







总目录
-
妮蒂·史蒂文斯对性染色体生物学领域的贡献
-
妮蒂·史蒂文斯在百年前破解性别之谜,却因性别而无缘荣誉
-
性别的起源、流动、进化
-
……
The contributions of Nettie Stevens to the field of sex chromosome biology
妮蒂·史蒂文斯对性染色体生物学领域的贡献
注:机翻,未校。
Philos Trans R Soc Lond B Biol Sci. May 9, 2022; 377(1850): 20210215.
Published online March 21, 2022. doi: 10.1098/rstb.2021.0215
PMCID: PMC8941642
PMID: 35306894
Sarah B. Carey, Conceptualization, Data curation, Formal analysis, Visualization, Writing – original draft, Writing – review & editing, 1 , 2 , † Laramie Aközbek, 1 , 2 , † and Alex Harkess, Conceptualization, Writing – original draft, Writing – review & editing 1 , 2
Author information Article notes Copyright and License information PMC Disclaimer
Associated Data 相关数据
Abstract 抽象
The early 1900s delivered many foundational discoveries in genetics, including re-discovery of Mendel’s research and the chromosomal theory of inheritance. Following these insights, many focused their research on whether the development of separate sexes had a chromosomal basis or if instead it was caused by environmental factors. It is Dr Nettie M. Stevens’ Studies in spermatogenesis (1905) that provided the unequivocal evidence that the inheritance of the Y chromosome initiated male development in mealworms. This result established that sex is indeed a Mendelian trait with a genetic basis and that the sex chromosomes play a critical role. In Part II of Studies in spermatogenesis (1906), an XY pair was identified in dozens of additional species, further validating the function of sex chromosomes. Since this formative work, a wealth of studies in animals and plants have examined the genetic basis of sex. The goal of this review is to shine a light again on Stevens’ Studies in spermatogenesis and the lasting impact of this work. We additionally focus on key findings in plant systems over the last century and open questions that are best answered, as in Stevens’ work, by synthesizing across many systems.
1900 年代初期在遗传学方面取得了许多基础性发现,包括重新发现孟德尔的研究和染色体遗传理论。根据这些见解,许多人将研究重点放在性别分离的发育是否具有染色体基础上,或者它是由环境因素引起的。Nettie M. Stevens博士的《精子发生研究》(1905年)提供了明确的证据,证明Y染色体的遗传启动了黄粉虫的男性发育。这一结果证实了性别确实是具有遗传基础的孟德尔特征,并且性染色体起着至关重要的作用。在《精子发生研究》(1906年)的第二部分中,在数十个其他物种中发现了一对XY,进一步验证了性染色体的功能。自这项形成性工作以来,大量的动物和植物研究已经研究了性别的遗传基础。这篇综述的目的是再次揭示史蒂文斯在精子发生的研究以及这项工作的持久影响。此外,我们还关注上个世纪植物系统中的关键发现,以及通过跨多个系统进行综合来最好回答的开放性问题,就像史蒂文斯的工作一样。
This article is part of the theme issue ‘Sex determination and sex chromosome evolution in land plants’.
本文是主题问题“陆地植物的性别决定和性染色体进化”的一部分。
Keywords: sex chromosomes, dioecy, cytology, genomics
1. Introduction 1. 引言
For over a century, uncovering the genetic basis for the development of the separate sexes has been a lively area of research. How a single species develops two strikingly different forms captivated early naturalists, like Carl Linnaeus and Charles Darwin, but it was not until the early 1900s that sex was shown to have a genetic basis. The pivotal study that provided this evidence was Studies in spermatogenesis (1905) by Dr Nettie M. Stevens [1]. In this two-part piece, Stevens showed, through careful cytological examination, that the inheritance of the Y chromosome is correlated with male development in dozens of insect species. Despite the importance of this work, and over 6000 peer-reviewed articles on the topic of sex chromosomes since (Web of Science, accessed 20 August 2021), Studies in spermatogenesis has been cited fewer than 100 times (Google Scholar, accessed 24 July 2021). Here, we aim to reilluminate interest in this eloquent body of work and the decisive importance of Stevens’ research to the topic of sex chromosomes. We next discuss the outpouring of studies on sex chromosomes in diverse plant systems after the publication of Studies in spermatogenesis, and posit that the future of studying sex chromosomes should follow the lessons of past and current researchers by examining many independent evolutions across kingdoms.
