




为了活下去,它们一直在欺骗同类
原创 环球科学
一群病毒社会学家正在探索一个 70 多年前就藏在鸡胚中、奇怪的病毒繁殖现象。而其中的关键系于病毒在自然繁殖中会自发产生的缺陷病毒。这些缺陷病毒因缺乏一些基因,只能在宿主中等待完整病毒,再通过 “欺骗” 繁殖自身。不过,基因组短的优势让缺陷病毒能充分利用完整病毒的资源,并抑制它们的生存。随着各种病毒特别的 “社交” 故事被揭开,科学家开始从一些新的视角来理解病毒的行为,并尝试利用这些微观的 “社交” 来攻克极其危险的病毒。
在这篇文章中,我们将跟随卡尔・齐默的讲述,看看最新的研究揭示了一个怎样的病毒社交世界。
撰文|卡尔・齐默(Carl Zimmer)
翻译|赵建元
自 19 世纪末期科学家发现病毒以来,他们就将病毒与其他生命体区分开来。病毒远小于细胞,它的蛋白质外壳内除了一些基因外,几乎什么都没有。它们无法生长、无法靠自身复制基因和进行其他任何活动。研究人员认为每个病毒都是独自在世界上漂泊的颗粒,只有恰巧入侵合适的细胞后,它们才能复制。
新加坡科技研究局(Agency for Science, Research and Technology)传染病实验室的病毒学家马尔科・维纽齐(Marco Vignuzzi)说,最初病毒正是因为这样的简单性吸引了众多科学家,“我们都想要成为还原论者”。
这种还原主义也得到了回报。有关病毒的研究对现代生物学的诞生至关重要。得益于病毒缺乏复杂的细胞结构,科学家揭示了基因发挥作用的基本原则。不过维纽齐表示,这是有代价的:如果你假设病毒很简单,就有可能蒙蔽自己的双眼,忽视它们在某些方面可能会很复杂,而我们对此一无所知。
举例来说,当你把病毒当作一个孤立的基因集时,想象它们拥有社会生活就会显得荒谬。但是,维纽齐和一群志同道合的病毒学家完全不会这么认为。最近数十年来,他们发现病毒拥有一些奇怪的特征,无法用它们是一个独立的颗粒这种观点来解释。他们正在揭示一个奇妙且复杂的病毒社会。这些病毒社会学家(sociovirologist,有时研究人员会这样自我称呼)认为,病毒只有作为病毒社群中的成员时才有意义。
诚然,病毒的社会生活一定不会与其他物种完全相同。它们无法像人类一样在社交媒体上发布自拍照,在食品银行做志愿者或盗窃其他人的身份。它们也不会像雄性狒狒为了统治群体而那样相互打斗,不会像蜜蜂收集花蜜来喂养蜂王。它们甚至不会像一些细菌会凝聚成一层黏滑的垫子来共同抵御环境压力。即便如此,一些病毒社会学家认为,病毒确实会欺骗、合作,以及用别的方式和其他病毒互动。
病毒社会学尚处于发展早期,是一个规模较小的研究领域。2022 年,专门讨论病毒社会生活的会议首次举行,第二次会议已于今年 6 月举行,共有不到 80 人出席这次会议。尽管如此,病毒社会学家认为,这个新领域或许会产生深远的影响。如果我们认为病毒之间相互孤立,那么像流感这样的疾病似乎不应该存在。反之,如果我们能揭开病毒的社会生活,或许能利用这一点来反击一些病毒引发的疾病。
线索触手可及
近一个世纪以来,一些有关病毒社会生活最重要的证据其实一直摆在人们眼前。20 世纪 30 年代初,科学家发现流感病毒后,就想到将它们注射到鸡胚中,以此让它们繁殖和储存。随后,研究人员便可以用这些新病毒来感染实验动物,借此开展研究,或者将它们注入新鸡胚中继续培养。
20 世纪 40 年代末期,丹麦病毒学家普雷本・冯・芒努斯(Preben von Magnus)在培养病毒时发现了一些奇怪的现象。他发现,将一个鸡胚中培养的许多病毒转接到新的鸡胚后,它们常常会无法复制。到第三轮转接时,只有万分之一的病毒还会继续复制。不过在此之后的转接中,有缺陷的病毒会越来越少,那些能复制的病毒又会卷土重来。芒努斯怀疑这些无法复制的病毒没有发育完全,因此称它们为 “不完全” 的。
在后续数年中,病毒学家将病毒在培养中表现出 “不完全” 的兴衰过程称为 “冯・芒努斯效应”(the von Magnus effect)。在他们看来,这虽然很重要,但只是一个需要解决的问题。由于没有人在实验室培养物之外见过缺陷病毒(也称为不完全病毒),因此病毒学家认为它们是人造的,并想出了摆脱它们的方法。
美国加利福尼亚大学戴维斯分校的病毒学家萨姆・迪亚斯 - 穆尼奥斯(Sam Díaz-Muñoz)回忆起当时在该领域存在一个普遍观点。他说:“如果你不希望缺陷病毒干扰实验,就必须从实验室的库存中剔除它们。因为它们不是‘天然存在’的。”20 世纪 60 年代,研究人员观察到缺陷病毒的基因组会比典型的完整病毒,更短。这一发现也加深了许多病毒学家原有的观点,即缺陷病毒是有缺陷的 “怪胎”,缺乏复制所需的基因。但在 21 世纪的第二个十年,价格低廉且强大的基因测序技术清晰地揭示,人体内实际上存在着大量缺陷病毒。
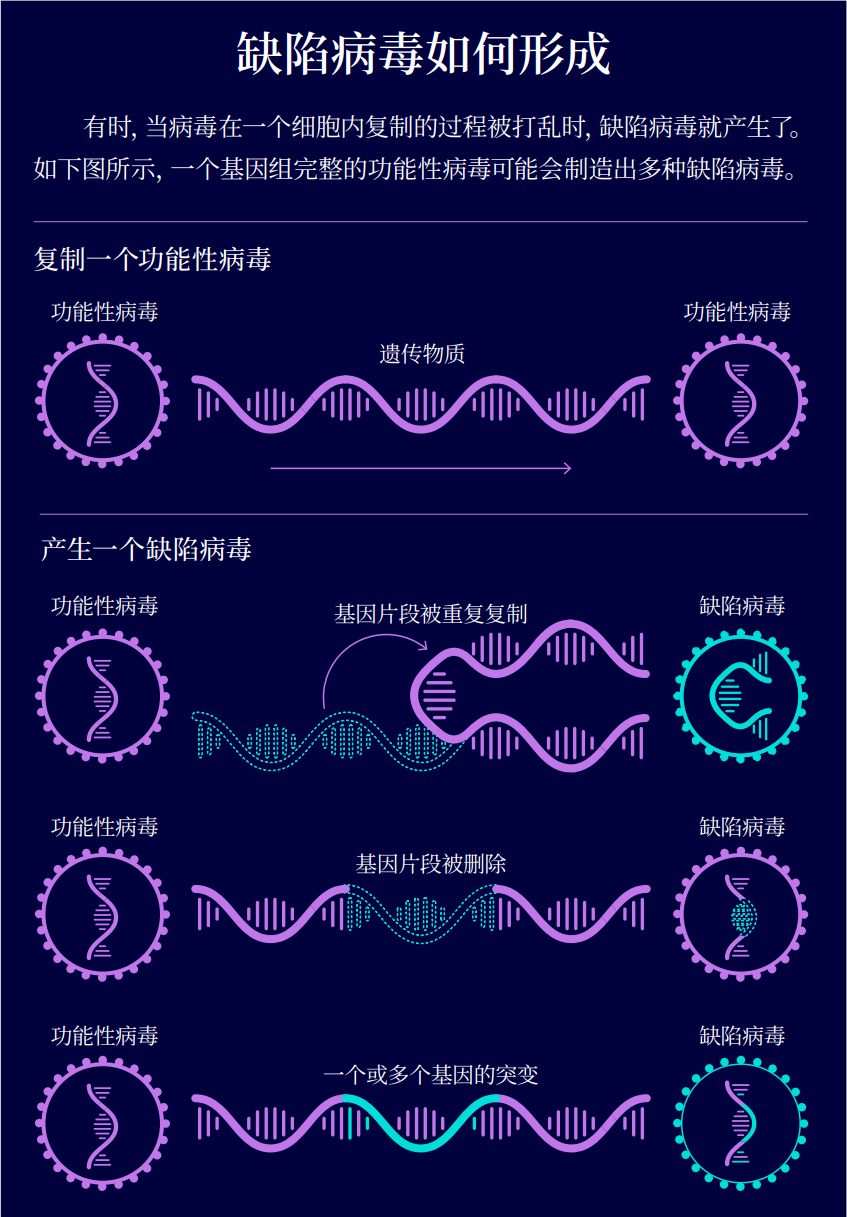
