






急性肾损伤(AKI)通常由缺血、败血症或肾毒性损伤引起,常与死亡率升高和慢性肾病(CKD)风险增加相关。急性肾损伤导致近端小管细胞(PTCs)功能障碍或死亡,触发了一种尚不完全了解的自我修复程序。修复缺陷与长期过渡到慢性肾病相关。
我们进行了轻度至中度缺血-再灌注损伤(IRI)实验,以模拟在多种临床环境中反映肾损伤的反应,包括肾脏移植手术。通过对在IRI后7天(“早期”)和28天(“晚期”)的基因标记损伤近端小管细胞(PTCs)进行单核RNA测序,识别了损伤-修复过渡中具体的基因和通路活动。特别地,我们在晚期损伤阶段识别了Vcam1+/Ccl2+近端小管细胞,具有明显激活的NF-κB、TNF和AP-1信号通路。这些近端小管细胞表现出与衰老相关的分泌表型特征,但没有显示G2/M期细胞周期停滞,与其他报道的肾损伤后适应不良的近端小管细胞有所不同。
命运追踪实验揭示了这些细胞的空间和时间上不同的起源。在伤害发生的皮质-髓质交界处(CMB),大多数Vcam1+/Ccl2+近端小管细胞源自早期复制的近端小管细胞。相反,在皮质区域,只有一部分Vcam1+/Ccl2+近端小管细胞可以追溯到早期修复的细胞,提示次生近端小管损伤的晚期发生部位。
综合这些数据表明,即使是中度缺血再灌注损伤,也会导致持续的损伤,并且这种损伤从CMB区域扩展到皮质区域。剩余的修复失败的近端小管细胞可能是慢性疾病进展的触发因素。**
关键词 急性肾损伤 | 近端小管 | 修复 | 转录组学 | 单核RNA测序
急性肾损伤(AKI)是全球范围内的重大公共卫生问题,年发病率约为1330万例(1)。AKI的主要原因包括缺血-再灌注损伤(IRI),通常与外科手术、败血症和肾毒性药物相关(2)。AKI与医院内死亡率升高、住院时间延长以及更高的医疗费用相关(3)。此外,AKI还可能导致慢性肾病(CKD)的发展,当前约有9.1%的人口受到CKD的影响,并导致每年120万例死亡(4)。肾脏移植患者的肾活检分析发现,肾脏移植患者的转录组和细胞学特征与严重IRI后观察到的相似,提示即使是轻度肾损伤,也可能导致长期的疾病进展(5-7)。迄今为止,尚无药物策略可以治疗AKI或防止AKI向CKD的进展。因此,全面理解AKI的病理生理机制以及驱动AKI向CKD转变的因素是至关重要的。
近端小管细胞(PTCs)是哺乳动物肾脏中的主要细胞类型。上皮近端小管负责维持电解质平衡和液体稳态。PTCs具有高代谢活性,特别容易受到缺血损伤,并在CKD的病理生理中发挥重要作用(2)。事实上,在损伤后,幸存的PTCs可以激活内源性修复过程,重新进入有丝分裂,重建肾小管上皮,恢复肾脏的结构和功能(8-11)。然而,肾脏修复是有限的;即使肾功能在急性损伤后恢复到基线水平,肾脏中的残留炎症和纤维化过程仍可能促进CKD的发展(2,9,12)。动物模型中的研究将疾病进展与PTCs在G2/M期细胞周期停滞相关联,这些细胞分泌促炎症和促纤维化的细胞因子(12,13),但驱动AKI向CKD转变的潜在机制仍待阐明。
单细胞和单核mRNA测序(snRNA-seq)提供了一种无偏的方式来表征与肾损伤和修复相关的转录组特征。在这项研究中,我们采用了针对PTCs的遗传标记策略,专门研究了在中度IRI模型中受损的PTCs。谱系追踪实验揭示了一种具有促炎症、促纤维化、衰老相关分泌表型的细胞类型,这些细胞来源于未能在原发损伤部位修复的PTCs,并可能在皮质损伤部位形成,后者可能是由于初次IRI事件后的损伤扩展所致。这些数据揭示了一种动态的损伤反应,识别了相关细胞类型、细胞通路及分子靶点,为治疗评估AKI向CKD转变提供了潜在的方向。