一个多世纪以来,揭示两性分离发育的遗传基础一直是一个活跃的研究领域。一个物种如何发展出两种截然不同的形式吸引了早期的博物学家,如卡尔·林奈和查尔斯·达尔文,但直到 1900 年代初,性别才被证明具有遗传基础。提供这一证据的关键研究是Nettie M. Stevens博士的《精子发生研究》(1905)[1]。在这篇由两部分组成的文章中,史蒂文斯通过仔细的细胞学检查表明,Y染色体的遗传与数十种昆虫的雄性发育有关。尽管这项工作很重要,并且自那时以来有超过 6000 篇关于性染色体主题的同行评议文章(Web of Science,2021 年 8 月 20 日访问),但精子发生研究被引用的次数不到 100 次(Google Scholar,2021 年 7 月 24 日访问)。在这里,我们旨在重新阐明人们对这部雄辩的工作的兴趣,以及史蒂文斯的研究对性染色体主题的决定性重要性。接下来,我们讨论了在《精子发生研究》发表后,对不同植物系统中性染色体的研究的大量涌现,并认为研究性染色体的未来应该遵循过去和现在研究人员的经验教训,通过研究各个王国的许多独立进化。
2. Nettie Stevens’ career 2. 妮蒂·史蒂文斯的职业生涯
Nettie Maria Stevens was born on 7 July 1861 in Cavendish, Vermont, USA (figure 1). Stevens started her education at Westford Academy (1872–1880) and Westfield State Normal School (now Westfield State University; 1881–1883), to prepare for a career in teaching, and for the next decade or so Stevens worked as a teacher or librarian [2]. She saved enough money to continue her education, and in 1896 she began at Stanford University (then called Leland Stanford Jr University), earning both Bachelor and Master degrees (1896–1900). It was during this time at Stanford that Stevens’ cytological and histological research took off while spending her summers working at the Hopkins Marine Station. In 1901, she published her first manuscript histologically describing ciliates, where through her detailed observations across the life cycle, she identified two new species [3].
内蒂·玛丽亚·史蒂文斯于1861年7月7日出生于美国佛蒙特州卡文迪什(图1)。史蒂文斯在韦斯特福德学院(1872-1880)和韦斯特菲尔德州立师范学校(现为韦斯特菲尔德州立大学;1881-1883)开始接受教育,为教学事业做准备,在接下来的十年左右的时间里,史蒂文斯担任教师或图书管理员[2]。她攒下了足够的钱继续她的学业,并于 1896 年开始在斯坦福大学(当时称为利兰斯坦福大学)学习,获得了学士和硕士学位(1896-1900 年)。正是在斯坦福大学的这段时间里,史蒂文斯在霍普金斯海洋站度过暑假时,细胞学和组织学研究开始起飞。1901 年,她发表了第一份组织学描述纤毛虫的手稿,通过对生命周期的详细观察,她发现了两个新物种 [3]。

Figure 1.
Photographs of Dr Nettie M. Stevens. (a) Stevens looking through her iconic microscope (1909), Bryn Mawr College Special Collections, PA_Stevens_Nettie_005. (b) Alice Boring, Nettie Stevens and colleagues at a beach near Capo di Messina (1909), Bryn Mawr College Special Collections, PA_Stevens_Nettie_001.
Nettie M. Stevens博士的照片。(a) 史蒂文斯通过她的标志性显微镜观察(1909 年),布林莫尔学院特别收藏,PA_Stevens_Nettie_005。(b) Alice Boring、Nettie Stevens 及其同事在 Capo di Messina(1909 年)附近的海滩上,布林莫尔学院特别收藏,PA_Stevens_Nettie_001。
The turn of the twentieth century was a transformative time for cytogenetic studies. Gregor Mendel’s foundational research on heredity in pea plants, establishing the laws of segregation and independent assortment in reproductive cells [4], had recently been rediscovered by Carl Correns, Erich von Tschermak and Hugo de Vries [5–7]. Only a few years later Walter S. Sutton and Theodor Boveri independently showed that the behaviour of chromosomes during meiosis could be the basis for such Mendelian inheritance [8,9]. Though not all biologists were sold on the role of chromosomes in heredity based on these works alone, Stevens was quick to adopt these findings into her research.
