






点击蓝字 关注我们
调控肠道菌群的宿主源代谢分子概述

iMeta主页:http://www.imeta.science
研究论文
● 原文链接DOI: https://doi.org/10.1002/imt2.88
● 2023年2月13日,瑞典卡罗林斯卡医学院杜鹃、Åsa Sjöling、武圣儒团队,西北农林大学姚军虎团队在iMeta在线发表了题为“An overview of host-derived molecules that interacted with gut microbiota”的文章。
● 本研究综述了宿主调节肠道菌群的机制,涉及肠源性分子,包括肠源性免疫系统分子、肠源性粘膜分子相关来源、肠源性外泌体 ncRNA 以及来自肠道以外其他器官的分子。这为了解肠道菌群和宿主的相互作用提供了系统的概述,为操纵肠道菌群的潜在方法提供了全面的来源,并为未来利用肠道菌群的个性化治疗奠定了坚实的基础。
● 第一作者:张晨光
● 通讯作者:杜鹃 ( juan.du@ki.se)、Åsa Sjöling (asa.sjoling@ki.se)、姚军虎 (yaojunhu2004@sohu.com)、武圣儒 (wushengru2013@163.com)
● 合作作者:刘慧峰、孙蕾、汪悦、陈晓东
● 主要单位:西北农林大学动物科学与技术学院,瑞典卡罗琳斯卡医学院微生物、肿瘤与细胞生物学系
亮 点
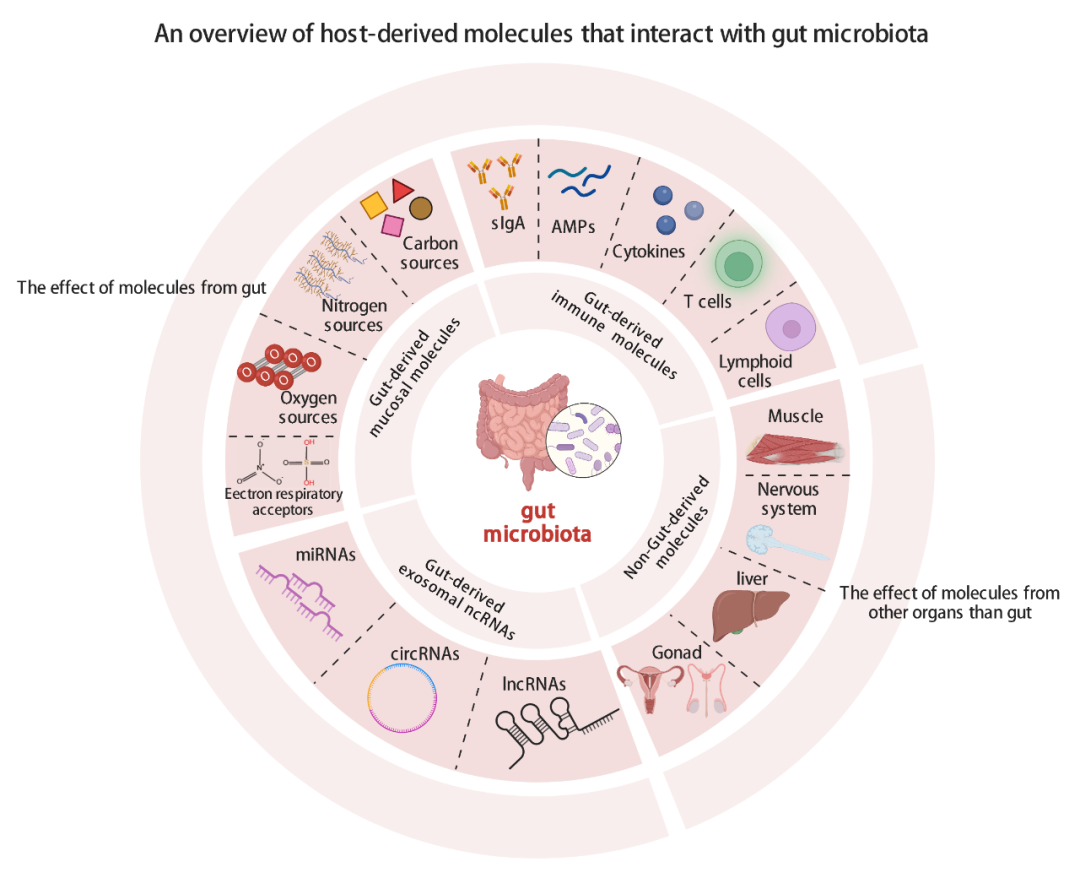
● 总结了调控肠道菌群的宿主源性分子影响肠道菌群的机制
● 归类综述了调控肠道菌群的宿主源性分子,包括肠道来源的免疫分子、肠道粘膜分泌的小分子、肠道源性外泌体ncRNA以及其他器官产生的分子
● 了解宿主源性分子如何调节肠道微生物群并影响疾病发病率,有助于开发新的预防和治疗干预措施,提高粪便微生物群移植的治愈率,甚至有助于预测个体的疾病易感性
摘 要
肠道中共栖的微生物包括细菌、古细菌、真菌、原生生物和病毒,它们彼此之间以及与宿主细胞之间存在相互作用。稳定的肠道菌群对调节宿主代谢和维持机体健康至关重要,而失调的肠道菌群可能诱发不同种类的疾病。此外,饮食是影响肠道菌群的主要因素,而宿主也可以通过其他因素塑造肠道菌群结构。这里,我们综述了宿主调节肠道菌群的机制,涉及肠源性分子,包括肠源性免疫系统分子(分泌性免疫球蛋白A、抗菌肽、细胞因子、CD4+效应T细胞和先天淋巴样细胞),肠源性粘膜分子(碳源、氮源、氧源和电子呼吸受体),肠源性外泌体ncRNA(miRNAs、circRNA和lncRNA),以及来自肠外器官的分子(雌激素、雄激素、神经激素、胆汁酸和乳酸)。这为了解肠道菌群与宿主互作提供了系统的概述,为调控肠道菌群的潜在方法提供了借鉴,并为未来利用肠道菌群的个性化治疗奠定了坚实的基础。
视频解读
Bilibili:https://www.bilibili.com/video/BV1id4y1J7ra/
Youtube:https://youtu.be/5415_b5G7i4
中文翻译、PPT、中/英文视频解读等扩展资料下载
请访问期刊官网:http://www.imeta.science/
全文解读
引 言
由细菌、古菌、真菌、原生生物和病毒组成的微生物群在食物消化、改变药物作用、神经信号、代谢过程、训练宿主免疫和调节肠道内分泌功能等多方面的作用受到越来越多的关注。一些信号连接,如肠-肾、肠-肝和肠-脑轴,已被确定并证明参与了肠道微生物群影响宿主表型的过程,无论是通过细菌与不同的肠细胞之间的直接作用,还是通过与微生物成分或者其代谢分子的间接相互作用。随着高通量测序技术的发展和无菌猪、无菌小鼠、基因敲除小鼠模型的建立,人们逐渐认识到肠道菌群对宿主健康和疾病发生的重要性。大量分子已被证明可驱动宿主与其微生物组之间的串扰,特别是微生物组在塑造宿主表型变化中的单向调控。相比之下,宿主衍生分子直接塑造肠道菌群的机制研究值得更多关注。
肠道菌群被认为是由宿主遗传、饮食、地理、社会接触模式等因素共同塑造。在这些因素中,饮食是一个主要的驱动因素,可直接影响肠道微生态。近年来,越来越多的研究发现,在相同的饮食或环境条件下,肠道微生物差异导致了宿主出现不同的生理反应,这提示肠道菌群的差异可能来源于宿主效应。此外,肠道菌群的代际遗传以及宿主遗传与肠道菌群之间的关联表明存在宿主因素塑造肠道菌群的潜在方式。
尽管越来越多的证据明确支持宿主塑造肠道菌群,但其潜在机制很少被讨论和系统地总结。值得注意的是,微生物组全基因组关联研究(mGWAS)已经确定了一系列与微生物群落定殖相关的基因。然而,潜在的机制主要集中在一些已经明确的来自肠道上皮的宿主源性分子对肠道菌群的影响,而直接作用背后的机制缺乏系统性梳理和总结。因此,本综述主要关注宿主源性分子与肠道微生物互作并调控肠道微生物群的机制,并进一步将这些相关机制分为4个部分:1)肠道源性免疫系统分子,2)肠道源性粘膜分子相关来源,3)肠道源外泌体非编码RNA(ncRNA)调控,4)肠道外器官源分子。总之,本综述的目的是系统地总结宿主源性分子调节肠道微生物群的机制,并加深我们对宿主-微生物群互作的理解。
肠源性免疫系统分子
免疫系统,包括先天免疫细胞和适应性免疫细胞。适应性免疫系统主要是对病原体的侵袭做出反应,随后诱导多种细胞因子响应微生物源性信号,启动炎症反应,清除致病菌。本综述对构成微生物群的所有个体微生物间的复杂网络不进行扩展,而是将重点放在整个微生物群上。