研究意义
一次急性肾损伤事件会增加慢性肾病(CKD)发展的风险。通过将单核RNA测序与受损近端小管细胞的遗传追踪结合,研究揭示了缺血-再灌注损伤后空间动态演变的损伤反应。未能修复的近端小管导致了一种持续存在的促纤维化、促炎症的 Vcam1+/Ccl2+ 细胞类型,该细胞展现了衰老相关分泌表型,并且NF-κB和AP-1通路的转录激活显著增强,但未见G2/M期细胞周期停滞的迹象。此研究的发现有助于制定改善肾脏修复并防止CKD进展的策略。
结果与讨论
在之前关于急性肾损伤(AKI)到慢性肾病(CKD)进展的长期双侧缺血-再灌注损伤(IRI)模型研究中,我们鉴定了角蛋白-20(Krt20)作为受损近端小管细胞(PTCs)的标记物(5)。为了能够在肾损伤中分离和追踪 Krt20+ 细胞,我们通过CRISPR-Cas9切割小鼠胚胎干细胞中的Krt20,并通过同源定向修复(HDR)生成了Krt20T2A-CE小鼠系,从而生产了胚系嵌合体(如图1A所示)。HDR促使在Krt20表达的细胞中生成CRE-ERT2(CE)融合蛋白,通过在Krt20的羧基末端插入T2A自切割肽。将Krt20T2A-CE小鼠与Gt(ROSA)26Sortm5(CAG-Sun1/sfGFP)Nat小鼠交配,这些小鼠在Cre的存在下会表达绿色荧光蛋白(GFP)与核膜蛋白相连,并进一步称为INTACT小鼠(特定细胞类型中带标签的核的分离,INTACT)(14,15)。此交配使得在肾损伤的近端小管细胞中可以依赖他莫昔芬标记核。
为了模拟在各种非生命威胁性的临床情境中可能发生的中度肾损伤,如肾移植手术过程中,我们测试了不同的缺血时间间隔。在双转基因雄性小鼠中,18分钟的缺血时间间隔导致48小时后基础肌酐和血尿素氮(BUN)水平显著增加(2到3倍)(附录SI,图S1 A和B)。肌酐和BUN水平在IRI后28天恢复到基线水平。对IRI的小鼠在IRI后第5和第6天,或第26和第27天分别注射了他莫昔芬,肾脏在IRI后的第7天(指定为“早期”,n = 2)或第28天(指定为“晚期”,n = 3)收集(图1B)。对于单细胞核RNA测序(snRNA-seq),我们使用荧光激活细胞排序(FACS)分离GFP+核,并通过10× Genomics分析和下一代测序识别snRNA-seq数据。作为对照数据集,我们将我们之前发布的FACS分选的GFP+核的snRNA-seq数据与额外的重复数据结合,后者来自这个小鼠系(包含1个未手术和3个假手术对照小鼠)(16)。
免疫荧光研究证实了Krt20表达细胞的成功标记,标记效率约为34%(图1C和F)。在IRI后的第7天和第28天的数据比较中,标记效率是一致的。大多数标记细胞对近端小管损伤标志物Havcr1呈阳性(图1F和附录SI,图S1E)(17)。尽管从第7天到第28天,标记的Krt20表达的近端小管细胞的总数减少,但这些细胞的主要位置从早期损伤样本中的皮髓交界区(CMB)(附录SI,图S1C中高亮显示的区域)转移到第28天时的外皮质(图1D和E,附录SI,图S1D和F)。IRI和对照的单细胞RNA测序(snRNA-seq)数据集使用Seurat R包进行整合(18),并从主数据集中去除低质量的样本,分析了24,086个核型谱,通过无监督聚类进行分析。最终得到的13个不同的簇通过二维统一流形近似和投影(UMAP)图呈现(图1G)。