







文 | OwlLite
感性和理性的冲突是人类认知 / 性格中的一个基本特点,其原因植根于人脑结构和功能的基本特征 (神经和生理性的),虽然问题是‘为什么人的大脑有时非常理性,有时非常不理性 (感性)?’,但实际中各人的平静状态倾向于感性还是理性是非常稳定的,不会轻易因为人的主观意识而变化,只有在受到外部扰动时候才会触发变动,并构成短期理性 / 感情的认知状态的锁定。其详细原理大致如下:
人脑在平静时候的工作拓扑状态可以成为默认网络 (Default Mode Network,DMN). 默认网络 [1] [2] 是神经科学领域的一个基本概念,不过它的历史并不算长,是随着正电子发射断层扫描(Positron Emission Tomography,PET) [3] 和静息态磁共振(fMRI) 技术应用研究的深化而发展出来的,这两类技术通过研究大脑各个区域之间的代谢水平来反映功能特征。默认网络的初衷在于研究大脑代谢水平的基准线 (Baseline),从而据此研究大脑在处理注意力的、目的性的、外源性的任务时候的代谢水平。但是 PET 研究发现,人类大脑在处理这些任务时候,一些区域的代谢水平会降低。因此 Baseline 并不存在一个全局的值,而是对不同的区域有不同的 Baseline. 这些区域代谢水平降低在不同的人和不同的注意力任务中体现了较好的一致性,反应出的是固定的模式,这个模式被称为默认网络,实质也就是下图中这些红色和黄色区域 [4]:
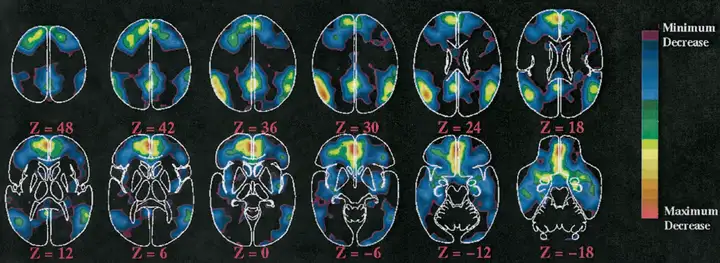
此外,默认网络还被发现牵涉情境记忆(episodic memory processing) [5] [6],这一事实的日常意义在下文中会解释。
人脑在处理注意力的、目的性的、外源性的任务时候,DMN 的代谢水平会降低,与此同时,另一些区域的代谢水平会升高,这些区域被称为任务正激活网络 TPN (Task Positive Network) [7] [8]:
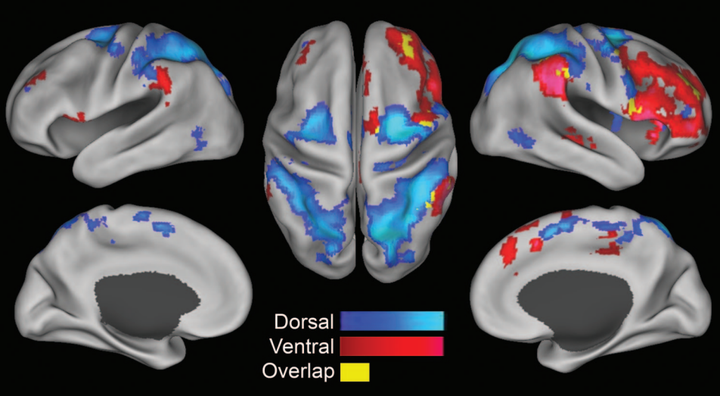
不难理解的,在进行任务时候,大脑中的 TPN 活跃,DMN 受抑,在休息时候则 DMN 活跃,TPN 受抑。也就是,这两个网络的代谢水平一般呈负相关关系 [9]:

换一种说法,这两个区域组(网络)之间是对抗关系[10],无论是在静息(大脑休息)还是任务(大脑活跃工作)状态下。
但是,大脑皮层每一个区域从来都不是单功能,或者说任务特定的。也就是,每个区域都会参与多种任务。有趣的是,大脑中的社交任务相关区域与 DMN 区域重合很大,而理性分析(数学的、机械的)的区域则跟 TPN 区域基本重合 [11]:
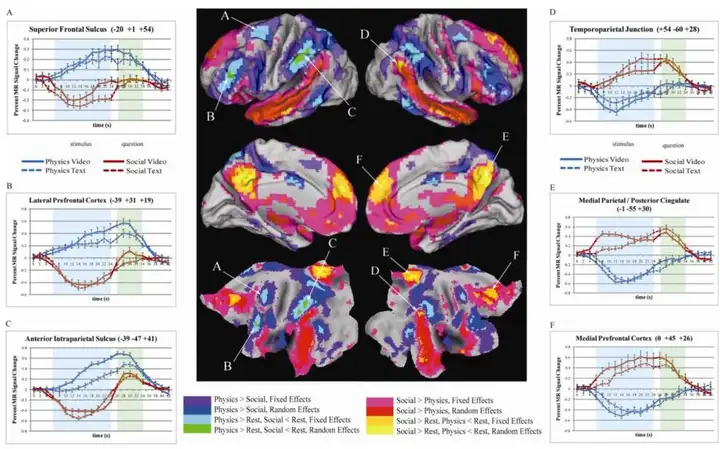
更多的研究证明,DMN 主要与共情能力、情绪自觉 (emotional self-awareness)、社交认知和伦理决策 (ethical decision making) 这一类能力相关。也就是说, 它是人类感性认识的基础结构,是我们认识自我反思自我的核心区域。而 TPN 则是分析性的、理性的,是我们得以认识客观世界的核心区域 [12]。同时,我们在处理一类任务时候,处理另一类任务的能力会降低,也就是,我们的感性和理性表现出二元对立特征。
这一对立的现实结果和表现是,当我们怒发冲冠时候,情绪性的区域高度激活,这使得我们不能客观的分析问题,不能合理的做出决策,表现出冲动型人格。而当我们长时间处理理性分析任务(投资分析、编程、数序和物理等),我们的共情能力又受到抑制,无法设身处地的去理解他人,我们的人类性因此湮没了,表现出机械性的木讷。
