





“神经液体”一词被引入用以定义大脑和脊髓中的所有液体,如血液、脑脊液和间质液。在过去的千年中,神经科学家们稳步识别出大脑和脊髓中几种不同的液体环境,这些液体环境以同步和谐的方式相互作用,以确保所需的健康微环境,从而实现最佳的神经胶质功能。神经解剖学家和生物化学家提供了大量证据,揭示了血管周围空间、脑膜和胶质的解剖结构及其在神经元废物排泄中的作用。由于非侵入性成像方式的有限可用性,能够提供大脑神经液体高时空分辨率的描述,人类研究受到了限制。因此,动物研究在推进我们对液体时间和空间动态的了解方面起到了关键作用,例如通过注射具有不同分子量的示踪剂。这类研究激发了识别人类疾病中神经液体动力学可能中断的兴趣,如小血管疾病、脑淀粉样血管病和痴呆症。然而,在将这些发现外推以理解人类大脑时,应考虑啮齿动物与人类生理学之间的关键差异。越来越多的非侵入性MRI技术被开发出来,以识别改变的排水路径的标志物。在2022年9月于罗马举办的国际医学磁共振学会组织的为期三天的研讨会上,几位杰出的国际专家讨论了这些概念,为已知内容和仍缺乏证据的领域奠定了基础。我们设想,在下一个十年中,MRI将能够成像人类大脑中神经液体动力学和排水路径的生理学,以识别疾病背后的真实病理过程,并发现用于早期诊断和治疗的新途径,包括药物递送。本文发表在Journal of Magnetic Resonance Imaging杂志。
引言
“神经液体”一词首次出现在2019年的文献中,作为一个总括性术语,用以定义大脑和脊髓中的所有液体;包括动脉血、静脉血、脑脊液(CSF)和间质液(ISF)。身体中液体的重要性在公元二世纪由伯尔伽门的盖伦描述。他描述了维持生命的四种液体或体液,并假设这些液体的紊乱会导致疾病。然而,直到13世纪,解剖剖析被合法允许,才揭示了更为复杂的大脑和脊髓的宏观解剖学。在随后的几个世纪中,随着伽利略显微镜的发展,神经解剖学家如蒙迪诺·德伊·卢齐、马切洛·马尔皮吉、安东尼·范·列文虎克、尼尔斯·斯藤森、弗雷德里克·鲁伊斯奇和多梅尼科·科图尼等人描述了更精细的显微解剖学。马尔皮吉提出灰质(GM)由微小的腺体组成,白质(WM)由管道组成。17世纪中叶,朱塞佩·赞贝卡里假设一种与血液分离的液体进入大脑以允许神经活动发生。这种液体被称为动物精神或“sugo de nervi”(神经汁)。18世纪中叶,多梅尼科·科图尼鉴定出填充脑室的脑脊液。在同一时期,亚历山大·蒙罗和乔治·凯利提出了著名的学说,即颅骨的固定体积决定了血液的进出流量相等,然而他们完全忽视了脑脊液的贡献。随后,神经外科医生如哈维·卡辛和神经学家如朱利奥·比佐泽罗,以及解剖学家如卡米洛·高尔基,对于定义血液(动脉和静脉)与脑脊液流动之间的相互关系起到了关键作用。20世纪中叶,海伦·切尔率先研究了间质液的动力学,为现代神经液体的研究奠定了基础。
自1980年代初第一批临床扫描仪开发以来,已经开发出几种基于磁共振成像(MRI)的技术来成像神经液体(见表1)。这些方法包括使用外源性(例如基于钆的成像技术,GBCA)和内源性(例如动脉自旋标记方法)示踪剂的方法。脑脊液流动动力学的研究主要基于相位对比技术,包括较新的4D流动方法。最近,扩散张量成像(DTI)、功能性MRI(fMRI)和其他技术被应用于评估微结构和神经血管振荡以及不同液体隔间的相互作用,导致学术产出显著增加。
表1. 成像神经液体的不同MRI技术
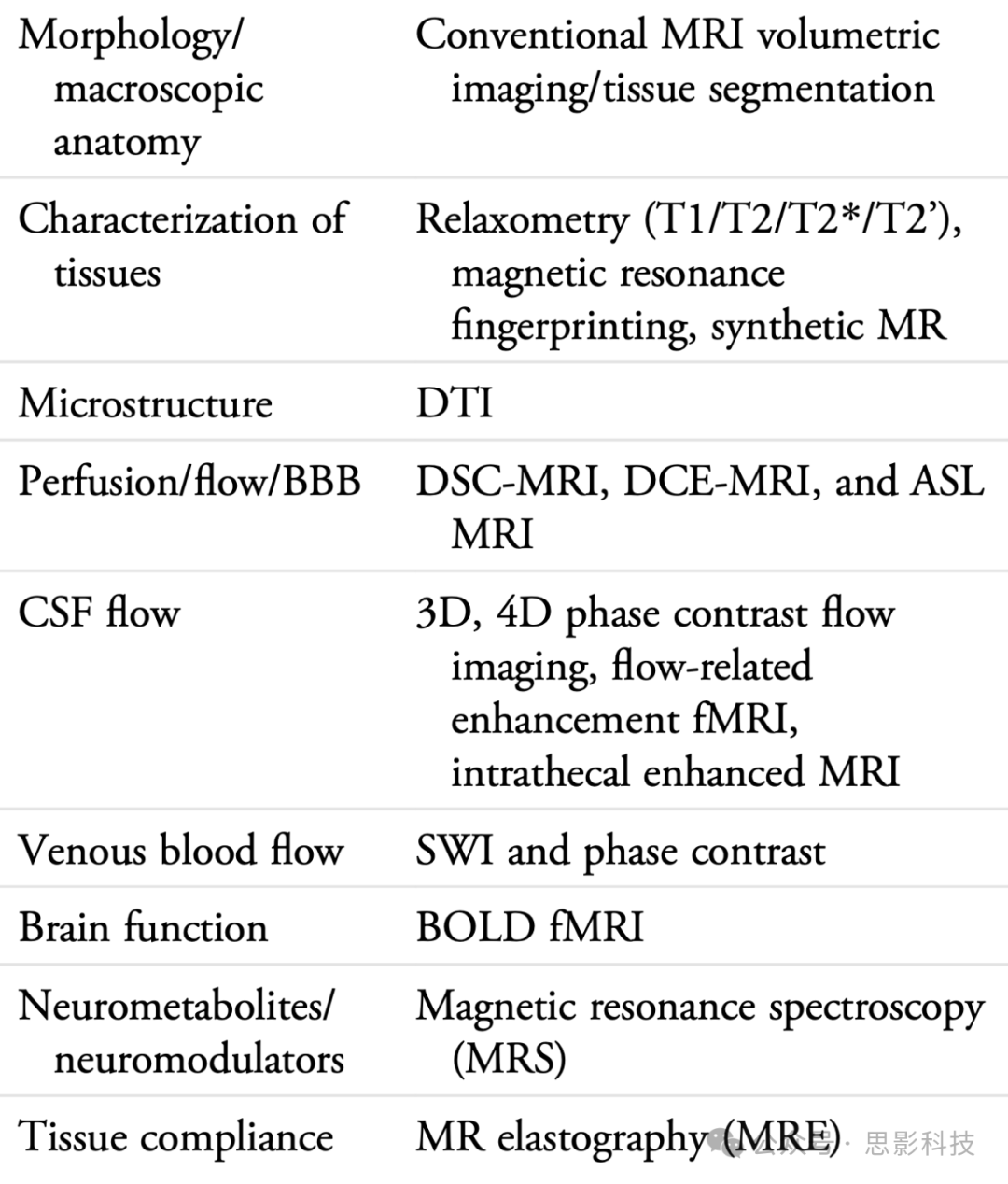
缩略语:DTI = 扩散张量成像;MRF = 磁共振指纹识别;DSC = 动态敏感性对比;DCE = 动态对比增强;ASL = 动脉自旋标记;SWI = 敏感性加权成像;fMRI = 功能性磁共振成像;MRS = 磁共振波谱。
在国际医学磁共振学会内部,2019年成立了一个新的研究小组“神经液体成像”,吸引了全球众多磁共振科学家的兴趣。第一次研讨会于2022年9月举行,旨在汇集关键科学家和临床医生,系统地阐明已知内容和尚待揭示的领域。本综述文章反映了此次研讨会的成果。毫无疑问,更好地理解神经液体的动态具有极大的潜力,可以显著增强我们对大脑废物清除生理路径的理解,了解其在神经疾病中的作用,并开发用于诊断和治疗的新途径(见图1)。
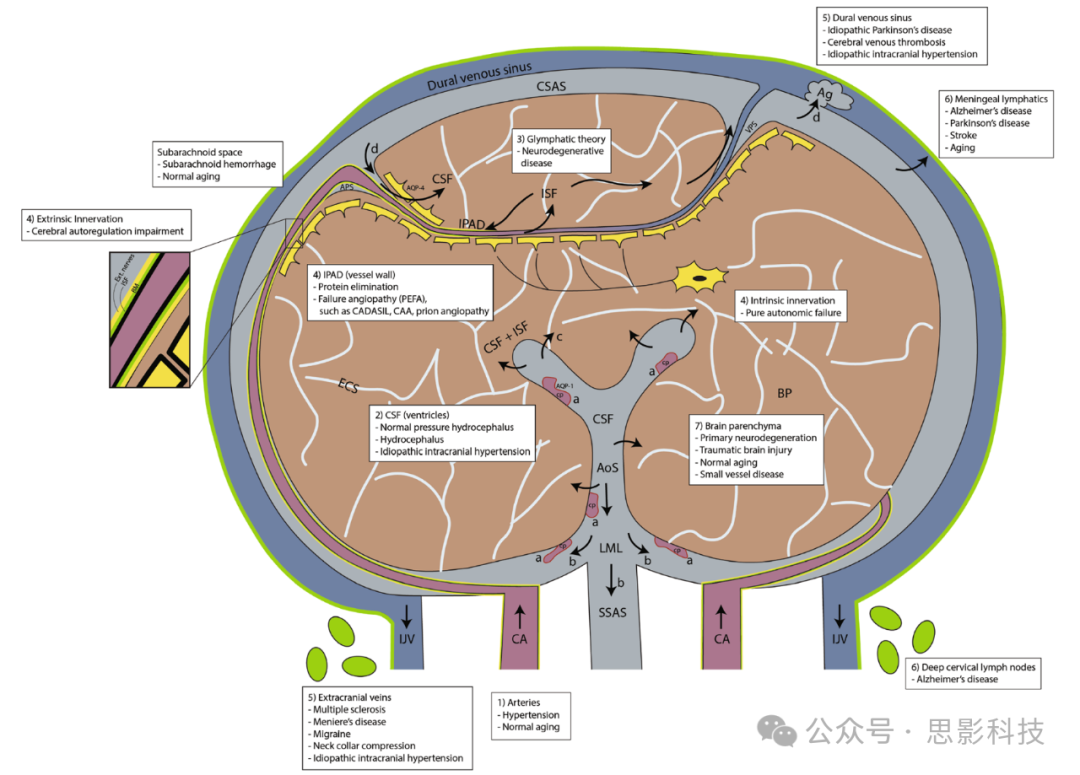
图1.大脑冠状切面的示意图,展示神经液体隔室与脑实质(BP)之间的相互作用,以及由于其紊乱可能导致的神经疾病(白色文字插入):脑膜下动脉和穿透性小动脉从颈动脉(CA)分支出来,并在大脑蛛网膜下腔(CSAS)皮质表面形成密集的网络。脑脊液(CSF)(a) 从脉络丛(CP)产生,(b) 向尾侧流入西尔维乌斯导水管(AoS),通过卢施卡孔和马根迪孔(LML)进入CSAS并流向脊髓蛛网膜下腔(SSAS),(c) 可能通过脑室上皮壁流向BP(脑实质),(d) CSF通过动脉(Ag)排入硬脑膜静脉窦。
CSF可以通过动脉周围空间(APS)和AQP-4水通道重新循环进入BP(脑实质)。总体流动和/或扩散过程将间质液(ISF)/CSF从BP(脑实质)移动到静脉周围空间(VPS),将代谢废物运输到CSAS(大脑蛛网膜下腔)并进入硬脑膜静脉窦。IPAD(壁内动脉周围引流通路)路径是ISF沿动脉基底膜(BM)逆向动脉流动的过程(放大框),并沿CA壁离开颅内腔进入深颈淋巴结。动脉和毛细血管壁分别有外在和内在神经支配,是脑自我调节的决定因素。静脉系统位于CSAS(大脑蛛网膜下腔)表面。皮质静脉排入浅静脉窦系统(硬脑膜静脉窦)。静脉血通过颈内静脉(IJV)和椎静脉(未显示)离开颅内腔,流向心脏。脑膜淋巴管是排列在硬脑膜静脉窦、颅神经鞘、筛板和基底静脉丛的通道,通过神经鞘排出CSF。BP(脑实质)由80%–85%的细胞和ISF组成。