在一项发表于 2013 年的研究中,美国匹兹堡大学(University of Pittsburgh)的研究人员用棉签擦拭了流感患者的鼻子和嘴巴。他们从这些棉签样本中提取了流感病毒的遗传物质,发现其中一些病毒会缺少部分基因。以流感病毒为例,在被感染的细胞中,当完整病毒的聚合酶错误复制自身的基因组或者意外跳过一段基因时,就会产生这样发育不良的病毒。
其他的一些研究也证实了这一发现。他们还揭示了缺陷病毒形成的其他方式。例如,一个被感染的细胞可能会在复制病毒的基因组时突然中途逆转,然后反向复制回到起始点。这样病毒会携带混乱的基因组。此外,缺陷病毒的形成还可能是由于一些突变破坏了基因序列,无法产生功能性蛋白质。这些研究也推翻了此前的假设,即冯・芒努斯的缺陷病毒仅仅是实验室中的人造产物。“它们是病毒生物学中一个自然的组成部分,” 迪亚斯 - 穆尼奥斯说。
这些关于人体内存在缺陷病毒的发现,也再次激起了科学家的兴趣。流感病毒的情况并非个例:许多病毒都会以缺陷的形式存在。研究人员在呼吸道合胞病毒(RSV)和麻疹病毒等的感染者体内,发现了大量的缺陷病毒。他们还为这些缺陷病毒取了新的名字,其中一部分人会称它们为 “有缺陷的干扰病毒颗粒”,还有人称它们为 “非标准的病毒基因组”。而迪亚斯 - 穆尼奥斯和同事们给它取名为 “骗子”。
病毒式欺骗
通常情况下,缺陷病毒可以进入细胞,但它们无法自行复制,这是由于它们缺乏必需的基因,例如一种编码聚合酶(这种酶会参与基因复制)的基因。为了复制,它们必须 “欺骗”,必须利用同类病毒。对于缺陷病毒来说,还算幸运,因为细胞经常被含有不同基因组的病毒感染。如果缺陷病毒所在的细胞中存在一个功能性病毒(拥有完整的基因组),这个病毒就会产生聚合酶,而它们也可以借用这些聚合酶来复制自己的基因。
缺陷病毒的欺骗
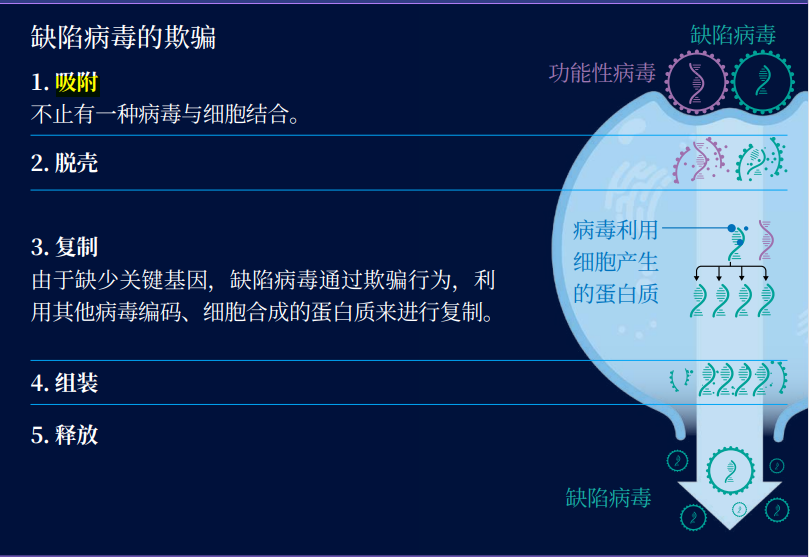
在这样的细胞中,两种病毒都会尽可能多地复制自己的基因组。不过缺陷病毒拥有一个巨大的优势:它们需要复制的遗传物质更少。这也意味着,聚合酶可以更快复制完缺陷病毒的基因组,而不是功能性病毒的基因组。
随着缺陷病毒和功能性病毒一起从一个细胞转移到另一个细胞,它们在感染过程中的优势也会逐渐放大。“如果你的基因组只有一半的长度,并不意味着你只会有双倍优势,” 美国耶鲁大学的博士后研究员阿舍・利克斯(Asher Leeks)说,他主要研究病毒的社会演化。他说,“这可能意味着缺陷病毒可以获得 1000 倍甚至更多的优势。”
其他一些缺陷病毒可能拥有有效的聚合酶,但缺乏基因来制造蛋白质外壳,包裹其遗传物质。它们常会在细胞中等待功能性病毒的出现,然后偷偷将自身的基因组运送到功能性病毒产生的衣壳中。一些研究表明,相比于功能性病毒,缺陷病毒的基因组可以更快地进入衣壳中。无论缺陷病毒采用何种策略来复制,结果都是一样的。即便它们利用与其他病毒的合作来为自己谋利,也无需承担合作的代价。
“单独一个缺陷病毒并无优势,但与其他病毒合作优势就会凸显。不过,如果都是缺陷病毒,那么就没有功能性病毒可以利用了。” 迪亚斯 - 穆尼奥斯说,“从一种演化论的角度来看,这就是我们定义病毒欺骗的证据。”
然而,这个定义的最后一部分也引出了一个疑问。如果缺陷病毒可以获得如此惊人的成功 —— 事实上也确实如此 —— 它们应该会将病毒推向灭绝。随着一代代病毒从旧细胞中释放,感染新细胞,缺陷病毒应该会越来越普遍。它们会不断复制,欺骗功能性病毒直到后者消失。而如果没有任何功能性病毒留下,缺陷病毒也会无法自行复制,最终整个病毒群体应该会被彻底消灭。
当然,一些病毒(例如流感病毒)显然躲过了这种迅速灭绝,这也意味着它们的社会生活中除了如死亡螺旋般的 “欺骗” 外,可能还存在其他未知。美国华盛顿大学医学院(Washington University School of Medicine)的病毒学家卡罗莱娜・洛佩斯(Carolina López)认为,在病毒社会中,一些病毒看起来像在 “欺骗” 群体,但实际上可能扮演着更温和的角色。它们并不是在利用同类病毒,相反是在与它们合作,帮助整个种群生存。
模糊的界限
病毒社会学家正在试图弄清楚在病毒世界里有多少的欺骗和合作。或许研究动物行为的科学家能感同身受这些研究有多难。一个个体在某些情况下可能会欺骗,但在其他情况下又可能会合作,而且一种看似合作的行为也可能是从自私的欺骗演化而来。
利克斯赞同缺陷病毒可能是病毒群落中富有功能的一部分。但他认为,我们需要始终重视这样的可能性,那就是即使它们看起来是在合作,实际上可能仍然在欺骗。根据演化论,病毒的基因组非常小,因此经常会有欺骗行为。“在病毒群体中,冲突占主导,” 利克斯说。