分泌性免疫球蛋白A
派尔集合淋巴结是肠粘膜免疫的重要组成部分,可产生分泌性免疫球蛋白A(sIgA)。sIgA是由IgA、连接链(J-chain)和分泌组分(SC)组成的二聚体,其中J-chain将两个IgA单体连接在一起,SC通过二硫键与IgA结合。除了针对侵入性病原体外,sIgA还广泛针对非侵入性共生生物。缺乏sIgA-包被的细菌可能对宿主造成侵害。重要的是,生命早期的微生物定植是一个连续的过程,与肠道sIgA的克隆结构平行变化。在成年人稳定的肠道菌群建立之前,sIgA多克隆抗体广泛覆盖共生菌。因此,IgA和微生物群之间的共同进化在生命的各个阶段影响着肠道菌群结构。
针对特定细菌抗原(例如,脂多糖和鞭毛)的sIgA对于维持肠道菌群的稳态至关重要。在正常生理条件下,sIgA主要通过4种方式调节肠道菌群:(1)sIgA包被的共生体(图1A-(1)):被sIgA包被的共生菌会被限制在肠腔内,使肠道免疫系统与无数微生物相关抗原分子的促炎作用最小化,从而避免了补体级联的激活。值得注意的是,sIgA包被的共生菌入侵肠道上皮时,sIgA可以通过与促炎吞噬细胞上的Fcα受体I(FcαRi)交联而引起炎症。(2)凝集(图1A-(2)):sIgA通过凝集作用限制肠道共生菌在肠腔内的运动。对于病原体,高亲和性sIgA可以阻止病原体的分离和扩散。(3)免疫包涵体(图1A-(3)):sIgA通过其SC的聚糖,将包被的共生菌锚定在肠道上皮的外黏膜层。(4)细菌基因表达调控(图1A-(4)):粘膜相关功能因子(mucus associated functional factor,MAFF)基因可上调共生菌对多糖的利用活性,增加产丁酸菌的定殖。丁酸盐等有益代谢产物可直接刺激结肠上皮细胞的增殖和再生,以应对上皮损伤。此外,针对单个微生物,sIgA有一系列调控措施,包括代谢调节、免受胆汁酸或噬菌体的影响以及运动性改变。值得注意的是,sIgA可通过调控细菌基因转录来影响细菌的形态、组成、生长、侵袭和免疫代谢功能。最后,调节T(Treg)细胞主导的IgA反应扩大有益菌(厚壁菌门),同时抑制潜在致病菌(变形菌门),从而调节肠道菌群的组成。
抗菌肽
与高亲和力sIgA相比,由多种组织(如瘤胃、肠道、肺、生殖道等)上皮细胞表达的宿主源性抗菌肽(AMPs)具有广谱抗菌活性。尽管AMPs在序列、结构和来源方面是一组不同的分子,但几乎所有AMPs都有一些共同的特性。AMPs含有特定的阳离子结构域,这归因于赖氨酸和精氨酸(有时是组氨酸)残基的存在,这导致AMPs通常带有净正电荷。此外,AMPs通常含有肽序列中的疏水残基(典型为50%)。阳离子残基和疏水残基之间的平衡决定了两亲性,这使得AMPs具有极性和非极性两端。
宿主源性AMPs通过破坏细菌膜和隔离必需营养物质来调控微生物群和防御致病菌感染。在胃肠道中,宿主来源的AMPs主要来源于肠上皮细胞或潘氏细胞,它们是位于小肠隐窝中的特化上皮细胞,与所有不同谱系肠道上皮细胞起源的干细胞非常接近。因此,宿主肠上皮的紊乱直接影响AMPs的分泌和肠道菌群的稳态。
宿主来源的AMPs通过与细菌膜成分互作直接调节肠道微生物。主要方式是捕获或杀死病原体。这类AMPs包括防御素、凝集素、核糖核酸酶(RNAse)、盲肠毒素和溶菌酶。宿主利用sIgA调节肠道微生物区系主要是为了增加共生菌的生态位竞争力,而AMP则提高了对病原菌的抵抗力。最近的几项研究发现,AMPs表达的缺陷将导致宿主对肠道病原菌感染的易感性增加,因为肠道微生物区系更容易受到干扰。同时,不同AMPs所针对的病原体也是不同的,主要包括:(1)防御素(图1B-(1)):防御素由潘氏细胞产生,缺乏成熟β-防御素的小鼠对Salmonella Typhimurium(S.Typhimurium)口服挑战的易感性增加。防御素也可以形成纳米网络来捕获S.Typhimurium。(2)凝集素(图1B-(2)):抗菌凝集素可在革兰氏阳性菌膜上形成六聚体孔,阻止细菌到达肠道黏液层。此外,再生(Reg)蛋白家族是一组可溶性的凝集素,Reg蛋白表达的缺陷增加了宿主对Listeria monocytogenes(L.monocytogenes)、Enterococcus和Yersinia Pseudotuberculosis(Y.Pseudotuberculosis)肠道感染的易感性,但这类蛋白的表达反而可以延长S.Typhimurium的感染。(3)核糖核酸酶(图1B-(3)):核糖核酸酶在体外对革兰氏阳性和革兰氏阴性病原体均表现出活性。在体内,T细胞受体δ(TCRδ)-/-小鼠表达的核糖核酸酶血管生成素4(Ang4)水平显著降低,这导致口服S.Typhimurium攻毒后,Ang4的产生受到抑制。(4)Cathelicidins(图1B-(4)):新生小鼠的Cathelicidins相关AMP(CRAMP)表达缺陷将导致大肠杆菌(E. coli)在体内过度繁殖,并增加成年后胰腺的自身免疫反应和糖尿病易感性。(5)溶菌酶(图1B-(5)):溶菌酶由潘氏细胞产生,在S.Typhimurium感染期间,潘氏细胞可以重组溶菌酶的产生以保持其抗菌能力。
AMPs调节肠道菌群的间接机制则包括必需营养素的隔离。例如,铁载体是大肠杆菌分泌的一类铁结合化合物,它可以与宿主的铁结合蛋白竞争铁离子,为病原体的生长提供必需的营养物质。(6)脂质运载蛋白-2(图1B-(6)):为了对抗细菌铁载体,宿主细胞可以产生脂质运载蛋白-2来隔离铁载体。