CADASIL = 脑部常染色体显性遗传性动脉性小血管病伴皮质下梗死和白质脑病。
CAA = 脑淀粉样血管病。
神经液体基础
脑血流动力学
大脑在很大程度上依赖于良好调节的血液供应,以匹配神经元和胶质细胞不断变化的代谢需求。脑血管的结构和功能特性旨在确保其获得足够的血液供应,并保持其微环境的稳态。结构上,主要的脑血管在大脑底部汇合于Willis环,这允许在动脉阻塞时通过替代路径进行补偿性血流,而脑表面的脑膜微循环形成一个密集的吻合网络,允许血液通过多个路径到达大脑深部。功能上,神经血管系统:(a) 在一定压力范围内防止血压下降时潜在有害的血流减少,(b) 将氧气和葡萄糖的供应与活跃大脑的能量需求相匹配,使神经活动增加时相应地增加活跃区域的血流,(c) 有助于清除大脑产生的代谢废物,(d) 通过跨胞吞和分子转运蛋白调节血液与大脑之间的双向分子交换, (e) 参与免疫细胞进出大脑的运输,以及 (f) 生成关键的生长因子以维持神经元和胶质细胞的活力。在脑疾病中,神经血管系统的这些关键特性受到破坏。例如,脑血管老化、血管风险因素(如高血压、高脂血症和糖尿病)以及动脉粥样硬化导致的大血管和微血管阻塞,分别导致急性或慢性血流减少,引发缺血性中风或血管性认知障碍。神经血管调节的变化也发生在神经退行性疾病中,如阿尔茨海默病(AD)、额颞叶痴呆和其他蛋白质病(淀粉样β(Aβ)、tau、α-突触核蛋白、TDP43等)。在这些情况下,通常观察到脑灌注、免疫失调、血脑屏障破坏、营养供给失败和清除路径的改变,这些可能导致大脑功能障碍和认知损害。因此,神经血管健康对于维护大脑健康至关重要。保护和促进脑血管健康的措施仍然是预防血管性认知障碍策略的必要组成部分。
血管周围间隙
使用高分辨率双光子成像的体内实验表明,动脉周围间隙是允许微球在啮齿动物中快速单向前流的巨大通道。小鼠动脉周围间隙(PVS)中微球的速度约为10–20 μm/秒。无法获得穿透性小动脉周围PVS的动力学数据,但由于这些空间在几分钟内被荧光右旋糖酐示踪剂(4–100 kDa)填充,速度可能在相同范围内。在组织学切片中,PVS表现为动脉壁外侧与脑实质(BP)之间的空间,可以通过脑内注射印度墨水颗粒或荧光聚苯乙烯微球实验性扩展。当动脉进入大脑皮质表面时,脑膜从大脑表面反射到蛛网膜下腔(SAS)的血管上,从而将其与大脑皮质分开。组织学研究无法在正常大脑中观察到皮质动脉周围的PVS(动脉周围间隙),因为平滑肌层、基底膜(BM)和一层脑膜下层在组织固定时被压缩在一起。然而,在白质和基底神经节中,动脉周围有两层脑膜下层,之间有一个PVS。(见图2)这些组织学研究尚未通过体内研究得到证实。最近的一项研究记录到,脑膜反射的PVS分离没有功能意义:CSF示踪剂独立于脑膜覆盖程度,沿动脉周围间隙深入进入大脑。使用正常成年犬脑组织证明,在白质中,动脉具有两层脑膜下层,之间存在潜在空间。脑表面的脑膜下动脉的分支穿过大脑皮质,通常不分支,供应下方的白质。需要注意的是,充满液体的空间在死亡后基本消失,因为能量耗竭导致膜电位丧失,引起细胞不可控的肿胀。其中一个最佳例子是,体内约占总组织体积20%的细胞外空间在组织学切片中完全丧失。因此,在开发大脑液体运输模型时,应谨慎使用组织学切片中的观察结果。事实上,注射到CSF中的示踪剂由于死亡后颅内压(ICP)的快速变化,将以高度非生理的方式移动。引文23 该研究展示了在用4%多聚甲醛进行心脏灌注期间,CSF示踪剂从PVS移向血管壁内部的动画(视频S1)。
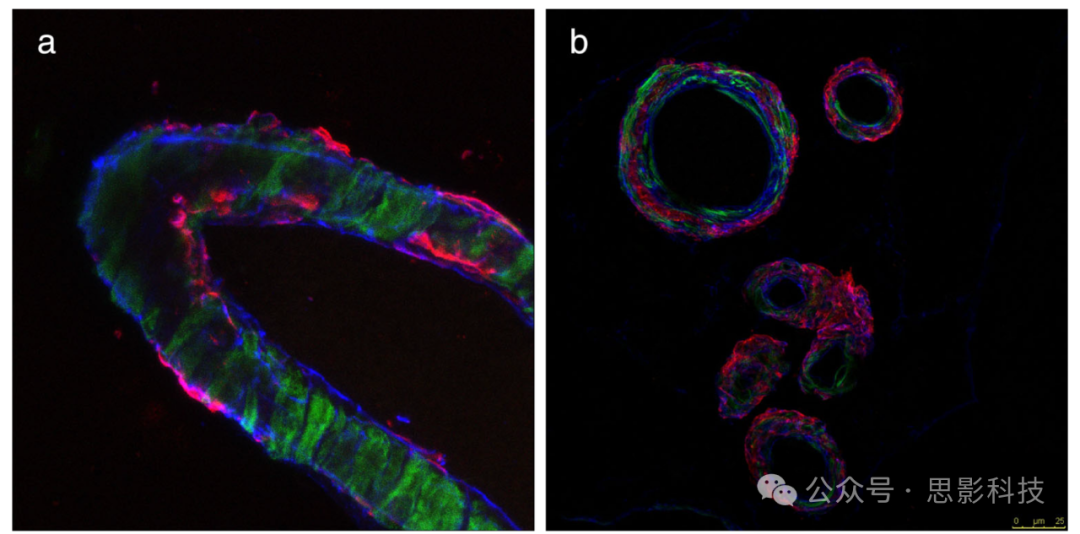
图2 Aβ在血管周围排泄路径(IPAD)中的积累示例。
(a) 显示人类CAA( 脑淀粉样血管病)的组织学样本。Aβ(红色)沉积在脑膜下动脉壁的基底膜(胶原IV为蓝色)中。
(b) 脑膜下动脉的横截面显示平滑肌。肌动蛋白为绿色,Aβ共定位为红色。(未发表的数据)
缩略语:DTI = 扩散张量成像;MRF = 磁共振指纹识别;DSC = 动态敏感性对比;DCE = 动态对比增强;ASL = 动脉自旋标记;SWI = 敏感性加权成像;fMRI = 功能性磁共振成像;MRS = 磁共振波谱。
PVS在MRI上表现为直径小于3毫米的脑脊液(CSF)信号的小线条或点状结构,位于皮质下灰质和白质中,并且其方向与穿通小动脉一致(见图3)。当动脉周围有一些液体,通常为CSF信号时,PVS才可见,并且在基底节和半卵圆中心的白质中最为清晰。T2加权成像是观察PVS最敏感的序列;T1加权序列的敏感性较低,因此通常只能识别较少且较大的PVS。尽管大脑中的静脉周围也存在空间,但这些通常在MRI上不可见。PVS在MRI上的可见性随着年龄的增长而增加,并且PVS的出现具有高度遗传性。然而,PVS的可见性在很大程度上依赖于序列参数和场强。因此,使用更敏感的序列时,更多的PVS会在MRI上可见。PVS可以通过视觉评分或计算方法进行量化,后者能够评估PVS的数量、总量和单个PVS的体积、宽度、长度、聚集和方向。这些定量指标可以用于跟踪PVS特征与组织损伤发展的关系,例如白质高信号(WMH)的形成。PVS量化的临床诊断价值目前是神经流体研究的一个活跃领域。在MRI上观察到的扩大PVS(EPVS)中的液体积累可能源自脑淀粉样血管病(CAA)中穿通动脉壁中Aβ的积累,但也可能是废物蛋白清除受损、血管周围炎症以及血脑屏障整体功能障碍的间接标志。
