







2024年2月12日,中医科学院黄璐琦、崔光红课题组联合中山大学巫瑞波课题组在ACS Catalysis上发表了题为 “From Functional Plasticity of Two Diterpene Synthases (IrTPS2/IrKSL3a) to Enzyme Evolution”(从两种二萜合酶(IrTPS2/IrKSL3a)的功能可塑性到酶的进化)的文章。
第一作者:Baolong Jin,Kangwei Xu,Juan Guo
通讯作者:Guanghong Cui,Ruibo Wu,Luqi Huang
DOI:doi.org/10.1021/acscatal.3c05918

1
01
背景
萜类化合物具有多种多样的结构和生物活性,其复杂性很大程度上源于萜烯合酶(terpene synthases,TPS)。这些酶可催化碳正离子的级联反应,然而其中负责淬灭最终正离子的基团在很大程度上是未知的。冬凌草Isodon rubescens来源的 IrKSL3a 和 IrTPS2 具有 98% 的序列同源性,并且都以(+)-CPP为底物,但不同的猝灭策略使其产生不同的产物,其中 IrKSL3a 通过去质子化直接生成Isopimaradiene(1),而 IrTPS2则是通过加水生成羟基化的Nezukol (2) (Figure 1b)。
1
02
研究结果
一、IrKSL3a 和 IrTPS2的特异性由一个关键氨基酸残基决定2
序列比对显示在活性口袋附近IrKSL3a 与 IrTPS2之间仅有5个残基的差异(Figure 1c),作者将IrTPS2五个位点的残基一一替换为IrKSL3a的残基。同先前在其他同源蛋白中得到的结果相同,IrTPS2:A523I突变体几乎完全产生1,而其余四种突变则未影响产物选择。然而研究人员用与异亮氨酸大小相似的亮氨酸进行同样的突变,IrTPS2:A523L 却继续产生羟基化产物 2(Figure 2a)。
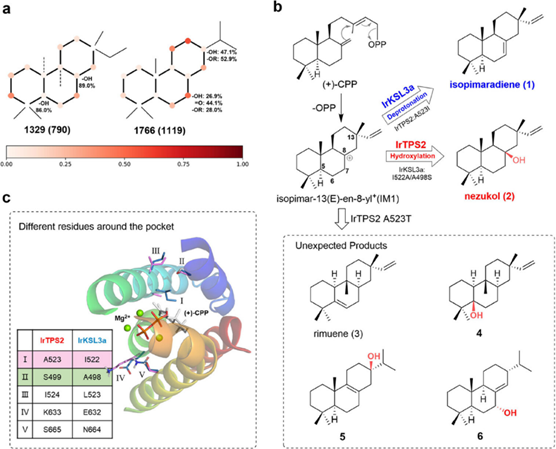

二、酶中存在对邻近的 β-甲基敏感的水结合二联体
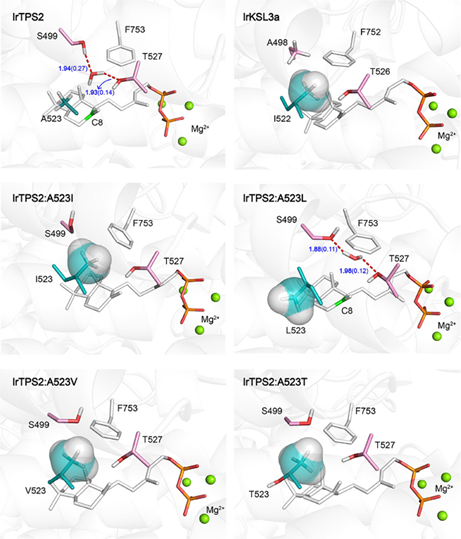
为了进一步研究酶结构与功能的关系,作者将底物 (+)-CPP与三个二价镁 (Mg2+) 辅因子一起对接到建模的野生型和突变体中,并进行了分子动力学 (MD) 模拟和QM/MM优化。野生型IrTPS2的T527 和 S499 均能与C8附近 (∼4Å)的水分子形成氢键,作者推测该水分子可能是羟基供体(Figure 3)。
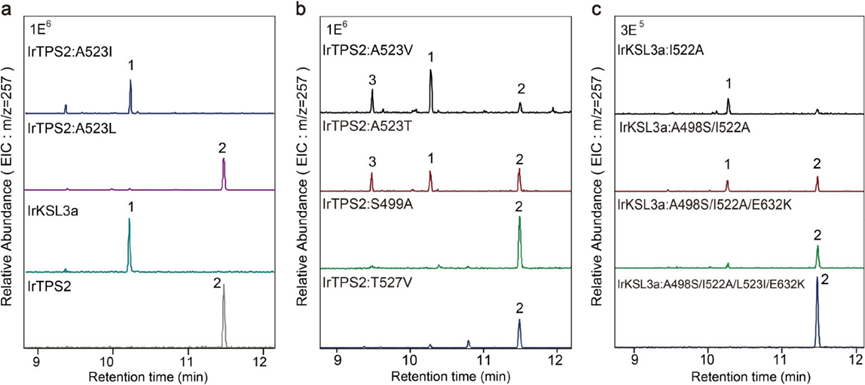