事实上,欺骗行为可以演化出适应性,让病毒之间看起来像在合作。利克斯最喜欢的案例之一是擅长隐藏冲突的矮缩病毒(nanovirus)。这种病毒会感染欧芹和蚕豆等一些植物,而它们的复制方式十分惊人。它们总共有 8 个基因,但每个病毒颗粒只含有一个基因。这也意味着,只有当 8 个携带不同基因的矮缩病毒颗粒同时感染同一株植物时,它们才有可能进行复制。植物细胞可以根据这 8 个基因来制造蛋白质,并将病毒基因的新拷贝包装进新的病毒衣壳中。
合作的多成分病毒
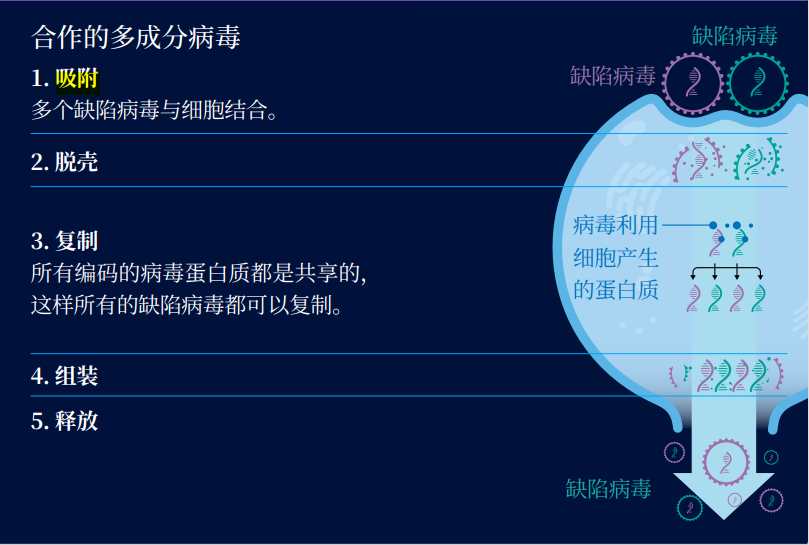
你可能会认为矮缩病毒是一个教科书式的合作案例。毕竟,其中任何一种病毒想要复制,都需要和其他的病毒协同工作。这个过程有点类似于蜂群中的分工,工蜂们需要分开去采集花蜜,照顾幼虫和为蜂巢寻找新的迁移地点。不过,利克斯和同事们已经绘制了矮缩病毒以及其他的多成分病毒(multipartite viruses)如何在它们合作的面具之下隐藏欺骗。
试想一下,最初矮缩病毒的祖先将所有 8 个基因都打包在一个病毒基因组中。然后,这种病毒却意外产生了一种只含有一个基因的缺陷病毒。当功能齐全的病毒复制基因,这种缺陷病毒会茁壮成长。而如果演化出第二个携带另一个基因的缺陷病毒,它也可以从完整的病毒那里得到好处。
当利克斯和同事们建立数学模型来模拟这个演化情景时,他们发现病毒可以十分轻易分裂出更多的变种。它们会持续分裂,直到能独立复制的原始病毒彻底消失。如今的矮缩病毒需要相互依赖才能继续生存,仅仅是因为它们的祖先曾相互利用。想要理清病毒社会的本质或还需要数年的时间,不过解开这个谜团将能带来巨大的回报。一旦科学家充分了解病毒的社会行为,他们就有可能让它们相互对抗。
扭转局面
在美国纽约大学,本・特诺弗(Ben tenOever)和同事们正在设计一种可能更有效的流感病毒 “骗子”。他们利用了病毒的生物学特点:每隔一段时间,感染同一个细胞的两种病毒的遗传物质会包装在一起,形成一种新的病毒。他们想知道是否可以制造出一种欺骗性的病毒,可以很容易入侵一个功能性流感病毒的基因组。
这个研究小组从受感染的细胞中收集了缺陷病毒。从这一批病毒中,他们发现了一种超级病毒 “骗子”,它们非常善于让自己的基因渗透进功能完整的流感病毒中。得益于这种缺陷病毒的干扰,研究人员获得的杂交病毒的复制能力都较差。
通过观察这种病毒 “骗子” 如何作为抗病毒药物发挥作用,特诺弗和同事将其包装成了一种鼻喷雾剂。在实验中,他们先用一种致命的流感病毒感染小鼠,随后将这种病毒 “骗子” 喷到小鼠的鼻子里。结果发现,这种病毒 “骗子” 非常擅长利用功能性病毒,并且会减缓它们的复制。这些小鼠在数周内就从感染中康复了。然而,如果没有这些病毒 “骗子” 的帮助,这些动物就会死亡。
研究人员发现在小鼠被感染之前,将这些超级病毒 “骗子” 喷进小鼠的鼻子里,效果会更好。这些病毒 “骗子” 会一直潜伏在小鼠体内,一旦功能性流感病毒感染小鼠,就会立即被病毒 “骗子” 攻击。在这之后,特诺弗和同事们开始用雪貂进行实验。雪貂感染流感病毒的过程和人类更接近:尤为关键的是,不同于小鼠,一只生病的雪貂可以很轻易地将流感病毒传染给相邻笼子里健康的雪貂。他们发现鼻内喷雾剂也能迅速减少受感染的雪貂体内的流感病毒数量,出现和小鼠实验相似的结果。然而,当科学家观察受感染的雪貂传染给健康雪貂的病毒时,他们发现它们不仅传播了功能性病毒,还包括隐藏在病毒衣壳内的病毒 “骗子”。
这一发现也引出了一种令人激动的可能性:这些超级病毒 “骗子” 或许可以阻止一种新型的流感病毒传播。如果人们接受含有超级病毒 “骗子” 的喷雾治疗,就有可能迅速从感染中恢复。如果他们确实将新病毒传播给了其他人,那么这些超级病毒 “骗子” 将一同传播出去,进而阻止新病毒传播。“这是大流行病的中和剂,” 特诺弗说。
至少,这种可能性在概念上是成立的。特诺弗需要进行一次临床试验,来观察这种在动物中有效的方式是否也会在人类中起效。然而,他提到监管机构对于批准这样的试验可能会感到不安,因为这不仅是给人们一种具有抗病毒作用的药物,这种药物还会传染给其他人,无论他们是否同意。“这似乎是一个死亡之吻,” 特诺弗说,他希望可以将病毒社会学的研究应用于医学。
迪亚斯 - 穆尼奥斯认为,考虑到我们对病毒社会学还有许多的未知,谨慎地利用病毒社会学无疑是正确的。采用惰性分子制造药物和利用病毒的社会生活完全是两回事。“这些有缺陷的病毒是活生生的,且不断在演化,” 迪亚斯 - 穆尼奥斯说。
本文节选自《环球科学》2024 年 9 月刊、原刊于西蒙斯基金会发起的 Quanta Magazine 的文章:《病毒的隐秘社交》
via: 为了活下去,它们一直在欺骗同类 环球科学 2024 年 09 月 06 日 21:30 北京
https://mp.weixin.qq.com/s/4tne5NwUSouaWKtJ_Dp5XQ
「细菌」和「病毒」有什么区别?
