





与老年期(认知和大脑健康下降最为明显,但干预的范围可能有限)相比,中年期在历史上一直是一个缺乏研究的人生阶段。然而,最近的研究表明,中年期可能标志着大脑老化的一个转变。我们回顾了多个分析层面的新兴证据,这些证据表明中年期是一个由独特的中心和外周过程定义的时期,这些过程塑造了未来的认知轨迹和大脑健康。根据人类和动物模型的衰老研究和寿命研究的最新进展,我们强调了在具有广泛受试者年龄范围的研究样本中模拟非线性变化的效用,以区分特定生命阶段的过程和那些在整个生命周期中线性作用的过程。本文发表在Trends in Neurosciences杂志。
要点
-
与更高龄相比,中年(大致相当于人类寿命的第五和第六个十年)在历史上一直缺乏研究,但它正在成为大脑老化的一个可能转折点,这可以预测未来的认知健康,同时仍然可以通过干预来改善。
-
一些与年龄相关的大脑变化在整个成年生活中表现出近似线性的轨迹,而另一些则以特定生命阶段的方式非线性地变化。中年期的特征是步态、反应时间、记忆力、功能连接性、白质完整性和神经炎症,以及基因和蛋白质表达的变化加速。
-
大脑以外的系统和器官(如体循环)的衰老过程会影响大脑衰老,甚至可能加速大脑衰老。
-
对女性而言,更年期是中年的一个决定性特征,它与大脑的非线性变化有关,其中一些变化对认知老化有影响。
关键词:神经影像学 组学 认知 机器学习 神经退行性变 炎症
中年期:大脑老化的可能转折点
尽管公众的理解各不相同,但中年通常是指40岁到60-65岁之间,它不仅仅代表寿命的“中点”,而且是一个被认为向“老年”过渡的时期。从健康科学的角度来看,中年期被认为是预测未来健康状况的一个时期。然而,与老年期(影响可能更大,但干预可能为时已晚或效果降低)相比,中年期的研究仍然不足。就大脑而言,中年期与未来痴呆风险的特定且可改变的风险因素相关(专栏1)。这些是重要的观察结果,因为到2050年,全球60岁以上人口将翻一番,这将导致痴呆症病例数量随之增加,并给患者和护理人员带来巨大的困难。尽管衰老自然与一定程度的认知能力下降有关,但个体间存在相当大的差异。我们重点介绍了在大脑衰老过程中可能与中年期特别相关,并被证明可预测未来认知结果的过程。
| 专栏一 中年生活方式和可改变的(风险)因素 虽然生物衰老可能不可避免,但其几个潜在过程很可能是可改变的,并且可能成为延缓功能衰退的目标。例如,生活方式因素,如吸烟、饮酒和饮食,已被证明与根据神经影像学估计的中年到老年大脑年龄变化有关。此外,2020年柳叶刀委员会关于痴呆症预防的报告指出了一些痴呆症风险因素,包括高血压、肥胖和饮酒,这些因素与中年期尤其相关,并被认为是特定生命阶段的可改变风险因素。由于中年期个体认知变化的纵向轨迹差异很大,并且部分独立于早期生活和先天因素,因此这一时期可以作为干预的切入点。 例如,体育活动对几个身体系统都有有益的影响,这些影响可能延伸到大脑和认知。对运动(尤其是需氧运动)干预的荟萃分析表明,运动可以减缓健康老年人左海马体的年龄相关体积损失,并改善整个成年期(包括中年期)额叶和颞叶束中白质完整性的各种测量指标。鉴于海马体积和联合白质体积从中年开始加速下降,这一点尤其重要。与此相关的是,较高的体育活动水平与中年和老年人情景记忆的纵向下降减少有关。此外,教育程度和社交接触频率对认知的保护作用似乎部分可以通过与更高水平的自我报告体育活动之间的关联来解释。这些发现可能对公共卫生预防痴呆症的努力有用。需要注意的是,一项对24项荟萃分析的综述发现,各种形式的运动对认知的影响(无论年龄范围如何)尚无定论,并且可能受到发表偏倚的影响。此外,有证据表明,38岁时心肺健康与认知功能之间的关系可能可以用儿童期认知得分来解释。这突出了对运动干预对认知老化的纵向影响进行进一步研究的必要性。 最后,当前的老年科学致力于识别驱动机体衰老的常见分子和细胞通路。通过使用药物化合物(例如二甲双胍、senolytics和雷帕霉素)靶向已识别的"衰老标志",正在进行各个阶段的临床研究,以对抗痴呆症和神经退行性变,尽管这些主要针对老年人,并且取得了不同程度的成功。 |