由于各人的生活经历、教育和工作的关系,在日常中会更多的处理一类任务:更多的社交性的需要感性的任务或者更多的需要理性的分析性任务。前者对应文科类的教育和工作,而后者大体对应理科类的。我们长期处理某一类任务,会增强大脑中对应区域的能力,从而在发育过程中强化其生理结构,也就是大脑功能的结构化,体现的是认知功能网络和结构网络的因果性 [13],这一因果性是我们学习新能力的基础。因此也能成就我们的外化性格:更感性还是更理性,更面向社交还是更面向分析,更情绪化还是去情绪化的。
此外,长期处理一类任务还会影响内分泌,表现为催产素、睾酮等分泌水平的变化 [14]。这些激素不仅影响性征和性格,还影响神经认知和神经发育。分别的促进社交性的和分析性的认知能力。从而形成反馈性的自锁因果环,感性的人因此越来越感性,而且更难以逆转;反之亦然。
总结来看,感性和理性在认知结构上存在对抗关系,处理一类任务会降低处理另一类任务的能力,进入感性或者理性状态的短期锁定。这种对抗导致,长期处理一类任务后,即使在休息情况下,也会降低处理另一类任务的能力,也就是形成结构性固化,表现为性格的决然不同;也就有了各人在平静状态下感性认知和理性认知的差异。人性有常,感性和理性的状态变化只是昙花一现的波动。
参考
- Costandi, Moheb. “Default Mode Network.” 50 Schlüsselideen Hirnforschung. Springer Berlin Heidelberg, 2015. 160-163
- Buckner, Randy L., Jessica R. Andrews‐Hanna, and Daniel L. Schacter. “The brain’s default network.” Annals of the New York Academy of Sciences 1124.1 (2008): 1-38.
- Raichle, Marcus E. “Positron emission tomography.” Annual review of neuroscience 6.1 (1983): 249-267.
- Raichle, Marcus E., et al. “A default mode of brain function.” Proceedings of the National Academy of Sciences 98.2 (2001): 676-682.
- Greicius, Michael D., et al. “Functional connectivity in the resting brain: a network analysis of the default mode hypothesis.” Proceedings of the National Academy of Sciences 100.1 (2003): 253-258.
- Greicius, Michael D., et al. “Default-mode network activity distinguishes Alzheimer’s disease from healthy aging: evidence from functional MRI.” Proceedings of the National Academy of Sciences of the United States of America 101.13 (2004): 4637-4642.
- Fransson, Peter. “Spontaneous low‐frequency BOLD signal fluctuations: An fMRI investigation of the resting‐state default mode of brain function hypothesis.” Human brain mapping 26.1 (2005): 15-29.