差异基因表达分析识别了簇特异性的标志基因,并使用已知的肾脏细胞类型、免疫细胞、血管和间质细胞的标志基因对这些簇进行了注释(图1H和附录SI,图S2B)。正如预期,我们的数据集在对照组和IRI组中都强烈富集了近端小管细胞核。在对照和IRI数据集中,少数非近端小管细胞签名的簇很可能代表了在FACS分选过程中对GFP+细胞的相对宽松选择。通过使用已知的标志基因,我们明确识别出三个主要的簇作为近端小管段。另两个密切相关的簇表现出近端小管特异性损伤标志物Havcr1的强表达,并被识别为受损的近端小管细胞(17)。其中一个受损的近端小管簇通过细胞周期基因的富集表达而被区分出来,表明其处于细胞周期状态。受损的近端小管簇以及免疫细胞簇几乎完全来自IRI组的重复实验(附录SI,图S2A)。
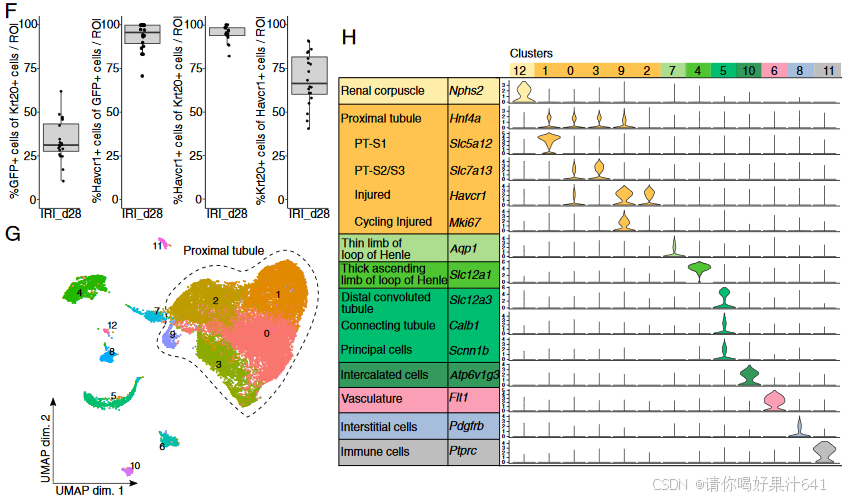
图1. Krt20可诱导Cre-LoxP系统追踪IRI后肾脏中的受损近端小管细胞(PTCs)。
(A) 通过CRISPR将CRE-ERT2(CE)敲入Krt20基因组位点的示意图。
(B) 实验设置。
© IRI样本中GFP报告基因与Krt20和Havcr1的共定位(比例尺,20μm)。
(D) 每个感兴趣区域(ROI)中GFP+细胞的定量分析,每个肾脏切片有五个ROI(n = 2)。
(E) 皮质和皮髓交界区(CMB)每个ROI中GFP+细胞的定量分析,每个肾脏切片有10个皮质和CMB ROI(n = 2)。
(F) 每个ROI中GFP报告基因、Krt20和Havcr1的共定位定量分析,每个肾脏切片有10个ROI(n = 2)。
(G) 综合IRI和对照组单核RNA测序数据集的UMAP图。
(H) 按细胞类型排列的标志基因小提琴图;簇编号标示在图上方。
在D–F中,灰色框表示中间50百分位,水平线表示中位数,胡须表示1.5倍四分位距。***P < 0.001。
为了深入表征不同的近端小管细胞(PTC)状态,我们对所有PTCs(在图1G中标记的)进行了重新聚类分析,共分为12个簇。数据以UMAP图和最具差异表达基因的列表呈现(图2A和数据集S1)。对照组和IRI样本的重复数据一起出现在12个簇中的7个簇(0, 2, 3, 6, 7, 8, 9),而其他5个簇则主要由IRI样本组成,进一步称为IRI簇(1, 4, 5, 10, 11)(图2C)。将可变基因聚类到共同调控的基因模块中,显示出这两组簇之间转录谱的明显区别(SI附录,图S3)。由对照组和IRI样本组成的簇,Hnf4a(一种调控近端小管分化的关键转录因子)及其靶基因Lrp2的表达较高,而主要来源于IRI样本的簇则表达较低(图2B)(19, 20)。这与之前描述的在损伤后正常近端小管转录特征的下调一致(5, 21, 22)。表达Lrp2的簇进一步通过段特异性标志基因(如Slc5a12(S1段)、Slco1a1(S2段)和Slc7a13(S3段))进行细分(图2B)。有趣的是,一些簇显示出正常的S1、S2和S3段特征,但通过不同的近端小管标志基因的富集,显示出PTCs的异质性。例如,S3簇8在IRI样本中显示出Slc7a12的异位激活,这是一种女性特异性的S3段标志基因(23),同时也显示出Cyp7b1等典型的男性S3段特异性基因的表达(图2B)。通过RNAscope原位杂交实验确认了Slc7a12在IRI后男性PTCs中的上调(图3A)。一些Slc7a12+的PTCs对Slc22a7(一种标志女性和男性S3段的标志基因)呈阳性,而其他一些则共表达血管细胞粘附分子-1(Vcam1),这是已知的炎症性肾脏疾病和AKI的标志基因(21, 24)。