最近的实证研究表明,生物衰老既不是一个跨个体一致的线性过程,也不是在任何给定的实际年龄在整个身体中以相同的速度进行。相反,它被认为包含许多细胞、组织和器官特异性的子过程,这些子过程可以在系统上相互作用,并由在多个生物学水平上运作的内部和外部损伤驱动。组织甚至可能在整个生命周期中经历不同的衰老阶段,每个阶段都由特定的分子特征和时间动态定义,其中一些是非线性的。人生中的一个这样的时期似乎是从中年到老年期的过渡。在人类大脑中,与突触、炎症和长寿相关通路(如哺乳动物雷帕霉素靶蛋白(mTOR))相关的基因和蛋白质的表达被发现在这一过渡期间经历加速或突然的变化,对啮齿动物的研究也报告了中年期类似的非线性。在女性中,其中一些效应可能与更年期的复杂变化有关。鉴于大型神经影像学研究显示中年期结构和功能变化加速,其中一些已被发现可以预测未来的认知衰退,并且可能由体循环中的分子驱动,这些发现尤其有趣。
本文回顾了来自人类和啮齿动物研究的多个分析层面(从行为到分子)的新兴证据,表明中年期可能标志着大脑衰老的转变,这种转变是由与未来认知健康相关的中心和外周过程驱动的。我们认为,将衰老研究的最新趋势应用于这一人生时期,可以揭示新的生物标志物和可能的干预措施,以对抗日益老龄化的全球人口中的认知衰退。我们强调非线性分析方法与广泛受试者年龄范围相结合的效用,以便更好地将特定生命阶段的变化与终生衰老生物标志物区分开来(图1)。
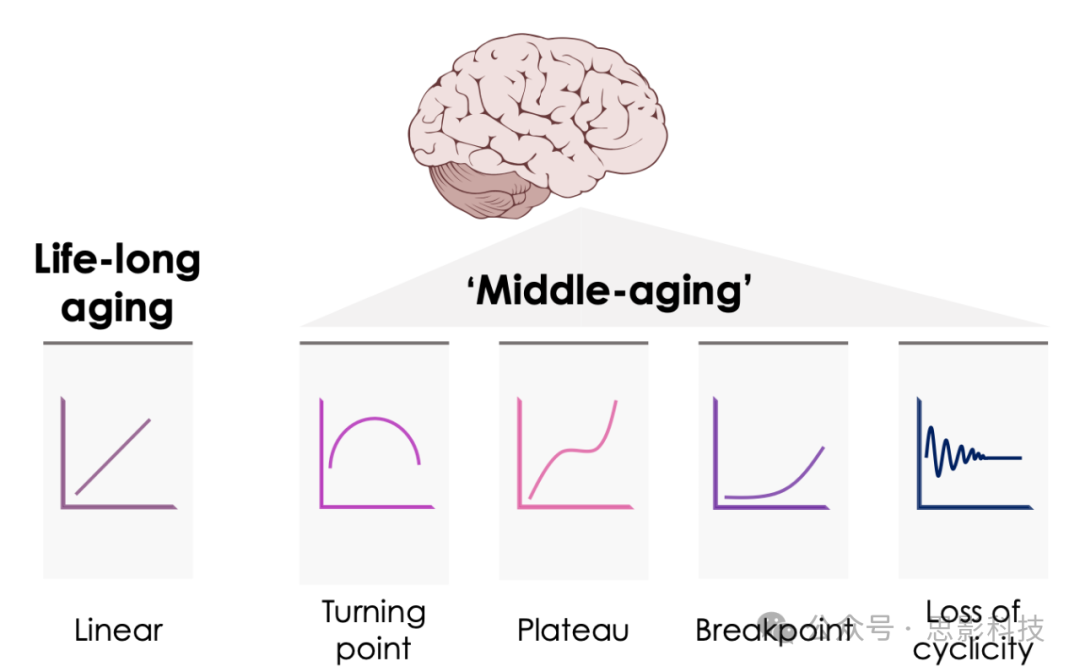
图1 整个生命周期和大脑内部不同衰老动态
大脑衰老包含许多在不同生物学层面运作的过程,这些过程可以作为衰老的生物标志物(例如,基因、蛋白质、代谢物、神经网络)。这些过程可以根据不同的轨迹变化:有些是线性的和恒定的(表明一个逐渐的过程,跟踪实际年龄并随着时间的推移而累积),而另一些是非线性的,并且可以在人生的不同阶段突然变化(这里以转折点、平台期、突然断点和周期性丧失为例)。这些不同的动态可能可以用来区分终生衰老过程和特定生命阶段的衰老过程(这里以中年特有的过程为例)。重要的是,由于不同的大脑系统可以以不同的速度衰老,因此可以说它们在任何给定的实际年龄都有不同的生物学或功能年龄。这可能对干预措施的时间安排有影响,具体取决于它们的目标过程。通过这种方式,中年可以提供一个干预点,以防止大脑加速衰老和认知能力下降。
中年大脑的认知变化
在美国进行时间最长的衰老研究(巴尔的摩衰老纵向研究,始于1958年)得出的一个教训是,生理和神经影像学表型通常显示出与年龄高度不同且非线性的关系。因此,在试图识别在任何特定生命阶段可能特别容易受到影响的认知功能时,我们可能会寻找加速变化的证据(专栏2)。大多数认知领域都表现出从20多岁后期开始的近似线性下降轨迹,但对于某些领域,包括记忆力、推理能力、反应时间指标以及认知负荷对步态的影响,下降似乎会加速,尤其是在中年后期。值得注意的是,在从中年到老年期的过渡中,情景记忆(即对日常事件的记忆)的纵向轨迹似乎特别不稳定,这表明一个人的表现可能会开始突然改变,这可能反映了环境影响的增加。事实上,快速变化的社会关系(例如,父母的丧失和家庭义务)以及职业轨迹的个体差异(例如,退休)是这一人生阶段的显著方面。此外,据报道,处理速度的个体差异在同一过渡期间会放大(引文[41,42];参见[43])。另一个有趣的观察结果是,“练习效应”(即在不同的研究场合重复测试)的好处在60岁左右的情景记忆纵向研究中开始消失。鉴于最近的一项发现,即中年和老年人言语记忆练习效果的下降与认知障碍的严重程度有关,这可能具有临床意义。因此,中年可能是一个未来认知轨迹个体差异开始出现的点,例如与早期认知衰退有关的差异,只要有足够敏感的测量方法。事实上,一项使用大型人群认知老化研究(例如引文[37])中使用的测试组合的研究发现,载脂蛋白E (APOE)基因的ε4等位基因(已知的阿尔茨海默病(AD)风险因素)对中年人而非年轻人的情景记忆有负面影响。因此,一项更大的研究发现,在中年人十年间的纵向言语记忆轨迹中,APOE ε4剂量存在可测量的差异。