除了病原体的易感性外,宿主源性抗菌肽还可以改善家畜的生产性能。最近的一项研究发现了一个核心瘤胃上皮基因(DEFB1),它属于β-防御素家族。有趣的是,DEFB1的抑菌活性与莫能菌素(一种提高饲料转化效率的添加剂)相似。
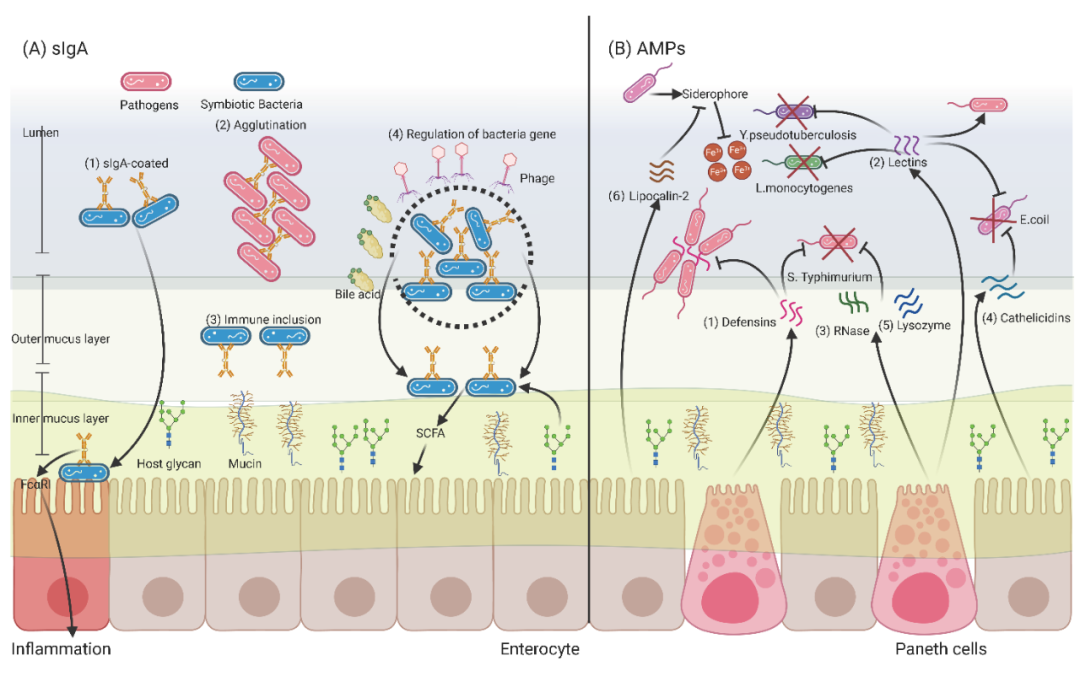
图1. 调控肠道菌群的宿主源免疫分子(sIgA和AMP)
细胞因子
免疫细胞因子可分为促炎细胞因子、抗炎/促溶解和趋化细胞因子。白细胞介素(IL)-1β、IL-18、IL-6、肿瘤坏死因子(TNF)、干扰素(IFN)、IL-33、IL-17、IL-21和IL-13为促炎细胞因子,IL-10、IL-22、IL-4和转化生长因子-β(TGF-β)为抗炎细胞因子。这些促炎和抗炎细胞因子与肠道菌群之间的关系已被系统地讨论和回顾。大多数细胞因子是分子量小于25KD的单链形式的糖蛋白。本文主要就细胞因子在影响肠道菌群中的作用进行综述。例如,IL-1β可激活IL-1R 2型,进而抑制结肠内AMPs的产生,并触发促炎肠道菌群的生长(图2-(1))。作为IL-1细胞因子家族的一员,高水平的IL-18与AMPs的产生相关,这表明其在调节肠道菌群中的作用(图2-(2))。此外,大多数抗肿瘤坏死因的作用已被证明可以部分通过调节肠道菌群组成来改善炎症。例如,抑制TNF-α的产生可降低厚壁菌门与拟杆菌门的比例(图2-(3))。研究还发现,IFN-β可以调节多发性硬化患者肠道菌群的组成,并增加Prevotella copri(P.copri)(图2-(4))的丰度,这提示IFN-β也可能参与调节肠道菌群。
肠上皮细胞上缺乏IFN受体(IFNAR1+-/-)的小鼠在结肠炎实验模型中表现出肠道微生物群组成的显著改变和对肿瘤发展的更强的易感性。此外,最近发现IL-33是肠道菌群的重要调节因子,因为IL-33缺陷小鼠的结肠免疫球蛋白A(IgA)减少(图2-(5))。IL-21也可影响IgA生产调节B细胞分化(图2 -(6))。以上细胞因子的功能间接影响肠道菌群的调节,因为IgA负责控制肠道共生菌和致病菌的水平,并参与肠上皮稳态。同时,已证实了共生微生物能够调节IL-17和IL-22的表达(图2-(7)),进而刺激维持黏膜完整性的固有机制,如AMPs和粘蛋白的产生和组织修复,进而参与肠道菌群的调节。此外,研究发现TGF-β信号通过增强IgA分泌进入管腔,有助于控制肠道菌群组成和维持肠道屏障完整性(图2-(8))。综上所述,上述结果证明肠道细胞因子可通过不同途径参与肠道菌群组成的调控,而细胞因子参与肠道菌群组成调控的潜在差异途径更多,有待进一步研究。
CD4+效应T细胞和先天淋巴样细胞
此外,CD4+效应T细胞(cluster of differentiation 4+,CD4+)和先天淋巴细胞(innate lymphoid cells)在调节肠道菌群中的作用受到了广泛关注。在粘膜组织中存在着多种功能独特的CD4+ T细胞,即Forkhead box protein P3(Foxp3)调节性T细胞(Treg)、辅助T细胞(Th)1、Th2、Th17和滤泡辅助T细胞(Tfh)。其中,Th1、Th2和Th17是CD4+效应T细胞的主要亚群,参与调控肠道菌群。在启动免疫系统对微生物抗原的反应后,IFN-γ和IL-12启动Th1细胞的分化(图2-(9)),加速病原体的去除,IL-4启动Th2细胞的分化(图2-(10)),帮助B细胞产生抗体,这是调节肠道菌群的关键。值得注意的是,IFN-γ和IL-4在不同水平上相互拮抗,因此Th1和Th2的发育被认为是互斥的。此外,IFN-γ还能促进肠上皮产生活性氧和氮,为某些肠道微生物提供生态位和营养物质(见3.4电子呼吸受体)。最新发现Th17细胞的亚群,通过分泌IL-17A、IL-17F和il - 22诱导肠上皮细胞(IECs)产生AMPs和紧密连接蛋白。同时,由于肠道微生物抗原的刺激,肠道Th17细胞与多种炎症性疾病相关。有趣的是,肠道Th17细胞在炎症反应中的作用取决于微生物抗原的类型。共生细菌诱导肠道驻留稳态Th17细胞表达IL-17和IL-22细胞因子(图2-(7)),后者维持非炎症和粘膜免疫屏障的状态(通过IECs刺激AMPs的分泌)。致病菌则诱导Th17细胞分泌促炎细胞因子,诱导炎症反应(图2-(11))。此外,Orphan nuclearreceptorγ+(RORγ+)Tregs是结肠Tregs的主要亚型,在调控免疫耐受和炎症反应稳态中发挥重要作用。在RORγ+ Tregs缺失的模型动物中观察到微生物代谢失调和炎症Th17细胞增加。因此,肠道Tregs的诱导似乎是宿主-微生物免疫适应的一个标志。
先天淋巴样细胞(ILCs)是最近发现的先天细胞类型,由DNA结合抑制剂(Id2)依赖的淋巴样祖细胞发育而来,并与分化的T细胞共享功能特征。ILCs可以通过产生细胞因子来调节肠道菌群。关于肠道CD4+ T细胞室如何对菌群组成的变化做出反应,以及CD4+ T细胞亚群可塑性是否在适应菌群变化中发挥作用仍在进一步与研究中。微生物定植影响肠道中存在的各种免疫细胞。因此,在组织微环境中存在复杂的相互作用,一种细胞类型分泌的细胞因子进一步影响其他细胞类型的效应功能,反过来,免疫抗原也可以反馈并影响肠道菌群。
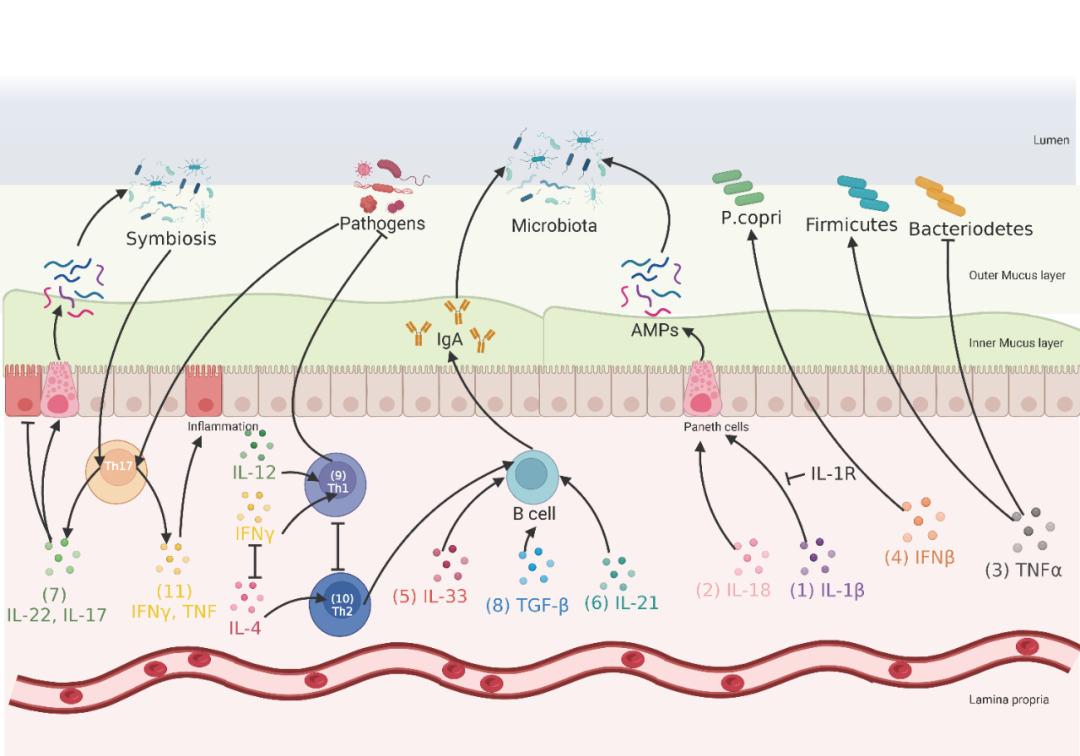