- Fox, Michael D., et al. “Spontaneous neuronal activity distinguishes human dorsal and ventral attention systems.” Proceedings of the National Academy of Sciences 103.26 (2006): 10046-10051.
- Fox, Michael D., et al. “The global signal and observed anticorrelated resting state brain networks.” Journal of neurophysiology 101.6 (2009): 3270-3283.
- Uddin, Lucina Q., et al. “Functional connectivity of default mode network components: correlation, anticorrelation, and causality.” Human brain mapping30.2 (2009): 625-637.
- Jack, Anthony I., et al. “fMRI reveals reciprocal inhibition between social and physical cognitive domains.” NeuroImage 66 (2013): 385-401.
- Boyatzis, Richard Eleftherios, Kylie Rochford, and Anthony Ian Jack. “Antagonistic neural networks underlying differentiated leadership roles.” Frontiers in human neuroscience 8 (2014): 114.
- Greicius, Michael D., et al. “Resting-state functional connectivity reflects structural connectivity in the default mode network.” Cerebral cortex 19.1 (2009): 72-78.
- Crespi, Bernard J. “Oxytocin, testosterone, and human social cognition.” Biological Reviews (2015): 000-000.
via:为什么人的大脑有时非常理性,有时非常不理性? - 知乎 OwlLite 发布于 2020-10-27 17:24
https://www.zhihu.com/question/427548455
风险选择行为
作者:生物流
问题太大,人的非理性行为太多,每一种非理性行为的脑机制差异也很大,很难在一篇问答中说清楚。
这里,单说一种非理性行为背后的脑机制——risky choice behavior,风险选择行为。风险选择行为是冲动行为(impulsivity)的一种。
什么是风险选择行为?
我们的人生中面临着许多选择。
例如,你打算将积蓄存进安全但收益低的银行,还是投进收益可能很高,但也有可能让你一贫如洗的股市?
例如,高考填报志愿时,你会选择一所100%能被录取的二本院校,还是选择一所不一定能被录取的985高校?
再例如,猎豹捕猎时,他是选择一头病怏怏跑不动的老绵羊,还是去追逐一匹浑身腱子肉的青年绵羊?老绵羊手到擒来,但肉少味寡。青年绵羊肉肥味美,但很可能猎不到。
以上的例子,主人公都面临着两种选择,一个选择奖励少,但确定能得到。而另外一种奖励大,但不一定能拿到,有一定风险。
正常情况下,大多数人会选择稳妥的方式。但有些时候我们会冒风险去搏一搏更大的收益。
风险选择行为即选择大奖励低概率的选项。
风险选择行为虽然在很多情况下会把事情搞砸,但它之所以被进化保留,自然有其有利的一面。它是一种冲动行为,会让我们更快地做出选择,避免了优柔寡断,延误战机,空手而归。同时,它也有一定概率让我们一夜暴富。

目前,科学家发现许多脑区都在风险选择行为中起作用,这些脑区大都属于大脑边缘系统。
伏隔核和眶额叶
为了模拟生活中常见的风险选择场景,科学家给动物或人类被试提供了两种类似的选择。

选择一,奖励小,但是有很高的概率或者确定能拿到。
选择二,奖励大,但是有一定的概率什么都得不到。
科学家发现,在人类被试做高风险的选择之前,即选择低概率的大奖励之前,大脑内的伏隔核会活跃起来。
伏隔核位于大脑深部,属于基底神经节,是腹侧纹状体的一部分。