| 专栏2 研究中年需要的不只是中年受试者 生物衰老机制,如炎症,具有线性和非线性过程。线性过程跟踪实际年龄,可以说在整个生命周期中都适用,而非线性过程可能在某些生命阶段尤其相关,特别是当进展加速时,这可能表明相反的生物过程的平衡发生了变化。需要注意的是,线性轨迹可以通过阈值模型与表型(例如,疾病发作)耦合,其中一个逐渐的过程在超过某个阈值后累积地产生一个离散的结果,正如衰老细胞负荷与年龄相关疾病发病率之间的联系所提出的那样。因此,对整个生命周期中生物过程的非线性建模可能有助于识别特定生命阶段的变化(而不是终生的变化)。例如,最近一项关于人类皮肤的转录组学研究发现,女性的非线性基因表达轨迹明显多于男性,并且几个基因在50岁左右出现转折点,作者推测这可能与更年期开始有关。 非线性模型的选择对于估计转折点发生的年龄至关重要。例如,与局部平滑算法相比,二次多项式对年龄范围高度敏感。事实上,一项关于人类海马体积的研究发现,提高年龄范围的下限会使拟合断点移动多达三十年,而平滑样条稳健地将其置于47岁左右。此外,包含更大的年龄范围可以抵消与参数化程度更高的样条模型相关的过度拟合风险。相比之下,先前对生命周期神经影像学研究的回顾发现,大多数研究采用多项式模型和55-85岁的参与者。尽管一些非线性模型的效用目前受到样本量的限制,但其他模型(如广义加性混合模型(GAMM))正越来越多地用于稳健地捕捉复杂的生命周期轨迹(例如,引文[27])。 在“组学”应用中,最近出现的差异表达滑动窗口分析(DE-SWAN)允许通过在研究人群年龄范围内以给定分辨率进行连续测试来识别外周蛋白质组和表观基因组的变化时期。这强调了衰老研究中线性和非线性建模的必要性,因此需要两个以上的年龄组,最好是大部分生物体生命周期的代表性样本。 |
情景记忆的一个方面可能对衰老特别敏感,那就是模式分离(或在人类行为评估中称为“记忆辨别”)。海马体的这一功能允许最小干扰存储和回忆高度相似的相关记忆。众所周知,它会随着年龄的增长而下降,对亚临床认知障碍敏感,并且与几种AD生物标志物相关(在[48]中进行了综述)。尽管记忆辨别的下降在整个成年期似乎大致呈线性,但分箱表明,最早可检测到的差异出现在中年早期,并在60岁左右趋于平稳(引文[49,50];参见[51])。一项研究使用超高场磁共振成像(MRI)来评估海马子区的结构和活动如何与整个成年期的记忆辨别相关联。值得注意的是,它发现在齿状回体积、CA3活动(对模式分离最重要的两个区域)和记忆辨别之间的关系中,在40岁到60岁之间存在非线性转折点。这些发现被认为反映了一个可能的过渡点,在此期间,与未来认知衰退相关的神经变化开始出现。根据最近对小鼠的研究,我们很容易推测,这可能部分是由于成年海马神经发生的减少(这被认为对模式分离至关重要),据报道,这种神经发生在人类成年期逐渐下降。然而,重要的是要注意,与啮齿动物相比,人类成年海马神经生成的存在和可能的功能相关性是一个正在进行的辩论的问题,并且与模式分离的确切机制联系(主要通过啮齿动物研究理解)仍有待完全阐明(引文[60]讨论了流行的理论)。
中年记忆力下降也可能延伸到模式生物。一项涉及数千只小鼠和大量行为任务的研究发现,海马体依赖性记忆下降的某些方面(例如,空间记忆和上下文恐惧条件反射)仅在8-12个月大的组中出现(10-14个月对应于生殖衰老,是人类中年的鼠类类似物)。
综上所述,一些认知领域,尤其是情景记忆,似乎在中年期经历加速变化,这可能反映了新出现的认知衰退的个体轨迹,这可能是由大脑的非线性变化所支撑的。
中年大脑的结构和功能变化