图2. 调控肠道菌群的宿主源免疫分子(细胞因子)
肠源性黏膜分子
来自黏液层的宿主源营养物质和电子呼吸受体在粘膜层上为微生物生长提供了生态位。因此,共生菌与病原体对生态位的竞争决定了宿主对病原体感染的抵抗力。
碳源
在单胃动物中,膳食糖在小肠中被利用,这意味着定殖后肠的微生物只以宿主聚糖和膳食纤维作为碳源。值得注意的是,反刍动物利用前胃发酵膳食纤维,这凸显了宿主聚糖对肠道菌群调节的重要性。在这里,我们主要讨论宿主来源的碳源(宿主聚糖),如岩藻糖、半乳糖、唾液酸等。这些碳源都来源于肠上皮细胞的黏液层。其中岩藻糖为己糖,在第6个碳原子上缺少羟基,疏水性较强。唾液酸是九碳糖的衍生物,一般位于糖蛋白和糖脂类等缀合物的末端。抗生素治疗可以增加宿主来源的游离唾液酸。Bacteroides thetaotaomicron(B.thetaiotaomicron)可以将粘膜上的唾液酸分解进入肠腔变为游离唾液酸,但是B.thetaiotaomicron本身不能吸收利用唾液酸。游离唾液酸为病原体生长提供碳源,导致宿主对感染易感性增加(图3-(1))。宿主来源的碳源与宿主基因型密切相关。ABO血型的N-乙酰-半乳糖胺(GalNAc)转移酶基因的2.3 kb缺失降低了GalNAc的浓度,从而降低了在肠道中利用GalNAc作为碳源的Erysipelotrichaceae 的丰度(图3-(2))。另一方面,宿主也有策略地避免病原体对生态位的利用。例如,免疫调节部分讨论了sIgA增加共生菌多糖利用活性,从而增加有益菌的黏膜生态位竞争力(“免疫”部分,图1A-4)。此外,改变宿主聚糖的分子结构是另一种策略。拟杆菌也可将岩藻糖从粘膜层释放到肠腔内,这将增加宿主对S.Typhimurium 感染的易感性。有趣的是,由活化T细胞分泌的白细胞介素-22(IL-22)是宿主固有的防御机制之一,它刺激半乳糖苷2-α-L-岩藻糖基转移酶2(FUT2)和α(1,2)- 岩藻糖基化(糖基化)的上皮表达,而不被B.thetaiotaomicron 在黏液层切割,因此不被S.Typhimurium 用作碳源(图3-(3))。
氮源
宿主的粘蛋白层,包括跨膜粘蛋白和形成凝胶的粘蛋白,这也是一些肠道微生物的重要氮源(图3-(4))。在静脉注射稳定同位素标记的苏氨酸(13C-和15N标记)后,在Bacteroides acidifaciens(B.acidifaciens)和Akkermansia muciniphila(A. muciniphila)的中检测到大量的13C和15N标记,这些微生物被证明是利用宿主来源的粘蛋白的主要采集者。A.muciniphila 是疣微菌门中一种降解粘蛋白的细菌,其丰度已被证明与几种疾病状态呈负相关,包括炎症性肠病(IBD)、急性阑尾炎和肥胖。A.muciniphila 在肠道黏膜层具有独特的生态位优势,不仅增强了先天免疫反应和肠道屏障功能,而且抑制了病原体在黏膜层的定植。
氧源
最近有人提出了氧代谢和氧屏障调控肠道菌群组成的概念。在IBD中,腔内氧水平的增加是由于上皮细胞被破坏,这导致血红蛋白所携带的氧气释放到微生物群定植的黏液层。氧含量的增加破坏了上皮细胞的厌氧环境。这进一步为兼性厌氧菌或潜在的需氧菌提供生态选择优势,这使致病菌繁殖时更具竞争力(图3-(5))。例如,在厌氧环境紊乱的情况下发现沙门氏菌等致病菌有氧扩张。重要的是,腔内氧水平的增加不仅是由于物理屏障的破坏,而且还由于厌氧糖酵解的增加减少了跨细胞途径的氧消耗,特别是在结肠上皮中。与小肠上皮更易利用的葡萄糖和谷氨酰胺不同,成熟的结肠上皮主要通过氧化丁酸等短链脂肪酸来产生能量,使粘膜表面缺氧。然而,如果结肠上皮细胞转换为更倾向于利用葡萄糖时,剩余的氧气可能扩散到肠腔,并最终导致兼性厌氧菌(如肠杆菌科)的扩张。事实上,新生儿在出生时肠道是有氧的。新生儿肠道中相对较高的氧气水平有利于兼性厌氧菌(如肠杆菌科、肠球菌和链球菌)的存活。这些早期的定殖者消耗可用的氧气,从而在肠道中创造厌氧微环境,并促进专性厌氧菌(如双歧杆菌、梭状芽胞杆菌、拟杆菌、细孔菌、真杆菌和瘤胃球菌等)的形成。所有这些证据都支持氧水平可调节宿主肠道菌群。
电子呼吸受体
另一种来源于肠道的代谢物是电子呼吸受体,它为兼性厌氧菌的有氧和厌氧呼吸提供了条件。肠道中氧含量的增加与炎症反应和抗生素治疗有关,不利于厌氧微环境。然而,厌氧硝酸盐呼吸和硫酸盐呼吸是病原体在肠道定植的常见策略。促炎细胞因子IFN-γ可激活肠上皮的Nos2基因产生活性氧和氮(如一氧化氮),具有抗菌作用,但会在管腔内形成硝酸盐。肠杆菌科的大多数成员表达硝酸盐还原酶基因,该基因将硝酸盐还原与用于厌氧呼吸的节能电子传递系统耦合,这一过程被称为硝酸盐呼吸(图3-(6))。有趣的是,致病性沙门氏菌不能利用生态位中上皮来源的硝酸盐,但可以利用吞噬细胞浸润产生的硝酸盐。值得注意的是,肠道内专性厌氧共生微生物不能表达还原酶基因,这为兼性厌氧病原体提供了生态位优势。此外,部分细菌可以粘附在肠细胞上,并借助一种被称为III型分泌系统(T3SS)的注射器状装置影响宿主细胞的功能。通过T3SS,细菌可以利用肠道来源的粘膜营养物质(如乙醇胺和岩藻糖),避免不必要的能量消耗。然而,在体内稳态和体外条件下,S.Typhimurium不能利用乙醇胺。抗生素链霉素可激活内源性硫化合物(硫代硫酸盐)氧化形成四硫酸盐,后者作为电子呼吸受体增强鼠伤寒链球菌对乙醇胺的利用能力(图3-(7)和3-(8))。虽然使用替代电子呼吸受体的能力可以为兼性厌氧菌的定植提供条件,但宿主也可以利用这种能力在稳态期间限制厌氧共生体与肠上皮的接触。柠檬酸杆菌种类(病原体)可以通过厌氧过氧化氢(H2O2)呼吸在未发炎的肠道中生长。而IECs的NADPH氧化酶1(Nox1)产生的H2O2可使粘液内层基本无细菌,从而避免粘液内层破坏和厌氧细菌在稳态期间渗透到隐窝(图3-(9))。据观察,电子呼吸受体对肠道菌群的调节作用取决于肠道的生理状态(病原体感染或稳态)。