二十世纪初是细胞遗传学研究的变革时期。格雷戈尔·孟德尔(Gregor Mendel)对豌豆植物遗传的基础研究,建立了生殖细胞中分离和独立分类的定律[4],最近被卡尔·科伦斯(Carl Correns),埃里希·冯·切尔马克(Erich von Tschermak)和雨果·德弗里斯(Hugo de Vries)[5\u20127]重新发现。仅仅几年后,Walter S. Sutton和Theodor Boveri独立地证明了减数分裂过程中染色体的行为可能是这种孟德尔遗传的基础[8,9]。尽管并非所有生物学家都仅凭这些工作就对染色体在遗传中的作用感到满意,但史蒂文斯很快就将这些发现纳入了她的研究。
Nettie Stevens continued her education at Bryn Mawr (Pennsylvania, USA), which by many accounts was an ideal place for biological research. Bryn Mawr was a relatively new school at this time, established in 1885 as one of the Seven Sister Schools, but had employed two well-known biologists in succession: Edmund Beecher Wilson, who would later author the acclaimed The cell in development and inheritance (1896) [10] and Thomas Hunt Morgan, future Nobel Laureate (1933) and ‘Father of Modern Genetics’. Though Wilson left for Columbia University before Stevens started, Morgan became Stevens’ doctoral advisor, and the three collaborated closely. Soon after starting, in 1901, Stevens received the Bryn Mawr President’s European Fellowship, which provided funding to research at Naples Zoological Station with Theodor Boveri, who at the time was working on his contributions to the chromosomal theory of inheritance. Stevens’ doctoral thesis built on her Master’s work, expanding to new species and varieties of ciliates, where she described microanatomy and regeneration [11]. In 1903, Nettie Stevens received her PhD.
妮蒂·史蒂文斯(Nettie Stevens)在布林莫尔(美国宾夕法尼亚州)继续她的学业,许多人认为这里是生物学研究的理想场所。布林莫尔大学在当时是一所相对较新的学校,成立于1885年,是七所姐妹学校之一,但先后聘请了两位著名的生物学家:埃德蒙·比彻·威尔逊(Edmund Beecher Wilson),他后来撰写了广受赞誉的《发育与继承中的细胞》(1896年)[10]和托马斯·亨特·摩根(Thomas Hunt Morgan),未来的诺贝尔奖获得者(1933年)和“现代遗传学之父”。虽然威尔逊在史蒂文斯开始之前就去了哥伦比亚大学,但摩根成为了史蒂文斯的博士生导师,三人密切合作。在开始后不久,在1901年,史蒂文斯获得了布林莫尔总统的欧洲奖学金,该奖学金为那不勒斯动物园与西奥多·博韦里(Theodor Boveri)的研究提供资金,当时西奥多·博韦里(Theodor Boveri)正在研究他对染色体遗传理论的贡献。史蒂文斯的博士论文建立在她的硕士工作的基础上,扩展到纤毛酸盐的新物种和变种,在那里她描述了显微解剖学和再生[11]。1903年,妮蒂·史蒂文斯(Nettie Stevens)获得了博士学位。
Over the next several years, Stevens continued her upward trajectory and notability as a scientist. In 1903, Stevens applied for and received a grant to specifically study sex determination by chromosomes [12], the research published in Studies in spermatogenesis. In 1904, she became a postdoctoral research assistant with the Carnegie Institute of Washington and then returned to Bryn Mawr as a research associate. Her research continued to focus on cytological analyses throughout spermatogenesis, development and regeneration. Interestingly, Stevens may have also been one of the first scientists to discover B chromosomes, suggesting a possible relationship between these and sex chromosomes [13,14]. In 1905, her manuscript focusing on the germ cells of aphids won the Ellen Richards Prize given by the Association for Maintaining the American Woman’s Table at the Zoological Station at Naples [15]. In 1910, Stevens was listed in the top 1000 ‘men of science’, being one of 18 women recognized that year [16]. By 1912, Stevens was finally offered a research professorship at Bryn Mawr, but before she began this new role, she died of breast cancer at the age of 50 (4 May 1912).
在接下来的几年里,史蒂文斯继续她作为科学家的上升轨迹和知名度。1903年,史蒂文斯申请并获得了一项资助,专门研究染色体的性别决定[12],该研究发表在《精子发生研究》上。1904 年,她成为华盛顿卡内基研究所的博士后研究助理,然后回到布林莫尔担任研究助理。她的研究继续集中在精子发生、发育和再生过程中的细胞学分析。有趣的是,史蒂文斯也可能是最早发现B染色体的科学家之一,这表明这些染色体与性染色体之间可能存在关系[13,14]。1905年,她以蚜虫生殖细胞为重点的手稿获得了那不勒斯动物站维护美国妇女餐桌协会颁发的艾伦·理查兹奖[15]。1910 年,史蒂文斯被列入前 1000 名“科学男士”之一,是当年公认的 18 位女性之一 [16]。到 1912 年,史蒂文斯终于获得了布林莫尔的研究教授职位,但在她开始这个新角色之前,她在 50 岁时死于乳腺癌(1912 年 5 月 4 日)。
Without a doubt, despite her life and career tragically being cut short, Stevens made an extraordinary impact on the field of biology. In the 11 years between Stevens’ first publication and her passing, she published at least 38 manuscripts [2]. Stevens’ contributions have not been completely lost to time. In 1994, Stevens was inducted into the National Women’s Hall of Fame and in 2017 Westfield State University opened the Dr Nettie Maria Stevens Science and Innovation Center. Stevens was a remarkably accomplished scientist with many foundational discoveries, though her best-known is about the role of sex chromosomes.