这些数据表明,Slc7a12的表达在受损的Vcam1+、Slc22a7−的PTCs中上调,并且提示Slc7a12的表达可能在恢复的PTCs中持续存在,恢复Slc22a7并丧失Vcam1。另一方面,Slc7a12也可能在Slc22a7+的细胞中异位激活,而不表现出强烈的损伤特征。通过可搜索数据库(humphreyslab.com/SingleCell/)分析了一个已发布的单细胞RNA测序数据集,该数据集来自单侧输尿管梗阻后14天的雄性肾脏,结果也显示在PTCs中富集了Slc7a12的表达(25)。因此,Slc7a12在雄性肾脏中因AKI激活类似女性的表达模式,并不限于IRI模型。
基因本体(GO)术语分析显示,在每个近端小管簇中差异表达基因的GO术语富集,S1和S2/3簇在所有GO类别中均富集了与正常近端小管结构和功能相关的术语。例如,在生物学过程类别中,“有机阴离子转运”和“药物转运”富集,而在细胞组分类别中,“刷状膜”和“细胞顶端部分”过度代表(图2D和F及数据集S2)。有趣的是,S1簇7和S2/3簇3富集了“呼吸链”、“线粒体内膜”和“细胞色素c氧化酶活性”(数据集S2),这可能表明这些细胞的能量需求增加。Spag5(编码一种有丝分裂纺锤体蛋白,在癌症中的氧化应激反应以及人类足细胞的高葡萄糖处理反应中已被上调(26, 27))在这些簇和簇8中也过度表达(图2B)。在IRI簇中,我们基于簇中IRI样本的主要组成,识别了两个“早期受损”簇、两个“早期+晚期受损”簇和一个“晚期受损”簇(图2B和C)。早期受损簇显示出Havcr1和编码钙粘蛋白的基因Cdh6与Cdh13的富集表达。在正常肾脏中,Cdh6和Cdh13分别标记着构成外髓内条带下行薄支的细胞(皮质和肾髓质交界部的肾小管)(https://cello.shinyapps.io/kidneycellexplorer/)(23)。免疫荧光研究证实了Cdh6和Cdh13在受损的近端小管细胞中的新上调(图3B和C)。早期受损簇10以高表达DNA复制相关基因(如Mki67和Top2a)为特征,表明细胞处于分裂状态(图2B和3D)。强烈的复制反应局限于早期受损簇,这与损伤引发的复制修复一致。早期和晚期受损簇1和4的特征是富集表达Cald1和Pdgfd,而Kcnip4在簇4中特别上调。Cald1编码钙调蛋白1,是平滑肌和非平滑肌收缩装置的肌动蛋白连接调节蛋白,在健康肾脏的成纤维细胞中表达,并已与糖尿病肾病相关(28, 29)。血小板衍生生长因子D(Pdgfd)已知在肾纤维化中发挥作用(30),而Kcnip4编码钾通道相互作用蛋白4,最近被描述为在一种独特的促炎、促纤维化的近端小管细胞状态中上调,该状态在AKI后数周持续存在(21)。与此发现一致,Kcnip4在晚期受损簇11中也有表达,尽管其水平低于簇4。簇11与其他簇的区别在于强烈表达Vcam1和单核细胞趋化蛋白1编码基因Ccl2。Ccl2与小鼠11号染色体上的Ccl7和预测的蛋白编码基因模型Gm17268紧密关联。所有三种转录本都是簇11晚期受损细胞群体的最特异性标志物之一(数据集S1)。通过免疫荧光和RNAscope研究验证了AKI后4周受损肾脏中Ccl2+、Vcam1+、Krt20+细胞的存在(见图5A和B及SI附录,图S4C)。