大脑的结构在整个成年期持续演变,其特征是线性和非线性轨迹,由转折点划分,这些转折点可能反映了退行性和发育过程之间平衡的转变。海马体和邻近的海马旁回是两个这样的区域,在人类五六十岁时会出现体积转折点,一些海马体子区域萎缩得比其他区域更快更早。重要的是,这种加速下降及其个体差异程度与整体认知状态以及从中年到老年过渡期间的纵向情景记忆下降有关。在更大范围内,脑白质和脑室体积分别在大约六十岁时表现出非线性下降和增长,这些结构的个体差异与轻度认知障碍和阿尔茨海默病病例对照状态有不同的关联。事实上,大多数神经束(尤其是联络纤维)在整个生命周期中都呈现倒U型曲线关系,转折点在40至50岁之间。这些发育动态可能是反应时间在整个生命周期中镜像趋势的基础,反应时间在六十岁时呈指数级减慢。额顶叶长程和短程纤维的传输速度在30至40岁之间达到最小传导延迟,后者成熟较慢,这支持了这一观点。此外,一项小型研究在健康的50岁和40岁成年人中建立了跨领域认知衰退与扣带束和右侧额枕下束恶化之间的初步关联(尽管受认知储备能力的影响)。这与这些纤维在五十岁时的结构转折点相吻合。
与大多数白质束的生命周期轨迹一致,一些网络(例如额顶网络和默认模式网络)的结构和功能连接性表现出负二次趋势,下降始于40岁左右。其他网络(例如腹侧注意网络)的正二次趋势与此平行,这被认为反映了代偿机制。功能连接性(即用功能性磁共振成像评估的相关神经活动)的个体差异也反映了注意网络和额顶网络分别呈现相反的正负二次关系趋势,转折点大约在六十岁。这种变异性与流体智力和外显运动行为有关。一般来说,功能连接的平均强度和数量似乎在50岁左右达到峰值,然后下降。这种老化期间的下降尤其与长程功能连接的丧失平行。功能连接的平均解剖距离已被证明在大约相同年龄达到峰值,这支持了这一观点。大脑区域之间的连接性使它们能够形成模块,这些模块共同构成更大的功能分离系统(例如,注意网络和默认模式网络)。这是通过系统内更密集的连接和系统间更稀疏的连接之间的平衡来实现的。随着年龄的增长,系统之间的相关性总体上有所增加,导致系统分离度降低,并且可能导致功能特化丧失(这被称为“去分化”)。
值得注意的是,高级联络系统(即额顶叶、注意、默认模式)之间的分离丧失似乎是非线性的,断点在50岁左右(与感觉运动系统的终身线性趋势相比),这种丧失独立于年龄预测情景记忆下降,并且个体差异显著。最近的一项纵向研究重复了这种联络系统和感觉运动系统中功能分离的非线性丧失与线性丧失的对比,尽管它将非线性断点置于稍晚的58岁。类似于社会中高度“连接”的个体相互形成网络的现象(称为“富人俱乐部”),大脑在其最密集的枢纽区域之间表现出功能互连性,这种组织似乎在中年达到顶峰。事实上,最近一项对144项静息态功能成像研究的系统综述提出,四五十岁可能是大脑网络组织的转折点,其特征是最佳效率、系统分离和模块化,随后这些特性加速衰减。有趣的是,一种针对认知测试领域的连接组方法(即使用类似的图论指标)发现,从五十岁到六十岁的过渡期间,效率和模块化也出现了类似的损失。
综上所述,有充分的证据表明,人脑在中年期经历非线性的结构和功能变化,这对认知功能有影响,而这些过程的变化可以解释认知老化的个体轨迹。最终,这些变化可能源于最基本生物学水平的复杂动态。
细胞和分子在中年大脑中的变化
在过去十年中,使用大规模生物特征检测(如蛋白质、代谢物、(非)编码RNA和DNA修饰,统称为“组学”)对机体衰老的研究越来越多(引文[73]综述)。一个基础示例包括通过机器学习算法构建所谓的生物“时钟”,这些算法根据此类生物特征预测实际年龄,最显著的是使用血液样本中的DNA甲基化水平。尽管使用的模型通常是线性的,但时钟在不同器官中的“滴答”速度可能不同,并且在整个生命周期中遵循不同的“滴答”功能(在数学意义上),其中一些是非线性的。一个原本线性的时钟可能包含以非线性方式跟踪实际年龄的子特征,有时以断点或平台的方式。事实上,基于人脑组织的表观遗传时钟通过考虑非线性关系得到了改进。重要的是,表观遗传年龄也被发现可以预测中年期的纵向认知功能。因此,正如时钟所捕捉到的,生物衰老是具有不同动态的复杂过程的混合(并非所有过程都会立即造成损害),这开启了识别那些与特定生命周期相关的过程的可能性。