伏隔核是大脑奖励系统重要的组成部分,它接受前额叶,中脑多巴胺,海马,杏仁核等重要脑结构的信息输入。
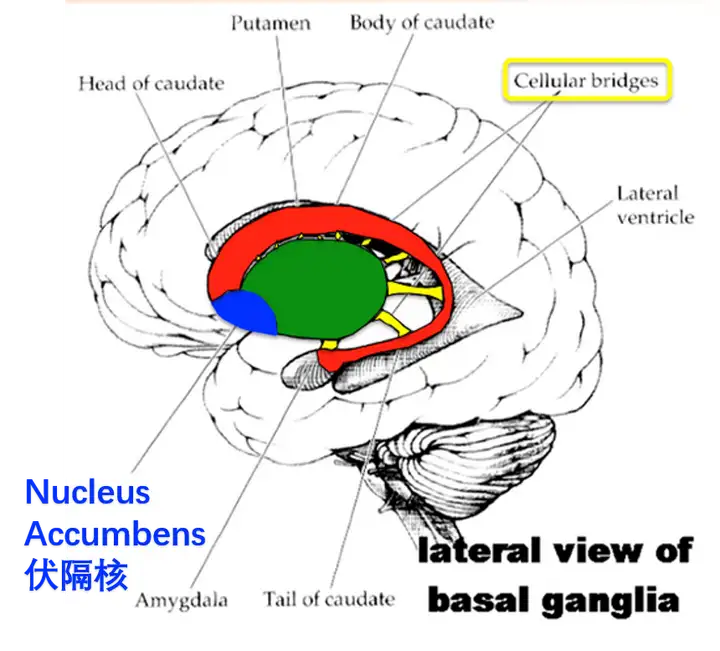
人脑内的基底神经节和伏隔核
如果将老鼠的伏隔核核心损毁,老鼠就不再那么频繁地做风险选择了。
这似乎跟伏隔核在动机产生方面的作用有关。如果动机足够强,总想一口吃个胖子,总想一夜暴富,那么我们就更倾向于高风险选择。而如果伏隔核坏了,动机就弱了,也就想安安稳稳做小日子,拿小奖励,不想再冒险了。
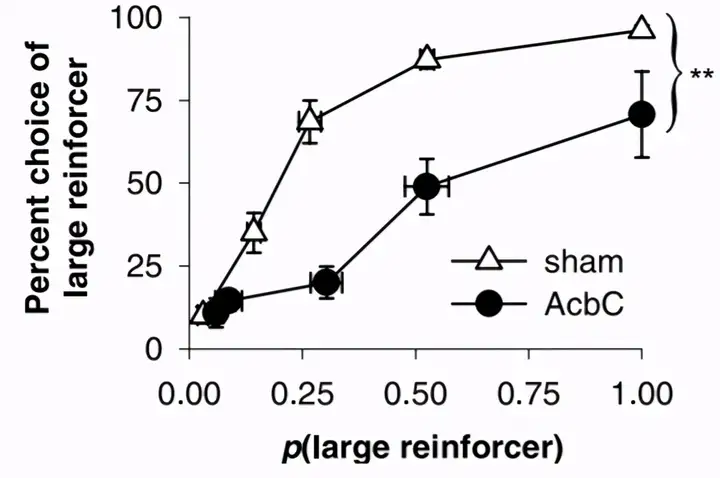
伏隔核: Nucleus accumbens core (AcbC)
而老鼠的内侧眶额叶,似乎起着相反的作用。损毁内侧眶额叶,老鼠的高风险选择显著增多了。

内侧眶额叶
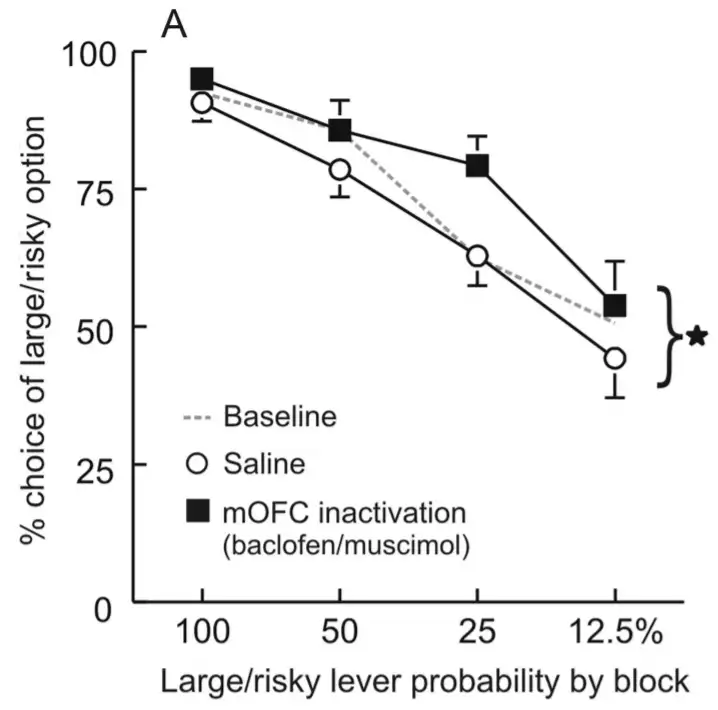
我们知道,眶额叶在价值计算,价值比较方面起着重要作用。可能的解释是,内侧眶额叶的损毁,会让老鼠无法比较不同选项的收益,无法清晰地计算每个选项的平均收益。老鼠只会看到当前的收益。当老鼠选择高风险选项,并且恰好获得大奖励时,它的眼里只有大奖励,无法将大奖励的低概率整合进来。这会让它在接下来的trial里,更频繁地选择高风险高收益的选项。
这种策略也称为“win-stay”,即赢了就保持原样,保持原来的选择。
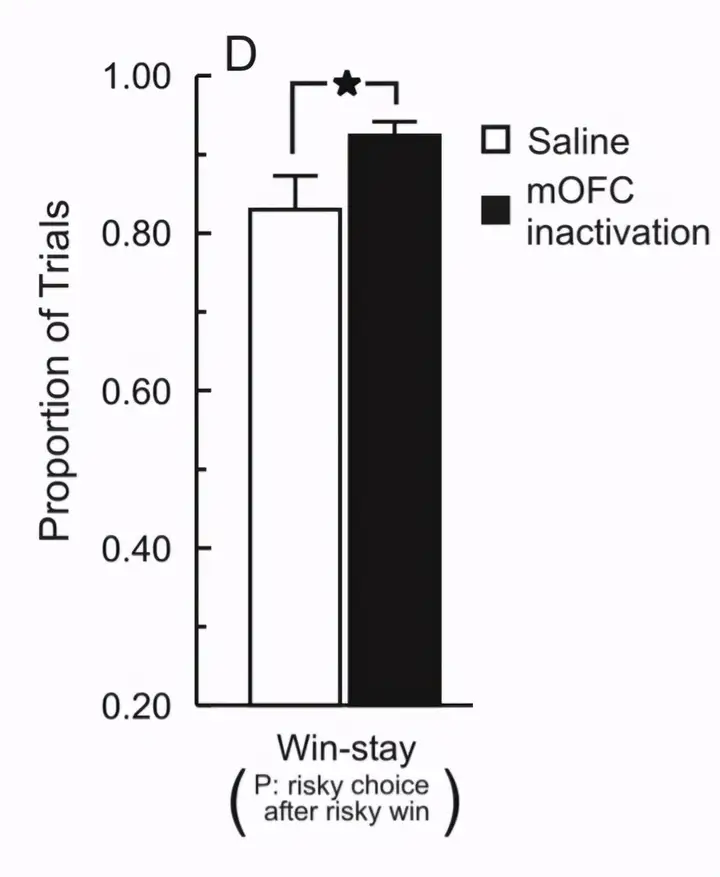
综上,从行为表现来看,伏隔核核心促进高风险选择,而内侧眶额叶则降低高风险行为。当它们损毁后,大鼠的行为朝反方向发展。
除此之外,科学家还发现基底外侧杏仁核-伏隔核,内侧前额叶-杏仁核这两条通路也在高风险行为中起着重要作用,并且是相反的作用。
基底外侧杏仁核-伏隔核,内侧前额叶-基底外侧杏仁核
为了阻断AB两个脑区之间功能连接,一种常用的手段是抑制一侧半球的A脑区,同时抑制另一侧半球的B脑区。这样,两侧半球内,AB之间都无法形成有效的信息交流。而对于脑区A和B,都在某一侧半球内是完整可用的。
利用此手段,科学家研究了基底外侧杏仁核-伏隔核,以及内侧前额叶-基底外侧杏仁核之间的信息交流,在风险选择中的作用。
科学家发现,当基底外侧杏仁核-伏隔核之间的功能连接被抑制时,大鼠的高风险行为减少。
