










转自:http://blog.sciencenet.cn/blog-815628-1054663.html
本文来自倪乐意科学网博客。
链接地址:http://blog.sciencenet.cn/blog-815628-1054663.html
在所有的感觉信息中,视觉机制可能是最复杂的了(图1)。我们每个人都能轻而易举地欣赏大自然的美景——青翠的草木、飞舞的蝴蝶、苍茫的白雪……但我们并不知晓大脑是如何浮现曾经目睹过的外部世界的各种图像的。
在美国哈佛大学的两位神经生物学家——美裔加拿大人休伯尔(David Hunter Hubel)与瑞典人维泽尔(Torsten Nils Wiesel)自1958年开始对视觉机制进行了长达25年的合作研究,共同获得了1981年诺贝尔生理或医学奖。他们首次用微电极研究外侧膝状体和视皮层神经细胞感受野,并提出视觉信息是通过三条独立的通道进行加工的。

David Hunter Hubel (1926-2013)
他们将动物(猫)麻醉,将头固定于立体定向头架中,将一个细金属电极插入到视皮层(visual cortex)中,并尽量靠近(不损伤细胞膜)单个神经细胞或纤维以检测由神经冲动产生的电流。同时使猫的眼睛保持张开并阻止眼球转动。将猫面向几米外的屏幕,使用幻灯机在屏幕上投射与背景不同几何性质的图形(如线条等)(图1)。通过利用这样的装置来研究动物神经细胞对不同类型光刺激的反应(电信号)。
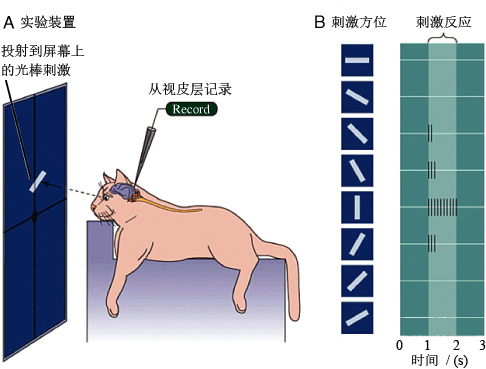
图1 Hubel & Wiesel的实验示意图(引自Purves et al. 2004)