与此相关的是,分子过程(即使是那些属于相同衰老特征的过程)在整个生命周期中都表现出线性和非线性轨迹。例如,对人类或啮齿动物组织进行的转录组学研究(采用广泛的受试者年龄)已经发现了支持这一观点的发现。在人类皮层和海马体中,有一些证据表明基因间的共表达在第六和第七个十年之间经历了指数级变化。这涉及免疫相关基因表达的增加和一些突触基因表达的减少。特别是,当评估不同数据集和大脑区域中个体间基因表达的异质性时,中年期似乎出现了非线性变化,与轴突引导和mTOR信号传导(一个与长寿始终相关的、高度保守的通路)相关的基因在60岁左右出现指数级增长。一项对人类海马体的研究也类似地发现,在从第六个十年到第七个十年的过渡期间,神经传递和免疫相关基因的表达异质性呈指数级增长。人们很容易推测,这种差异表明中年期个体衰老轨迹的差异,有些人比其他人衰老得更快。尽管基因表达变异性的原因仍有待完全解释,但在60岁之后(最早在40岁时也被检测到)快速增加的方差已通过不同的方法和技术得到重复验证。有趣的是,一项分析多个数据集的研究发现,由mTOR和线粒体通路组成的基因表达谱与人脑、肌肉和皮肤的年龄呈线性相关(主要是负相关),但这种关联在大约60岁时突然消失。这可能是从中年到老年过渡期间这两个通路调节中非线性断点的证据,正如对表皮组织中mTOR的建议。
与前面提到的中年期脑室体积的指数级增长相关,一项对认知健康参与者的脉络丛(脑脊液的主要来源)的转录组学研究发现,最大的基因表达变化发生在第六个十年。这些变化独特地富集了细胞屏障相关通路(例如,间隙连接相关基因的下调),而轴突和神经元投射通路则富集了那些持续变化(主要增加)到生命第九个十年的基因。尽管未纳入50岁以下的参与者,这增加了效应可能更早开始的可能性,但鉴于最近发现克隆性T细胞募集到脑脊液与六至九十年生命周期中健康和非健康衰老期间的神经轴突分解有关。有趣的是,这项研究发现,年龄在50-80岁左右的认知障碍参与者的T细胞受体库与68岁以上健康参与者的T细胞受体库更相似,而不是与54-67岁的健康参与者更相似,这意味着加速生物衰老是认知障碍的一个特征。与此相关的是,老年期的认知健康被认为反映了中年期海马结构的转录衰老轨迹。一项对人类海马体和枕叶皮质突触体的小型蛋白质组学研究(即对总蛋白质群的分析)显示,一些蛋白质在40-50岁和70岁以上之间表现出断点样差异丰度,并且这些蛋白质富集于转化生长因子β (TGF-β) 通路。特别是在海马体中,该通路被突然上调,作者推测这可能代表通过促炎性小胶质细胞信号传导的年龄相关区域突触脆弱性机制。至关重要的是,在大型独立队列中,背外侧前额叶皮质中的线粒体、突触和炎症蛋白表达(包括小胶质细胞和TGF-β通路)被发现与从中年晚期到老年期的个体纵向认知轨迹相关,这些发现与来自老年AD病例对照比较中同一脑区的基因表达数据一致。
支持中年期是跨物种加速变化时期的观点,最近一项对小鼠器官中非编码RNA的转录组学研究发现,在12个月和18个月时,大脑中最强的影响是以miRNA共表达转折点的形式出现的。一个在大脑中具有这种轨迹的非编码转录本(miR-29c-3p)被发现对通过异时异种共生(即通过手术连接年轻和年老动物的循环系统)的全身再生敏感,并与免疫衰老有关,可能通过TGF-β信号传导。对编码RNA的类似研究发现,在小鼠大脑中年期,神经元和突触(平台期)、免疫(断点增加)和蛋白质折叠(断点减少)通路中存在非线性轨迹。虽然这些效应可能不一定转化为蛋白质水平的变化,但有趣的是,小鼠海马体中的突触和蛋白稳态通路被发现在12个月时在蛋白质水平上失调(可能是由于昼夜节律表达模式的丧失(参见[89]关于人类海马体中保留周期性的证据)。
总之,大脑似乎在中年期经历了与mTOR、线粒体、突触和炎症相关通路的非线性变化,基因表达异质性的加速可能标志着人类生命第七个十年的开始。有趣的是,并非所有变化都源于大脑本身,也可能由外周和其他器官驱动。
大脑"中年"的可能外周调节因素
