG中delta和beta振荡之间功率的大小与皮质醇和睾酮等激素水平有关;因此,研究静息状态下的社交焦虑是一个值得研究的好主意。先前的研究(Miskovic et al., 2010)发现了证据,表明额叶SW(慢波)和FW(快波)之间的谱耦合水平可用于区分社交焦虑个体,并量化社交互动期间SAD的严重程度。具体而言,在静息状态下,HSA成年人右额叶电极(右半球)的delta-beta耦合显著大于LSA成年人(De Pascalis et al., 2020)。
预期状态下的Delta-Beta耦合
最近的一项研究(Poole and Schmidt, 2019)发现,具有不稳定和易怒气质前兆的SAD学龄前儿童表现出相对较高的额叶delta-beta相关,如Knyazev等(2019)和Poole等(2020)所报告的。显然,虽然这项研究中SAD儿童的整体额叶跨频率耦合更强,但在检查单独的半球耦合时,这种模式似乎主要由右额叶控制。这项研究中呈现的结果与先前观察到的相同,HSA参与者在社交表现任务期间显示出低于LSA参与者的显著幅度-幅度相关,而LSA参与者在早期预期和所有条件下都显示显著值(Morillas-Romero et al., 2015)。Delta-beta耦合作为SAD的电皮质测量,被认为在社交焦虑参与者预期社交压力时更有前途。与LSA个体相比,HSA组在认知治疗前的预期期间显示出正delta-beta相关的增加。SAD患者在预期期间SW和FW之间正跨频率耦合的增加在CBT后减少,高或低SAD个体之间没有差异(Miskovic et al., 2011a)。SAD患者在预期期间表现出比低SAD个体更高的正delta-beta相关(Miskovic et al., 2010)。相反,有报告称SAD患者比低SAD患者表现出更大的负delta波和beta波之间的耦合(Harrewijn et al., 2016)。此外,在预期自我展示时,HSA发现更高的右额叶大脑功率。人们认为,负SW-FW耦合可能是由大脑内部(如杏仁核和岛叶)和外部(如皮质)位点之间连接在相反方向上增加所解释的。已知的广泛性社交焦虑障碍(GAD)(Bishop, 2007)和SAD(Miskovic and Schmidt, 2012)患者皮层下和皮质网络之间的失衡,可能导致负delta-beta耦合。
当代电皮质指数表明,脑电图中的中额theta(4-8 Hz)振荡为大脑处理社交排斥提供了新的见解(Knyazev et al., 2019)。这些发现表明,中额theta(4-8 Hz)振荡对社交排斥非常敏感,但只有当同伴排斥是意料之外的时候,这表明额叶theta受到广泛不同神经网络的控制,涉及显著性感知和冲突检测(van der Molen et al., 2017)。Delta和beta之间的相关性被证明对外部影响因素非常敏感,因为它区分了完美和不经意的表现条件(Miskovic et al., 2011b)。
恢复状态下的Delta-Beta耦合
虽然事后处理是社交焦虑的一个非常重要的方面,但很少有研究调查预测社交压力条件后的delta-beta耦合(恢复)。在Harrewijn等(2016)的研究中,与较低SAD的个体相比,较高SAD的个体在展示自己的优点和缺点后表现出更大的负delta-beta相关。这种效应反映了大脑外部(皮质)和内部(皮层下)在事后处理状态下的不可测性。这与先前对参与社交事件背景下事后反刍的SAD个体的认知行为研究结果一致(Clark and McManus, 2002; Brozovich and Heimberg, 2008)。根据Poppelaars等(2018)的研究,额叶delta-beta是高或低社交焦虑测试分数女性的压力调节指标。结果证明,delta-beta耦合在社交表现任务的预期阶段区分了HSA和LSA。LSA参与者在所有条件下(静息、预期和恢复)表现出更显著的差异,表明额叶delta-beta对特质焦虑敏感,反映了适应性压力神经调节机制(Poppelaars et al., 2018)。因此,增加社交表现范式中恢复状态的研究数量似乎是值得的,未来的研究应验证恢复期间的delta-beta相关是否可以作为SAD的假设EEG测量。