GO术语分析显示了正常S1、S2/S3簇和IRI簇之间的功能性明显区别(图2D和F及数据集S2)。在IRI簇中,没有任何与正常近端小管结构和功能相关的术语富集,而与上皮状态变化、去分化、迁移和纤维化相关的新术语则过度代表(图2D)。此外,已知在细胞粘附和迁移中起作用的细胞组分,如“肌动蛋白细胞骨架”和“粘附连接”,在IRI簇中富集(图2F和数据集S2)。早期增殖簇显示了细胞周期相关GO术语的高度簇特异性富集,强调了这一簇细胞周期状态的局限性(图2D和数据集S2)。为了进一步了解每个簇中细胞的调控状态,我们使用单细胞调控网络推断和聚类(SCENIC)重建了基因调控网络(图2E和数据集S3)(31)。S1、S2和S3簇显示了与近端小管发育相关的调控因子的活性,包括Hnf4a、Hnf1b和Esrra(19, 20, 32)。这些与核心近端小管功能相关的调控因子的活性在IRI簇中被预测为下调,这与正常近端小管基因表达谱的显著丧失一致。有趣的是,被注释为S1和S3的两个近端小管簇,簇7和簇3,显示出Hsf1(编码热休克蛋白的主转录调控因子,参与应激和癌症过程(33))和Hcfc1(宿主细胞因子C1,一种转录调控因子,依赖于Hsp90稳定性来靶向细胞周期基因(34))的调控因子活性。此外,簇3与IRI簇共享一些调控因子的活性。簇3和簇7可能代表了功能减弱的近端小管细胞,表现为Hnf4a活性降低和应激转录状态。
增殖中的早期受损细胞显示出E2f3调控因子活性,与这些细胞的复制状态一致。基因调控网络分析进一步表明,Hippo信号通路可能在早期受损细胞中起作用:Tead1调控因子的上调,Tead1是Hippo信号处于非激活状态时的核效应因子,与细胞增殖相关。Hippo信号通路已知能响应上皮组织中张力和机械约束的变化(35),通过调节细胞增殖和凋亡来控制器官发育和生长。YAP和TAZ是关键的转录共激活因子,它们与Tead DNA结合伙伴一起发挥作用,已被证明在囊性肾病进展和AKI后肾纤维化的发展中起重要作用(36)。与Hippo信号在早期受损细胞中的参与一致,差异基因表达分析显示与YAP去激活和Hippo信号传导相关的调节因子(例如Sav1、Stk3)的上调,以及Hippo信号沉默时转录反应效应因子(例如Yap1、Tead1)和知名的YAP靶基因如Axl和Ctgf的上调。SCENIC还预测了IRI簇特异性的调控因子活动,如Nfat5,它是一种转录因子,介导细胞对高渗透压和缺氧的应答,已被描述为通过减少半胱天冬酶-3依赖性凋亡减轻IRI后的肾损伤(38)。Nfat5的表达在所有IRI簇中均有富集(数据集S1)。簇11中的晚期受损细胞显示出与NF-κB(Nfkb1、Nfkb2、Rela、Relb、Bcl3)和激活蛋白-1(AP-1;Jun、Junb、Jund和Fosl2)信号通路相关的调控因子活性的强烈上调。类似的基因调控网络已在许多人的癌症中被描述,进一步证明这些细胞存在促炎状态(39)。此外,Smad3调控因子的活性也显示出上调,Smad3是由转化生长因子-β(TGF-β)激活的细胞内信号转导因子,提示这些近端小管细胞可能也响应TGF-β信号通路,TGF-β与驱动肾纤维化相关,主要作用于肾小管旁的系膜细胞和邻近间质细胞类型(40, 41)。IRI簇还显示了Myc调控因子的活性,Myc编码原癌基因c-Myc,已知其与小管细胞凋亡有关,并且Tfcp2l1的活性上调,后者与肾小管发育相关(42, 43)。

















 1103
1103

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