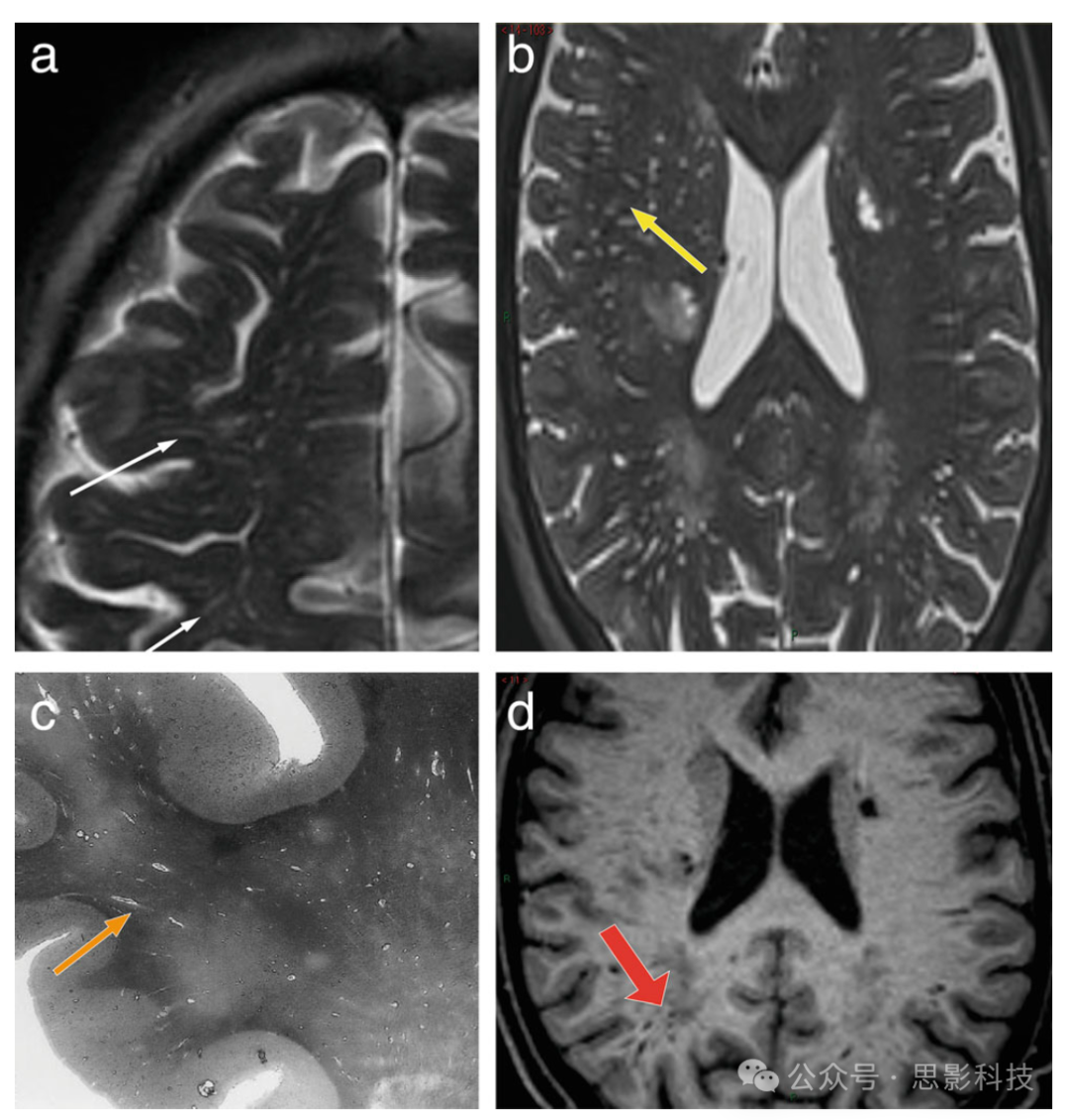
图3 常规MR图像上PVS的表现
(a) 位于半卵圆中心水平的轴位T2加权图像,显示多个细长的线状结构,呈现类似CSF信号的高信号(白色细箭头);
(b) 位于侧脑室水平的轴位T2加权图像,显示一些点状结构,呈现类似CSF信号(黄色箭头)。PVS的形态(线状与点状)取决于血管与图像平面的平行或垂直关系。
(c) 一张组织学切片,显示多个充满液体的线状PVS(橙色箭头)。
(d) T1加权轴位图像显示线状PVS,具有类似CSF信号的低信号(红色粗箭头)。
间质液(ISF)组成
在大脑的液体中,间质液(ISF)可能与神经元和胶质细胞的通信最为密切。尽管ISF的确切来源尚不清楚,但据信其起源于血管、脑脊液(CSF)和细胞源。ISF是水基的,包含一系列离子、蛋白质、神经递质和其他信号分子。此外,ISF中还含有大脑代谢产物和其他废物,这些都是中枢神经系统高代谢周转的结果。这些产物的组成和浓度受到诸多因素的影响,如神经活动、昼夜节律、生物性别和年龄。在这些成分中,诸如Aβ、tau、α-突触核蛋白和亨廷顿蛋白等蛋白质因其与神经退行性疾病(如AD、帕金森病(PD)和亨廷顿病(HD))的关联而受到特别关注。Aβ肽是在淀粉样前体蛋白经过蛋白水解切割后分泌到ISF中的。Aβ易于聚集成神经毒性纤维,构成了AD特征性的细胞外老年斑块,并能吸引存在于ISF中的可溶性Aβ。因此,老年斑块的密度和生物化学组成可以影响ISF中Aβ的浓度。与Aβ不同,tau、α-突触核蛋白和亨廷顿蛋白主要在神经元和胶质细胞内聚集。然而,体内外的证据表明,这些蛋白质的致病性聚集体被主动分泌到ISF中,这种分泌可能促进了AD、PD和HD中病理在大脑区域间的细胞内传播和扩散。ISF、CSF与壁细胞活动之间的通信在疾病进展过程中对蛋白质聚集物浓度和扩散的贡献程度,是神经液体研究的一个活跃领域。
脑脊液(CSF)动力学
经典的CSF生理学理解是,脉络丛(CP)、脑池空间和硬脑膜窦中的蛛网膜颗粒(Ags)是决定CSF持续生成、单向流动和主要被动吸收进入硬脑膜静脉窦的关键解剖结构。然而,在脑室内注射示踪剂后,显著数量的示踪剂进入邻近的中枢神经系统组织,并在CSF脉动驱动下分散到CSF系统的所有隔室。示踪剂扩散也可能发生在相反的方向,即从蛛网膜下腔(SAS)向侧脑室,这支持了除传统“脑室-脑池灌注”外的CSF生成和吸收的替代模型。这种CSF从脑室向BP(脑实质)的移动符合代谢废物清除的概念。此外,传统认为蛛网膜颗粒是CSF吸收的主要部位,必须根据新的发现进行修正,这些发现表明其数量、大小和解剖分布在整个人类生命过程中不断变化。实验、临床和放射学发现指出,经典的CSF生理学假说无法解释许多现象和病例,显示出这一概念需要更新的必要性。例如,具有长回波时间的ASL MRI最近已识别出蛛网膜下腔CSF与硬脑膜动脉之间的水交换。
CSF的脉动流动可能是清除机制的关键贡献者。每一次心跳,少量CSF在颅-脊系统的两个主要隔室,即颅腔和脊髓管之间来回移动。自从1825年马根迪发现“CSF”以来两个世纪,人们认识到这些CSF-振荡在维持大脑健康中的关键作用。这些振荡可以促进血液通过脑血管系统的循环,主要是在仰卧姿势下,当颅骨的顺应性降低时。尽管CSF振荡并不属于原始的蒙罗-凯利学说的一部分,但在收缩期和舒张期的这些振荡仍然是维持恒定颅内压(ICP)的基础。相位对比(PC)MRI研究清楚地显示,在收缩期,颅内CSF体积减少以适应动脉血流的流入,反之亦然。CS系统中的CSF振荡也可能促进代谢废物从大脑中清除,这主要发生在睡眠期间。事实上,仰卧姿势下的CS脑血量(SV)是直立姿势的两倍以上。颅骨和脊髓管隔室组织的机械特性在调节ICP以及CSF振荡的动力学中起着重要作用。与脊髓管中的硬脑膜相比,颅骨中的硬脑膜受到颅顶的更紧密限制,尤其是在腰椎区域。这使得CSF脉冲压力能够进一步传播到脊髓管。需要认识到,颅骨和脊髓系统都参与了CSF的有效振荡。
神经液体生理学:啮齿动物与人类的差异
啮齿动物和人类在解剖学和生理学上存在重要差异,在将鼠类模型的结果扩展到人类时必须加以考虑。从宏观层面看,鼠脑宽度约为10毫米,而人脑宽度约为140毫米。人类大脑皮层具有褶皱,即脑沟和脑回,而鼠类大脑皮层是平滑的。ISF的流动在人类中发生在广泛且复杂的空间几何结构中。鼠类研究表明,这种流动主要沿着主要白质路径进行,而人类中这些路径丰富,而鼠类中则较为稀疏。尽管实验证据有限,质量传输模型支持人类中的液体运输可能由平流和扩散共同描述,而仅由扩散可能足以解释鼠脑的情况。在微观层面上,定位于血管周围星形胶质细胞末端的星形胶质水通道AQP4支持血管周围的液体和溶质交换。有趣的是,鼠类中内皮细胞与实质中的AQP4通道比例为7:1,而在人类中仅为2:1,这表明星形胶质水运输和实质在介导更大人脑液体运输中可能起更大的作用。鼠类模型中的液体运输在睡眠期间更为活跃,越来越多的证据表明人类也可能如此。然而,鼠类是夜行性的,具有高度碎片化的睡眠结构,每10–15分钟经历一次快速眼动(REM)睡眠,而人类则是昼行性的,每90–120分钟经历一次REM睡眠。鼠类是否具有明确的非快速眼动(NREM)阶段尚不清楚;人类具有可以通过脑电图(EEG)识别的明确NREM睡眠阶段。随着年龄增长,鼠类的睡眠驱动力呈增加趋势,而人类则呈下降趋势。这些以及其他关键差异,包括动脉和静脉的分支、脑膜淋巴管、CSF体积和周转率等,在将动物观察结果转化为人类状况之前,需要得到充分表征。