毫无疑问,尽管史蒂文斯的生活和职业生涯不幸地被缩短,但她对生物学领域产生了非凡的影响。从史蒂文斯首次出版到她去世的11年间,她至少出版了38篇手稿[2]。史蒂文斯的贡献并没有完全被时间遗忘。1994 年,史蒂文斯入选国家女性名人堂,2017 年,韦斯特菲尔德州立大学开设了内蒂·玛丽亚·史蒂文斯博士科学与创新中心。史蒂文斯是一位成就卓著的科学家,拥有许多基础性发现,尽管她最著名的是关于性染色体的作用。
(a) Studies in spermatogenesis (a) 精子研究
The development of the sexes was an area of substantial interest by the end of the ninteenth century. As Wilson described it, ‘The phenomenon of sex is so nearly a universal one that it may be assumed to make some appeal to the interest of biologists in every field of inquiry’ [17, p. 53]. Many researchers began investigating the leading theories behind sex determination, principally whether there is a genetic underpinning or if external environmental factors are involved. While today there are some species for which a form of environmental sex determination has been identified [18,19], most species with gonochory or dioecy have a genetic basis.
到十九世纪末,两性的发展已成为人们极为关注的领域。正如威尔逊所描述的那样,“性现象几乎是一种普遍现象,以至于可以认为它在某些研究领域都引起了生物学家的兴趣”[17,第53页]。许多研究人员开始调查性别决定背后的主要理论,主要是是否存在遗传基础,或者是否涉及外部环境因素。虽然目前已经确定了一些物种的环境性别确定形式[18,19],但大多数具有雌雄异株或雌雄异株的物种都有遗传基础。
The beeline that resulted in the identification of sex chromosomes started in 1891, when Hermann von Henking found in the firebug, Pyrrochoris apterus, that during meiosis half of the sperm inherited 11 chromosomes and the other half 12. Von Henking called this twelfth chromosome the ‘X-element’ [20]. Less than a decade later, in 1899, McClung proposed the term ‘accessory chromosome’ for this element [21] and in 1902 he presented a theoretical framework for the involvement of this sperm accessory chromosome in the sex of an organism,
导致性染色体鉴定的直线始于 1891 年,当时赫尔曼·冯·亨金 (Hermann von Henking) 在萤火虫 Pyrrochoris apterus 中发现,在减数分裂过程中,一半的精子继承了 11 条染色体,另一半继承了 12 条染色体。冯·亨金(Von Henking)称第十二条染色体为“X元素”[20]。不到十年后的1899年,McClung为这一元素提出了“辅助染色体”一词[21],并在1902年提出了这种精子辅助染色体参与生物体性别的理论框架,
A most significant fact, and one upon which almost all investigators are united in opinion, is that the element is apportioned to but one half of the spermatozoa. Assuming it to be true that the chromatin is the important part of the cell in the matter of heredity, then it follows that we have two kinds of spermatozoa that differ from each other in a vital matter. We expect, therefore, to find in the offspring two sorts of individuals in approximately equal numbers, under normal conditions, that exhibit marked differences in structure. A careful consideration will suggest that nothing but sexual characters thus divides the members of a species into two well-defined groups, and we are logically forced to the conclusion that the peculiar chromosome has some bearing upon this arrangement. [22]
一个最重要的事实,也是几乎所有研究人员都一致认为的事实,是该元素只分配给精子的一半。假设染色质确实是细胞在遗传方面的重要部分,那么就可以看出,我们有两种精子,它们在重要问题上彼此不同。因此,我们期望在正常条件下,在后代中发现两种数量大致相等的个体,它们在结构上表现出明显的差异。仔细考虑会发现,只有性特征才能将一个物种的成员分为两个定义明确的群体,我们从逻辑上被迫得出结论,即特殊的染色体与这种安排有一定关系。[22]
Regarding the accessory chromosome, McClung (1902) also writes that ‘Its careful and uniform division during the mitoses of all the spermatogonia suggests anything but an unimportant structure’ [22]. Studies focusing on identifying accessory chromosomes in diverse systems swelled. Louise Wallace identified a double accessory chromosome system in the spider Agalena naevia [23]. Frederick Paulmier considered the accessory chromosome to be degrading and disappearing from a species because he observed that it fails to divide and is not equally represented in the final spermatocyte mitosis [24]. Likewise, Thomas Montgomery thought the accessory chromosomes ‘… are in the process of disappearance, in the evolution of a higher to a lower chromosomal number’ [25]. Discussion on whether the accessory chromosomes were involved in sex determination continued, but the direct evidence for its role had yet to be shown.