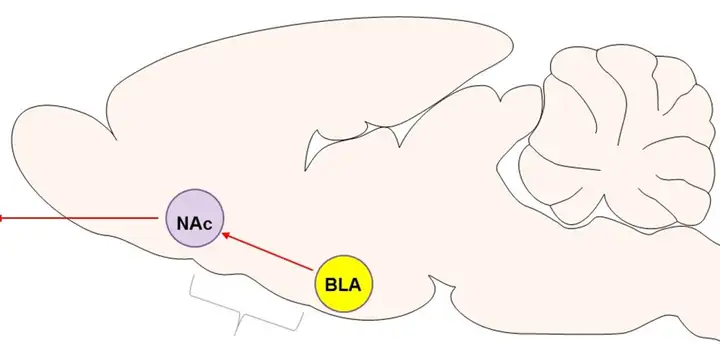
基底外侧杏仁核和伏隔核在老鼠脑内的位置
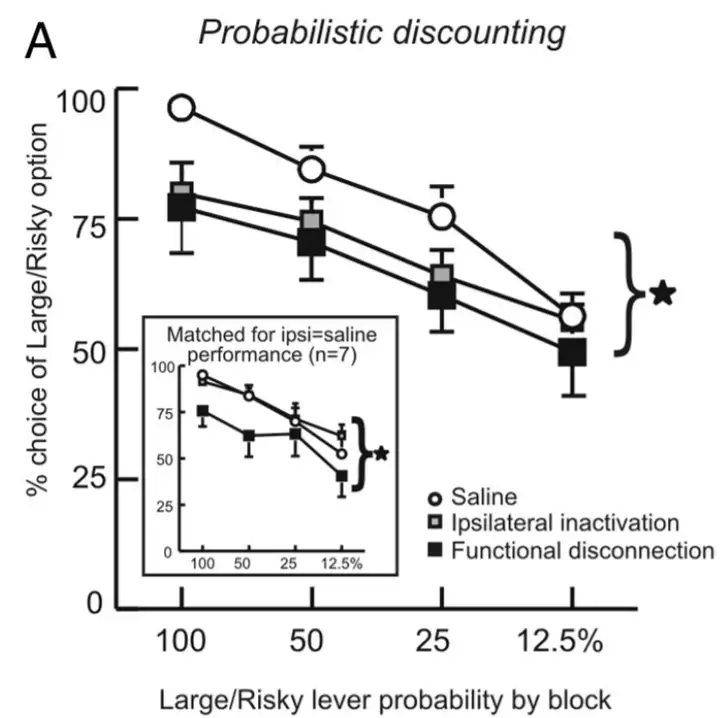
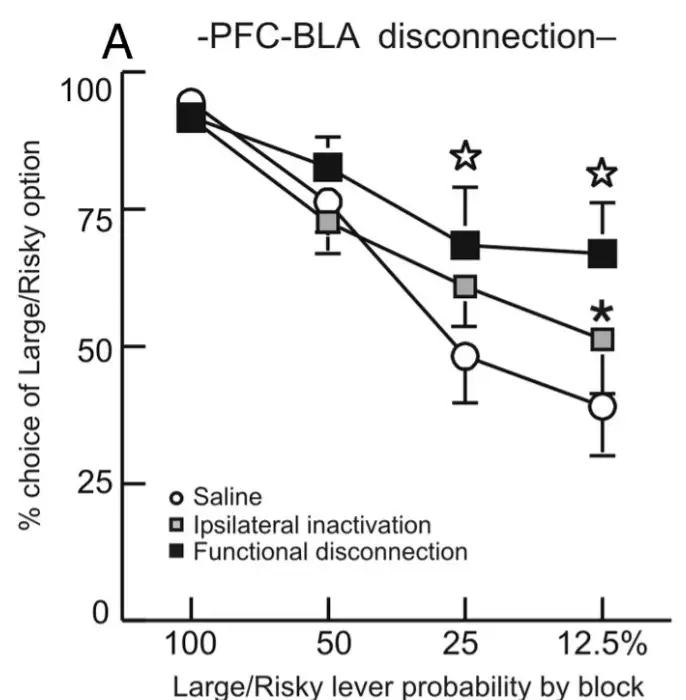
相反,当内侧前额叶-基底外侧杏仁核之间的功能连接被切断后,大鼠的高风险行为增加。
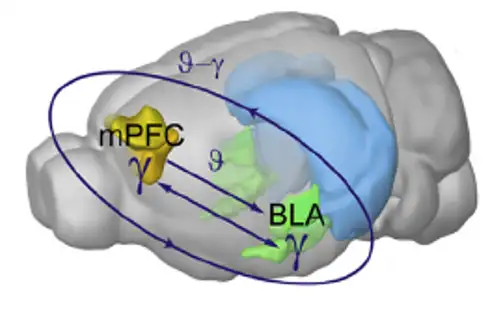
基底外侧杏仁核和内侧前额叶在老鼠脑内的位置
总结一下
科学家在大鼠上发现,从行为表现来看,伏隔核核心以及基底外侧杏仁核-伏隔核之间的信息交流促进风险选择,而内侧前额叶以及基底外侧杏仁核-内侧前额叶之间的功能连接,降低风险行为。
神经递质,例如多巴胺/五羟色胺也在风险选择行为中扮演着复杂的角色,后面我会继续更新。
相关文章:
参考文献:
Separate Prefrontal-Subcortical Circuits Mediate Different Components of Risk-Based Decision Making
Effects of lesions of the nucleus accumbens core on choice between small certain rewards and large uncertain rewards in rats
Selective Involvement by the Medial Orbitofrontal Cortex in Biasing Risky, But Not Impulsive, Choice
Dissociable Contributions by Prefrontal D1 and D2 Receptors to Risk-Based Decision Making
Receptor-Specific Modulation of Risk-Based Decision Making by Nucleus Accumbens Dopamine
编辑于 2020-11-10 08:04
via: 为什么人的大脑有时非常理性,有时非常不理性? - 知乎 生物流BioStream
https://www.zhihu.com/question/427548455/answer/1567177988