作者:林鱼儿
细菌和病毒区别较大,不仅仅是定义上,其实从大小,形状,进化,生物活性,治疗手段等很多方面都有很大的不同。
1、细菌
1.1 定义
广义的细菌即为原核生物。是指一大类细胞核无核膜包裹,只存在称作拟核区(nuclear region)(或拟核)的裸露 DNA 的原始单细胞生物,包括真细菌(eubacteria)和古生菌(archaea)两大类群。人们通常所说的即为狭义的细菌,狭义的细菌为原核微生物的一类,是一类形状细短,结构简单,多以二分裂方式进行繁殖的原核生物,是在自然界分布最广、个体数量最多的有机体,是大自然物质循环的主要参与者。
1.2 大小及分类
1.2.1 大小
目前已知最小的细菌只有 0.2 微米长,因此大多只能在显微镜下看到它们;而世界上最大的细菌可以用肉眼直接看见,有 0.2-0.6 毫米大,是一种叫纳米比亚嗜硫珠菌的细菌。
1.2.2 分类
细菌可以按照不同的方式分类。细菌具有不同的形状。大部分细菌是如下三类:杆菌是棒状;球菌是球形(例如链球菌或葡萄球菌);螺旋菌是螺旋形。另一类,弧菌,是逗号形。
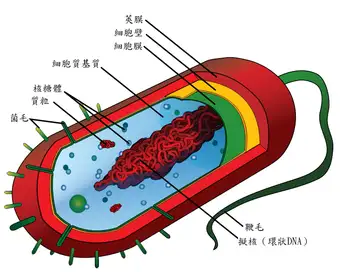
细菌的结构十分简单,原核生物,没有膜结构的细胞器例如线粒体和叶绿体,但是有细胞壁。根据细胞壁的组成成分,细菌分为革兰氏阳性菌和革兰氏阴性菌。“革兰氏” 来源于丹麦细菌学家汉斯・克里斯蒂安・革兰,他发明了革兰氏染色。
这是一般情况下细菌的结构
这是众多细菌的形状
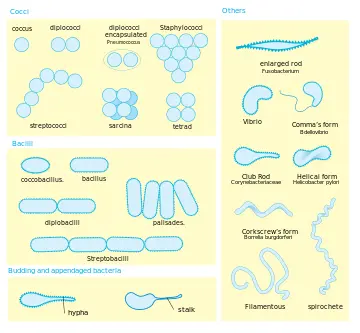
1.3 代谢特征繁殖方式及进化
1.3.1 代谢
细菌的营养方式有自养及异养,其中异养的腐生细菌
是生态系统中重要的分解者,使碳循环能顺利进行。部分细菌会进行固氮作用,使氮元素得以转换为生物能利用的形式。细菌也对人类活动有很大的影响。一方面,细菌是许多疾病的病原体,包括肺结核、淋病、炭疽病、梅毒、鼠疫、砂眼等疾病都是由细菌所引发。然而,人类也时常利用细菌,例如乳酪及酸奶和酒酿的制作、部分抗生素的制造、废水的处理等,都与细菌有关。在生物科技领域中,细菌有也著广泛的运用。
1.3.2 繁殖
细菌以无性方式进行繁殖,最主要的方式是以二分裂法这种无性繁殖的方式:一个细菌细胞细胞壁横向分裂,形成两个子代细胞,在分裂的时候可以产生遗传重组。单个细胞也会通过如下几种方式发生遗传变异:突变(细胞自身的遗传密码发生随机改变),转化(无修饰的 DNA 从一个细菌转移到溶液中另一个细菌中,并成功整合到该细菌 DNA 或质粒上,使之具有新的特征),转染(病毒的或细菌的 DNA,或者两者的 DNA,通过噬菌体这种载体转移到另一个细菌中),细菌接合(一个细菌的 DNA 通过两细菌间形成的特殊的蛋白质结构,接合菌毛,转移到另一个细菌)。细菌可以通过这些方式获得基因片段,通过分裂,将重组的基因组传给后代。许多细菌都含有异源的 DNA 片段。
当细菌处于温度、湿度、空气、营养等丰富的环境中时,会快速繁殖,呈指数级增长,可以形成肉眼可见的集合体,例如菌落(colony)。
有些细菌可以形成芽孢结构,芽孢能够耐受高温、干旱、强辐射等极端恶劣,有利于其度过严峻的环境,保持自身的延续。
1.3.3 进化(演化)
现今的细菌是从 40 亿年前的单细胞生物演化而来。在此后的 30 亿年间,细菌和古细菌都是主要的生物。虽然细菌有化石存在,如叠层石等,但这些化石缺乏有效的形态学证据,很难与现生的细菌共同建构出细菌的演化史。幸运的是,日益成熟的基因定序技术让我们有机会建立演化的树状图,这些研究使我们明了了细菌演化的第一次大分歧是在真核及原核之间之后,细菌又发生了第二次的剧烈演化,有一部分的古细菌与其他细菌内共生,成为了现今真核生物的祖先。真核生物的祖先吞下了一种 α- 变形菌门
的细菌,成为后来的线粒体,或是氢酶体。之后,有些已经拥有线粒体的生物,吞下了类似蓝菌类的生物,形成了后来的叶绿体,这一支后来演化成了藻类和植物。另外,有些藻类还有可能再吞入其他藻类进行内共生,此现象称为二次内共生。
1.4 治疗手段及顽强程度
1.4.1 治疗手段
很多致病细菌均可通过抗生素治疗,现阶段人类经常用的青霉素,头孢类,万古霉素,红霉素,四环素…… 都属于抗生素类
1.4.2 顽强程度(适应环境的能力)
细菌耐高温,耐高压,耐盐碱(嗜盐菌),对于极端恶劣环境适应能力极强,有人发现火山口,海底热泉附近都有细菌存在,细菌分布极为广泛,从大气层上界到海底均有分布。甚至外太空及外星球都有细菌分布。一般来说灭菌彻底要这样做:在 121 摄氏度,0.1 兆帕的环境下灭菌 21 分钟才能杀死细菌芽孢,或者直接在火焰上炙烤才能彻底灭菌。
2、病毒
2.1 病毒的定义
病毒(virus)是由一个核酸分子(DNA 或 RNA)与蛋白质构成的非细胞形态,靠寄生生活的介于生命体及非生命体之间的有机物种,它既不是生物亦不是非生物,目前不把它归于五界(原核生物、原生生物、菌物、植物和动物)之中。
看见了吧,这是比较科学的病毒的定义,病毒是不是生物还没确定!