关于副染色体,McClung(1902)还写道,“它在所有精原细胞的有丝分裂过程中仔细而均匀的分裂表明,它不是一个不重要的结构”[22]。专注于识别不同系统中的辅助染色体的研究激增。路易丝·华莱士(Louise Wallace)在蜘蛛Agalena naevia中发现了双副染色体系统[23]。弗雷德里克·保尔米尔(Frederick Paulmier)认为副染色体正在降解并从物种中消失,因为他观察到它无法分裂,并且在最终的精母细胞有丝分裂中没有得到平等的代表[24]。同样,托马斯·蒙哥马利(Thomas Montgomery)认为副染色体“…正在消失的过程中,在从较高的染色体数数进化到较低的染色体数的进化中“[25]。关于副染色体是否参与性别决定的讨论仍在继续,但尚未显示其作用的直接证据。
Studies in spermatogenesis was published as a two-part book, with the first part released in 1905 (figure 2) [1]. Importantly, Stevens tracked the behaviour of the accessory chromosome across different orders of Coleoptera and deduced its inheritance pattern through cell division. Part I of Studies in spermatogenesis included chromosome squashes from termites (Termopsis angusticollis), sand crickets (Stenopelmatus spp.) and croton-bugs (Blattella germanica), and indeed, in Stenopelmatus and B. germanica, Stevens found evidence of the accessory chromosomes. But it is mealworms (Tenebrio molitor) that Stevens described as the most interesting group studied in her 1905 publication, for what she found differed from that of the accessory chromosomes described by McClung. She writes that ‘In both somatic and germ cells of the two sexes there is a difference not in the number of chromatin elements, but in the size of one, which is very small in the male and of the same size as the other 19 in the female’ [1]. Stevens reasons that,
《精子发生研究》以两部分的形式出版,第一部分于1905年出版(图2)[1]。重要的是,史蒂文斯追踪了鞘翅目不同目次的副染色体的行为,并通过细胞分裂推断出其遗传模式。精子发生研究的第一部分包括来自白蚁(Termopsis angusticollis)、沙蟋蟀(Stenopelmatus spp.)和巴豆虫(Blattella germanica)的染色体南瓜,事实上,在Stenopelmatus和B. germanica中,史蒂文斯发现了辅助染色体的证据。但是,史蒂文斯在她1905年的出版物中描述为最有趣的一组研究的是黄粉虫(Tenebrio molitor),因为她发现与McClung描述的辅助染色体不同。她写道:“在两性的体细胞和生殖细胞中,染色质元素的数量不同,而是一个元素的大小,雄性非常小,而雌性则与其他19个相同”[1]。史蒂文斯认为,
Since the somatic cells of the female contain 20 large chromosomes while those of the male contain 19 large ones and 1 small one, this seems to be a clear case of sex-determination, not by an accessory chromosome, but by a definite difference in the character of the elements of one pair of chromosomes of the spermatocytes of the first order, the spermatozoa which contain the small chromosome determining the male sex, while those that contain 10 chromosomes of equal size determine the female sex. This result suggests that there may be in many cases some intrinsic difference affecting sex, in the character of the chromatin of one-half of the spermatozoa, though it may not usually be indicated by such an external difference in form or size of the chromosomes as in Tenebrio. [1]
由于女性的体细胞包含 20 条大染色体,而男性的体细胞包含 19 条大染色体和 1 条小染色体,这似乎是性别决定的一个明显案例,不是通过辅助染色体,而是通过一阶精母细胞的一对染色体元素的特征存在明确差异, 包含决定男性性别的小染色体的精子,而包含 10 条相同大小的染色体的精子决定了女性性别。这一结果表明,在许多情况下,可能存在一些影响性别的内在差异,即一半精子的染色质特性,尽管通常可能不会像Tenebrio那样通过染色体的形式或大小的外部差异来表示。[1]
Figure 2.
Key events for visualizing sex chromosome research in plants over time. Purple circles indicate empirical findings and yellow squares technological advances that have set the foundation for discovery in sex chromosome research. The timeline begins with Stevens’ discovery of sex chromosomes, followed by the wave of cytological research that followed her, including the first descriptions of a heteromorphic sex chromosome pair in a liverwort (1917) and in angiosperms (1923). With the development of PCR and modern sequencing techniques, the identification of sex chromosomes diverged from traditional cytological techniques and moved toward marker-based as well as whole-genome approaches. This has led to a new renaissance of sex chromosome research not unlike the one Stevens began in 1905. An expanded timeline can be found in electronic supplementary material, table S1. NGS, next-generation sequencing.