体循环中的因素(主要是各种器官分泌的蛋白质)被认为在认知、可塑性、成年神经发生和神经炎症方面介导大脑衰老(最近在[90]中进行了综述)。因此,通过改变这种外周特征(例如,间接通过运动,或直接通过异时异种共生),可以加快或减缓大脑细胞时钟上的生物学时间。鉴于此,特别有趣的是,最近的表观基因组学(即对表观遗传修饰和染色质结构的大规模表征)和蛋白质组学研究发现,在整个成年生命周期中,一些最大的变化峰值发生在中年期,如血浆、外周血单核细胞和肌肉中所测量的。这些峰值中有许多涉及炎症和免疫相关通路。此外,一些在中年期出现转折点的血浆蛋白共表达轨迹富集了神经系统相关通路,在肌肉和单核细胞蛋白质组和甲基化组中也发现了类似的结果。这表明,中年期体循环的状态可能可以预测个体认知衰老轨迹。事实上,一项大型纵向研究表明,在中年期基线测量的四种炎症标志物的综合水平预测了长达20年后的认知衰退斜率,这种效应对于言语情景记忆尤其可靠。在同一队列的后续研究中,对中年期采集的样本进行血浆蛋白质组学分析,发现了与25年后发生的痴呆症、5年后的神经影像学表型(例如,白质高信号)和脑脊液炎症负荷相关的突触、蛋白稳态以及免疫和小胶质细胞相关蛋白质。有趣的是,中年期的蛋白共表达模块与近期和长期痴呆症发作的关联不同,并且富集了不同的生物学通路和组织。中年期识别的痴呆症相关蛋白质很少与其他老年队列中识别的蛋白质重叠,这表明认知衰老的外周特征可能以特定生命阶段的方式进化。
一项对英国出生的晚期中年人(60-64岁)的类似队列研究建立了言语记忆/处理速度的横断面和纵向测量与血浆代谢物(主要是脂肪酸和脂质)之间的关联,这些主要由童年认知和教育来解释。对相同代谢组学数据的基于网络的分析显示,涉及酰基肉碱和鞘脂的共表达模块(即相关的代谢物子集)分别与5-9年的纵向记忆和处理速度以及横断面海马体积相关,并且对早期生活因素具有稳健性,而其他关联主要由童年因素解释。外周代谢物水平的一些变化可能是由于中年期人类肠道微生物群组成(即居住在胃肠道中的数万亿微生物)的非线性变化,这可能会影响神经炎症和认知,正如对啮齿动物的研究表明的那样。因此,中年认知轨迹的预测因素似乎既来自持续(加速)的衰老过程,也来自早期生活的决定因素,这些可能通过纵向和横断面研究设计来区分(专栏3)。
| 专栏3 横断面研究和纵向研究定义的中年期可能不同 多年来,认知能力随年龄增长自然下降的轨迹一直存在争议,因为横断面研究设计估计下降始于三十岁,而纵向研究设计则认为下降始于七十岁。最终,这种差异可以用纵向重复测试效应(已知会持续数年)、队列效应以及两种设计从根本上捕获不同变异来源来解释。因此,设计的选择可能会使年龄相关关联的估计产生偏差。最近,在生命周期神经影像学研究中也出现了类似的讨论,其中横断面研究设计在历史上和最近都是常态。横断面研究对结构变化的估计似乎大大低估了实际的纵向下降轨迹,而这些差异部分归因于技术混淆和参与者的生活方式因素。此外,大脑年龄的横断面和纵向估计(即使用机器学习模型从神经影像学表型预测的实际年龄)并不相关(独立于统计模型和研究人群),并且似乎反映了不同的影响,前者(而非后者)尤其与早期生活和遗传因素相关。与心理学中的发现相似,年龄差异并不等同于年龄相关变化引起的差异,这强调了对人类(例如,引文[36])和模型生物(例如,引文[142])进行纵向研究以阐明大脑衰老的因果驱动因素的必要性。这种对由持续衰老和先天或早期生活因素引起的生物学年龄的解析,让人想起Leonard Hayflick对衰老(认为是由于熵增引起)和寿命(认为由遗传因素决定)的区分。然而,纵向生物学年龄指标也捕捉到了早期生活和遗传影响,但至关重要的是,可以在成年期持续变化的估计中对此进行调整。 重要的是,由于生物学年龄估计(或“衰老时钟”)本质上是根据生物学特征(例如,DNA甲基化、神经影像学表型、蛋白质水平)预测实际年龄的统计模型,因此上述注意事项将独立于所测量的特征而适用。然而,迄今为止,大多数衰老时钟都是基于横断面数据(参见[9]中关于具有非线性轨迹的纵向示例),尤其是那些依赖组学技术的数据(最近在[73]中进行了综述)。为了充分阐明中年期的生物学过程如何塑造未来的认知轨迹和大脑健康,纵向研究将非常重要。 |