上述研究为神经元delta-beta谱相关作为事后处理中神经元电生理活动量化的可能性提供了洞察,并提供了证据,表明SAD中delta-beta子带之间的相干性比任何其他频域生物标志物表现得更强。预期和事后状态下的SW-FW耦合似乎是SAD个体电活动测量的更合适候选。经验上,SW-FW耦合被认为代表与焦虑相关的皮质(beta)和皮层下(delta)区域之间的动态通信(Knyazev et al., 2006; Knyazev, 2012)。SAD个体中增加的SW-FW耦合与一些fMRI研究的结论相似,这些研究发现GAD个体的皮质和皮层下区域更相同,而在SAD个体中更是如此(Miskovic and Schmidt, 2012)。皮质层和皮层下层之间的失衡也与心理治疗研究中发现的事后处理偏差一致(Kashdan and Roberts, 2007)。另一个重要发现是,边缘(皮层下)和边缘旁(皮质)层的神经增大似乎与社交威胁的功能机制(包括对情感处理的更多注意)相关(Miskovic and Schmidt, 2012)。
社交焦虑中的额叶Alpha脑电图不对称
半球不对称和情绪理论是一个有影响力的理论,指出具有不同特征的个体之间的差异。可以通过EEG额叶alpha不对称功率的不对称模式来研究情绪的神经基础(Davidson, 1992)。特别是,相对较大的EEG左额叶功率与趋近行为相关,而较大的右EEG额叶功率与回避行为相关(Meiers et al., 2020)。然而,应该注意的是,回避/趋近效应与自信/抑制行为之间并不存在直接一致性。例如,愤怒是一种与趋近行为相关的自信感觉,也被发现与较高的左额叶皮质活动相关(Harrewijn et al., 2019)。
额叶大脑alpha不对称通常通过测量左额叶电极上的EEG alpha功率减去右半球同源电极的方式来表示(Davidson, 1998)。因此,alpha功率与大脑活动呈负相关,负的不对称功率分数反映较高的相对右侧(即减少的右额叶alpha功率),正值反映较强的相对左额叶激活(即减少的左额叶alpha功率)(Zhang et al., 2020)。额叶EEG大脑alpha不对称已被广泛研究,以检查大脑中的趋近行为和情绪处理系统(Harmon-Jones et al., 2010)。此外,额叶alpha不对称(FAA)已被证明与行为回避和抑制有关(Allen et al., 2004)。相反,许多研究已经证实FAA与行为抑制(社交焦虑)之间的关系是复杂和不相关的(Coan and Allen, 2004)。因此,在表2中我们列出了与SAD相关的FAA的相关EEG研究。
表2. 与SAD相关的FAA(额叶Alpha不对称)工作总结
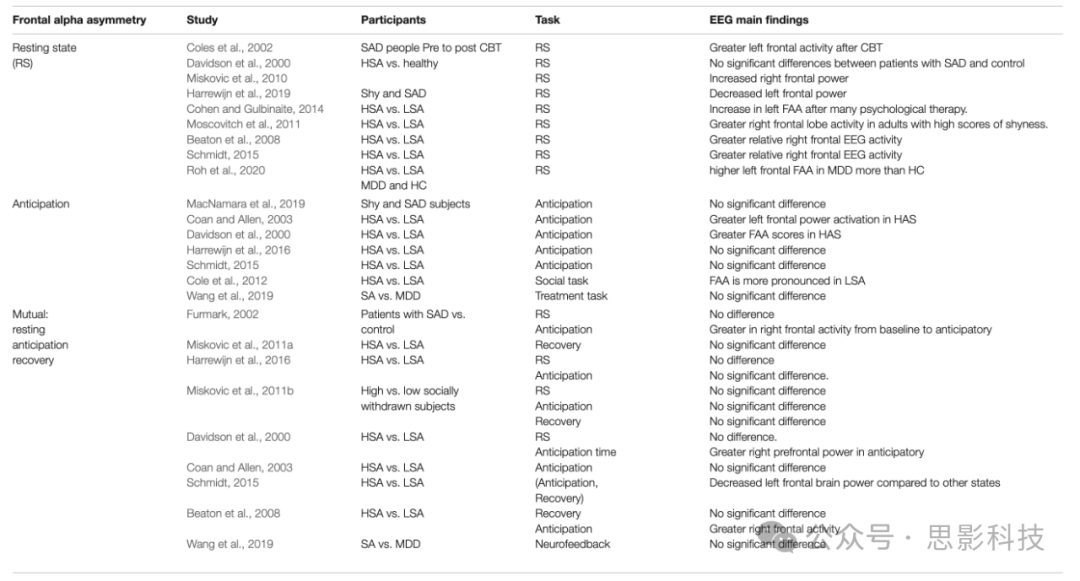
静息状态下的额叶Alpha不对称
FAA(额叶Alpha不对称)已经在静息状态下进行了研究,要求参与者坐下并眨眼或闭眼一段时间。关于SAD静息基线状态下FAA不对称性的文献似乎各不相同且不一致(Roh et al., 2020)。例如,在一项研究中,SAD个体在多次心理治疗后左FAA增加(Cohen and Gulbinaite, 2014)。然而,该研究没有将其结果与对照组进行比较。静息状态下的FAA也在与社交焦虑相关的症状(如害羞)的背景下进行了研究,观察到害羞得分高的成年人比得分低的成年人右额叶活动更高(Moscovitch et al., 2011)。相反,其他研究发现SAD个体与正常健康对照以及重度和中度SAD患者之间在静息状态FAA方面没有显著差异(Davidson et al., 2000)。还发现,静息状态下害羞与右额叶EEG alpha不对称之间的关联只有在控制同步和并发抑郁情绪后才会出现。在控制并发抑郁后,自我报告害羞程度高和社交退缩程度高的个体在静息状态下表现出比轻度社交退缩个体更大的相对右额叶EEG活动(Beaton et al., 2008; Schmidt, 2015; Harrewijn et al., 2016)。