大脑情绪控制系统——边缘系统
作者:生物流
边缘系统英文名字为limbic system。limbic 源自于拉丁语,意为“边界”,意指边缘系统位于大脑皮层的边界位置。边缘系统在情绪反应中起重要作用,同时也在学习、记忆、行为中起关键作用。
边缘系统命名来源
大脑边缘系统最早是由美国神经学家Paul D. MacLean提出并命名的。边缘系统也叫古哺乳动物脑,是Paul提出的三重脑模型的“一重脑”。另外“两重脑”分别是爬行动物脑、新哺乳动物脑(或叫新皮层)。
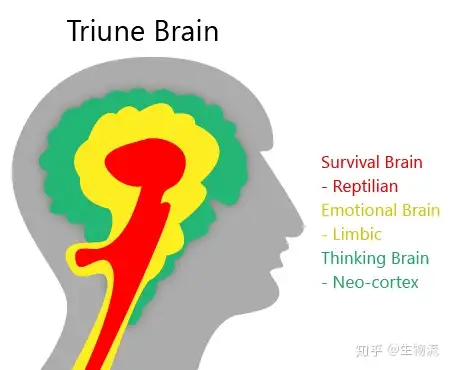
三重脑模型
三重脑模型是从大脑进化的角度划分的。
爬行动物脑位于大脑的最底部,主要是中脑和后脑组成的脑干。爬行动物脑在进化上也最早出现。爬行动物脑控制着动物的攻击、领地意识、等级意识等跟生存密切相关的行为,Paul认为这些行为起源于爬行动物。
最顶部是新哺乳动物脑,也即大脑新皮层。
夹在新哺乳动物脑和爬行动物脑之间,即脑干和新皮层之间的一系列脑组织,便是边缘系统。

边缘系统位于脑干和新皮层之间
边缘系统的构成
控制情绪的边缘系统,究竟包括哪些脑结构,并没有一个大家都一致认可的答案。但大体上,边缘系统可分为两类,一类隶属于皮层,也称为边缘皮层(limbic lobe)。另一类是一系列皮层下结构。
我们分别来看。
边缘皮层
边缘皮层,形象地看,即为大脑皮层的边界(limbic),位于大脑皮层和皮层下结构之间。
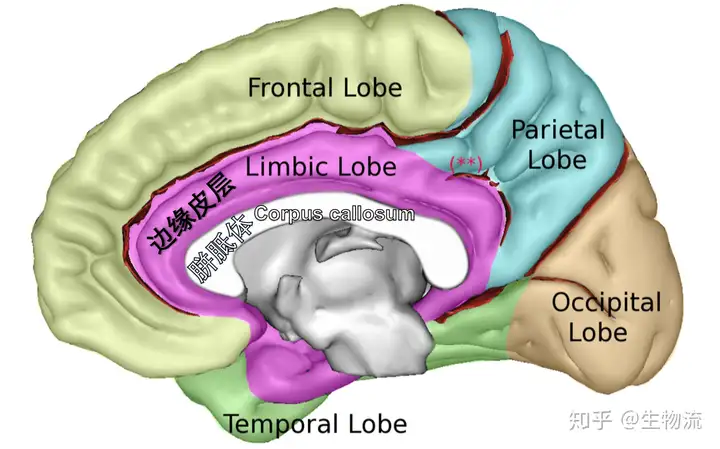
大脑边缘叶(limic lobe),红色部分
边缘皮层包围着胼胝体,形成一个指环状结构。这一系列结构包括扣带回、胼胝体下区(Subcallosal area)、海马旁回、海马、齿状回等。
也有人认为,边缘系统也包括眶额叶、岛叶。
扣带回既可以调节血压心跳等身体的自主功能,又参与认知、注意以及情绪的调节,这些功能参与情绪的表达。
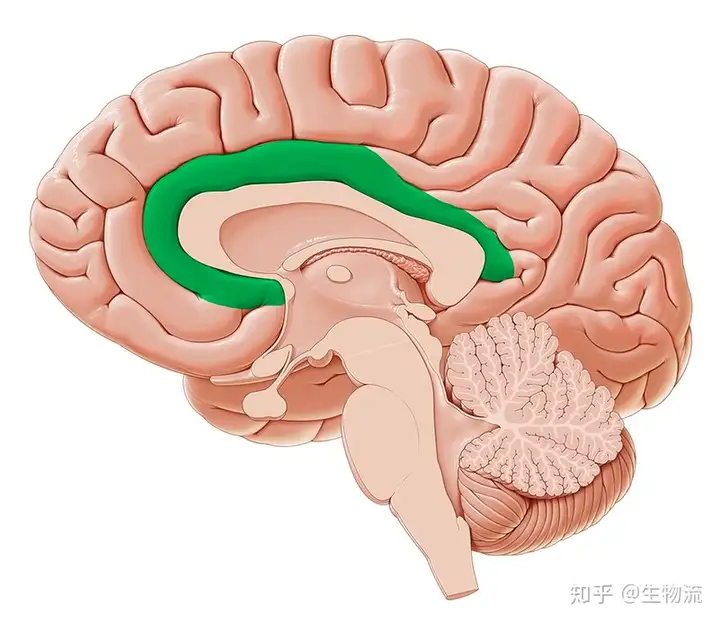
扣带回,绿色部分
海马位于颞叶深处,是最重要的记忆脑组织,参与长期记忆的形成、存储和提取,当然也包括情绪记忆。

大脑海马