另外该定义来自 [维基百科](https://zhida.zhihu.com/search?q = 维基百科 & zhida_source=entity&is_preview=1),是比较老的定义,现在人们已经发现了只有蛋白质的病毒种类 —— 朊病毒**。**
2.2 大小形状及分类
2.2.1 大小
大多数病毒的直径在 10-300 纳米(nm)。一些丝状病毒的长度可达 1400nm,但其宽度却只有约 80nm。大多数的病毒无法在光学显微镜下观察到,而扫描或透射电子显微镜是观察病毒颗粒形态的主要工具,常用的染色方法为负染色法。病毒比细菌小多了。二者相差约 1000 倍。
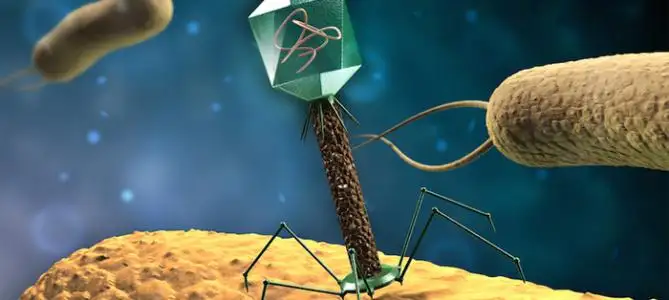
这是正在感染的噬菌体(病毒的一种),看得出来,就是战斗机和航母的差别。(实际比这还要大)。
2.2.2 形状(具体不再详述,看看图吧)
2.2.2.1 螺旋形
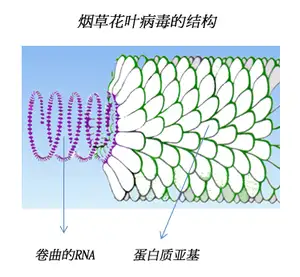
2.2.2.2 正十二面体
大多数动物病毒
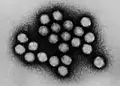
2.2.2.3 包膜型
流感,HIV,带状疱疹
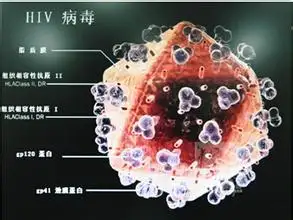
2.2.2.4 复合型
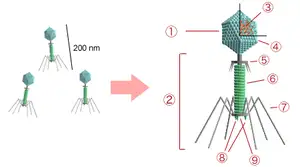
噬菌体
一个典型的有尾噬菌体的结构:①头部,②尾部,③核酸,④头壳,⑤颈部,⑥尾鞘,⑦尾丝,⑧尾钉,⑨基板
2.2.3 分类
按照感染源不同分为:
2.2.3.1、DNA 病毒
以单链或双链 DNA 为基础加蛋白质外壳组成的病毒,如天花,登革热
2.2.3.2、RNA 病毒
以单链或双链 RNA 为基础加蛋白质外壳组成的病毒,如烟草花叶,HIV
2.2.3.3、蛋白质病毒
只有蛋白质为基础,如朊病毒
2.2.3.4、纯感染性 RNA
后来人们发现某些裸露的小片段 RNA 也有感染能力,后来发明了 RNAi
还有其它分类方法,参见《微生物学》沈萍著
2.3 进化(演化)
对于病毒是如何演化或者说进化而来的,科学界存在广泛争议,甚至对于病毒是否能归为生物也说法不一,目前有代表的假说认为:(只是假说,没有事实证据或证据很少)
逆向理论(Regressive theory):病毒可能曾经是一些寄生在较大细胞内的小细胞。随着时间的推移,那些在寄生生活中非必需的基因逐渐丢失。这一理论的证据是,细菌中的立克次氏体和衣原体就像病毒一样,需要在宿主细胞内才能复制;而它们缺少了能够独立生活的基因,这很可能是由于寄生生活所导致的。这一理论又被称为退化理论(degeneracy theory)。
细胞起源理论(有时也称为漂荡理论):
一些病毒可能是从较大生物体的基因中 “逃离” 出来的 DNA 或 RNA 进化而来的。逃离的 DNA 可能来自质粒(可以在细胞间传递的裸露 DNA 分子)或转座子(可以在细胞基因内不同位置复制和移动的 DNA 片断,曾被称为 “跳跃基因”,属于可移动遗传元件)。转座子是在 1950 年由巴巴拉・麦克林托克在玉米中发现的。
共进化理论:病毒可能进化自蛋白质和核酸复合物,与细胞同时出现在远古地球,并且一直依赖细胞生命生存至今。
看见了吧,连病毒怎么来的都不知道。病毒起源于何时尚不清楚,因为病毒不形成化石,也就没有外部参照物来研究其进化过程,同时病毒的多样性显示它们的进化很可能是多条线路的而非单一的。 分子生物学技术是目前可用的揭示病毒起源的方法;但这些技术需要获得远古时期病毒 DNA 或 RNA 的样品,而目前储存在实验室中最早的病毒样品也不过 90 年。
另外关于命名也没有细菌系统:
病毒的命名并无绝对的规则,常依病毒的型态、感染对象、最初发现地点。例如感染动植物的病毒可能依感染的对象、病征来命名,例如麻疹病毒、狂犬病毒,以发现地点命名的包括埃博拉病毒。噬菌体的命名常依实验室内编号命名,例如 T1 噬菌体。
2.4 繁殖(增殖)方式
由于病毒是非细胞的,无法通过细胞分裂的方式来完成数量增长;它们是利用宿主细胞内的代谢工具来合成自身的拷贝,并完成病毒组装。病毒并不是严格生物学意义上的繁殖,而且每次释放个体数量巨大,故而称其为 —— 增殖。不同的病毒之间生命周期的差异很大,但大致可以分为六个阶段:附着,入侵,脱壳,合成,组装,释放。详细不再赘述,前面已有人回答。
2.5 顽强程度
因为就是蛋白质等大分子,所以体外很脆弱,怕热不怕冷,怕湿不怕干,一般 55-60 摄氏度,加热 13 分病毒就会变性死亡(参考煮鸡蛋,鸡蛋清变性),表面活性剂(洗衣粉),氧化剂(84 消毒液)亦能有效抑制或杀死病毒。一般病毒可保存于零下 196 度的液氮环境中,数年后依然有感染能力。湿度达到 50%~60% 基本病毒就会被水气沉降。所以冬天下场雪流感就少很多,干冷就容易爆发流感。
2.6 治疗手段
2.6.1 绝大多数病毒的感染我们束手无策!