在一项最新研究中,发现FAA调节了行为抑制与事件相关负性的社会效应之间的关系。FAA(额叶Alpha不对称)与行为抑制或焦虑无关,也不能缓解早期行为抑制与后续焦虑症状之间的关系(Cole et al., 2012)。高社交焦虑(HSA)和低社交焦虑(LSA)参与者在静息状态下的FAA有显著差异。HSA参与者在所有焦虑状态下的FAA也表现出差异,这意味着FAA可以用作SAD的指标。先前的研究(Harrewijn et al., 2019)显示更大的左EEG额叶大脑激活,这被认为与趋近行为相关,但与抑制行为分数无关。减少的左额叶活动被认为反映了情绪调节困难,如(Zhang et al., 2020)所报告的。
预期状态下的额叶Alpha不对称
认知模型强调了SAD个体预期暴露于可怕的社交情境时情绪处理的重要性。特别是,通过给予一个即兴自我展示任务来研究SAD中的预期时间,要求参与者准备一个关于一般主题或个人属性的演讲。SAD个体通常期望参与社交互动,这导致产生更多负面结果和更多自动负面情绪循环。社交焦虑的人担心他们会以不适当的方式行事,因为这可能导致周围人的负面评价(Hofmann, 2000; Clark and McManus, 2002; Hirsch et al., 2003)。
从科学角度来看,一直认为较大的左额叶功率不对称是抑郁和焦虑的持续生物标志,并在任何有影响的触发因素期间区分SAD个体和健康对照(Coan and Allen, 2003)。在最近的一项研究中(Coan and Allen, 2003),通过研究高低SAD参与者在既定情节回忆任务中的神经差异来评估FAA。结果表明,低SAD个体(特质焦虑)表现出显著高于高或中等SAD个体的右额叶EEG大脑半球激活。大多数研究似乎一致认为,FAA与参与社交互动之前的时间内的SAD相关(Davidson et al., 2000; Beaton et al., 2008)。例如,在先前的研究中(Davidson et al., 2000),评估了SAD个体在预期就未知主题发言时的前额FAA。SAD患者在任务准备期间表现出比静息状态更高的右额叶活动。同样,HSA参与者在观看同伴以焦虑方式说话的视频时,在预期自我展示期间表现出升高的右额叶和侧皮质功率(Beaton et al., 2008)。
先前的调查显示,在期待发表演讲时,重度焦虑和轻度焦虑的人之间SAD的效应没有显著差异(Schmidt, 2015; Harrewijn et al., 2016)。尽管(Schmidt, 2015; Poole and Schmidt, 2019)报告HSA和LSA之间没有显著差异,但害羞与右前额叶活动增加有关(Wang et al., 2019),仅在管理心理压力因素后才出现。这些神经生理EEG结果的差异可以通过各种解释来解释。首先,SAD的效应只能在由压力源(言语攻击)引起的极端社交焦虑等级上量化。极端社交压力源对重度SAD的影响比轻度SAD更高(Davidson et al., 2000)。然而,14名SAD参与者构成了一个统计上较小的人群,无法推广结果(Davidson et al., 2000);因此,这些结果需要谨慎解释。此外,只有抑制性HSA个体表现出右前额叶EEG alpha功率的增加。当HSA参与者暴露于社交挫折任务(观看同伴视频)时,正常样本中的群体效应被发现更为显著,这可能在即将到来的事件中预期(Cole et al., 2012)。没有压力诱导情况的社交任务可能不会引起FAA的激活上升,如先前研究所报告的(Harrewijn et al., 2016)。
其次,如果健康对照组在执行任务时不表现出焦虑,SAD的效应可能会显著可测量。例如,先前研究(Davidson et al., 2000)中的健康对照参与者在预期期间没有表现出主观焦虑特质的增加,而在另一项研究(Harrewijn et al., 2016)中,LSA参与者记录了特质恐惧症分数的增加。健康对照中特质恐惧症分数的增加可能阻碍FAA显著差异的识别。第三,一些研究者(Beaton et al., 2008)关注两种状态(预期和静息状态)之间的区别,而大多数研究只关注预期状态(Beaton et al., 2008; Schmidt, 2015)。然而,预期和静息状态数据之间没有报告社交焦虑的显著效应(Harrewijn et al., 2016)。第四,预期时间内SAD对FAA的影响可能与预期时间的差异有关。特别是,一些研究者使用了3分钟的预期时间和2分钟的计划会话(Davidson et al., 2000),这可能会在此阶段再次增加社交焦虑。即使使用相对较长的预期期(6分钟),也没有研究发现额叶不对称效应(Harrewijn et al., 2016; Wang et al., 2019)。这与先前报告的不对称效应结果相反(Schmidt, 2015)。一般来说,使用延长预期阶段的任务无效可能是由于熟悉效应。本质上,如果预期时间很长,SAD个体可能会习惯它并最终表现出较少的正面活动。在未来的研究中,在比较预期期间不同时间点或其他状态的FAA时,应考虑任务的潜在熟悉影响。