CSF介导的大脑废物排泄路径模型
传统模型
自2012年以来,“糖淋巴”(glymphatic,glial-lymphatic)一词实际上已成为用于描述通过动脉/血管周围路径进行大脑清除的总称。该概念于2012年提出,使用体内双光子显微镜和动态对比增强磁共振成像(DCE-MRI)(见图4)。糖淋巴系统被确定为一个宏观的大脑废物清除网络,利用由星形胶质细胞形成的独特复杂的血管周围通道清除BP(脑实质)的间质空间中的代谢废物。该系统涉及CSF沿着动脉周围间隙(PVS)的流入,随后通过AQP4介导的CSF与组织的交换,ISF“冲刷”大脑组织并吸收可溶性废物,随后通过静脉周围空间排出。在重要的后续研究中,显示这种由星形胶质细胞介导的CSF和ISF的全脑运输主要在NREM睡眠期间增强,当时慢波EEG活动较高。除了清除废物外,糖淋巴系统还可能有助于循环必要的营养物质和神经递质,并可能作为药物递送的路径。这表明,利用糖淋巴系统可以更好地治疗主要的急性和慢性大脑疾病。已经提出了多个额外的CSF排出清除路径,包括沿着颅神经和淋巴管。
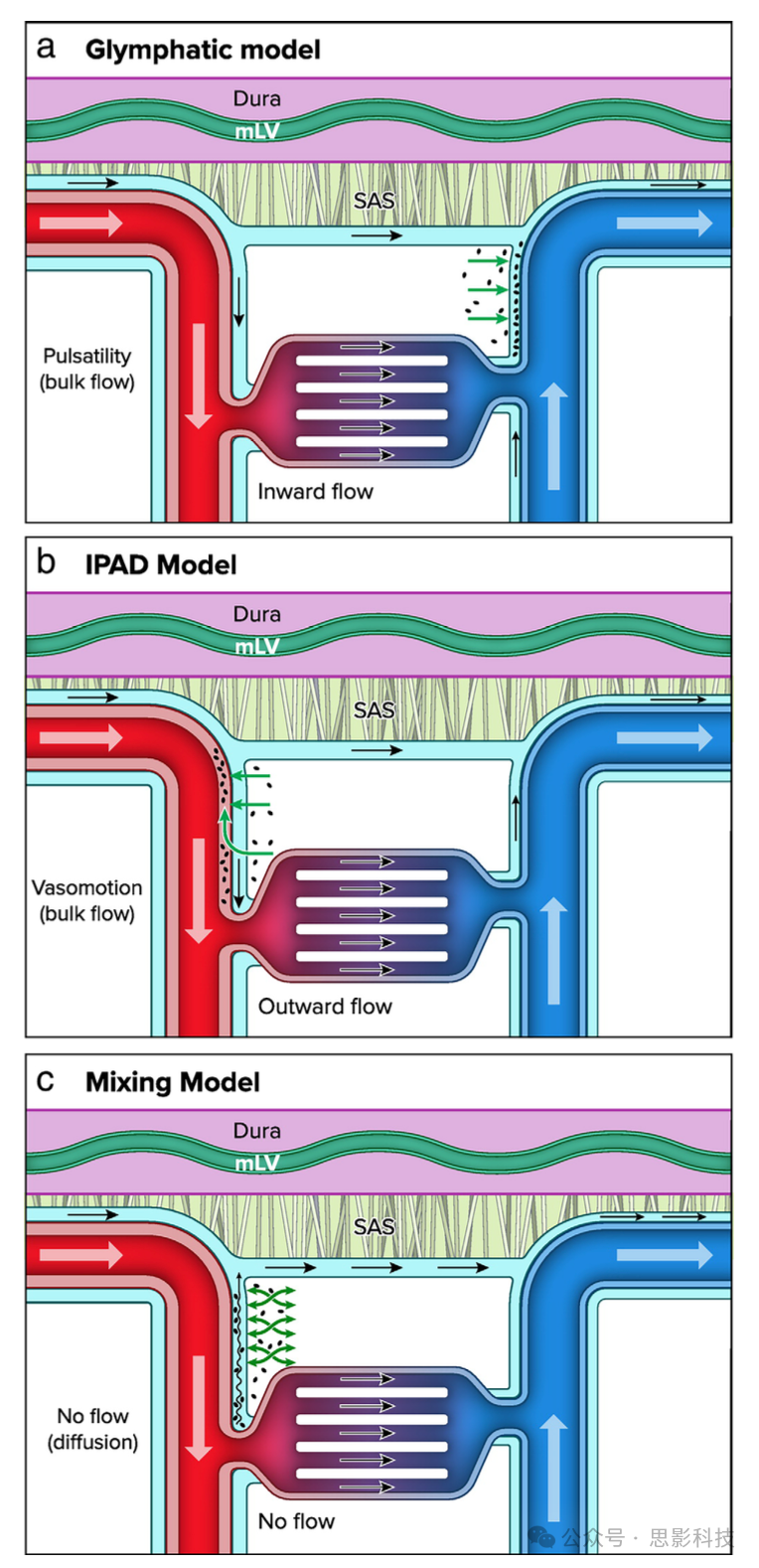
图4 大脑废物清除路径:(a) 糖淋巴模型描述了ISF和废物从动脉周围动脉空间向静脉周围空间的流动;(b) IPAD模型建议ISF和废物通过动脉周围空间沿动脉血流的反方向清除;(c) 混合模型建议穿透性血管没有流动,而是ISF和废物通过动脉脉动等生理过程“混合”。
作为糖淋巴模型的替代或补充,已提出了动脉内膜周围排泄路径(IPAD)模型。IPAD位于血管平滑肌细胞层和血管基底膜(BM)之间16(见图2)。该模型提出,排泄毒素如Aβ的ISF进入IPAD路径,并沿动脉血流的反方向在动脉内膜中移动,最终朝向颈部淋巴结。液体和废物通过离心方向从大脑中传播,这主要由平滑肌细胞的内在收缩活动(也称为血管运动)支持。
超越传统模型:混合假说
有强有力的证据表明,脑表面存在脑脊液的整体流动。然而,流动是否继续沿着穿透血管的PVS尚不清楚。蛛网膜下腔(SAS)水平的流动并不一定意味着沿着穿透血管的流动的继续,质量守恒可能通过CSF的多个平行排出路径得以维持,包括硬脑膜淋巴管、通过筛板沿嗅神经和脊神经。这些路径可能绕过大脑组织。最近的研究显示了鼠脑组织的电子显微镜细节,显示覆盖穿透动脉进入大脑的入口位点的脑膜。这些膜预计会阻止细胞和微米级颗粒的进入,但可能不会限制水和小溶质的流动。然而,这一解剖屏障限制了基于颗粒跟踪确定流动特征的方法,通常通过啮齿动物的体内成像技术进行。
尽管目前缺乏关于穿透血管PVS沿线的血管周围流动模式的实际数据,建模工作表明,整体流动并非增强CSF与脑ISF之间溶质交换的必要条件。流体混合也可能提供一种有效的溶质交换机制,既允许从大脑清除废物,也允许营养物质和其他可溶性因子沿同一路径从CSF进入间质。这一路径最有可能由穿透动脉沿线的空间所代表,因为静脉沿线的PVS似乎有限或非常小。在这种观点下,SAS(蛛网膜下腔)水平上“新”CSF的持续供应是提供废物清除浓度梯度所必需的。穿透血管沿线的PVS则提供了低阻力路径,在这些路径中振荡诱导混合,从而极大地增强了CSF与ISF之间的交换。
迄今为止,在啮齿动物中发现的增强示踪剂分散的振荡包括由心跳引起的血管脉动,以及血管运动。呼吸也被认为是一种驱动力,尤其在人类中。这些潜在的CSF–ISF交换驱动力已经在糖淋巴以及IPAD68 理论中被涉及。然而,这些概念在排水路径的解剖划分和流动方向上有所不同。方法学的差异,包括将示踪剂注射到CSF中与将示踪剂注射到实质中的差异,可能在其中发挥作用。因此,溶质分散可能取决于注射本身引起的压力梯度,或局部注射产生的陡峭浓度梯度。两者的方法学限制都可能轻易地模糊溶质移动的生理路径。因此,目前需要的是那些微创技术,这些技术将示踪剂注射与溶质移动分析分离,或者不依赖于示踪剂注射,包括直接在人体中研究CSF–ISF驱动力的需求。这些技术进步可能为穿透动脉PVS中是否存在整体流动或混合提供决定性的答案(见图4)。
MR成像技术
无对比增强技术
扩散张量成像
基于体素内非相干运动(IVIM)的扩散MRI和沿血管周围空间的DTI分析(DTI-ALPS)已被用于研究间质中的水分子运动,作为研究ISF动力学的间接方法。基于IVIM的ISF测量依赖于在广泛的b值范围内获取数据,更强调中间b值,因为认为ISF表现出的扩散值介于由随机定向的微血管床中的灌注引起的伪扩散值和组织扩散之间。DTI-ALPS依赖于侧脑室水平上PVS的特定解剖特征,其中髓质静脉与主要纤维轨迹垂直运行。DTI-ALPS的主要优点在于它使用传统的DTI序列,因此允许回顾性分析。然而,DTI-ALPS指数在多大程度上真正是糖淋巴功能的标志,仍然是一个积极争论的主题。这种技术的主要批评包括信号对CSF不具有特异性敏感性,缺乏足够的证据证明测量信号仅限于PVS,以及邻近侧脑室的可见PVS不是静脉周围的。然而,通过将运动敏感梯度与长回波时间读出相结合,MR信号可以限制在主要和穿透动脉周围的CSF,从而允许我们测量PVS中CSF的流动性。另一种利用扩散技术进行大脑清除成像的方法依赖于多b值采集和将信号分裂为三个隔室:灌注和扩散部分,以及第三个可能反映CSF或ISF的中间部分。