目前(2023 年)随着技术的进步,我们有了抵抗流感和部分病毒的专用药物
2.6.2 已知的有效手段
2.6.2.1 盐酸吗啉胍(病毒灵)
本品能抑制病毒的 DNA 和 RNA 聚合酶,从而抑制病毒繁殖。在人胚肾细胞上,1% 浓度对 DNA 病毒 (腺病毒,疱疹病毒) 和 RNA 病毒 (埃可病毒) 都有明显抑制作用,对病毒增殖周期各个阶段均有抑制作用。对游离病毒颗粒无直接作用。
2.6.2.2 利巴韦林(病毒唑)
利巴韦林为合成的核苷类抗病毒药。利巴韦林对呼吸道合胞病毒(RSV)具有选择性抑制作用。利巴韦林是一种前体药物 ,当微生物遗传载体类似于嘌呤 RNA 的核苷酸时,它会干扰病毒复制所需的 RNA 的代谢。它究竟如何影响病毒的复制,尚不清楚。
2.6.2.3 磷酸奥司他韦(达菲)
磷酸奥司他韦是其活性代谢产物的药物前体,其活性代谢产物(奥司他韦羧酸盐)是强效的选择性的流感病毒神经氨酸酶抑制剂。神经氨酸酶是病毒表面的一种糖蛋白酶,其活性对新形成的病毒颗粒从被感染细胞中释放和感染性病毒在人体内进一步播散至关重要。
磷酸奥司他韦的活性代谢产物能够抑制甲型和乙型流感病毒的神经氨酸酶活性。在体外对病毒神经氨酸酶活性的半数抑制浓度低至纳克水平。在体外观察到活性代谢产物抑制流感病毒生长,在体内也观察到其抑制流感病毒的复制和致病性。
本品通过抑制病毒从被感染的细胞中释放,从而减少了甲型或乙型流感病毒的播散。
2.6.2.4 法维拉韦(Favipiravir)
6 - 氟 - 3 - 羟基吡嗪 - 2 - 甲酰胺,英文名 Favipiravir,法维拉韦在机体细胞内经酶代谢为活性形式法维拉韦核苷三磷酸。法维拉韦核苷三磷酸可竞争性抑制流感病毒 RNA 依赖的 RNA 聚合酶。法维拉韦核苷三磷酸在 1mM 浓度下对人 RNA 聚合酶 α 无抑制作用,对人 RNA 聚合酶 β 和 γ 的抑制作用分别为 9.1〜13. 5% 和 11. 7〜 41.2%。法维拉韦核苷三磷酸对 RNA 聚合酶 Ⅱ 的 ⅠC50 值为 905μM。
2.6.2.5 瑞德西韦(Remdesivir)
Remdesivir(GS-5734)是氰基取代的腺苷核苷酸类似物前药,显示出对几种 RNA 病毒的广谱抗病毒活性。
在 Remdesivir抑制埃博拉病毒活性的实验当中,发现 Remdesivir 是前药,其活性分子为三磷酸化后的(Remdesivir-TP)形式。
2.6.2.6 干扰素
Interferons. Another group ofproteins that provide protectionare the interferons, whichinhibitthe replication of many—but not all—viruses.(抑制部分病毒的复制) Cells that have been infected with a virus produce interferon, which sends a signal to other cells of the body to resist viral growth.
干扰素是最早的抗病毒药物,他在丙型肝炎
中的主导地位目前没有任何药物能够撼动,干扰素联合利巴韦林是国际通用的抗 HCV 方案,特别是聚乙二醇干扰素联合利巴韦林,对于难治性丙型肝炎,一些蛋白酶抑制剂等将要相继上市,但大多也要和干扰素联合。
2.6.2.7 Paxlovid
Paxlovid 由 Nirmatrelvir(奈玛特韦,PF-07321332)和 Ritonavir(利托那韦)两部分组成,真正的活性成分是 Nirmatrelvir,是新冠病毒的 3CL 蛋白酶抑制剂,而 Ritonavir 在这里的作用是用于维持 Nirmatrelvir 的血药浓度。
2.6.2.8 玛巴洛沙韦
玛巴洛沙韦是一款创新的帽状结构依赖性核酸内切酶抑制剂,在进入体内后经芳基乙酰胺脱乙酰酶(AADAC)水解生成具有抗流感病毒作用的活性成分 - 巴洛沙韦。 巴洛沙韦通过与 Cap - 依赖性核酸内切酶结合,阻断病毒 mRNA 的合成,从而抑制病毒增殖,发挥抗病毒作用。与奥司他韦需要 5 天的用药周期相比,玛巴洛沙韦全程只需一次服药就能使病毒在 24 小时内停止排毒,缩短传染期并大幅减少流感症状持续时间。此外,玛巴洛沙韦还能够治疗对奥司他韦产生抗性的病毒株和禽流感病毒株。
其原研药企为日本盐野义,2016 年,罗氏制药
与盐野义达成合作,加入玛巴洛沙韦的研发和商业推广工作;2018 年 2 月和 10 月,该药先后在日本和美国获批上市;2021 年 4 月,成功登陆中国,商品名为速福达,适应症为用于治疗 12 周岁及以上急性无并发症的流感患者;同年 12 月成功进入新版医保目录。
林鱼儿:抗流感新药玛巴洛沙韦(速福达)

大部分都是抑制,根本就不能干掉,还有过程不明,但随着计算机辅助制药的进步,我们目前具有了专杀某些病毒的口服药。
PS:为什么感冒输液能好?
细菌病毒经常组团到来,感冒其实是复合感染。抗生素干掉了细菌,帮您节约了弹药,抗体自己干掉了病毒。自身抗体是干掉病毒的唯一有效途径!
PS:感冒吃药有什么用?