鉴于这些发现,有趣的是,外周器官的生物学年龄可能会影响大脑的生物学年龄。最近一项主要针对中年人的英国生物银行队列研究构建了大脑(使用神经影像学衍生表型)、其他七个器官(基于器官特异性临床生物标志物)和整体认知的线性年龄时钟(即根据生物学特征预测实际年龄的机器学习模型)。值得注意的是,较大的大脑年龄差距(即个体预测年龄与实际年龄之间的差异,可作为生物学年龄加速的指标)在横断面研究中与较大的器官年龄差距相关,尤其是在心血管、肺和代谢系统中。预测的大脑和身体年龄越大,也预示着认知年龄越大,并且发现外周器官会加速其他器官的纵向年龄差距。最后,另一组采用类似方法的研究发现,基于45岁时19种临床身体生物标志物的个体衰老速度与白质高信号负荷、海马体积和处理速度降低以及记忆力和感觉运动功能较差相关,即使在调整了二十年前的儿童智商(IQ)和基线健康状况后也是如此。
总而言之,中年期似乎是大脑外发生显著变化的时期,这可能通过循环中的因素(尤其是与炎症通路相关的因素)加速其衰老,并最终塑造个体的认知轨迹。
更年期——女性中年的一个决定性特征
在我们迄今为止的讨论中,尚未提及的一个中年重要方面是生殖衰老。在人类女性中,这自然发生在50岁左右,以更年期的形式出现,最近被定义为卵巢功能、生殖激素产生和生育能力的自然(或手术诱导的)永久丧失。根据女性血液样本的测量结果,更年期已被发现会加速表观遗传衰老。事实上,其发病年龄的遗传结构主要涉及DNA损伤反应,这可能通过表观基因组信息丢失来驱动身体、大脑和认知的衰老。生殖细胞完整性对体细胞衰老的因果影响似乎在整个生命中都是保守的,并且涉及前面几节中提到的许多通路,例如(m)TOR、TGF-β和蛋白稳态[106]。
在人类中,更年期通常伴有主观认知变化,尽管个体间差异很大。围绝经期到绝经后过渡期可能与多个认知领域的非线性下降有关,尤其是与言语情景记忆受损有关。这种过渡——与年龄无关——伴随着海马体积加速损失、脑葡萄糖代谢失调和白质高信号负荷。更年期年龄越早,大脑衰老的速度就越快,这是通过神经影像学衍生表型(如总灰质体积和功能连接)评估的。这可能是由复杂的激素动态驱动的。有趣的是,最近一项针对年轻成年女性的纵向研究表明,月经周期中的卵巢激素波动与海马亚区体积的非线性波动有关。一项针对雌性大鼠的研究支持了女性生殖衰老在驱动复杂大脑变化中的作用,该研究发现,周期性雌激素的丧失伴随着海马中TGF-β、干扰素和脂质代谢通路的差异调节(其中一些也出现在中年女性与年轻成年女性的比较中)以及白质可能受到免疫浸润。这表明更年期过渡期可能是几个大脑衰老过程中的一个转折点。然而,重要的是要注意,更年期的更传统定义,例如月经周期停止(即最后一次月经的时间),并不适用于大多数物种(包括啮齿动物),因此常见的模型生物无法捕捉到这一点。
虽然对更年期对大脑影响的深入回顾超出了本文的范围,但我们注意到,这种过渡可能是上述许多研究中非线性变化的来源,其本身值得认真考虑。
结语和未来展望
人类在五六十岁时,大脑功能和结构连接、情景记忆以及大脑和全身循环中基因和蛋白质表达的非线性变化轨迹表明,中年是塑造未来大脑和认知健康的独特过程的时期(图2)。这些发现主要来自对人类或模式生物的研究,涵盖了大部分成年期。这凸显了在研究与年龄呈复杂关系的生物学过程时,纳入广泛受试者年龄范围的重要性,因为简单的两组比较(例如,年轻人与老年人)无法充分捕捉这些过程。中年时期功能性脑网络和白质完整性的变化就是一个很好的例子。此外,越来越清楚的是,纵向和横断面研究有时会捕捉到不同的年龄相关变异指标。在本综述讨论的几项研究中,横断面研究对年龄效应的估计通常反映的是受试者早期的差异,而不是衰老引起的持续变化。因此,为了捕捉特定生命阶段的衰老过程,该领域将受益于利用更多纵向设计研究的结果。
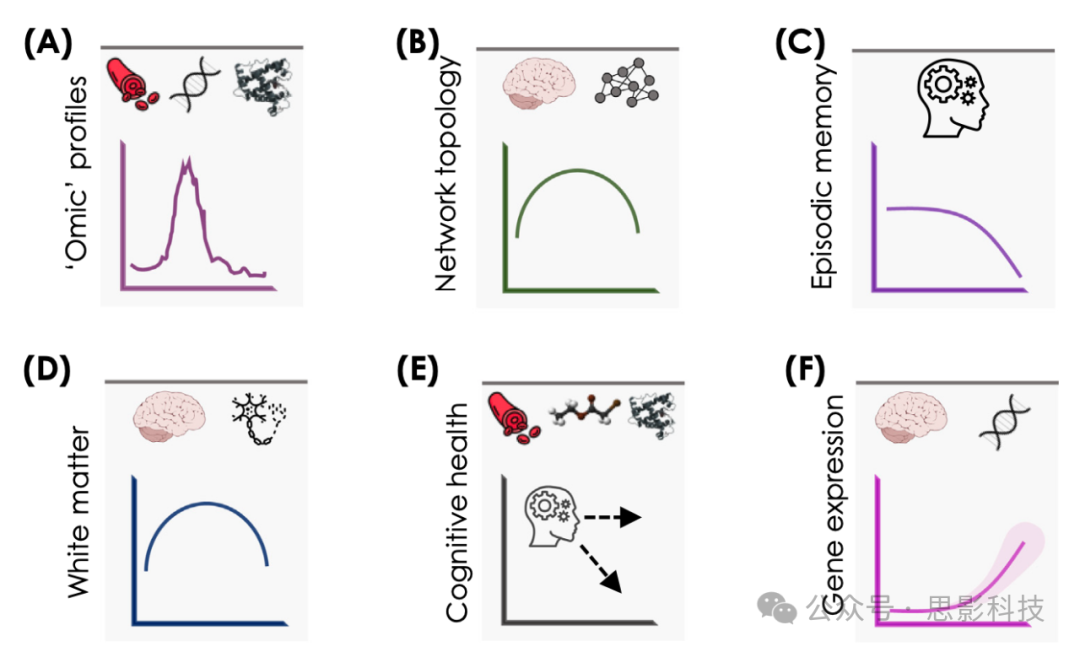
图2 中年时期认知、大脑和外周的变化
来自多层次分析的证据表明,中年期与可能塑造未来认知老化轨迹的独特过程相关。
(A) 一些外周组织和细胞类型(例如,肌肉和血液单核细胞)的血浆蛋白质组(包括炎症通路)和甲基化组经历了差异调节的波动,尤其是在生命的五六十岁之初。
(B) 功能性大脑模块(主要涉及联想网络)的组织达到峰值,然后开始恶化。