他们在反复多次的实验中发现认,视皮层中的神经细胞对光点或大面积弥散光刺激并无反应,但却在一次偶然中惊喜地观察到,这些细胞对一定朝向(或方位)的亮暗对比边、光棒或暗棒反应强烈(产生密集的电信号),但若偏离该细胞“偏爱”的最优方位,细胞反应停止或骤减。他们发现,绝大多数视皮层细胞都具有强烈的方位选择性,各个细胞的感受野位置连续地发生漂移(图2),即最优方位大致以10度/50μm的变化率按顺时针或逆时针方向发生连续变化,有时在旋转90~270度以后,旋转方向发生逆转。
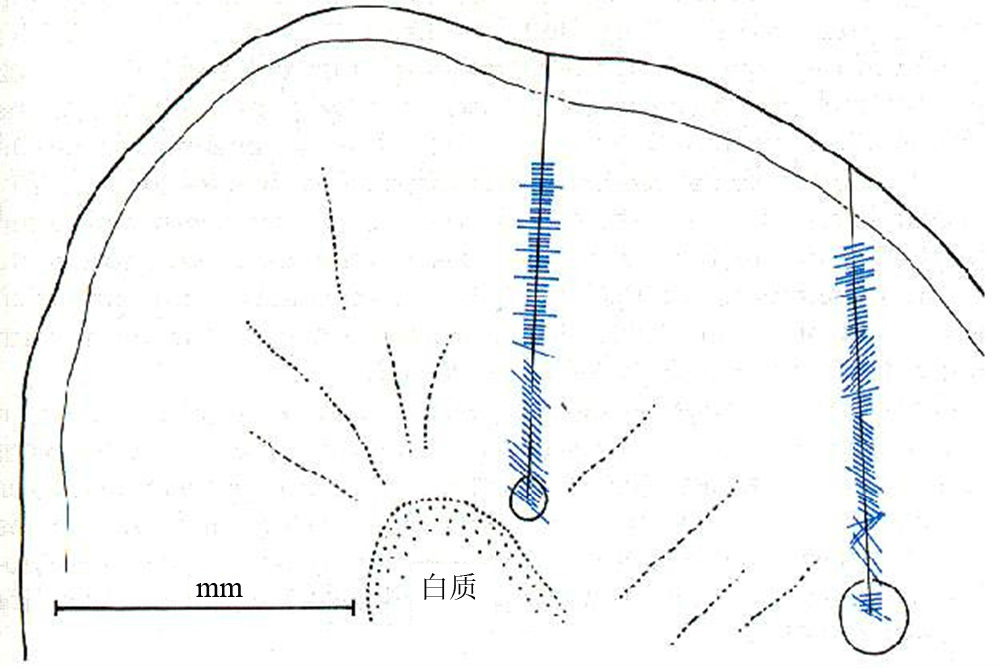
图2 1958年发现功能柱的实验,垂直和倾斜穿刺连续纪录得到的细胞最优方位分布,短线的长度代表该细胞反应的强弱,短线的朝向代表每一纪录到细胞的最优方位(引自Hubel and Wiesel 1962)
他们注意到,视觉信息在经由视网膜感受野→外膝体感受野→视皮层的传递过程中,视神经细胞的感受野(receptive field)发生了质的变化。其实,感受野并不神秘,譬如,视觉感受野就是指影响视觉神经元的刺激区。他们发现视网膜上神经节细胞的感受野与外膝状体神经元上的感受野是一一对应的,两种神经元对光点照射均呈现中心与周边相互拮抗式的响应模式(同心圆状的感受野),但是,视皮层神经元的感受野对应视网膜上的一个更大的区域,因为它是由若干个外膝体细胞的感受野共同会聚到一个视皮层细胞的感受野上的(图3)。他们宣称,简单细胞的感受野再汇聚成复杂细胞的感受野,后者再进一步汇聚成超复杂细胞的感受野。但笔者认为,感受野充其量只是一种电生理响应(发放),只能反映神经活动的一个有限的侧面。
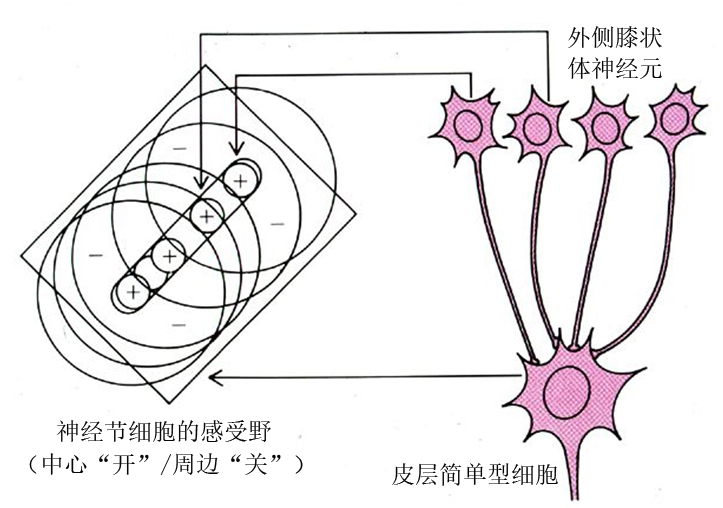
图3 简单细胞感受野与外侧膝状体神经元和神经节细胞感受野的关系(引自Hubel and Wiesel 1962)