恢复状态下的额叶Alpha不对称
对于SAD个体来说,从社交压力情境(如公开演讲)中恢复可能会导致事后处理中焦虑的增加。正如一些研究者所指出的(Clark and McManus, 2002; Brozovich and Heimberg, 2008),SAD的情绪事后处理以持续的思考为特征(例如,对先前社交互动的不利担忧)。增强痛苦记忆的检索和对负面假设的关注被认为会维持SAD的症状(Morrison et al., 2019)。心理任务(社交压力源)后的持续反刍可以通过FAA来研究。到目前为止,有两项研究对HSA和LSA参与者进行了自我展示后恢复状态的FAA测量。神经研究(Davidson et al., 2000)未能揭示SAD和健康对照个体之间的FAA关系。在HSA和LSA个体中研究了FAA与SAD之间的关系(Harrewijn et al., 2016)。尽管研究不足,但先前的研究表明,FAA不是恢复状态下SAD参与者情绪处理的候选生物标志物,因为FAA无法解释SAD中的情绪调节。静息和事后处理期间的FAA与SAD无关。然而,预期时间内的FAA似乎是SAD的潜在神经电生理候选生物标志物,但只有在焦虑严重时才如此。尽管如此,最近的研究发现,应用神经反馈后,抑郁和焦虑特质降低,但FAA在所有时间内保持稳定(Wang et al., 2019)。这可能表明FAA不是SAD的可能特质标记,但可能与某些明确的极端压力源存在时的SAD有关。FAA相关文献中的一项先前研究(Thibodeau et al., 2006)报告了不一致的发现,表明FAA可能是其他精神疾病(如重度抑郁障碍(MDD)和广泛性焦虑障碍(GAD))的潜在生物标志物。需要更多研究关注SAD与其他精神疾病的共存。
社交焦虑中与信息处理相关的ERPs
事件相关电位(ERPs)是头皮记录的电压波动,用于测量大脑对时间锁定事件的反应(Herrmann and Knight, 2001)。ERP反映了数千个同时进行的大脑过程,评估感知处理、注意选择、认知和感觉运动耦合。通常,ERP研究在SAD研究中用于描述与认知行为和视觉处理中涉及的刺激过程相关的电皮质测量(Klawohn et al., 2020)。ERP代表电皮质活动的变化,对特定刺激有时间锁定,并提供神经激活模式时间结构的精细描述(Koivisto and Revonsuo, 2010)。ERP有能力为神经元刺激处理的即时和后期阶段分配客观洞察(Herrmann and Knight, 2001)。
图3显示了一项使用自相关积极或消极社交反馈的研究中典型ERP信号及其最常见的组成部分(Cao et al., 2015)。刺激后100毫秒触发的ERPs可能受刺激物理因素的调节,而非感知因素(Herrmann and Knight, 2001; Sa, 2005)。然而,高度突出的刺激物或刺激呈现顺序的改变可能影响这些早期ERP组分(Meynadasy et al., 2019; de Bruijn et al., 2020),反映了对感知和注意的影响(Eimer and Driver, 2001)。某些研究报告,SAD对P1振幅显示出显著的敏感性(Hagemann et al., 2016; Fang et al., 2019; Torrence et al., 2019)。
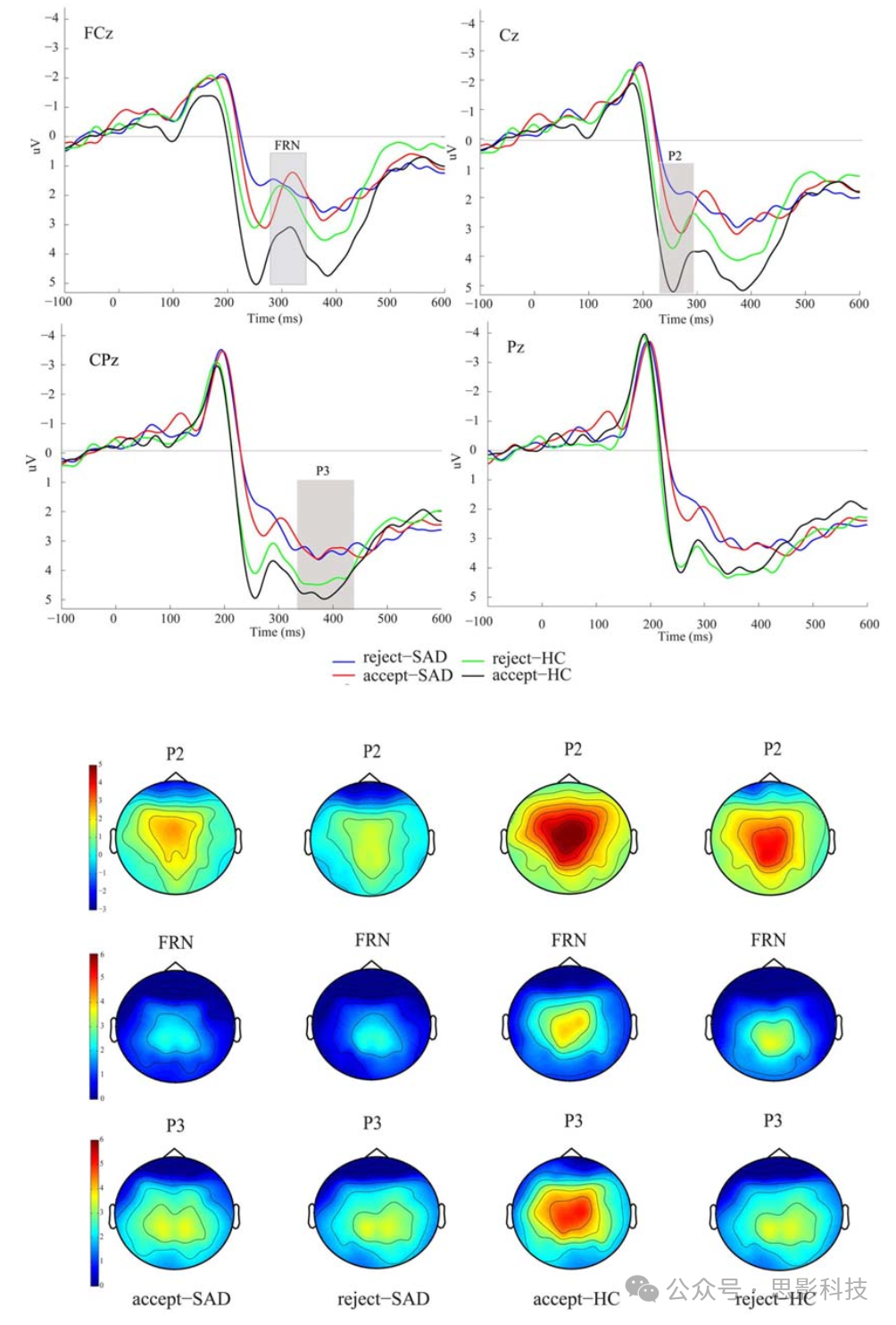
图3. SAD和HC组在中线电极(FCz, Cz, CPz和Pz)上两种反馈类型的总平均事件相关电位(ERPs)和地形图。[四种条件:拒绝-SAD(蓝色),接受-SAD(红色),拒绝-HC(绿色)和接受-HC(黑色)]。