眶额叶位于眼眶之后,是处理价值的重要脑区,在基于价值的抉择中起着重要作用。另外,眶额叶可以在适当的时候,抑制恐惧情绪,以避免不必要的情绪反应。眶额叶受损的猴子,面临危险时,会表现出异于常猴的恐惧反应。并且,即使长时间地面对一个假的危险,如橡皮蛇,其恐惧应激反应也无法很好地消退,即无法适应危险。眶额叶功能的异常,很可能跟焦虑有关。

眶额叶
皮层下边缘系统
皮层之下的边缘系统包括嗅球,下丘脑(Hypothalamus),杏仁核(Amygdala)、伏隔核(Nucleus accumbens)等。
杏仁核是情绪处理中心,参与情绪的产生以及表达。例如,焦虑、攻击、尤其是恐惧,还参与情绪记忆的形成和保存。

杏仁核(amygdala)
恐惧的面部表情图片,会激活杏仁核。杏仁核受损的人,无法再识别出一张脸上的恐惧。同时,他们也无法再拥有恐惧的感觉,他们变得极其大胆,已经不知道恐惧为何物。当然,恐惧触发的自主神经激活以及内分泌也荡然无存。
在所有的情绪中,之所以恐惧占据中心位置,是因为恐惧关乎动物的生死。看见天敌或经历危险时,如果动物没有任何应激反应,不逃跑也不去战斗,反而无所事事地凑近危险源,等待它的只有死亡。

情绪激发的身体反应,主要通过下丘脑来实现。下丘脑可以通过调节内分泌激素的产生和释放,进而调节自主神经系统。自主神经系统又称为植物神经系统,可以在无意识的情况下调节我们的血压、呼吸、心跳、消化、瞳孔反应、性唤醒等等。这些身体的自主反应对于情绪的表达必不可少。
伏隔核是腹侧纹状体的一部分,接收中脑腹侧被盖区(VTA)多巴胺神经元的投射,是大脑奖励系统的重要部分,介导动物的强化学习、以及动机(motivation)。伏隔核内的多巴胺释放,会使得动物有更强大的动力去完成或追求某件事情,例如,去寻找食物、努力学习去获得奖励等等。在药物成瘾中,VTA和伏隔核这条动机通路,介导了疯狂的药物寻找行为和情绪。

位于腹侧纹状体的伏隔核,绿色部分
总结
大脑皮层内和皮层下的边缘系统,组成了一个复杂的神经网络,来控制情绪的生成和表达,以及情绪记忆的形成、存贮和提取。
也有人认为,情绪的生成和调控极其复杂,仅仅归因于一个简单的边缘系统是远远不够的。
via: 大脑情绪控制系统——边缘系统 - 知乎 生物流BioStream 发布于 2020-09-03 22:42


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