随时间推移可视化植物性染色体研究的关键事件。紫色圆圈表示实证发现,黄色方块表示技术进步,为性染色体研究的发现奠定了基础。时间线始于史蒂文斯发现性染色体,随后是随之而来的细胞学研究浪潮,包括首次描述苔类(1917年)和被子植物中的异形性染色体对。随着PCR和现代测序技术的发展,性染色体的鉴定从传统的细胞学技术中分化出来,转向基于标记和全基因组的方法。这导致了性染色体研究的新复兴,这与史蒂文斯在1905年开始的研究不同。扩大的时间表见电子补充材料表S1。NGS,下一代测序。
One of the virtues of Nettie Stevens’ work is the diversity of species where she observed the segregation of different sex chromosome systems. Stevens published part II of Studies in spermatogenesis in June of 1906, where she studied the spermatogenesis of 23 more species in Coleoptera, and in August 1906 a footnote was added containing results for 19 more [26]. In this second part, Stevens found that 86% of the species studied are characterized by having heterochromosomes and the remaining had accessory chromosomes in male germ cells [26]. On the accessory chromosomes (referred to here as ‘odd chromosome’) Stevens writes,
妮蒂·史蒂文斯(Nettie Stevens)工作的优点之一是物种的多样性,她观察到不同性别染色体系统的分离。史蒂文斯于1906年6月出版了《精子发生研究》的第二部分,研究了鞘翅目中另外23个物种的精子发生,并于1906年8月添加了一个脚注,其中包含另外19个物种的结果[26]。在第二部分中,史蒂文斯发现,所研究的物种中有86%具有异染色体,其余的具有雄性生殖细胞中的辅助染色体[26]。关于副染色体(这里称为“奇数染色体”),史蒂文斯写道:
The odd chromosome, so far as it has been studied, behaves precisely like the larger member of the unequal pair without its smaller mate. In the growth stage it remains condensed and either spherical or sometimes flattened against the nuclear membrane. In the first maturation mitosis it is attached to one pole of the spindle, does not divide, but goes to one of the two second spermatocytes. In the second spermatocyte it divides with the other chromosomes, giving two equal classes of spermatids differing by the presence or absence of this odd chromosome. [26]
就所研究的而言,奇数染色体的行为与不等对中较大的成员完全相同,而没有较小的配偶。在生长阶段,它保持凝结,呈球形或有时在核膜上扁平。在第一次成熟有丝分裂中,它附着在纺锤体的一极上,不分裂,而是进入两个第二精母细胞之一。在第二个精母细胞中,它与其他染色体分裂,产生两类相等的精子细胞,这些精子细胞的不同之处在于这条奇数染色体的存在与否。[26]
In this section, Nettie Stevens uses the term ‘mitosis’ to describe what is now known as meiosis I and II in the spermatocytogenesis, where primary spermatocytes (2n) divide into secondary spermatocytes in meiosis I and spermatids in meiosis II. Interestingly, the term ‘meiosis’ (from the Greek μείωσις, ‘lessening’) was not coined until 1905 by cytologists John Farmer and John Moore, explaining the absence of this term in Stevens’ analysis [27]. She demonstrates that these divisions lead to the ‘odd chromosome,’ labelled as ‘x’ on her plates, segregating according to Mendelian principles in meiosis I and II. Likewise, this Mendelian behaviour was found for the pair of heterochromosomes that she labels as ‘l’ (for large) and ‘s’ (for small), which later became known as ‘X’ and ‘Y’ chromosomes.
在本节中,Nettie Stevens 使用术语“有丝分裂”来描述现在称为精子细胞生成中的减数分裂 I 和 II,其中初级精母细胞 (2n) 在减数分裂 I 中分裂为次级精母细胞和减数分裂 II 中的精子细胞。有趣的是,“减数分裂”一词(来自希腊语μείωσις,“减数分裂”)直到1905年才由细胞学家约翰·法默(John Farmer)和约翰·摩尔(John Moore)创造,这解释了史蒂文斯分析中没有这个术语[27]。她证明,这些分裂导致了“奇怪的染色体”,在她的板上标记为“x”,根据减数分裂I和II中的孟德尔原理进行分离。同样,这种孟德尔行为也被发现在一对异染色体上,她将其标记为“l”(代表大)和“s”(代表小),后来被称为“X”和“Y”染色体。
The implications of her observations and deductions are elegant and profound: these odd chromosomes (x) or heterochromosomes (l or s) follow Mendel’s laws of inheritance, and the presence (or absence) of these chromosomes corresponds to sex determination. The logic behind this is eloquently noted in her discussion, when she states that ‘It is therefore evident that an egg fertilized by a spermatozoon (1) containing the small member of an unequal pair or (2) lacking one chromosome, must develop into a male, while an egg fertilized by a spermatozoon containing the larger element of an unequal pair of heterochromosomes or the odd chromosomes must produce a female’ [26]. In 1905, the same year as part I of Studies in spermatogenesis was published, E. B. Wilson also published a study on the sex chromosomes in Hemiptera [28]. In his piece, Wilson also showed that males possess an unequal pair of chromosomes, the smaller of which he called the ‘idiochromosomes’. Wilson added a footnote to his 1905 piece acknowledging Stevens’ findings.