CSF流动的相位对比测量
非侵入性量化神经液体动力学存在诸多挑战,特别是流体速度的动态范围和隔室的大小。定量CSF流动测量通常通过相位对比MRI(PC-MRI)获得。来自PC MRI研究的一些显著观察结果如下(见图5)。CSF流动脉动性是由心动周期中颅内动脉流入和静脉流出之间的暂时性差异驱动的,并且进一步受到颅内顺应性、呼吸和胸内压力的调制。颅内顺应性越大,收缩期离开颅腔的CSF体积越少。静脉流出也发生在收缩期,并且似乎是CSF振荡的主要来源。静脉流出还受到呼吸的影响。呼吸与胸内压力的变化相关,这直接影响右心房和硬膜外静脉的压力。吸气时,胸内压力下降,在颅腔和胸腔之间产生反向压力梯度,促进静脉流动。呼气时,静脉回流被胸内压力的上升所阻断。硬膜外静脉体积的变化也驱动脊髓CSF流动。引文75对CSF振荡的起源和驱动力的深入研究导致了基于MR的颅内顺应性和压力量化的发展,如图5所示。
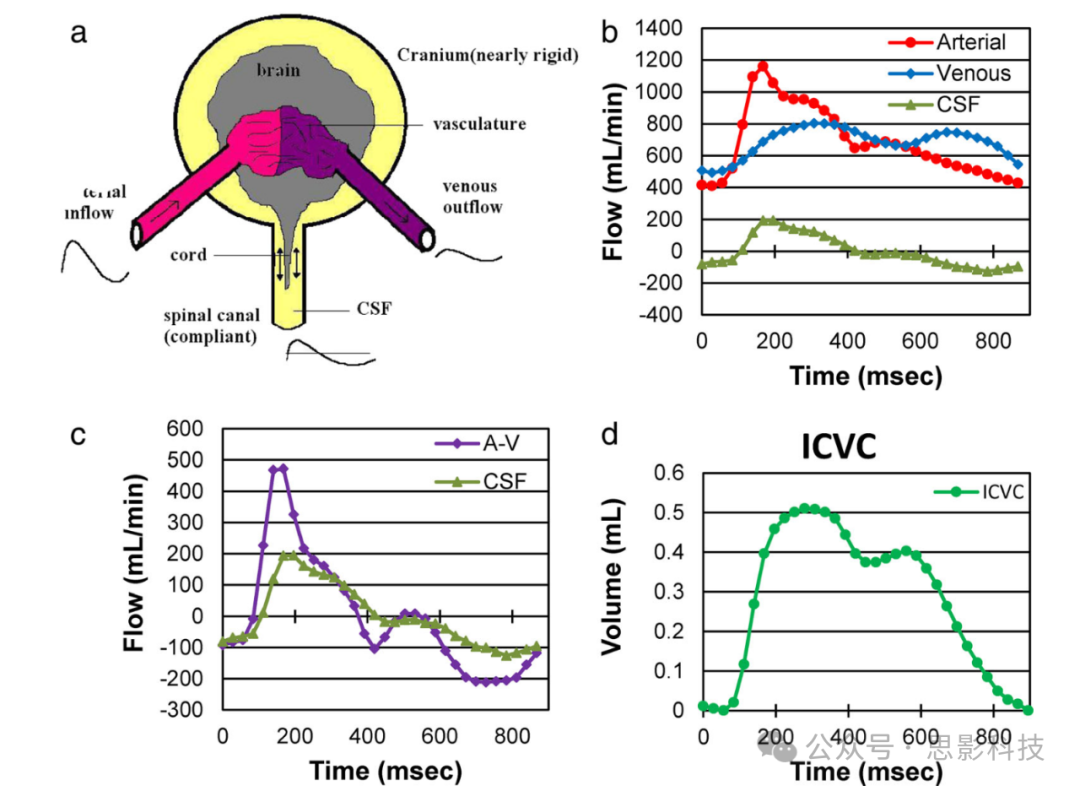
图5 (a) CS系统的简化模型。(b) 动脉流入、静脉流出和CSF流的体积流量波形。(c) CSF流波形与动脉减静脉(A-V)流波形的对比图。这两个波形不完全相同,意味着颅内容积(ICV)不是恒定的,因此颅骨具有顺应性。CSF波形遵循净颅内血流的模式,表明A-V流驱动了CSF脉动。(d) 通过对动脉减静脉流进行时间积分获得的心动周期内的颅内容积变化。
动脉自旋标记用于测量跨血脑屏障的水运输
脉络丛(CP)或血脑脊液屏障(BCSFB)被认为是脑中CSF分泌的主要来源,因此代表了大脑液体动力学的关键兴趣点。动脉自旋标记(ASL)MRI使用动脉血-水作为内源性对比剂,进行无创的灌注成像。ASL对脉络丛灌注的测量可能代表CP(脉络丛)功能的一个有价值的生物标志物。最近的研究应用了标准的ASL技术来估算CP灌注,显示出对衰老、血管生成和镰状细胞病的敏感性。另一种方法将ASL方法与长回波时间(TE)读出结合,用于量化动脉血-水通过BCSFB输送到脑室CSF的速率(见图6a)。这项方法由Petitclerc等人在人体大脑中进行了转化和扩展。 这种无创成像技术捕捉到了血液与CSF隔室之间快速且全脑范围的水交换,不仅在脑室中的BCSFB,也在SAS中(见图6b)。
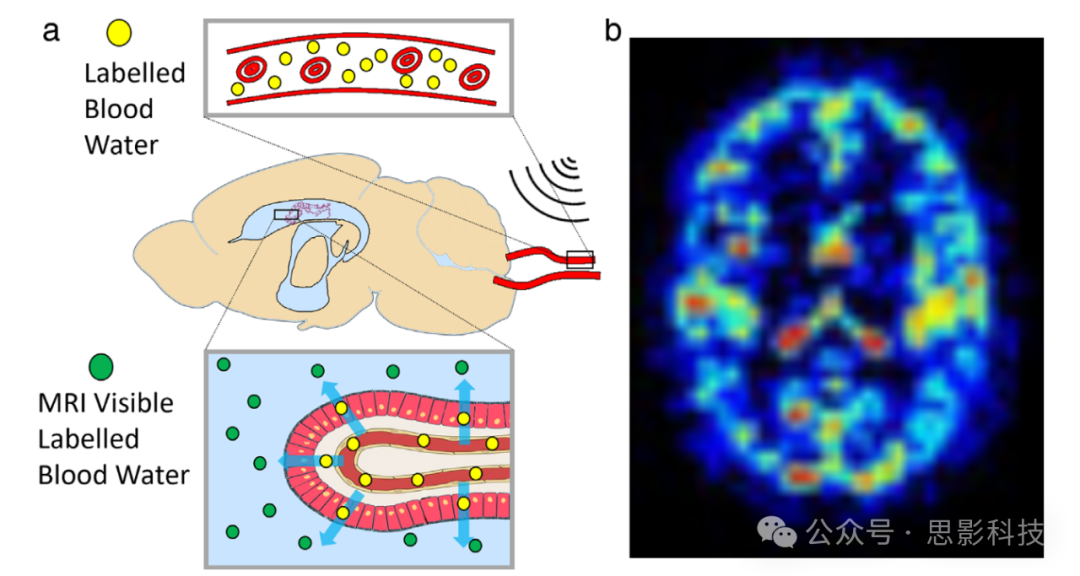
图6 (a) Evans等人提出的概念示意图,MRI信号来自被标记的动脉血水输送到CSF的情况。
(b) 人脑中动脉血-水输送到CSF的估计分布图,捕捉了血液到CSF的快速且全脑范围的水交换。
功能性磁共振成像(fMRI)测量CSF流
成像CSF流动的一个关键技术挑战是它随时间迅速变化,许多成像技术没有足够的时间分辨率来捕捉这些动态。此外,CSF流动相对于血液较慢,标准的速度测量方法如相位对比MRI对慢流的敏感性相对较差。最近的一项方法学发展是重新利用fMRI采集来高时间分辨率测量CSF流。使用短重复时间(TR,如<500毫秒)的快速fMRI会导致流动相关增强,在这种情况下,流动液体在成像体积(volume)的边缘切片上表现出更亮的信号(见图7)。
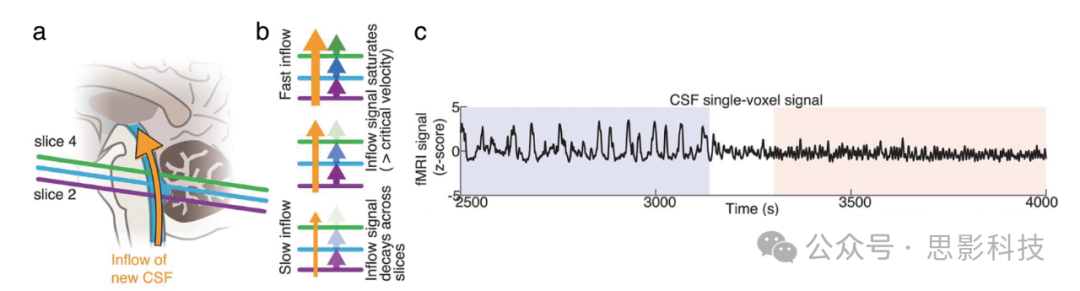
图7.使用流入效应在2D快速fMRI中测量CSF动态
(a) 将快速fMRI序列采集体积的底部切片定位以与第四脑室相交,导致新流入效应,上升流动液体生成明亮信号。
(b) 液体流速导致CSF在fMRI体积的底部切片上呈现不同的信号强度分布,基于流动CSF在到达某个切片前经历的激发脉冲次数。流速慢时,CSF在通过最低切片时会经历更多的RF脉冲,导致信号强度迅速降低,从而新流入效应消失。流速快时,新流入效应可以在多个切片上被识别。
(c) 该方法已被用于识别在NREM睡眠期间特异性出现的第四脑室内高速度脉动CSF流动。
这种现象是由于飞行时间效应:流入采集体积的新液体尚未经历RF脉冲,因此在边缘切片中产生明亮的信号。这一特性长期以来已知会在fMRI数据中导致高强度的血流信号,并且在快速时间尺度成像时这种效应特别强烈。虽然这些流动信号通常被认为是fMRI中的伪影,但也可以利用它们来测量与CSF流动相关的信号变化,通过将采集体积的边缘切片定位于与脑室相交。这些流动信号在使用短TR时最强烈;然而,也可以使用这种方法在更常规的采集中检测类似流动的CSF信号。这种方法的一个附加优点是能够同时使用血氧水平依赖(BOLD)fMRI信号测量皮层的血流动力学,从而允许研究CSF流动与全脑血管功能之间的耦合。MR成像CSF流的一个重要应用是理解睡眠的影响,因为睡眠与大脑废物清除率的提高有关,表明它可能在改变大脑液体动力学中发挥关键作用(见图7)。这些测量已经表明,睡眠转变了人脑中CSF的流动,并强调了基于MR的CSF测量技术如何发现大脑中液体流动的关键属性。
基于对比剂的技术
鞘内对比增强MRI