N1(称为N1)是第一个负向组分,基本上被认为捕捉早期感知编码和面部识别。N1通常在外部刺激触发后130-200毫秒达到峰值,主要在枕颞叶(Sa, 2005)。大量研究致力于研究N1组分。这些研究强调N1振幅与视觉刺激和情绪表达相关,而其他研究则没有显示这种对情绪的敏感性(Kolassa and Miltner, 2006)。负偏转ERP发现表明,当受试者识别不同面部表情时,视觉处理的初始阶段可能受到情绪表情的影响。N1似乎不受SAD对特定任务反应的调节。SAD参与者和健康对照在对情绪任务的神经反应中N1振幅没有显示任何显著差异(Kolassa and Miltner, 2006; Kolassa et al., 2009)。在对情绪面孔的视觉任务反应中,HSA和LSA组之间的N1振幅没有差异(Peschard et al., 2013; Hagemann et al., 2016)。只有一项研究显示,在情绪视觉任务中分类愤怒肖像时,SAD组在右顶叶的N1振幅增加,而HC组在同一位置表现出较小的振幅(Peschard et al., 2013)。有人得出结论,HSA和HC个体之间的差异主要观察到早期P1和N1组分(Staugaard, 2010)。与早期研究相反,有人指出SAD与更高的P1振幅相关,而N1保持不变(Hagemann et al., 2016)。最近关于N1振幅的研究显示,SAD和HC个体在对幸福和恐惧面部表情的反应中没有差异(Torrence et al., 2019)。
P2也称为P200,是第二个负向偏转,在前额皮层刺激开始后150-250毫秒达到峰值(Schmidt, 2015),被认为反映了大脑中神经过程的突触后活动。P2的振幅可能受到视觉刺激不同方面的调节,如感知处理和注意力。一般来说,P2振幅对一致目标的反应高于不一致目标或均匀刺激,P2组分通常随不寻常的目标刺激而增加(Kolassa et al., 2009; MacNamara et al., 2013)。通常,P2振幅对快乐和恐惧刺激的反应高于中性刺激(MacNamara et al., 2013)。P2振幅对情绪面孔的反应增加,这可以促进理解先前信息如何塑造未来反应,以及它在皮层中的情绪重要性(Normann et al., 2007; MacNamara et al., 2013)。
当要求SAD个体将认知注意力集中在面部肖像上时,P2组分似乎对视觉认知处理敏感,但对社交焦虑不敏感。SAD和HC组之间P2电压值偏转没有显著差异(Kolassa and Miltner, 2006)。此外,HSA和LSA个体在Stroop任务中P2振幅没有差异(Peschard et al., 2013)。HSA人群没有表现出任何注意偏差(Kolassa and Miltner, 2006)。在处理面部任务时,SAD和HC组之间P2振幅没有差异(Hagemann et al., 2016)。此外,认知任务后SAD和HC组之间第二个正向偏转组分(P2)没有差异(Kolassa et al., 2009),HSA和LSA之间也没有差异(Peschard et al., 2013)。在这项研究中,我们进一步强调了这些组分和潜在处理任务的差异,如表3所示,并研究了这些注意和认知任务对SAD个体的影响。
表3. 与SAD相关的ERPs工作总结。
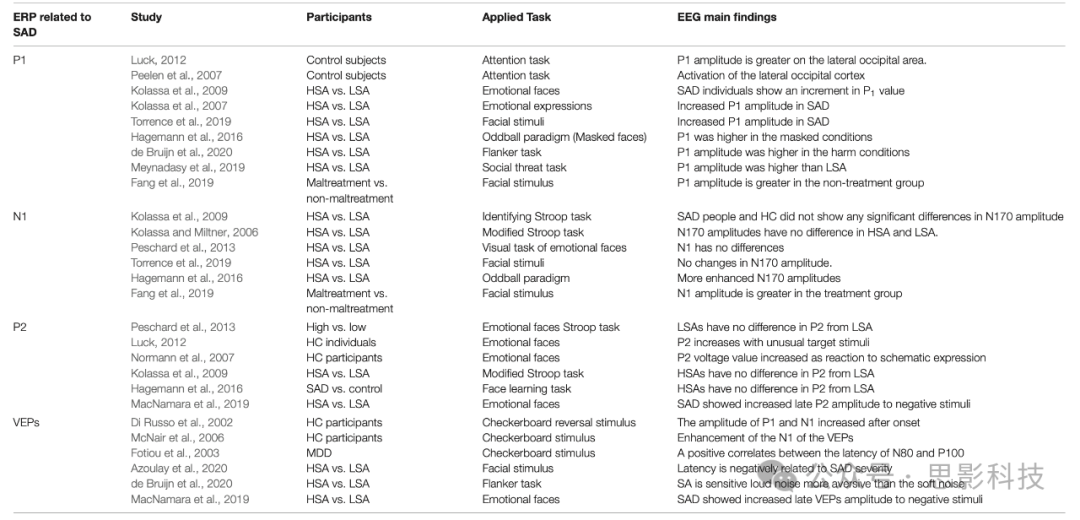
视觉诱发电位(VEPs)是记录枕叶电极视觉处理强度和量化注意力分布皮层电位相关物的理想工具,可同时从视觉皮层上方的头皮进行EEG记录。在先前的研究中(Normann et al., 2007),VEP被用作研究大脑皮层电位激活的可行范式。VEPs同时刺激平均EEG信号,主要反映神经元兴奋性和抑制性突触以及来自各种感觉信息位点的动作电位。VEP数据和fMRI模态的整合通过追踪第一VEP组分中视觉皮层的激活,导致电皮质活动源(Di Russo et al., 2002)。报告的结果显示,长期视觉刺激会在皮层反应中产生可塑性改变,刺激诱导开始后P1和N1的振幅显著增加。在一些实验研究中(Di Russo et al., 2002; McNair et al., 2006; Normann et al., 2007),使用9 Hz棋盘格反转刺激2分钟导致VEP的N1振幅增强。