她的观察和推论的意义是优雅而深刻的:这些奇数染色体(x)或异染色体(l或s)遵循孟德尔的遗传定律,这些染色体的存在(或不存在)对应于性别决定。在她的讨论中,她雄辩地指出了这背后的逻辑,当时她指出,“因此,很明显,由包含一对不等子的小成员或(2)缺少一条染色体的精子受精的卵子必须发育成雄性,而由包含一对不等异染色体或奇数染色体的较大元素的精子受精的卵子必须产生雌性”[26]。1905年,在《精子发生研究》第一部分发表的同一年,E.B.威尔逊也发表了一篇关于半翅目性染色体的研究[28]。在他的文章中,威尔逊还表明,男性拥有一对不等的染色体,其中较小的染色体他称之为“同色染色体”。威尔逊在他1905年的文章中添加了一个脚注,承认了史蒂文斯的发现。
The discovery, referred to in a preceding footnote, that the spermatogonial number of Anasa is 21 instead of 22, again goes far to set aside the difficulties [of McClung’s hypothesis] here urged. Since this paper was sent to press I have also learned that Dr N. M. Stevens (by whose kind permission I am able to refer to her results) has independently discovered in a beetle, Tenebrio, a pair of unequal chromosomes that are somewhat similar to the idiochromosomes in Hemiptera and undergo a corresponding distribution to the spermatozoa. She was able to determine, further, the significant fact that the small chromosome is present in the somatic cells of the male only, while in those of the female it is represented by a larger chromosome. These very interesting discoveries, now in course of publication, afford, I think, a strong support to the suggestion made above; and when considered in connection with the comparison I have drawn between the idiochromosomes and the accessory show that McClung’s hypothesis may, in the end, prove to be well founded. [28, p. 403]
在前面的脚注中提到的这一发现,即Anasa的精原数量是21而不是22,这再次大大消除了这里所强调的[McClung假设]的困难。自从这篇论文发表以来,我还了解到N.M.史蒂文斯博士(经她允许,我能够参考她的研究结果)在一种甲虫Tenebrio中独立发现了一对不等的染色体,这些染色体与半翅目中的同染色体有些相似,并经历了与精子的相应分布。她能够进一步确定一个重要事实,即小染色体仅存在于男性的体细胞中,而在女性的体细胞中,它由较大的染色体表示。我认为,这些非常有趣的发现现在正在出版中,为上述建议提供了强有力的支持;当结合我在同染色体和附属染色体之间的比较来考虑时,表明麦克朗的假设最终可能被证明是有根据的。[28,第403页]
While Wilson’s research was published a few months before Stevens’, some give Stevens the credit for the discovery of sex chromosomes because her conclusions were firmer [12]. Regardless of whether the discovery for the role of sex chromosomes should be shared between Stevens and Wilson, as the two independently arrived at these results in 1905, Stevens was certainly the first to concretely show that the Y chromosome was involved in sex determination. Her work provided a molecular and cytological framework for supporting the earlier hypothesis put forth by Carl Correns after crossing the dioecious vine Bryonia that sex was, indeed, a Mendelian trait [29]. McClung had incorrectly asserted that the accessory chromosome was a male determiner [22]. Wilson maintained environmental roles [29]. In Stevens’ own words,
虽然威尔逊的研究比史蒂文斯的研究早几个月发表,但有些人认为史蒂文斯发现了性染色体,因为她的结论更为坚定[12]。无论性染色体作用的发现是否应该在史蒂文斯和威尔逊之间共享,因为两人在1905年独立得出了这些结果,史蒂文斯无疑是第一个具体证明Y染色体参与性别决定的人。她的工作为支持卡尔·科伦斯(Carl Correns)在杂交雌雄异株葡萄藤(Bryonia)后提出的早期假设提供了一个分子和细胞学框架,即性别确实是孟德尔的特征[29]。McClung错误地断言副染色体是男性决定因素[22]。威尔逊保持了环境角色[29]。用史蒂文斯自己的话来说,
Wilson suggests as alternatives to the chromosome sex according to Mendel’s Law, (1) that the heterochromosomes may merely transmit sex characters, sex being determined by protoplasmic conditions external to the chromosomes; (2) That the heterochromosomes may be sex-determining factors only by virtue of difference in activity or amount of chromatin, the female sex chromosome in the male being less active. [26]
威尔逊认为,根据孟德尔定律,作为染色体性别的替代方案,(1)异染色体可能仅传递性别特征,性别由染色体外部的原生质条件决定;(2)异染色体可能仅通过染色质活性或数量的差异而成为性别决定因素,而男性中的女性性染色体活性较低。[26]