侵入性技术可以允许直接跟踪液体运输。宏环状MRI对比剂(gadobutrol)已被非标签地用作CSF示踪剂,剂量为0.25和0.50 mmol,注射到腰椎脊髓管内,最好与盐水混合,总量为1 mL。一般来说,尚未报告鞘内MRI对比剂在高达1.0 mmol剂量下的神经毒性,且未发现脑中钆的滞留迹象。由于gadobutrol高度亲水,并且在鞘内给药后位于完整的血脑屏障(BBB)外,因此可以被视为内源性脑溶质沿血脑屏障外路径从大脑和CSF中清除的替代标志物。使用注射后24–48小时内连续的T1w MRI扫描,可以监测示踪剂在CS隔室内的富集和清除特征(见图8)。
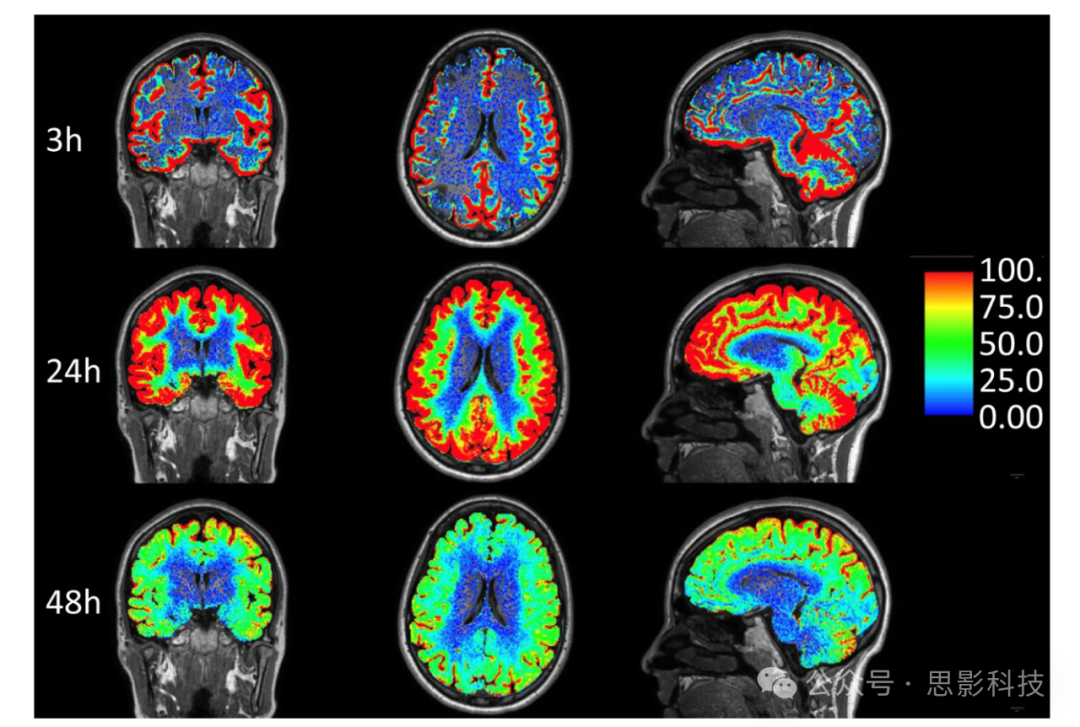
图8 鞘内增强MRI(gMRI):在鞘内注射MRI对比剂gadobutrol(0.5 mmol,分子量=0.6 kD)到腰椎水平后3、24和48小时,在脑外血管隔室中的归一化百分比T1信号增加。
脑膜下和脑室隔室中的示踪剂富集从图像中扣除。该方法可以展示通过鞘内途径注射的治疗药物到达脑血脑屏障外隔室的潜力,并用于评估大于水分子的分子的脑清除速率。人类的示踪剂研究表明,CSF清除是脑清除的一个限制步骤。脑组织中CSF示踪剂的富集高度依赖于示踪剂是否到达每个脑表面区域,这可能有很大差异。在此患者中,他正在接受松果体囊肿的检查,脑皮层和下部白质中发生了广泛的增强,但深部白质中未发生,这是典型的。因此,很可能深部脑白质的ISF来自于CSF以外的其他来源,并且CSF-ISF交换在深部结构的脑分子清除中没有主要作用。另一个需要注意的点是基底节中CSF示踪剂的非常有限的增强,这与啮齿动物研究显著不同,尽管在视觉上扩大的PVS中示踪剂增强明显(未显示)。
迄今为止,研究已经揭示了关于示踪剂从鞘内CSF空间进入BP(脑实质)的移动的有趣发现。首先,基于示踪剂的增强是鲜明的,并且早期在大动脉干如内侧颞叶附近的大脑表面区域到达。其次,鞘内gadobutrol可以以向心方式进入所有脑组织区域,从而绕过大脑完整的血脑屏障。第三,脑表面示踪剂增强的程度是不可预测的,并显示出显著的个体间差异,这对涉及CP和Ag的经典CSF生成和吸收概念提出了挑战。例如,针对特发性正常压力脑积水(iNPH)患者的鞘内研究显示脑室隔室内早期且持续的增强,而脑上凸部的示踪剂增强在整个扫描过程中有时完全缺失。此外,鞘内腰椎空间注射gadobutrol后,血液中gadobutrol的水平在脑上凸部示踪剂增强峰值之前达到峰值,表明CSF主要通过硬脑膜颗粒外途径重新吸收。
物种间的生理和解剖差异是显著的,在解释人类数据时必须考虑。例如,人类中鞘内钆的清除过程持续时间更长(天而非小时),通过鼻粘膜的CSF清除在人体中较小。同样,尽管睡眠剥夺严重阻碍了啮齿动物中的示踪剂清除,但在人类中这一影响更为温和。啮齿动物在硬脑膜窦中有真正的淋巴管,而在人类中,鞘内示踪剂显示CSF直接流向侧矢状硬脑膜,并在那里存在淋巴管,使得周边免疫系统与脑特定抗原在此位置之间可以进行交叉对话。基于gadobutrol的鞘内MRI有潜力作为多种神经退行性疾病如AD的CSF介导脑清除的替代标志物。
静脉对比剂基于的MRI
相比于鞘内给药,静脉注射的GBCA更少侵入性,并且在临床环境中更容易实现。IV给药的GBCA在健康人类中有一小部分进入BP和CSF,而血脑屏障泄漏在正常衰老和AD中更为普遍。使用IV-GBCA和流体衰减反转恢复(FLAIR)图像,可以在人体中可视化废物清除系统(即PVS和脑膜淋巴管)的流入和流出部分的对比剂增强。静脉注射的GBCA沿皮质静脉的皮下空间被认为是下游废物清除系统的一部分,静脉注射的GBCA从皮下空间泄漏到SAS随年龄增长而增加。在接近上矢状窦的皮质静脉周围形成囊状结构的报道虽然其意义尚未完全理解。基于IV-GBCA的成像已经清楚地显示了沿浅表硬脑膜静脉窦的脑膜淋巴管的存在。还有人建议,沿皮质静脉的皮下空间继续进入脑膜淋巴管。一项研究表明,静脉注射的GBCA在啮齿动物睡眠后清除增加。
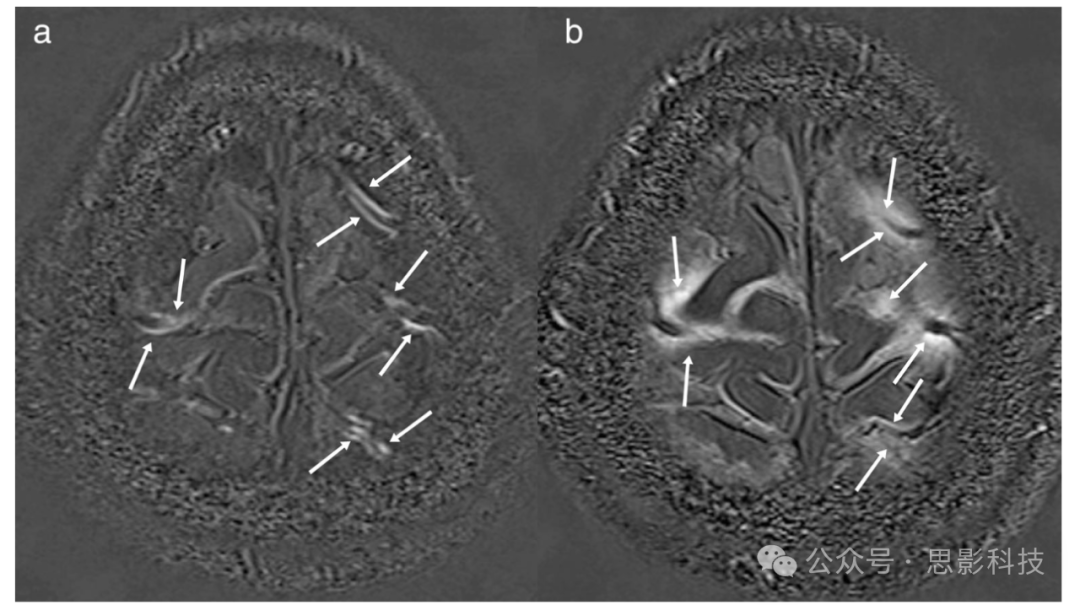
图9 静脉注射(IV)增强延迟MRI:在一名60岁男性疑似内淋巴水肿患者中,脑外血管隔室在鞘内注射单剂量钆基对比剂(IV-GBCA)后5分钟获得的归一化3D-real IR图像与鞘前对比。显示沿皮质静脉的对比增强线状结构(箭头)。这些线状结构被推测为皮下静脉排泄路径。
(b) 鞘前对比3D-real IR图像与IV-GBCA注射后4小时获得的3D-real IR图像的差分。对比剂增强扩散到皮质静脉周围的SAS(箭头)。钆基对比剂泄漏到CSF的程度已被报道随着年龄的增长而加速。
IV-GBCA基于的MRI也被用于研究眼睛和耳朵的清除路径。使用改良的IR序列对眼睛和耳朵进行延迟成像可能揭示青光眼和梅尼埃病的病理生理。线状GBCA已通过IV和鞘内给药显示在脑中沉积。环状GBCA被认为在人类中沉积量显著较小。至少,尚未显示出GBCA沉积的明显神经学效应。
计算流体动力学的计算模型