据报道,非典型患者的N1和P1潜伏期显著缩短,而抑郁症患者则延长(Fotiou et al., 2003)。N1和P1潜伏期之间存在正相关关系,抑郁指数与害怕的社交互动的发生呈负相关。模式反转潜伏期与害怕的社交互动分数之间报告了更大的负相关关系。一般来说,SAD个体比LSA更能检测到更快的视觉刺激偏移,如最近报道的那样(Azoulay et al., 2020),这意味着检测潜伏期与SAD严重程度呈负相关。
最后,可以得出结论,几种关于SAD的研究已经证明,SAD与P1和晚期正潜伏期振幅的增加以及N1振幅的恒定性更强相关(MacNamara et al., 2019)。必须强调的是,大多数先前的实验包括一个小规模人群(SAD为10-22人),这不足以推广最终的神经结果。SAD与ERP发现之间的关联在先前的综述中已经得到更深入的探讨(Harrewijn et al., 2017)。除了ERP研究,最近,SAD已通过大脑各部位之间连接图的变化进行了研究。在接下来的部分中,我们概述了各种类型的连接,并总结了基于这些连接的SAD最新发现。
大脑连接性
最近,研究人员对通过大脑区域间激活变化以及分布在不同大脑区域的神经元集合之间的相互作用来研究正常和病理大脑功能的兴趣日益增长(Beaty et al., 2019)。大脑各个区域之间的这种相互作用网络被称为大脑连接性(Busby et al., 2019)。大脑连接性可以从结构连接性、功能连接性和有效连接性方面来解释。物理上从一个大脑区域延伸到另一个区域的纤维通路代表结构连接性(Sokolov et al., 2019)。这些纤维束可以通过MRI和弥散张量成像(DTI)技术最好地观察到(Richards et al., 2015)。除了结构上的连接,大脑区域还可能在功能上连接,即在执行功能时不同大脑区域之间的神经活动在统计上变得相关。这种类型的统计依赖性通常被称为功能连接性(FC)(Greicius et al., 2009)。为了避免FC在双向性和易受第三方连接影响方面的根本缺陷,提出了一个相对较新的概念,称为有效连接性(EC)(Valdes-Sosa et al., 2011)。EC反映了驱动因素(启动外部力)和响应(被驱动系统)之间的因果关系;它对一个大脑区域的神经系统比对其他神经系统有更直接的影响,并定义了它们之间的动态定向相互作用(Valdes-Sosa et al., 2011)。这种影响可以通过信号直接估计,称为数据驱动EC,或根据指定不同大脑区域之间因果联系的模型命名,称为模型驱动EC(Bakhshayesh et al., 2019)。在接下来的两个小节中,概述了使用大脑FC和EC对SAD进行的最新研究和发现,如表4所示。
表4. 与SAD相关的大脑连接性工作总结

功能连接性
神经科学中的功能连接性代表大脑不同解剖单元内同时进行的活动。如果两个区域在神经系统内的活动随时间高度相关,则认为它们在功能上是连接的。FC基本上是一个统计概念,它捕捉分布式远程神经单元的统计依赖性变化。统计依赖性被过度应用于描绘大脑中对任何神经刺激或认知任务反应改变其激活水平的区域,可以通过量化相关矩阵、相干性或相位锁定来确定。在用于FC估计的模态中,EEG是首选,因为它比其他神经成像技术(如fMRI)具有更高的时间分辨率。FC估计主要依赖于使用不同大脑部位的静息状态fMRI数据。FC水平通常在大脑中所有神经活动单元之间进行量化,无论它们是否通过线性连接相连。与结构连接性不同,FC受时间影响很大:皮质振荡模式在几个特定时间范围内同时在统计上波动(10-100毫秒)。值得注意的是,FC并不明确指定特定的方向效应或基本物理模式。当代研究发现,在静息状态默认网络模式(DMN)中可以揭示视觉、运动、语言和工作记忆系统中各个回路单元之间的区域间连接。由于大脑边缘系统在情绪处理功能中的关键作用,许多研究将杏仁核和岛叶视为分析社交焦虑个体FC的主要感兴趣区域。