综上所述,宿主来源的粘膜代谢相关源可以影响细菌在肠道中的组成。在讨论涉及肠道菌群调节宿主上皮因子时,常讨论连接紧密的上皮物理屏障和微绒毛,但其在菌群调节中的作用主要归结于物理屏障功能。值得注意的是,宿主紧密连接和微绒毛的破坏导致宿主肠上皮内暴露于环境中,从而通过肠源性粘膜代谢相关源(包括肠道碳、氮和氧源)影响肠道菌群。
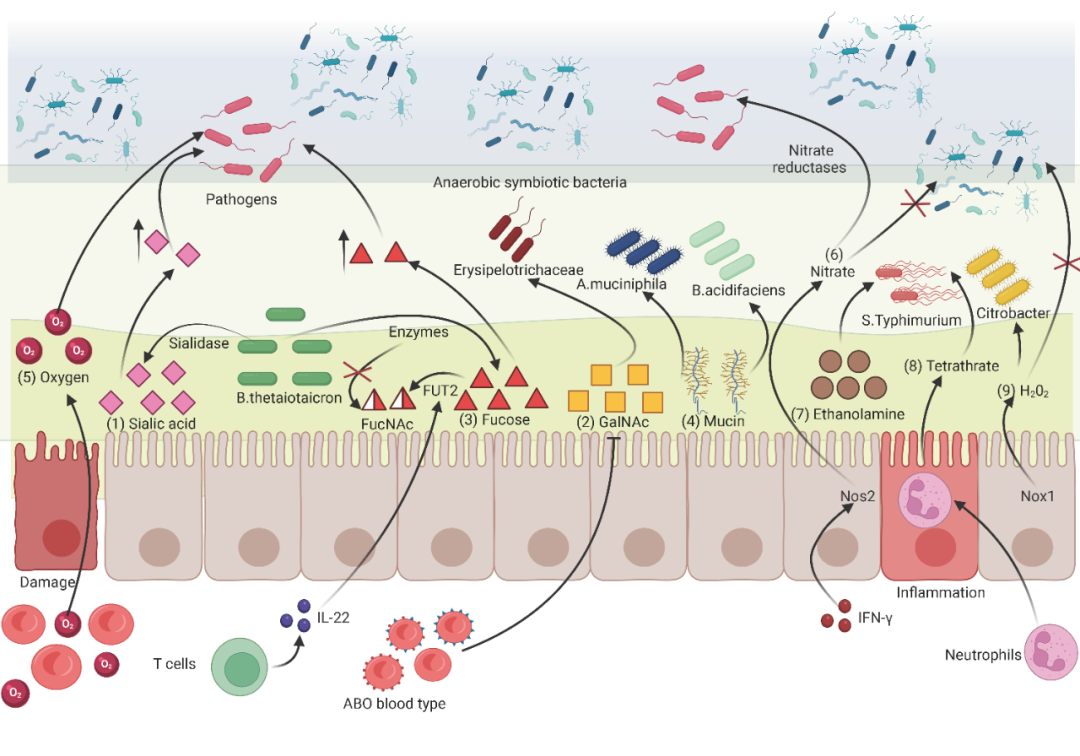
图3. 调控肠道菌群的宿主源粘膜分子
肠源性外泌体的<非编码RNA>
外泌体由几乎所有类型的细胞通过胞吐途径分泌,在宿主和肠道菌群之间建立串扰。外泌体的成分主要包括蛋白质、脂质、长链非编码RNA(lncRNA)、微小RNA(miRNA)和环状RNA(circRNA)。其中,外泌体蛋白质和脂质可能分别通过影响肠道中的碳源和氮源来影响肠道微生物群。因此,我们建议对宿主源性外泌体的蛋白质和脂类在调节肠道菌群中的作用进行更多的研究。相比之下,关于宿主源性外泌体的ncRNAs组分对肠道菌群影响的研究较多,因此本部分重点探讨了宿主源性外泌体ncRNAs组分对肠道菌群的影响。
miRNAs
影响腔内菌群生长的miRNAs主要来源于IECs和homeodomain-only protein homeobox(HOPX)阳性细胞。宿主源外泌体可携带miRNA,并作为载体实现原核和真核细胞间的跨界通讯。miRNAs可以通过不稳定的mRNA和抑制转录后调节蛋白的结合来调节细菌基因的表达。miRNAs通过细菌与外泌体的膜融合进入病原体(如 F. nucleatum 和 E.coli)后,特异性调控细菌基因转录物(图4-(1)和4-(2))。miRNA缺乏的小鼠表现出肠道菌群失调和结肠炎加剧,这可以通过粪便miRNA移植缓解。值得注意的是,高温处理的粪便移植也可具有治疗效果,因为miRNA是耐热的。宿主来源的miRNA除了能调控病原体外,还能调控共生细菌。在结肠炎耐受小鼠中,用硫酸右旋糖酐(DSS)刺激IECs分泌miR142a-3p,miR142a-3p通过与罗伊氏乳杆菌的特定靶点(位点标记LREU_RS06530[polA]和位点标记LREU_RS03575)结合,特异性促进罗伊氏乳杆菌(L.reuteri)的生长(图4-(3))。
circRNA
miRNAs和circRNAs都可以通过外泌体分泌,这对肠道微生物群和健康有潜在的影响。Zhu等报道无菌小鼠体内的微生物定殖(特异性无病原体微生物群或双歧杆菌)会影响血液和肺肿瘤微环境中circRNA的表达。具体而言,当无菌小鼠通过粪便移植接受SPF菌群或接种双歧杆菌时,血液中circ0000730的表达下调。靶点预测分析显示,circ0000730可以吸收miR-466i-3p和miR-466f-3p,这两种基因可以调节SOX9基因(一种发育调节因子和致癌基因)的功能。因此,作者提出circRNA-miRNA-mRNA轴(circ0000730-miR466i-3p-SOX9轴)抑制肺部肿瘤以响应肠道菌群。这一概念通过揭示调控circRNA之间的相互作用,增加了宿主-微生物互作复杂性的另一层面。Chen等利用过表达的circNF1-419腺相关病毒(adeno-associated virus,AAV)动物系统研究表明,大脑中circNF1-419过表达不仅会影响大脑胆碱能系统,还会改变新生小鼠肠道菌群组成、肠道稳态和生理以及肠道菌群轨迹。他们的发现证明了circRNA和肠道微生物组之间的联系,扩大了“微生物组-转录组”链接库,并提供了更多关于肠-脑轴的信息。迄今为止,关于circRNA和肠道菌群的研究非常有限。作为miRNA的负调控因子,需要对circRNA进行彻底的研究,以了解circRNA - miRNA -微生物组的相互作用(而不是仅仅关注circRNA -微生物组轴)。此外,同样重要的是了解circRNAs在不同组织和整体宿主健康中对其靶miRNAs表达的影响,以响应肠道微生物群,并了解circRNAs是否与宿主相关微生物组保持双向相互作用,类似于miRNAs。
lncRNA
除了miRNA和circRNA外,lncRNA是一组长度和结构各异的ncRNA分子。lncRNA具有高度的多样性,可以连接肠道菌群与肠外器官,如肠-胎盘轴、肠-肺轴、肠-脑轴。同时,用不同大肠杆菌菌株或粪便源菌群重新定植小鼠,可以通过IEC的lncRNA表达谱进行区分。因此,lncRNA显示出了与肠道菌群相互作用的可能性。然而,进一步研究lncRNA在微生物调控中的作用是非常重要的。
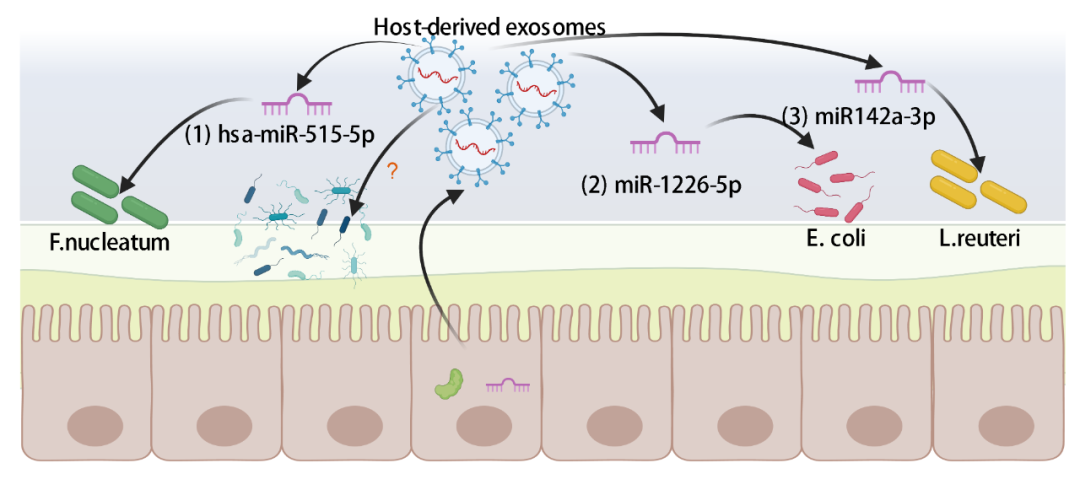
图4. 调控肠道菌群的肠道源外泌体的<ncRNAs>
来自除肠道外的其他器官的分子
性激素 (性腺)
性腺主要通过分泌类固醇激素(性类固醇)来调节宿主的生理活动,主要包括雌激素和雄激素,它们与疾病的二态性密切相关。
雌激素