之后,感受野的研究开始疯狂,因为Hubel and Wiesel(1962)的这篇文章(发表在The Journal of Physiology)的google引用已超过12000次!人们对类似实验重复的热情使我感到震惊。尤其令我惊讶的是,迄今为止,无人能够回答为何感受野会发生这样的变化!在生命科学领域中,类似的不可思议的事情司空见惯,譬如,人们对遗传密码子、光合作用和生化循环等的工作原理描绘得栩栩如生,但却一点都不知道它们是怎么来的。
20多年之后,休伯尔与其学生在Science上的一篇综述论文(Livingstone and Hubel 1988)中指出,我们的大脑会把一个视觉场景划分为各个子维度,例如颜色、深度、运动和形状,然后对各个维度同时加工(图4)。Hubel(1988)深信在视觉过程中,信息是先被分解后被整合起来了的,这被称之为视觉信息的加工。但他又无奈地感叹道,像形状、颜色与运动等特征由不同的脑部位所负责,这就产生了一个问题,即不同的信息是如何组合在一起而形成例如跳跃的红球这样的知觉的呢?除了负责捕捉球的运动神经之外,它们显然地要在脑中某个地方进行组合,但关于在哪里组合以及如何组合,我们则一无所知。
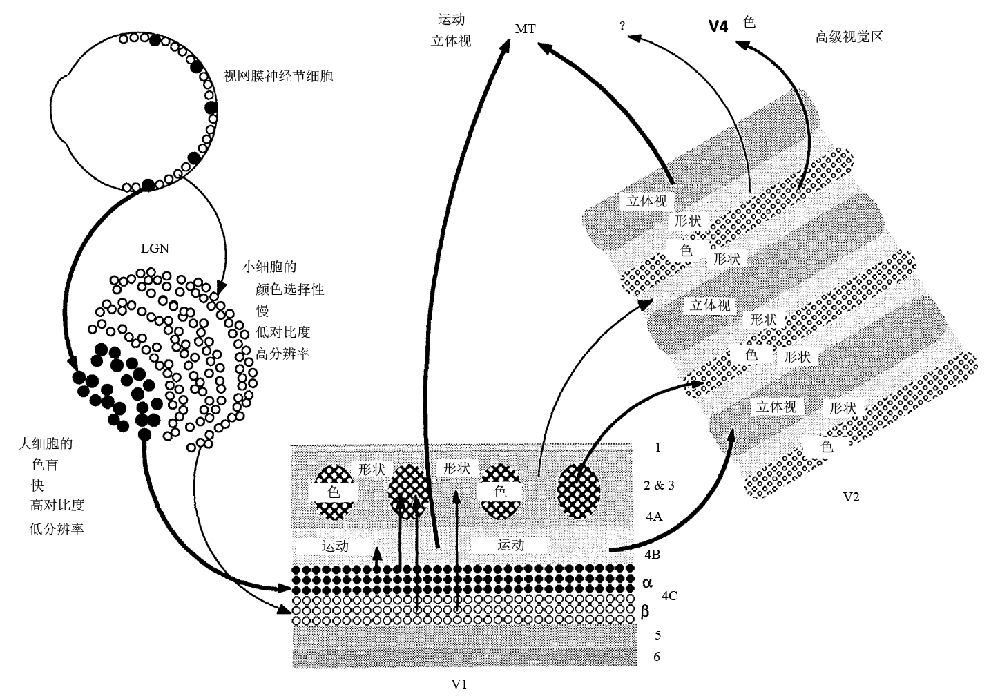
图4 猴子的V1视区和V2视区内加工形状、颜色、运动和深度知觉信息的分离处理通路(LGN:外侧膝状体,MT:颞中区)(引自Livingstone and Hubel 1988)
自休伯尔和维泽尔于1958年开始进行的开拓性的研究之后,已经过去了半个多世纪。最近,在Nature Reviews Neuroscience 的1篇综述论文之中,Nassi and Callaway(2009)指出,“并行处理是视觉系统的一个独有的特征,十几种类型的神经节细胞将射入的视觉信号解析与导入到在功能和解剖上特化的通道中,再平行地投射到LGN,再到V1。覆盖在视网膜上的这些神经节细胞提供一个可以传递到大脑的整个特征性视野的完整表征。一旦到了V1,这些平行的输入通道被整合成模块,这些模块具有空间定义以及局域连接,因此,形成一些新的平行信息通道,传送到大脑的其它区域。