在最近一项关于SAD的研究中,使用静息状态网络的FC,记录了32名广泛性社交焦虑障碍(gSAD)患者和32名统计上健康对照的EEG数据(Xing et al., 2017)。对EEG电极应用加权相位滞后指数(WPLI)显示,中前额区域振荡theta节律相干性的增强波动增加,证明gSAD患者在静息状态下的连接性高于健康对照。15名gSAD个体在应用CBT后,左边缘单元(杏仁核)的FC相对于基线显著降低(Zhou et al., 2017)。fMRI分类分析中的多变量模式算法应用于20名SAD和20名健康对照个体,显示用于区分社交焦虑受试者的链接的一致性广泛位于DMN、感觉运动神经元、视觉网络和突显网络内部或之间(Bhaumik et al., 2017)。正如预期的那样,SAD患者在情绪范式中记录了杏仁核过度激活和左杏仁核功能耦合减弱(Hahn et al., 2011)。同样,SAD患者在面对潜在的社交刺激、负面情绪(Etkin and Wager, 2007)和严厉的面部知觉(Birbaumer et al., 1998; Phan et al., 2006)时一直表现出更大的杏仁核激活。还观察到左右前扣带皮层之间的beta连接性降低(Imperatori et al., 2019)。然而,由于先前研究的样本量小,我们认为不可能对SAD和健康对照组的最终FC结果得出明确结论。此外,这项研究没有确定焦虑状态的变化,这使得这种解释特定于特质焦虑。
此外,实验证明SAD个体的岛叶皮层(岛叶)也出现过度活跃,激活强度被认为与焦虑水平相关。在应用适当的抗焦虑治疗后,岛叶皮层活动已经显示出降低。例如,进行了两项MRI研究来检查和比较SAD个体与健康对照个体的岛叶激活,结果发现SAD和健康对照组的岛叶体积存在统计学上显著的差异。SAD患者的双侧岛叶体积减少,表明SAD个体存在异常的神经解剖网络(Syal et al., 2012; Kawaguchi et al., 2016)。另一项FC研究表明,岛叶是内部和外部信息之间的通信链接,用于创建内部身体感觉的意识。异常的岛叶-前额叶连接性显示,与健康对照相比,SAD个体对威胁刺激的反应更为过度(Klumpp et al., 2012)。最近,功能性MRI研究报告,在社交表现任务中,SAD组个体的双侧后岛叶激活程度高于健康对照,这可能反映了自我知觉SAD的早期标志(Wang et al., 2019; Yuan et al., 2020)。SAD的功能神经成像实验主要关注对焦虑、挑衅或情绪面部线索反应的区域神经活动。
尽管fMRI和EEG技术已成为绘制人脑大规模网络的强大工具,但它们有一些重要的局限性。这些局限性包括确定网络节点、每个单独节点的连接分析、体素级全脑估计、节点组和FC变化。FC并不特别阐明大脑中神经信号的方向性影响。关键是要检查组间的可变性,fMRI对头部运动和扫描期间心理状态的变化敏感。
有效连接性
有效连接性被认为是物理连接和功能连接性的结合,因为EC可以有效地识别大脑中一个神经单元对其他神经单元的因果影响网络。基本上,动态影响是通过模型的系统性干扰推导出来的,因为原因必须在时间上超过结果,这是通过数学时域分析实现的。用于提取EC参数的几种方法需要一个特定的系统,涉及物理模式。EEG的几个有效特征(高时间分辨率、设备便宜和便携性)使其成为研究认知功能神经连接的适当模态。以前,研究了EEG功率谱以阐明情绪处理中不同频带的变化。功能性大脑连接可以通过多种技术来量化,例如,计算互信息、交叉相关、EEG信号之间的相互关系,或时域和频域中的相干性。计算EC比确定功能性大脑连接更具挑战性。如前所述,FC量化统计依赖模式,而EC确定一个神经单元对其他单元的定向影响网络。
目前正在不断研究和开发几种用于评估EC的数学模型。一种称为协方差结构方程建模的模型将EC强度分配给结构路径,这与在任何特定实验中发现的协方差完全相似(Friston et al., 2003)。这个过程的泛化被称为动态因果建模(DCM)(Tononi and Sporns, 2003),它在贝叶斯结构样本中运作,以评估和得出神经元素中的因果效应。不同的方法被用来识别各种神经位点之间的广泛大脑相互作用及其因果联系,包括使用扰动方法来捕捉两个大脑区域相互作用程度的有效数据驱动方法(Liao et al., 2010)。也可以使用时间序列分析来估计有效连接。这些方法中的一些基于Granger因果性(GC)的解释概念(Ding et al., 2006; Seok and Cheong, 2019)。此外,传递熵因果性测量已被广泛使用(Schreiber, 2000),旨在通过考虑一个神经单元的状态基于另一个神经单元的状态转移概率的有效性来揭示神经单元中定向输入数据的互换。EC因果性对数据减少、采样率、窗口模式和状态空间选择高度敏感。