相当于暂时性吸毒,缓解各种症状,使你不那么痛苦!我没危言耸听!盐酸布洛伪麻片(康泰克)本质是神马?就是大麻(类似物)!磷酸可待因本质是神马?就是海洛因(不成熟提取物)!只不过量少罢了。甘草片是神马?阿片粉,本质就是大烟粉!所谓止咳缓解鼻塞的道理明白了吧~
2024 年更新:
新一代感冒药如磷酸奥司他韦和玛巴洛沙韦,具有 “杀灭” 流感病毒的作用 —— 虽然也是竞争和抑制,但已经有确切的治疗作用了,并非只是缓解症状。
3、支原体、衣原体、立克次氏体
其实除了细菌和病毒还有一些介于二者之间无法归类的原核生物,有时他们的致病性一点儿也不比二者差。
支原体:支原体是 1898 年 Nocard 等发现的一种类似细菌但不具有细胞壁的原核微生物,能在无生命的人工培养基上生长繁殖,直径 50-300nm,能通过细菌滤器。过去曾称之为类胸膜肺炎微生物(pleuropneumonia-like organism,PPLO)。1967 年正式命名为支原体。
支原体(mycoplasma):又称霉形体,为目前发现的最小的最简单的原核生物。基因数量为 480。支原体细胞中唯一可见的细胞器是核糖体(支原体是原核细胞,原核细胞的细胞器只有核糖体)。
衣原体:衣原体为革兰氏阴性病原体,是一类能通过细菌滤器、在细胞内寄生、有独特发育周期的原核细胞性微生物。过去认为是病毒,现认为是介于立克次体和病毒之间的微生物。衣原体广泛寄生于人类、鸟类及哺乳动物。能引起人类疾病的有沙眼衣原体、肺炎衣原体、鹦鹉热肺炎衣原体。
立克次氏体:
1909 年,美国病理学家霍华德・泰勒・立克次(Howard Taylor Ricketts)(1871-1910 年) 首次发现洛基山斑疹伤寒的独特病原体并被它夺取生命,故名。
与其它细菌不同,立克次氏体只能在其它生物体(宿主)的细胞内生存和繁殖,而不能在自然界环境中生存。这些细菌中的许多种类生活在称为宿主的小型动物(如大鼠和老鼠)中。贝纳特氏立克次体以牛、绵羊或山羊为宿主,可导致 Q 热。普氏立克次体**以人类为常见宿主,可导致流行性斑疹伤寒。**作为储存宿主的动物感染后可以发病也可不发病。**立克次体常常通过先前寄生于感染动物的蜱、螨、蚤和虱(媒介)叮咬人而传播。蜱、螨、跳蚤和虱子被称为媒介,是因为它们将引起疾病的生物体从一个宿主传播(传染)到另一个宿主。**由贝纳特氏立克次体引起的 Q 热可通过空气或食物传播。每一种立克次体和相关细菌都有它自己的宿主和媒介。其中有一些立克次体(以及由其导致的疾病)发生在全世界各地。其它则仅发生在某些地理区域。在人体中,立克次体感染小血管壁的细胞,引起起血管的炎症和阻塞或向周围组织渗血。其它细菌(埃利希氏体属和无形体)进入 白细胞。损伤发生部位和身体的反应方式决定了症状的类型。(此段参考《默沙东诊疗手册》)
氯霉素和四环素类抗生素
对各种立克次体均有很好效果,能明显缩短病程,大幅度降低病死率。但某些立克次体病
的复发日渐增多,可能为药物未能杀死所有病原体的缘故。病原体的最终清除仍有赖于机体免疫机能(尤其是细胞免疫)。
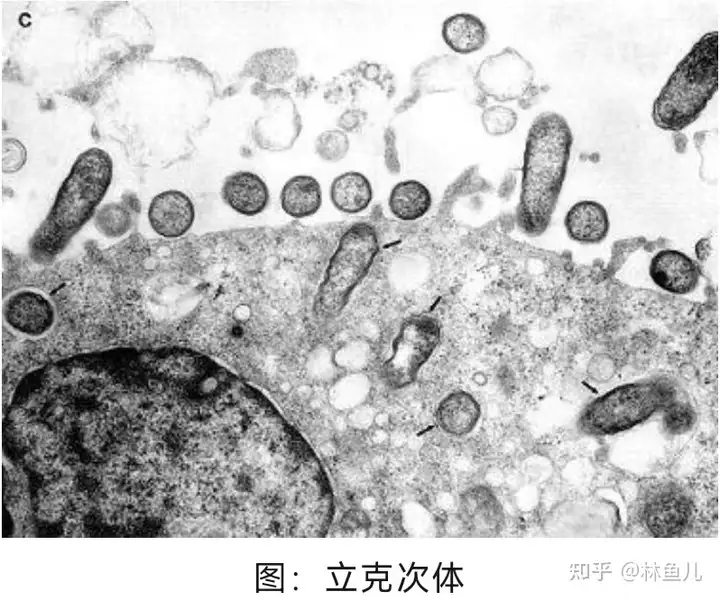
立克次氏体
比如撸猫感染立克次氏体的例子:
-
“撸猫” 需当心感染立克次体 – 健康・生活 – 人民网
http://health.people.com.cn/n1/2020/1125/c14739-31943301.html
家猫及时驱虫(内驱和外驱),是没有问题的,但野外的流浪猫要小心。
+++++++++++++
参考
微生物学是一门很广博的知识学科,前面是最简单的概括。
1. 维基百科相关词条
2.《微生物学》沈萍著
3.《伯杰氏鉴定细菌学手册》
(1923 年第 1 版,后于 1925、1930、1934、1939、1948、1957、1974 年相继出版了第 2 至第 8 版,每个版本都反映了当时细菌学发展的新成就。其中第 8 版有美、英、德、法等 14 个国家的细菌学家参加了编写工作,对系统内的每一属和种都做了较详细的属性描述。近年来,由于细胞学、遗传学和分子生物学的渗透,大大促进了细菌分类学的发展,使分类系统与真正反映亲缘关系的自然体系日趋接近。第 8 版(1984,1986)中实质性的变化,象征着细菌分类学的发展进入新的阶段。第一,手册更名,原书名为《伯杰氏鉴定细菌学手册》(Bergey’s Manual of Determinative Bacteriology),第 8 版由于内容增加,范围扩大,提高了手册的实用性,同时指出各类细菌间的关系,所以改名为《伯杰氏系统细菌学手册》;第二,由 1 卷分成 4 卷,这是考虑到能及时反映新进展和使用者的方便;第三,细菌在生物界的地位,8、9 版间无变动,但它们的高级分类单位有很大变化(见下表),尤其嗜盐细菌和产甲烷细菌,根据胞壁分析和 DNA 序列分折,另列疵壁菌门,古细菌纲;第四,趋近自然体系,在各级分类单位中全面应用核酸研究;在表型特征的基础上,以 DNA 资料给予决定性的判断。使人为的分类体系过渡到自然体系的理想进一步付诸实现。)
4. 默沙东诊疗手册
-
《默沙东诊疗手册大众版》
https://www.msdmanuals.cn/home -
MSD诊疗手册专业版
https://www.msdmanuals.cn/professional
病毒和细菌到底有什么异同?为啥不能用抗生素杀病毒?