计算模型已被成功用于测试关于神经液体运动的假说,这些假说目前无法通过实验进行评估,并生成未来实验测试的新假说。通过计算建模进行假说测试,近年来关于糖淋巴系统中实质内方面的可行性的热烈讨论是一个很好的例子。糖淋巴假说规定,溶质通过整体流向静脉周围空间被BP(脑实质)冲刷,基于在小鼠中使用体内活体两光子荧光显微镜追踪荧光示踪剂。然而,这种成像方式无法直接辨别示踪剂运动的驱动力(扩散或对流)。采用多种方法的计算模型一致表明,通过整体流动的实质内溶质运动在物理上是不合理的,而实验证据与扩散驱动的溶质运输大体一致。计算模型还确定了我们对中枢神经系统液体运动物理学理解的关键差距,并有助于阐明在何种条件下净血管周围液体运动可能发生,以及涉及的物理机制。例如,许多模型已表明,虽然可以发生振荡性血管周围整体流动,但实验上测得的溶质(或示踪剂)的净运输不能仅由心动周期相关的动脉壁运动实现,需要额外的静态或动态压力梯度,可能与扩散(即分散)结合。开发有意义的计算模型的一个关键挑战是需要基于动物和人类实验中关于体内CNS的解剖和机械参数的现实假设。模型中过于简化或不现实的假设不可避免地导致不可靠的结论,因此实验、临床和建模研究人员之间的紧密联系至关重要。
建模还可用于补充、验证和增强测量的神经液体流动数据。例如,尽管4D PC-MRI是体内测量CSF速度的强大工具,压力梯度是中枢神经系统周围和内部液体运动的主要驱动力。液体运动的驱动力压梯度可以使用计算流体动力学(CFD)从测量的流体速度中估算。此外,结合PC-MRI数据与CFD确保体内液体流动测量符合物理基本定律。正在进行的技术开发旨在将流体力学的关键概念整合到成像工作流程中,以提高从成像中获得的流动测量的质量,例如,通过在相位对比协议中强制执行无散度速度,以及使用CFD进行降噪的深度学习方法来降噪4D数据。
计算模型还可以用于验证和扩展神经液体研究的理解,尤其是在评估不同清除路径和流动模式的相对贡献方面。
当前临床应用
脑内应用
过去十年中,识别脑废物清除路径及其在神经退行性中的作用引起了极大的兴趣。两个主要的临床相关MRI生物标志物是扩大PVS(EPVS)和白质高信号(WMH)。CAA( 脑淀粉样血管病)和AD患者可能在半卵圆中心具有增加的EPVS,而高血压性小血管病(SVD)和脑出血患者则在基底节有EPVS。此类患者的EPVS可能由于动脉壁(IPAD假说)内Aβ沉积或在间质空间(糖淋巴假说)中的Aβ沉积所致(见图10)。SVD和脑出血似乎与基底节EPVS的增加有关,可能由于动脉僵硬、血脑屏障损伤和稳态改变导致组织ISF增加。T2加权MRI上见到的WMH是动态的,代表几种不同的病因,其中之一是ISF增加。特定的成像如T1映射和DTI分析可能进一步有助于识别WMH的性质。
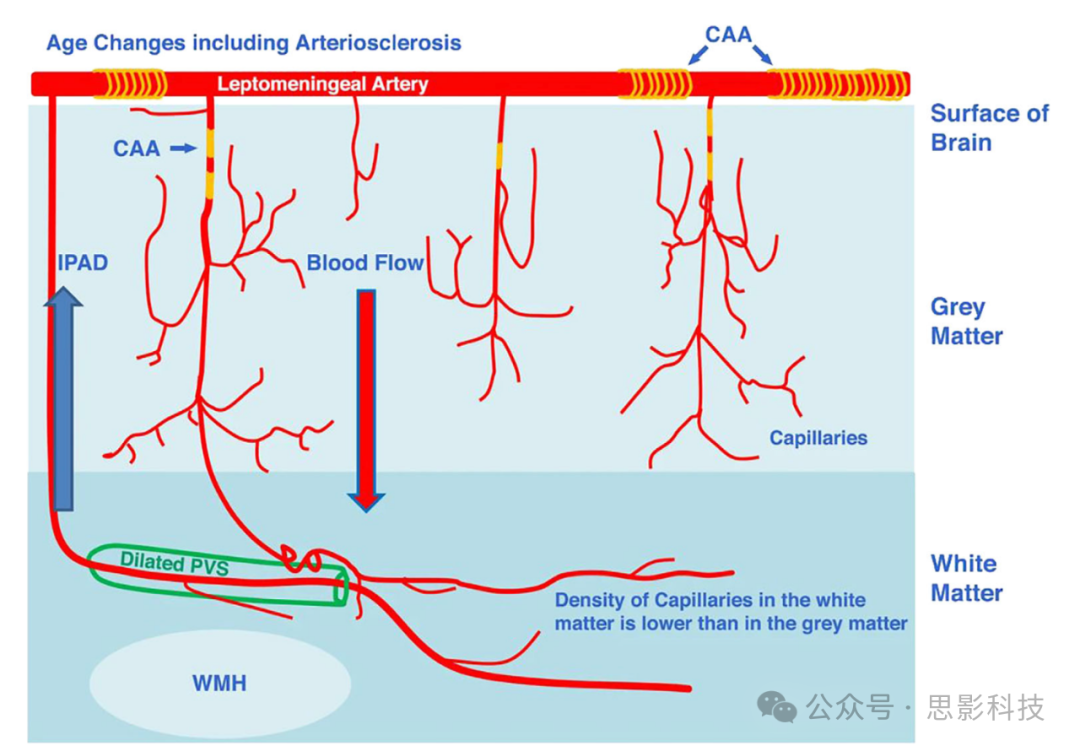
图10 一个示意图用来展示WMH与扩大PVS(EPVS)之间的建议关系。请注意,EPVS显示在有穿透性小动脉被双层脑膜下层包围的白质中,促进废物更容易在白质而不是灰质中积累。
CAA = 脑淀粉样血管病。IPAD = 动脉内膜周围排泄路径
在人类中成像脑废物清除路径的改变主要受限于MRI在空间和时间分辨率方面的不足。使用基于GBCA的成像在时间上识别出在iNPH、内淋巴水肿和青光眼患者中慢速清除区域似乎有一定前景。普遍关注于将DTI-ALPS应用于不同神经疾病患者,然而,解释时需要谨慎。脊前对比T2w FLAIR成像在研究脑膜淋巴管和可能的排出路径方面日益受到关注,并有潜力具有临床相关性。
除了MRI外,正电子发射断层扫描(PET)基于映射的CSF动力学在衰老和AD中也被讨论。PET研究表明,70%–85%的IV给药的脑渗透性放射性示踪剂从动脉血进入间质,并在90分钟内从大脑中清除。最近,使用PET观察到沿先前识别的CSF和静脉路径的CSF排出可被成像。最有趣的是具有最小特异性或非特异性脑结合位点的放射性示踪剂。在一系列使用有限结合PET示踪剂的研究中,定义了一个由示踪剂在脑室中减少的浓度组成的CSF清除指标,该指标以短10秒间隔采样。结果显示,与临床确定的轻度认知障碍和AD相关联的侧脑室放射性示踪剂清除减少,在测试淀粉样假说时,观察到脑室CSF清除与淀粉样病灶水平增加呈负相关。PET示踪剂时间活性曲线定义的CSF排出路径在很大程度上与黄金标准的鞘内给予钆对比剂MRI研究的发现相一致。PET排出路径已经被识别为上矢状窦、视神经和通过筛板向鼻甲的嗅神经通道(见图11)。如果这些观察结果得到复制和验证,表明可以通过现有的PET示踪剂收集CSF清除数据,以及通过鼻拭子采集CSF样本,从而可能避免进行腰椎穿刺以获取脑源性蛋白质。基于PET的方法仍然应被视为新方法,需要验证和额外的建模。目前,正在测试一种新型放射性示踪剂[11C]-Butanol,该示踪剂在大脑中不结合,作为CSF动力学的潜在生物标志物。
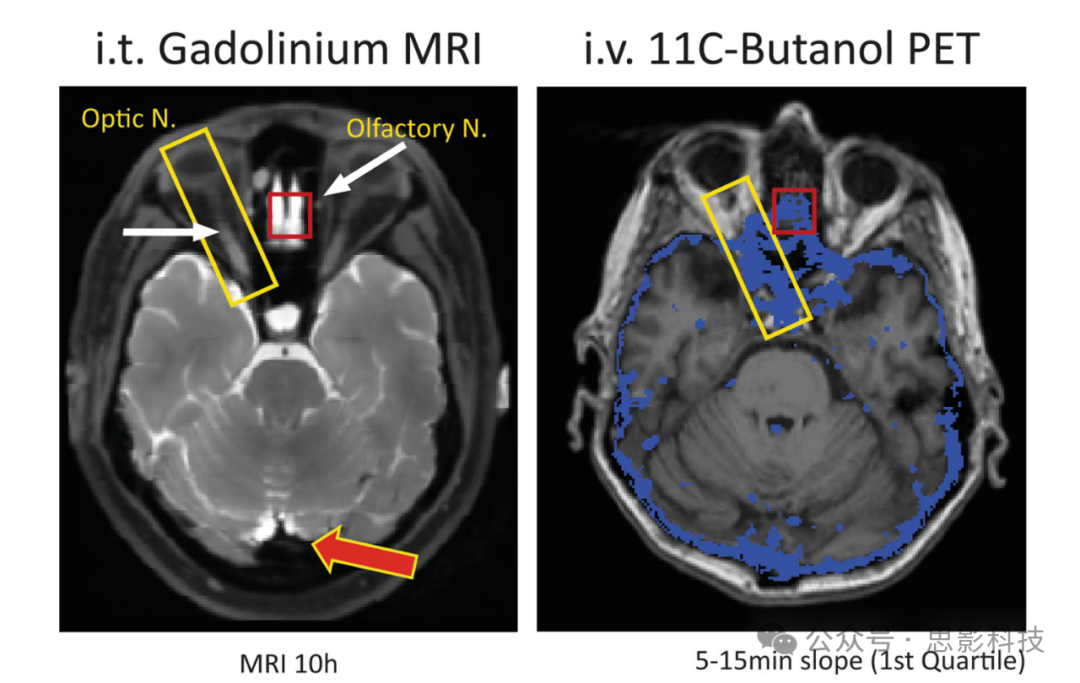
图11 显示了使用MRI对比剂钆(左)和11C-Butanol PET示踪剂(右)识别的CSF路径的解剖对应关系。
图中突出显示了几个通过MRI和PET识别的增强区域,包括视神经和通过颅底筛板的嗅骨板,嗅神经通过的鼻甲。左侧的MRI图像是在鞘内(腰椎)注射0.5 mL Magnevist(Gd-DTPA)后10小时获得的脂肪抑制T1图像,具有1×1×1 mm各向同性分辨率,重复时间=602 ms,回波时间=13.3 msec,视野=25.6 cm,由3 T GE MR750获得。右侧的MR-PET图像来自使用11C-Butanol的IV给药,通过SIEMENS Biograph 64进行动态(列表模式)60分钟成像,并与3 T SIEMENS Prisma的T1加权MPRAGE数据共配准。对于整个大脑,计算了注射后5至15分钟之间butanol斜率的体素级分布。右侧图像中的蓝色区域标识了“最慢四分位”的清除体素,叠加在相应的轴向MR图像上。