尽管侧链中甲基的位置仅有轻微差异,但模拟显示IrTPS2:A523I 与 IrTPS2:A523L 的结构具有非常显著的差异。IrTPS2:A523I 中的水位点被 I523 的 β-甲基占据,阻止了水分子淬灭 C8 碳正离子中间体。相比之下 IrTPS2:A523L 中的亮氨酸远离底物,水分子仍可与 T527 和 S499 结合(Figure 3),模拟结果与 C8 附近水分子的存在与否决定是否形成羟基化产物的假设一致。为了进一步研究 β-甲基的普遍影响,作者通过相同的计算策略发现 IrTPS2 的 A523T 和A523V 突变体也能阻碍水分子与 T527 和 S499 的结合。两突变体的功能验证进一步证实了计算结果(Figure 2b)。
虽然A523I突变对IrTPS2有影响,但IrKSL3a(I522A)的逆向突变仅产生极少量的产物2,同时产物1的量大大减少,其功能并没有完全逆转。作者假设这是由于 IrKSL3a 中的498位残基是丙氨酸而不是IrTPS2 (S499) 中与水形成氢键的丝氨酸。为此,作者进一步构建了 IrKSL3a:A498S/I522A。结果表明,这种双突变体可以产生更多的 2。在此基础上获得的三取代突变体 IrKSL3a:A498S/I522A/E632K 和四取代突变体IrKSL3a:A498S/I522A/E632K/L523I 可更特异性地产生 2(Figure 2c)。值得注意的是,作者观察到IrKSL3a:I522T/E632K突变体产生了四个额外的二萜产物,进一步测试表明,IrTPS2:A522T也可制备出这些物质(Figure 1b)。
这些实验和计算结果表明,S499 和 T527 的氢键结合能力是IrTPS2 合成 2所必需的,且可以通过关键氨基酸残基的突变实现与IrKSL3a 功能的转换。

三、IrTPS2 和 IrKSL3a 的淬灭机制
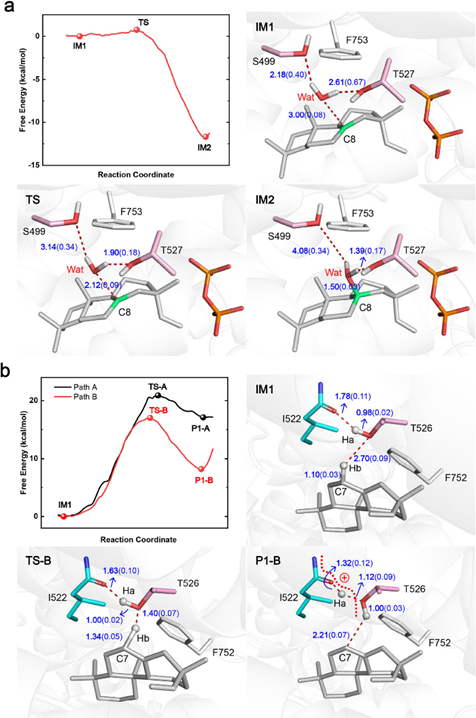
为了进一步研究 IrTPS2 的催化机制,作者通过QM/MM进行了自由能计算(Figure 4a)。将(+)-CPP替换为相关的碳正离子中间体,随着无机焦磷酸以及三重 Mg2+的释放,生成了第一个中间态 (IM1) 。自由能曲线表明,加水过程非常容易,因为它放热且几乎没有能量势垒。借助阳离子与C8碳正离子的极性相互作用,由S499和T527氢键连接的水分子(Wat)逐渐接近反应物,在过渡态(TS)中T527和Wat之间的氢键距离为1.9 Å,比IM1态的氢键距离(2.6 Å)短,而S499和Wat之间的氢键距离增加。这种转变在动力学上非常有利,其驱动力来自于加水后形成的烷氧基鎓反应物(IM2)使稳定性提高。
作者还用QM/MM计算了野生型IrKSL3a通过去质子化产生1的自由能曲线(Figure 4b)。模型结构表明,T526 的羟基是唯一可能进行去质子化的官能团。虽然最初的 QM/MM 自由能计算表明,这种去质子化的能量势垒约为 20.9 kcal/mol,使用 T526 去质子化生成 1需要吸收大量热(∼17.1 kcal/mol)(Figure 4b, path A),但还存在另一条路经,通过I522的酰胺键与T526形成氢键使T526的羟基在去质子化过程中正确定位,并通过稳定T526的质子化形式提高其碱性,从而使能量势垒降低到17.0 kcal/mol。(Figure 4b, path B)。此外,用其他氨基酸取代 T526 会显著降低催化活性进一步证实了T526的重要性。