一项基于fMRI的静息状态研究(Liao et al., 2010)是首次通过评估SAD个体的EC来揭示和量化异常大脑网络的研究。发现在SAD组中腹侧大脑皮层对杏仁核的影响显著低于HC组,而使用GC分析发现边缘系统(杏仁核组件)和视觉皮层之间的因果效应增加。在先前的研究中(Sladky et al., 2013),SAD患者显示前额皮层和杏仁核之间存在正相关,这表明存在兴奋性连接。基于EC,SAD患者中经常发现异常的杏仁核(Engel et al., 2001; Freitas-Ferrari et al., 2010)。一项基于静息状态FMRI数据的研究(Bae et al., 2017)提出了一个基于GC的网络属性模型,用于研究GAD患者和HCs的EC。因此,报告称GAD个体从皮质区域到杏仁核的EC降低(Qiao et al., 2017)。在先前的研究中(Minkova et al., 2017),招募了15名SAD患者和15名HCs,在情绪场景的认知任务期间估计杏仁核和前额皮质神经元中情感刺激的因果EC。发现在低认知负荷后,SAD个体的杏仁核激活高于HCs。EC可能受到认知任务处理期间认知负荷的影响,导致SAD参与者的EC与HCs不同。最近,在Dong等人(2019)的研究中,35名GAD患者和36名HCs进行了静息状态fMRI,并使用GC分析比较和研究全脑连接性和杏仁核通过EC。结果表明,相对于HCs,GAD个体从左到右杏仁核的GC影响被破坏。这项研究(Makovac et al., 2016)显示,SAD患者的整体连接性改变显著高于HCs。SAD患者表现出杏仁核两侧与右回之间的神经连接相对较低。此外,研究了20名SAD参与者和20名匹配的HCs之间前岛叶和前额叶皮质之间的EC,以探索激活差异。最近揭示,焦虑个体表现出岛叶和前额叶皮质之间的EC显著降低(Kandilarova et al., 2018)。
迄今为止,在不同的大脑成像技术中,fMRI模态在研究SAD的EC方面比其他任何神经成像技术更占主导地位,使用更广泛。EEG在探索神经活动时有几个优点。首先,EEG具有出色的时间分辨率(毫秒或更短),这使系统能够检测和捕捉电活动的任何突然变化。其次,一组EEG电极贴在受试者的头皮上,使EEG技术成为非侵入性的,因为不需要介入性医疗手术。第三,EEG设备相对便宜、操作简单,临床上可用。第四,EEG技术可以量化大脑不同区域之间的EC,而无需生理信息,并阐明不同大脑区域的活动如何相互影响。
结论和建议
总之,SAD是一种普遍的严重智力异常。然而,SAD的诊断仍然顽固地建立在个人评估和传统程序上。在SAD患者和HCs中研究了SAD对社交互动模型中认知行为和振荡动态的影响。EEG信号的功率谱密度分析和大脑连接性估计器是说明SAD神经相关性的鼓舞人心的技术,可能为传统大脑生物标志物提供一种权宜之计。尽管对SAD进行了广泛研究,但还需要进一步研究来验证当前结果并进一步评估预测性社交焦虑生物标志物。这些在创伤情况下观察到的神经生物标志物对探索社交焦虑的遗传基础、改善心理治疗和早期识别可能存在SAD风险的个体非常有效。然而,为这些目的改进的技术是无效的,因为较低的预后精确度或最终结果无法推广,因为样本量小。另一个挑战是倡导先前的经验事实和情境机制,如果纠正这些认知行为可以抑制具有遗传易感性的个体中SAD的扩展,则需要解决这些问题。以下建议强调了本研究的重要性及其对未来研究避免潜在挑战的有用性。
1. 样本量的选择是实验质量的重要决定因素,因为它确定了数据集的重要性并最小化了典型误差。由于样本量小,很难推广一些研究的发现并对最终结论得出明确结论。
2. 本研究讨论的几个生物标志物,如delta-beta相关、FAA(额叶Alpha不对称)、中线theta和大脑连接性,可以组合研究,以验证主要发现并设计有效的诊断方法。
3. 多模态神经成像技术广泛应用于神经科学,以更好地捕捉和可视化神经网络激活。例如,可以应用涉及PET、CT、EEG、fMRI和MEG、EEG-fNIR或fMRI和MEG同时测量的模型。EEG-fMRI整合将导致更高水平的时间分辨率和关于大脑动态过程的更准确数据。


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