入侵人体,细菌和病毒有不同的途径
细菌中有许多 “好人”,其中腐生细菌是生态系统中重要的分解者,使碳循环能顺利进行;部分细菌会进行固氮作用,使氮元素得以转换为生物能利用的形式。乳酪及酸奶和酒酿的制作、部分抗生素的制造、废水的处理等,都与细菌有关。在生物科技领域中,细菌也有着广泛的运用。当然,细菌中的 “坏人” 就是许多的病原菌了,包括肺结核、炭疽病、鼠疫等疾病都是由细菌所引发。
细菌和人体主要为寄生关系,咱们主要说说致病菌,由于大多数细菌有自己的代谢系统,细菌入侵人体后是可以寄生在人体正常细胞之间的。细菌夺取了机体所必须的营养物质;细菌的生长产生各种各样的代谢产物打乱了机体的生理平衡;甚至细菌的体积也成为致病的因素,干扰和破坏细胞的功能,因此,在一些疾病中,仅细菌的增殖就造成致命的后果。
病毒入侵人体可能是来源于一次打喷嚏,或者一次身体接触。病毒需要寄生在活的宿主细胞之内,依赖于宿主细胞提供病毒复制过程中所需要的原料体系、能量和场所。当一个病毒准备感染宿主细胞时,其需要以下六个步骤完成其增殖活动,分别是吸附、侵入、脱壳、生物合成、组装和释放。
-
吸附:病毒通过识别宿主细胞膜表面特有的受体蛋白分子来 “盯上” 目标细胞(比如新型冠状病毒识别的是人呼吸道和肺部细胞表面的血管紧张素转化酶 2-ACE2);
-
侵入:然后病毒要么通过某种方式进入宿主细胞(如膜融合),要么直接将遗传物质注入宿主细胞之内;
-
脱壳:紧接着病毒感染性核酸从衣壳内被释放出来;
-
生物合成:“马不停蹄” 地进行生物合成 —— 根据基因指令,并借助宿主细胞提供的原料、能量和场所来合成病毒的核酸和蛋白质;
-
组装:新合成的病毒核酸和蛋白质会组装成子代病毒;
-
释放:子代病毒释放到宿主细胞外。
细菌和病毒同是微小生物,二者大小相差约 1000 倍
细菌和病毒都是微生物。微生物作为地球上最小的生命形式,人们总是忽略它们的存在,可就是这微小的生物时时刻刻刷着 “存在感”,人类一不留神就会被它们感染。
细菌是细胞,有细胞壁,有 DNA,有细胞器,可以自行生产合成需要的酶并且代谢,可以自行分裂繁殖。而病毒比细菌小很多了,主要结构是蛋白质衣壳和内部的遗传物质(DNA 或者 RNA),而且病毒不能自我复制。病毒需要通过感染宿主细胞来复制自身的遗传物质,然后释放出更多的子代病毒去感染其他的宿主细胞。
细菌可以是无害甚至有益于人体健康的,是可以独立生存的。而病毒存在的目的就是复制自己,所以它不感染宿主就没有存在的意义,可谓是最纯粹的 “自私的基因”。
广义的细菌即为原核生物,虽为细胞结构,但和人的细胞结构还是有很大差异的,它们结构更加简单,没有细胞核,只有 DNA 的聚集区 —— 拟核。细菌一般为球状、杆状、螺旋状等等,人们给它命名的时候也不忘加上形状的描述,比如大肠杆菌、乳酸杆菌、金黄色葡萄球菌等等。
目前已知最小的细菌只有 0.2 微米长,因此只能在显微镜下看到它们;而世界上最大的细菌可以用肉眼直接看见,有 0.2-0.6 毫米大,是一种叫纳米比亚嗜硫珠菌的细菌。
病毒(virus)是由一个核酸分子(DNA 或 RNA)与蛋白质构成的非细胞形态,靠寄生生活的介于生命体及非生命体之间的有机物种,它进入细胞后表现的 DNA 的复制等新陈代谢确实是生命体的特征,而离开细胞后它只是一个没有生命的结晶体。大多数病毒的直径在 10-300 纳米(nm)一些丝状病毒的长度可达 1400nm,但其宽度却只有约 80nm。大多数的病毒无法在光学显微镜下观察到,而扫描或透射电子显微镜是观察病毒颗粒形态的主要工具。病毒和细菌相比,二者的大小相差约 1000 倍。
积极调动人体自身免疫力对抗病毒
不论细菌还是病毒都要突破人体的重重防线才能够完成入侵,但人体终将发现它们的存在,人类的免疫系统有着自己的防护措施和预警机制。
第一道防线:皮肤和黏膜。人类保护自己的方式首先是防御,通过皮肤和黏膜使人体形成了一个相对密闭的系统,当有害物质将要侵入人体时,皮肤和黏膜将外界致病因素阻挡在体外。
第二道防线:杀菌物质和吞噬细胞。黏膜表面和人体内部总是有着杀菌物质和吞噬细胞在 “巡逻”,防范病原体的入侵。以溶菌酶来举例,它能够破坏细菌的细胞壁,导致细胞壁破裂内容物逸出而使细菌溶解,还可与带负电荷的病毒蛋白直接结合,与 DNA、RNA、脱辅基蛋白形成复盐,使病毒失活。因此,该酶具有抗菌、消炎、抗病毒等作用。
第三道防线:特异性免疫。当强大的病原体突破了前两道防线后,人体的反攻才刚刚开始。通过吞噬细胞的吞噬和特殊的免疫细胞(T 细胞)的分析,人体生产出了可以特异性识别入侵物的抗体,抗体可以让病原体粘连在一起不再具有入侵性,最终暴露在细胞间的病原菌会被杀灭。但入侵了细胞的病毒是否安全了呢?非也!人体可以精妙地识别哪个细胞被病毒感染了,进而派出 “杀手” 杀死被感染的细胞,释放细胞内部的病毒,供抗原消灭。
但,人体的反击是需要时间准备的,病原体就会通过这样的时间间隙大肆入侵,占领人体。
细菌感染一度成为人类最大的敌人。比如当年鼠疫在欧洲叫做 “黑死病”,曾经 3 年间就使欧洲人口减少三分之一;我国现在还将鼠疫和霍乱列为甲类传染病。
直到抗生素的发现和推广,人类才控制了细菌感染的爆发。
抗生素主要对细菌有杀灭作用,通过破坏细菌细胞的结构,如细胞壁、细胞膜、改变内部代谢、阻碍核酸和蛋白质合成等,进而达到杀灭细菌的目的。
但是,抗生素对病毒是无效的!因为细菌和病毒的结构完全不同!
由于细菌拥有细胞壁,还有自己的核酸复制机器和核糖体,所以抗生素只要针对这些靶点设计,就能保证杀伤细菌而对人类副作用很小。但是,病毒没有细胞壁,没有自己的核酸酶,也没有核糖体,它所有的功能都依靠宿主细胞来完成。所以抗生素并不能杀死病毒(我们总不能制造一种杀灭宿主细胞的 “抗生素”)。
理想的抗病毒药物是既能作用于病毒增值周期的某个或几个环节,予以干扰或阻断,又不影响宿主细胞的正常代谢。如常见的药物病毒唑,提供了大量核苷酸类似物,偷梁换柱取代了正常的核苷酸,使病毒失去了复制能力,起到了抑制病毒扩增的作用(但对人体也有很大副作用哦)。另外,抗流感药物奥司他韦是通过阻断子代病毒的释放从而起效的。
针对这次的肺炎疫情,中国科学院武汉病毒所和军事医学科学院的科研人员也在细胞水平上初步筛选出了对新型冠状病毒(2019-nCoV)有较好抑制作用的药物。但面对病毒的治疗,人类始终没有找到像抗生素一样普适性特效药,积极的治疗都是调动人体自身的免疫能力去对抗病毒,因为只有生物本身才真正懂得如何对抗生物。
(作者:钟柯 单位:中国科学院微生物所)
via:
-
专家详说细菌和病毒 ---- 中国科学院 发布时间:2020-02-06
https://www.cas.cn/zt/sszt/kjgzbd/kpjd/202002/t20200206_4733615.shtml
-
科普丨细菌、微生物和病毒有什么不同?2018-06-25 17:35


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