四、蛋白工程获得了更多合成Nezukol的酶
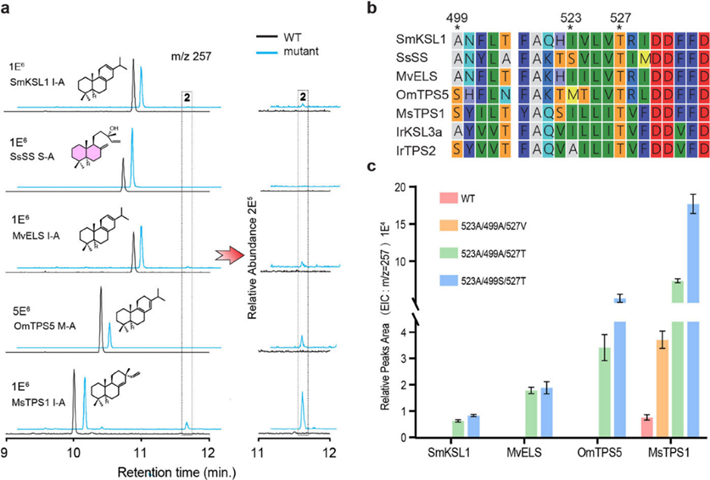
为了确定关键氨基酸位点功能是否普遍适用,作者对来自不同唇形科物种的同源蛋白进行了点突变获得变体 MsTPS1:I509A、OmTPS5:M353A、MvELS:I326A、SsSS:S333A和SmKSL1:I338A。结果表明,除SsSS:S333A之外的所有变体中均可检测到羟基化产物2,其产量按 MsTPS1:I509A > OmTPS5:M353A > MvELS:I326A > SmKSL1:I338A的顺序降低,显示 2 产量与酶和 IrTPS2 的同源性正相关(Figure 5a)。
为了研究水结合二联体(即 IrTPS2 中的 S499 和 T527)的作用,在上述四个单取代变体的基础上又进行了相关残基的突变。鉴于上述 T527 的保守性,所有四个变体中的 T527 都被缬氨酸取代,除此之外,用丙氨酸替换 MsTPS1 和 OmTPS5 保守的S499,用丝氨酸替换 MvELS 和 SmKSL1 的A499(Figure 5b)。与上文所示的 T527 的重要性相一致,除MsTPS1中仍能观察到少量的2外,其余突变体均无2的产生(Figure 5c)。与此相反,将 MvELS 和 SmKSL1 中的丙氨酸改为丝氨酸的影响微乎其微,虽然将 OmTPS5 和 MsTPS1 中的 S499 改为丙氨酸确实减少了 2 的生成量,但影响相对较小(Figure 5c)。这些结果进一步突出了3个残基的重要性,然而它们的效果进一步依赖于其他因素,IrKSL3a 也需要额外的突变来表现出2的选择性合成。这表明 IrTPS2中羟基化功能的获得经过了多步进化。

1
03
总结与讨论
作者通过对 IrTPS2 和IrKSL3a 的分子建模和分子动力学分析和 QM/MM 计算,发现了S499 和 T527两个影响酶羟化反应的关键二联体,且T527 很有可能参与酶催化过程,所有计算结果均通过突变实验进行了验证。将这些关键残基引入关系更远的 I 型 diTPSs 进一步检验了此结果的普适性和进化意义。具体来说,这三个关键残基(对应于 IrTPS2 中的残基位点 523/499/527)对于引入水催化羟化是必须的,但需要酶内的其他残基辅助才更有效,进而重塑功能,表明 IrTPS2 进化产生 Nezukol 的过程中存在选择压力。
研究展望
1、作者只对IrKSL3a的同功蛋白进行了进化上的正向突变,除此之外,还可以对IrTPS2的同功蛋白进行逆向突变已验证关键氨基酸的进化关系;
2、作者没有对新出现的产物进行进一步的研究,可以继续探究新产物产生的原因与机制;
3、尽管将本研究中发现的关键氨基酸进行突变可以使其产生羟化产物,但其产量极低且仍有大量烯烃产物,想要重塑Ⅰ型diTPS的功能还需要进一步的研究。

-END-
高颜值免费 SCI 在线绘图(点击图片直达)
最全植物基因组数据库IMP (点击图片直达)
往期精品(点击图片直达文字对应教程)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
















































 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