脊髓的应用
脊髓CSF的解剖学和生理学现已被理解得更加复杂,并且与传统的脊髓CSF生理学观点有所不同。额外的CSF生成可能发生在脊髓组织和血管内,CSF的吸收也通过神经根上的蛛网膜乳头和淋巴管进行。脊髓蛛网膜下腔(SAS)具有丰富的蛛网膜隔膜,有时可能阻碍CSF的正常流动。SAS中的流动受心脏脉动的影响,但更大程度上受呼吸运动的影响。CSF流入脊髓主要由呼吸压力驱动,而脊髓的液体外流则受心脏脉动的影响。蛛网膜炎或压迫导致SAS阻塞会增加液体流入脊髓。
脊髓CSF生理学的病理例子包括Chiari畸形、脊髓空洞症、蛛网膜囊肿和自发性脊髓CSF泄漏。这些疾病的病理机制尚不完全理解。脊髓空洞症是脊髓中CSF的积累,但这些囊肿如何形成仍然是一个谜。动态成像在临床上证明非常有用,并且结合计算建模,有助于理解这一状况。脊髓中CSF的成像具有挑战性,标准成像技术无法展示蛛网膜隔膜以及组织和液体的运动。心脏门控、重度T2加权扫描已揭示脑和脊髓组织的运动,并且在展示有时导致脊髓空洞症的异常蛛网膜隔膜方面极为有用(见图12)。
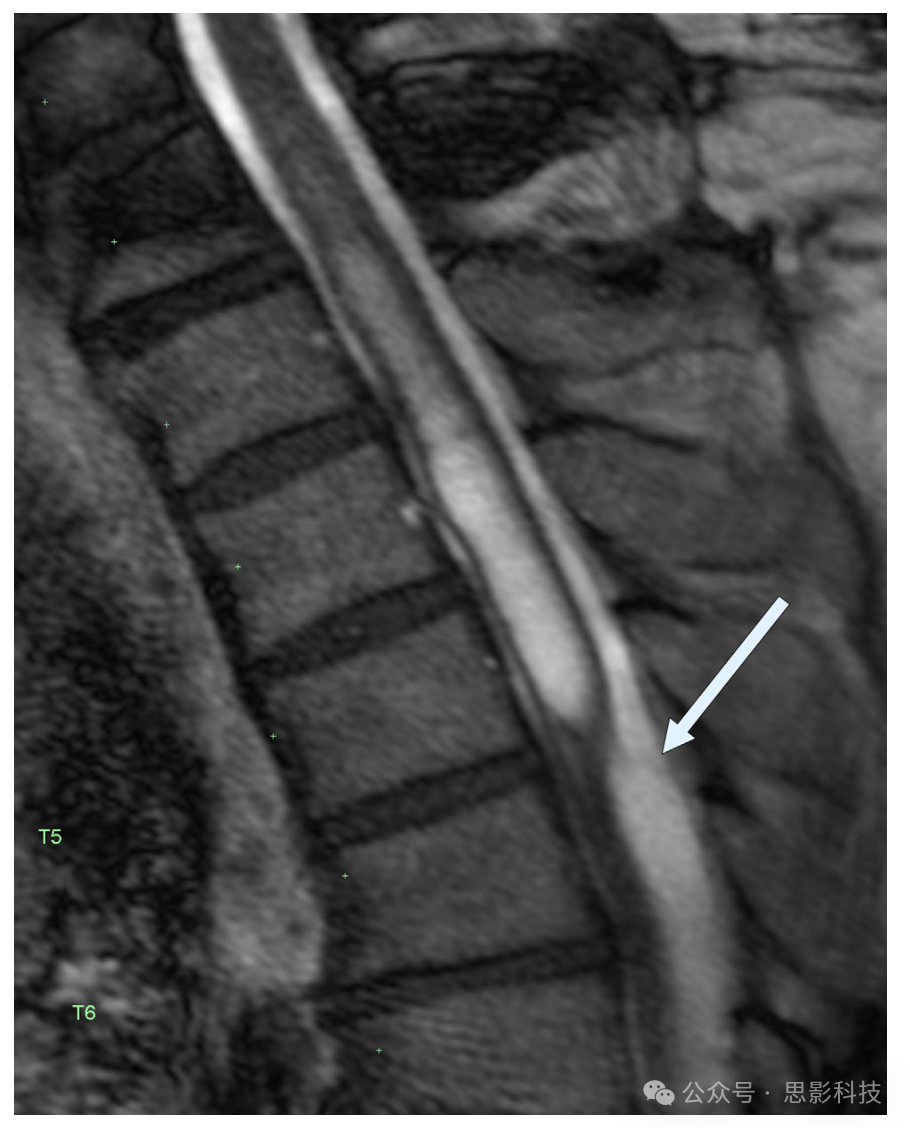
图12 心脏门控、重度T2加权矢状脊柱MRI的静止图像,展示了导致脊髓空洞症的蛛网膜隔膜(箭头)。该隔膜在标准T2 MRI上无法检测到。
总结
在2022年,神经液体国际医学磁共振学会(ISMRM)研讨会上,来自不同专业背景的国际专家概述了当前对神经液体解剖学、生理学及其动态成像的理解。脑血管不仅为我们的脑提供基本的营养物质,如氧气和葡萄糖,以确保神经胶质单元的最佳功能,而且在移除神经元活动产生的废物方面也是至关重要的。CSF似乎在大脑和脊髓内循环,提供了营养物质扩散、机械保护和废物清除的流体介质。一些CSF通过脑外实质路径如筛板和包围颅神经的鞘“逃逸”。在脑实质中,CSF与间质液(ISF)混合。所有神经液体的动态相互依赖,并以复杂的方式同步,这些动态可能受到诸如睡眠、呼吸、心脏活动、体位、脑和脊髓的顺应性以及血管平滑肌细胞的内在收缩活动(血管运动)等多个生理过程的影响。多种因素可以干扰这些过程,最终导致神经疾病。脑膜淋巴管和其他CSF排出路径的作用进一步增加了大脑清除路径的复杂性。体内MRI技术正在积极开发中,以识别清除路径改变的标志物。然而,我们对大脑清除的知识以及监测神经液体功能的MRI技术仍存在多个盲点。
EPVS(扩大的动脉周围间隙)可能是临床MRI上可用的阻塞性ISF和毒素排泄的最重要间接标志物之一。解剖学研究确实显示了EPVS与皮质中淀粉样沉积之间的联系,进一步强化了EPVS的临床重要性。然而,目前尚不清楚EPVS是否真正且始终具有病理性。
从成像的角度来看,MRI的一个最大挑战仍然是其有限的时间和空间分辨率以及信号的不特异性。IV钆基灌注研究允许监测示踪剂流入的最佳时间分辨率;然而,它们缺乏清晰评估对比剂所在隔室所需的空间分辨率。这引发了诸如“信号变化有多少是由于血管内增强?”和“有多少是由于示踪剂进入组织或CSF/ISF隔室?”等问题。鞘内注射清楚地显示了对比剂在CSF空间和脑组织内的分布,但存在长时间的时间足迹(24–48小时的间歇性观察)。对比剂向脑半球排出的整体影响使得即使是鞘内注射也难以清晰识别CSF从脑外实质的排出路径。非侵入性成像技术可以提供高时间和空间分辨率,但仅成像脑清除的溶剂,即CSF和ISF中的水,因此缺乏直接跟踪废物如淀粉样蛋白的敏感性和特异性。此外,在尝试隔离来自CSF或ISF的信号时,常常会产生残留的脑实质信号伪影。这可能会混淆测量并导致结果过度解释或缺乏显著性。
目前,鞘内使用MRI对比剂作为CSF示踪剂仅在研究中使用,因此迫切需要寻找非侵入性替代方法。研究表明,iNPH痴呆患者的CSF到血液的gadobutrol清除减少,尽管个体间存在显著差异。这一测量预计主要指的是CSF依赖的脑膜淋巴清除能力,这对tau的清除似乎特别重要。在未来的应用中,作为黄金标准的鞘内增强MRI可以与可能作为测量脑清除的非侵入性替代方法的MRI序列直接比较。在这里,DTI-ALPS成为了一种流行的,尽管备受争议的技术,通过区分沿白质PVS中的水运动与垂直于这些的轴突纤维中的水移动。然而,尚待确定的是,WM中水的移动是否是真实的脑皮层中更大溶质如AB和tau清除的有效替代标志。此外,DTI-ALPS信号的起源也是需要仔细解释的原因。最近,修改后的成熟MRI技术如ASL、fMRI和PC-MRI正在被研究,并在人体神经液体成像方面展现出前景。
研讨会上明显看出,在啮齿动物模型中进行十年的脑清除研究后,该领域正在快速迈向寻找成像模式以研究人类脑清除。许多最初为脑清除成像以外目的设计的MRI技术在该领域找到了新的角色,进一步展示了MRI作为理解我们复杂生理学的基本工具的多功能性。最后,研讨会表明,本文作者之间在一些关于脑清除成像和EPVS病因的概念上仍缺乏共识,这进一步表明需要协调的多学科努力以得出有意义的结论。


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