从V1和V2的输出进入了纹外皮层中的两个相互分离但相互关联的加工路径——外背侧通路和腹侧通路。这两个通路使用了一套相似的视觉属性,但进行了不同的计算,以介导非重叠的行为目的。但在每个通道中,每个纹外皮层区域可能使用了在V1中使用的同样的策略去重组与整合多端输入(multiple inputs),形成输送到下游的新的输出”。然而,依笔者之见,对视觉机制,并未见到革命性进展,虽然人们添加了若干新的视觉通道(图5),并还在继续挖掘中,但对视觉信息在这些通道中的具体传输机制依然是一无所知。
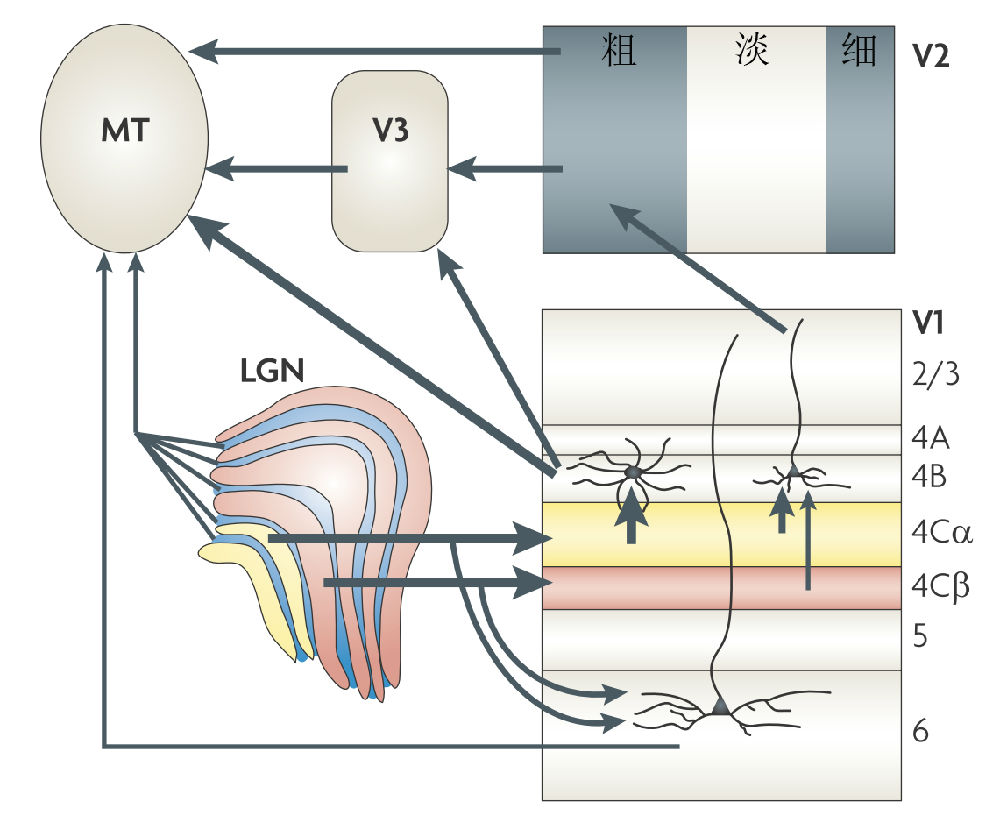
图5 视觉信息从丘脑内侧膝状体(LGN)到颞中区(MT)的多端传输,其中,粗:粗条纹区,细:细条纹区,浅:浅条纹区(引自Nassi and Callaway 2009)
沿着Hubel and Wiesel指引的方向,有人将视觉场景的加工进一步细分为三个层次——低水平加工是简单特征(如方位、颜色、对比度、视差、运动方向等)的分析,中等水平的加工用低水平特征解析视觉场景,包括轮廓整合、表面特征、形状区分、对象运动等,高水平加工就是运用表面和轮廓来辨识对象(Kandel et al. 2013)。
迈尔斯说,“要在头脑中表征这个世界,我们必须识别环境中的物理能量,并且将其编码为神经信号,这个过程通常被称为感觉(sensation)。同时我们必须选择、组织并且解释我们的感觉,而这个过程通常被称为知觉(perception)”(Myers 2004)。但笔者认为,即便在感觉过程中存在神经元产生电活动现象,也难以认定像视觉这样的感觉信息都被编码成了电信号。

















 7万+
7万+

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