在哺乳动物中,雌激素主要由卵巢和胎盘分泌,包括雌二醇(17β-雌二醇)、雌酮、雌二醇(16-羟雌二醇)和雌二醇(15α-羟雌二醇)。被称为雌激素组的肠道微生物可以产生β-葡萄糖醛酸酶,将结合的雌激素解耦为活性形式。有综述指出,雌激素组通过调节雌激素水平与多种疾病(如阿尔茨海默病、骨质疏松症、子宫内膜异位症、多囊卵巢综合征和癌症)有关。特别是女性在绝经后更容易患这些疾病,这凸显了雌激素对这些疾病的调节作用与肠道菌群的作用是分不开的。然而,从肠道菌群影响雌激素水平的角度来看,这可能是片面的。最近的一些研究发现,雌激素还可以通过调节微生物组成来影响女性对疾病的易感性。
调节肠道菌群的雌激素需要与肠组织中的雌激素受体结合,其中最经典的是雌激素受体α(ERα)和雌激素受体β(ERβ)。雌激素通过ERα和ERβ对肠道菌群具有不同的调节作用。敲除ERα的小鼠对DSS诱导的结肠炎具有更大的耐受性,提示ERα可激活肠内促炎级联反应(图5A-(1))。值得注意的是,炎症反应可增加病原体定植的可能性和宿主对疾病的易感性(见肠道源粘膜分子部分)。相反,ERβ可以调节肠上皮细胞的分化、紧密连接的形成和通透性。肝幽门螺杆菌(H.hepatcus)是一种引起肠道炎症的小鼠肠道病原体。在H.hepatcus感染的小鼠中,通过ERβ而非ERα发出信号,可显著减少盲肠炎症以及炎症细胞因子和趋化因子的表达。因此,宿主对肠道炎症的易感性可能与肠上皮中雌激素受体的类型有关。事实上,研究发现雌激素可以通过与ERβ结合来调节肠道微生物群。在相同的复合营养素饮食条件下,与ERβ+/+相比,ERβ-/-小鼠的变形菌门丰度显著增加。另一项研究也显示,与ERβ+/+小鼠相比,DSS诱导后ERβ-/-小鼠肠道菌群α-多样性下降幅度更大(图5A-(2))。
G蛋白偶联受体30(GPR30)也是雌激素的特异性受体之一,它不同于ERα和ERβ,存在于细胞膜上。GPR30可增加结肠运动,并可对抗炎症(图5A-(3))。因此,这些数据提示雌激素可以通过其受体维持肠道菌群的稳定,提高宿主对致病性感染的耐受性。
雌激素与感染易感性和肠道菌群密切相关。此外,雌激素可以通过影响IECs的代谢来调节肠道微生物群。一项研究发现,卵巢切除小鼠肠道中变形菌门和脂多糖(LPS)明显增多,这可能是由于雌激素刺激小肠中碱性磷酸酶的产生。另一项研究通过比较雄性和雌性小鼠的结肠转录组,确定了AMP基因表达的差异。有趣的是,一些AMPs的启动子区域包含雌激素反应元件。以上研究表明,雌激素- ER - AMPs -肠道菌群是宿主调节肠道菌群的途径之一(图5A-(4))。
雄激素
与女性相比,男性患代谢性疾病的患病率更高。例如,雄性小鼠的胰岛素敏感性低于雌性小鼠,并且更容易发生高脂饮食诱导(HFD-induced)肥胖和代谢综合征。除雌激素外,该疾病的性别二态性可能由雄激素驱动。雄激素主要包括雄烯二酮和睾酮,它们主要在睾丸中合成,而女性则在肾上腺皮质和卵巢中合成。在男性和女性的性腺、肾上腺皮质和肠道等非内分泌组织中,循环中的雄激素可通过芳香化酶转化为雌激素。
有趣的是,与雌激素不同,雄激素似乎在肠道菌群引起的代谢性疾病中表现出混合的结果。例如,成年雌性小鼠在新生儿雄激素暴露后肠道微生物多样性较低,厚壁菌门与拟杆菌门比率较高。最近的一项研究揭示了雄激素影响肠道微生物代谢的机制。除胰岛素敏感性增加外,阉割雄性小鼠的肠道菌群与雌性小鼠更接近。此外,雄激素降低了谷氨酰胺的循环,这可能会增加体外胰岛素敏感性。值得注意的是,雄激素对谷氨酰胺的作用部分取决于肠道菌群(图5B-(1))。然而,雄激素对肠道菌群的影响并不全是负面的。在非肥胖糖尿病(NOD)雌性小鼠中,结肠CRAMP的表达低于NOD雄性小鼠,这表明结肠CRAMP对糖尿病具有保护作用,因为雄性NOD小鼠对该疾病具有部分保护作用(图5B-(2))。因此,雄激素对肠道菌群的影响可能取决于特定的条件(如年龄和疾病类型)。这一点值得相关领域的研究人员关注。
神经激素(神经系统)
神经激素由神经内分泌细胞分泌,以响应神经元的输入。虽然它们被分泌到血液中以产生全身作用,但它们也可以作为神经递质。同时,神经激素也是调节肠道菌群的重要宿主源分子。它们对肠道菌群的作用主要是调节细菌的生长和基因表达。例如,儿茶酚胺可以改变细菌(如葡萄球菌)的生长、活力、生物膜形成或毒性。在宿主肾上腺素的作用下,沙门氏菌下调了对宿主抗菌肽的抗性,并诱导关键的金属离子转运系统,从而影响细胞内的氧化应激平衡。可以看出,与其他宿主源分子相比,神经激素主要直接作用于微生物,而不是通过宿主,Neumann等(2015)系统地综述了神经激素对肠道菌群的影响。
胆汁酸(肝脏)
胆酸(CA)和鹅去氧胆酸(CDCA)由肝脏以胆固醇为原料产生,然后肝脏将胆汁酸(BA)与甘氨酸或牛磺酸结合,经过两步反应产生初级BA。初级BA进入小肠后,在肠道菌群解偶联作用(去除甘氨酸或牛磺酸偶联物)下形成次级胆汁酸,其中95%的胆汁酸从肠道被重新吸收回肝脏,这个过程称为BA的肠肝循环。由此可见,肠道菌群在BA循环浓度中起着重要作用,BA循环浓度与脂质代谢密切相关。然而,肠道菌群与BA之间的相互作用不是单向的。BA一方面促进BA利用菌的生长,但通过破坏膜结构抑制BA敏感菌的生长(图5C-(1))。另一方面,BA可以与IECs相互作用,形成肠道菌群。BA可以诱导抗菌剂(如RNAse和一氧化氮)的转录,这些抗菌剂通过小肠的法尼氏X受体(FXR)影响肠道菌群(图5C-(2))。事实上,一项研究发现,BAs对IECs的免疫功能有深远的影响,从而进一步调节肠道菌群。石胆酸的衍生物(3-氧代石胆酸,LCA)和isoalloLCA可视为T细胞调节剂,其中3-氧代石胆酸可抑制Th17细胞分化,isoalloLCA可促进Treg分化。值得注意的是,Th17细胞的分化可调控肠道菌群(见关于CD4+效应T细胞和先天淋巴样细胞(ILCs)一节)。此外,肠道BA池可通过BA核受体(维生素D受体)调节结肠Treg稳态,改善宿主对炎性结肠炎的易感性(图5C-(3))。由此可见,肠道BA池在Treg细胞稳态中起着重要作用,进而影响宿主对感染的易感性。