(C) 情景记忆在从中年到老年过渡期间表现出加速下降,并且可能存在不稳定的个体纵向轨迹(例如,[37,38,41])。
(D) 结构(可能还有功能)连接,尤其是联络纤维,达到峰值,然后开始下降(例如,[64,67])。
(E) 中年时期循环中的蛋白质和代谢物可以预测纵向认知健康轨迹,有时甚至可以预测二十年后的情况(例如,[32,35,97])。
(F) 已经观察到大脑中基因表达和异质性的突然变化(涉及炎症、突触和长寿相关通路),尤其是在中年后期(例如,[18-20,79])。
虽然我们已经讨论了与认知老化相关的发现,但重要的是要注意,某些关联本质上是复杂的,可能代表代偿反应,而不是驱动老化本身的过程。事实上,某些血浆蛋白候选认知下降标志物就是这种情况。最终,科学家们将希望找到新的治疗靶点来减轻不健康的认知老化。由于针对最终证明是代偿性的年龄相关过程可能会造成有害影响,因此识别哪些“中年期”过程会导致大脑健康下降非常重要(参见未解决的问题)。另一个复杂性来自于观察到某些生物老化过程似乎是部分可逆的,例如人类和啮齿动物暴露于不同压力源导致的表观遗传年龄加速,但在逆转加速的能力方面存在显著的个体差异。如果这种变异性扩展到大脑的再生,那么在早期预测对干预的反应可能在临床上有用。
目前仍然缺乏针对不健康认知老化的干预措施。体育锻炼通常被认为是一种经济有效且易于操作的替代方案。与本文的讨论相关的是,运动已被证明会导致许多不同的因子(例如,蛋白质和代谢物)从身体器官分泌到全身循环中。其中一些因子对衰老大脑有益,正如在小鼠身上看到的那样。有趣的是,在年轻人[和中年人中,都发现大脑可塑性相关通路在对急性运动的血浆蛋白质组反应中富集,这表明结果可能扩展到人类。因此,研究衰老和运动相关分子过程的交叉点可以揭示新的治疗靶点。
最后,重要的是要注意,与衰老相关的过程中存在显著的性别差异,例如痴呆症的患病率差异(在[120]中进行了综述)。分子衰老轨迹也被发现存在差异。例如,大脑基因表达周期性的维持以及中年时期基因表达轨迹的非线性程度可能存在性别二态性。这些差异需要在人体研究中纳入不同性别的参与者并分析年龄-性别交互作用,以及使用动物模型对衰老中性别差异进行机制研究。
人类的五六十岁正在成为不仅仅是成年和老年之间的插曲。相反,多层次生物学的非线性轨迹表明,大脑可能正在经历一个“中年期”阶段。最终,这些变化可能会塑造未来的认知老化。因此,应该重新考虑这个以前未被充分研究的生命时期。
未解决的问题:
1,中年期能否以生物学信息的方式(重新)定义?
2, 鉴于在整个生命周期中变化并影响未来健康结果的众多过程,哪些过程在中年时期具有独特的预测性?
3,由于任何器官的整体衰老速度都包含许多子过程,大脑是否存在一个或多个(生物学)中年期?如果存在,它们能否被差异化地靶向?
4, 哪些干预措施特别适合在中年实施以减轻未来的认知衰退?
5,中年时期哪些与大脑相关的变化与认知衰退在机制上相关联?哪些(如果有的话)是中性的、适应性的或代表大脑或认知储备的变化?
6,中年时期哪些大脑和认知变化是由全身驱动因素介导的?其中一些能否作为干预的分子靶点?
7,在横断面设计中,一些分子过程在中年时期呈现非线性趋势,但哪些是由于加速的纵向衰老造成的?哪些是群体效应(例如,队列效应)或早期生命和先天因素造成的?
8,一些人的大脑衰老轨迹是否比其他人更容易干预或逆转?如果是,为什么?
9,当前的衰老科学致力于识别支撑生物衰老的共同且可靶向的通路和机制,并且人们认为这些机制与普遍的年龄相关疾病的机制重叠,这表明健康衰老和衰老疾病处于一个连续体中。如果是这种情况,那么本文讨论的正常、非线性(“中年期”)过程与通常在中年出现的表现为神经退行性过程的病理过程(例如,β-淀粉样蛋白斑块负荷)有何关系?
10,外周组织已被发现以性别依赖的方式呈现线性和非线性基因表达轨迹,尤其是在中年时期。是否存在类似的性别依赖动态的大脑衰老过程?


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