乳酸(肌肉)
对于宿主来说,绝大多数乳酸代谢处理发生在肝脏中。一项研究利用13C3乳酸注射到小鼠体内后发现,来自肌肉运动的血清乳酸可以通过上皮屏障进入肠腔。有趣的是,优秀运动员长期的高乳酸环境可以为Veillonella atypica(V. atypica)的生长创造一个肠道优势生态位。V. atypica可将乳酸代谢为丙酸,在运动过程中为机体提供更多能量(图5D)。因此,肌肉运动产生的乳酸可能是宿主调节肠道菌群的潜在分子。
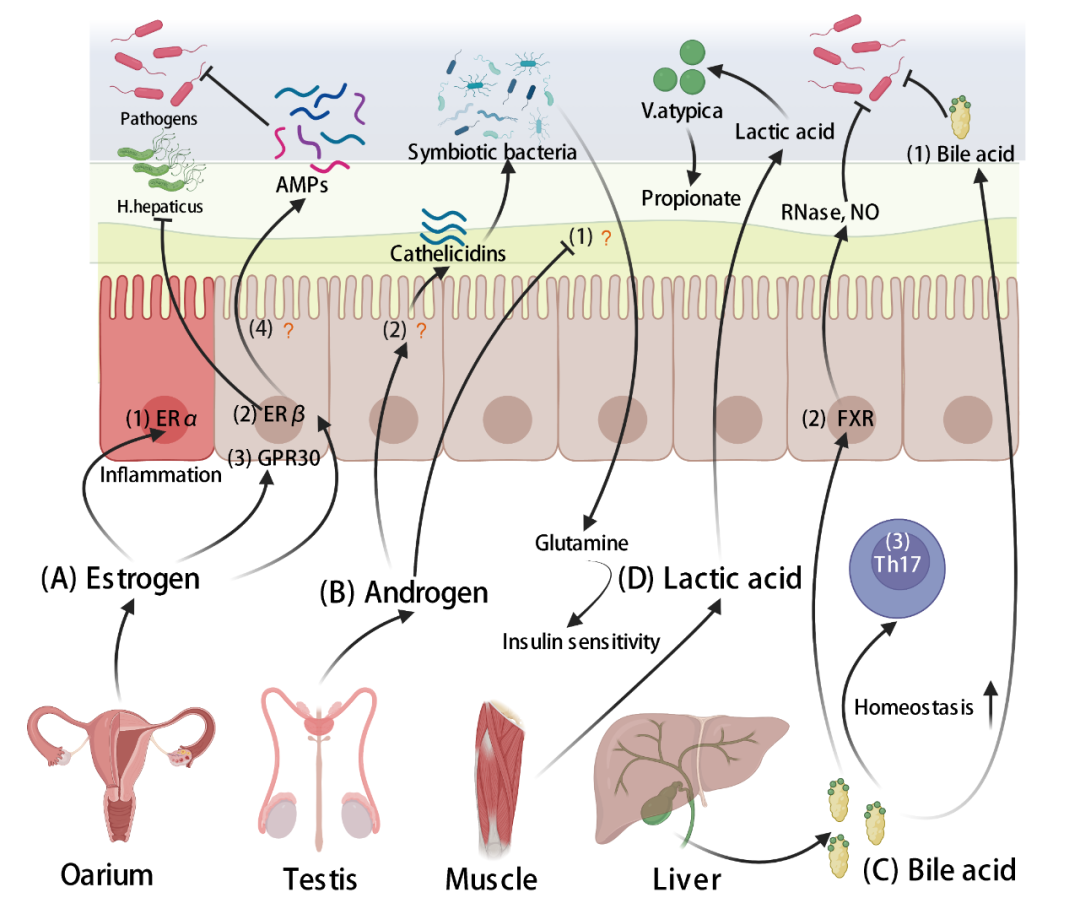
图5. 调控肠道菌群的来自肠道以外的其他器官分子
不同宿主衍生分子之间的相互作用
宿主和肠道菌群形成复杂的网络,重要的是要考虑各种宿主衍生分子之间的关系。如IL-33、TGF-β、IL-21和IL-4可通过刺激B细胞增加IgA的分泌;IL-22、IL-17、IL-18和IL-1β可通过刺激潘氏细胞触发AMPs的分泌。IL-22可刺激FUT2的分泌,促进岩藻糖的糖基化,防止岩藻糖被病原体利用。此外,生殖器官的雌激素和雄激素可能影响潘氏细胞的AMPs分泌。同时,宿主源分子对肠道菌群的调控与IECs的功能密切相关。大部分分子通过IEC分泌的IgA和AMPs调控肠道菌群,是宿主和肠道菌群相互作用的重要中介。
宿主调控肠道菌群的临床见解
扩展关于宿主因素如何塑造肠道菌群的知识,可为个性化调控肠道菌群提供更多机会,在临床和行业中具有巨大的应用潜力。
一般认为,肠道菌群失调(微生物多样性和丰富度降低)增加了感染的易感性。此外,肠道菌群失调与肠道微环境变化密切相关,而肠道微环境变化是由宿主源分子建立并受其影响。例如,肝硬化患者腔内IgA分泌的减少可增加菌血症和自发性细菌性腹膜炎的易感性。此外,结肠RegIIIγ表达降低也与DSS诱导的结肠炎易感性增加有关。对于非肠源性代谢分子,体内雌激素水平降低导致肠道菌群失调,增加代谢综合征易感性。因此,确定维持肠道微生物群稳态的宿主因素为降低病原体易感性等疾病的风险提供了额外的解决方案。
粪便菌群移植(FMT)是一种将整个肠道菌群转移到受者体内的新方法,目的是恢复肠道菌群稳态并治疗与菌群失调相关的疾病。一项荟萃分析研究表明,FMT治疗艰难梭菌感染(CDI)的治愈率高于常规治疗。然而,FMT治疗CDI患者的失败率为5.2%(结肠镜检查)- 21.9%(鼻胃管检查),这给FMT的应用蒙上了阴影。除FMT的剂量和频率外、给药方法,供体和受体的宿主因素也至关重要。一项研究表明,FMT的治疗效果由供体携带的宿主源性分子(如miRNA)反映。确定来自供体的治疗作用的成分将减少治疗的不良结果。此外,受体肠道生态位中的宿主源性分子也可以决定来自供体的粪便微生物群是否定植和发挥作用。
工作展望
在这篇综述中,我们详细总结了与宿主遗传学(基因型)密切相关的、塑造肠道菌群的宿主源分子。mGWAS可用于识别与菌群定殖相关的一系列基因。此外,双向孟德尔随机化(MR)可以利用通过mGWAS获得的宿主遗传变异信息作为工具,探索宿主基因组与肠道菌群之间的因果关系。更多像mGWAS这样的工具为研究人员或临床医生提供了支持性信息,以预测可能在宿主遗传控制下调控肠道微生物群的宿主源分子。当宿主遗传与肠道菌群的相关性被建立和验证后,研究者或临床医生就可以根据宿主基因型来评估定植在其中的肠道菌群。克罗恩病可根据宿主基因型和肠道微生物组成分为不同的疾病亚型。面对不同的疾病亚型,临床医生应采取个性化治疗,以最大限度地提高治疗成功率。值得注意的是,需要进行精心设计的临床队列研究,特别是大数量和纵向样本,以评估基于宿主基因型和肠道微生物组成的特殊微生物治疗对不同疾病亚型诊断的有效性。
为了研究宿主源分子与肠道菌群之间的因果关系,需要适当的实验模型。近年来,类器官芯片模型的发展为考虑多种细胞类型和肠道菌群提供了解决方案。例如,微生物、益生菌或侵入性细菌的共培养可以提供一个无菌环境,在这种环境中,微生物组和免疫因素可以被包括进来,以看到多细胞交叉作用。而且,在同一系统中,宿主细胞和细菌可以在不同的氧气和营养浓度下培养,满足了细胞和细菌的不同需求。然而,进一步的机制和体外模型研究将是必要的,这对该领域具有重要意义。
肠道菌群恢复力被定义为肠道菌群从饮食变化、病原体感染、炎症反应、抗生素治疗等外部应激因素中恢复的能力。研究人员利用稳定性景观框架模拟抗生素对肠道菌群的干扰以及随后肠道菌群的恢复。一项基于1000名西方成年人的研究表明,健康细菌的丰度受到衰老和超重等宿主因素的影响,而不是短期的饮食干预。在未来的研究中,需要探索影响肠道菌群时间动态的宿主因子,特别是受外界因素干扰的宿主因子。
总 结
本文就宿主因子调控肠道菌群的分子机制进行综述。虽然大量研究已经将宿主对疾病的易感性与肠道菌群和饮食行为联系起来,但宿主影响某些疾病肠道菌群的确切机制尚未完全破译。了解宿主因子如何调节肠道微生物群并影响疾病发病率可能对开发新的预防和治疗干预措施,甚至预测个体的疾病易感性很重要。应当指出,本综述中提到的机制并不单独存在;这些机制之间存在着上下游调控关系。因此,面对肠道菌群紊乱的科学问题,研究人员需要关注肠道组织的遗传差异,而不仅仅是肠道菌群。迄今为止,我们对宿主遗传学在调节肠道菌群方面的理解主要基于体外研究和啮齿动物模型。为此,必须进行精心设计的临床试验或收集临床数据,以填补临床和实验知识之间的巨大空白,并将从动物模型中获得的概念证明转化为临床环境。因此,这些研究有可能应用于益生菌、益生元,并在未来应用于有效治疗肠道菌群紊乱相关疾病。
引文格式:
Chenguang Zhang, Huifeng Liu, Lei Sun, Yue Wang, Xiaodong Chen, Juan Du, Åsa Sjöling, Junhu Yao, Shengru Wu. 2023. An overview of host-derived molecules that interact with gut microbiota. iMeta e88. https://doi.org/10.1002/imt2.88
作者简介

张晨光(第一作者)
● 西北农林科技大学2021级博士研究生
● 目前研究方向为反刍动物瘤胃微生物与宿主互作机制,相关学术成果已发表于iMeta、Journal of Dairy Science等期刊

武圣儒(通讯作者)
● 瑞典卡罗琳斯卡医学院微生物博士后,西北农林科技大学动物科技学院副教授,硕士生导师
● 现围绕反刍动物胃肠道健康调控与碳水化合物高效利用的宿主-微生物互作机制开展研究。近5年来以第一作者及通讯作者(含并列)在iMeta、npj Biofilms and Microbiomes、Proceeding B、JCI insight、Journal of Animal Science and Biotechnology 和 Animal nutrition 等杂志发表中科院大类一区论文13篇,Top期刊论文20余篇。荣获中国畜牧兽医学会奖、中国畜牧兽医学会动物营养学分会青年学者论坛优秀奖等荣誉。

杜鹃(通讯作者)
● 典卡罗林斯卡医学院副教授,博士生导师
● 主要研究微生物菌群对包括肿瘤在内的疾病的发生发展的相互作用。团队建立了国际上领先的微生物菌群临床和基础实验研究平台。以包括第一作者及通讯作者身份在内发表相关文章40篇,荣获瑞典国家研究委员会,瑞典国家委员会/中国国家自然科学基金委,瑞典战略研究基金会, 欧洲Eurostars等23项科研项目。并参与微生物菌群转化医学中心和华大基因合作的”百万微生物”项目

Åsa Sjöling (通讯作者)
● 哥德堡大学和瑞典卡罗林斯卡医学院教授,博士生导师
● 围绕微生物基因组与微生物互作开展工作。H指数24,i10指数45;在Nature Microbiology、npj Biofilms and Microbiomes 和 iMeta 等杂志发表SCI论文112篇,累计引用3315次。

姚军虎(通讯作者)
● 西北农林科技大学教授,博士生导师,国务院第七、八届畜牧学科评议组成员,中国畜牧兽医学会动物营养学分会副会长,中国动物营养指导委员会委员,第四届全国畜牧学科高峰论坛秘书长,陕西省饲料饲草产业体系首席科学家,西北农林科技大学动物营养与饲料学科学术带头人
●以奶畜能量高效利用的营养调控原理为主要科学问题,围绕瘤胃健康与高效发酵、小肠淀粉消化调控、肝脏能量利用、乳腺生物学等开展系统研究,系统揭示了奶畜碳水化合物代谢机理,建立了碳水化合物平衡指数体系(CBI)和能量高效利用一体化调控技术。
更多推荐
(▼ 点击跳转)
iMeta | 德国国家肿瘤中心顾祖光发表复杂热图(ComplexHeatmap)可视化方法
iMeta | 浙大倪艳组MetOrigin实现代谢物溯源和肠道微生物组与代谢组整合分析

第1卷第1期

第1卷第2期

第1卷第3期

第1卷第4期
期刊简介
“iMeta” 是由威立、肠菌分会和本领域数百位华人科学家合作出版的开放获取期刊,主编由中科院微生物所刘双江研究员和荷兰格罗宁根大学傅静远教授担任。目的是发表原创研究、方法和综述以促进宏基因组学、微生物组和生物信息学发展。目标是发表前10%(IF > 15)的高影响力论文。期刊特色包括视频投稿、可重复分析、图片打磨、青年编委、前3年免出版费、50万用户的社交媒体宣传等。2022年2月正式创刊发行!
联系我们
iMeta主页:http://www.imeta.science
出版社:https://onlinelibrary.wiley.com/journal/2770596x
投稿:https://mc.manuscriptcentral.com/imeta
邮箱:office@imeta.science
往期精品(点击图片直达文字对应教程)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|




























后台回复“生信宝典福利第一波”或点击阅读原文获取教程合集




















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