Over the next several years, more studies in spermatogenesis were undertaken by Stevens and her colleagues. Stevens was the first to identify the heterochromosomes of Drosophila melanogaster (then called Drosophila ampelophila) and other flies [30,31]. Even more heteromorphic pairs were found in earwigs (Forficula auricularia) [32] and guinea pigs [33]. Stevens’ rigorousness and tenacity to uncover the role of sex chromosomes were apparent, and her depth of knowledge of the field unmatched. Upon learning about lagging chromosomes, Stevens carefully re-examined aphids, revealing the lagging member was in fact a heterochromosome, revoking her previous findings that these species lacked evidence for any [34]. But, not in all species could heterochromosomes be identified; such was the case in mosquitoes [35]. At Bryn Mawr, Stevens advised doctoral student Alice M. Boring (figure 1), who notes in her dissertation that while at Woods Hole in 1905, Stevens suggested Boring study the spermatogenesis of many more species of insects [36]. Indeed, Boring’s PhD research focused on the spermatogenesis of 22 species, finding that all had the ‘odd chromosomes’ [36]. Later Boring would study chicken spermatogenesis, where a clear pair of neither heterochromosomes nor accessory chromosomes could be identified [37]. Over a decade after Stevens’ death, Boring found Stevens’ notes on her independent examinations of chicken [38]. As it turns out, the lack of heterochromosomes found in chicken spermatogenesis is because they have a ZW system, which was shown by Michael F. Guyer’s studies in oogenesis in 1916 [39].
在接下来的几年里,史蒂文斯和她的同事们对精子发生进行了更多的研究。Stevens是第一个鉴定出黑腹果蝇(Drosophila melanogaster,当时称为果蝇(Drosophila ampelophila)和其他果蝇异染色体的人[30,31]。在耳蕊(Forficula auricularia)[32]和豚鼠[33]中发现了更多的异形对。史蒂文斯在揭示性染色体作用方面的严谨和坚韧是显而易见的,她对该领域的知识深度无与伦比。在了解了滞后的染色体后,史蒂文斯仔细地重新检查了蚜虫,发现滞后的成员实际上是异染色体,推翻了她之前的发现,即这些物种缺乏任何证据[34]。但是,并非所有物种都能鉴定出异染色体;蚊子就是这种情况[35]。在Bryn Mawr,Stevens为博士生Alice M. Boring(图1)提供建议,她在论文中指出,1905年在伍兹霍尔(Woods Hole)时,史蒂文斯建议Boring研究更多昆虫种类的精子发生[36]。事实上,Boring的博士研究主要集中在22个物种的精子发生上,发现所有物种都有“奇怪的染色体”[36]。后来,Boring研究了鸡的精子发生,在那里可以鉴定出一对既不是异染色体也不是副染色体的清晰染色体[37]。史蒂文斯去世十多年后,博林发现了史蒂文斯关于鸡肉独立检查的笔记[38]。事实证明,在鸡精子发生中发现的异染色体的缺乏是因为它们具有ZW系统,Michael F. Guyer在1916年的卵子发生研究中表明了这一点[39]。
Across these foundational research pieces and more, many different terms were used to describe what we now refer to as sex chromosomes, an issue raised by researchers of the time. ‘Since the discovery of peculiarly modified chromosomes in certain of the insects a great variety of names has been proposed for them, and most of these suffer from a quite unnecessary length. My own earlier terms “heterochromosome” and “chromatin nucleolus” were cumbersome, and “accessory chromosome” and “heterotropic chromosome” sin equally in this regard, while “special chromosome” and “idiochromosome” are no way self-explanatory.’ [40]. In 1906 Wilson first used the term ‘sex chromosome’ [41] and by 1909 used ‘X’ and ‘Y’ to delineate between the heteromorphic pair [17]. Confusion about the term ‘sex chromosome’ and what it represents continues today (for a discussion on definitions, see [42]). By casting such a wide net of species diversity, scientists from each of these independent and complementary studies had stumbled upon the foundation of the modern diversity of sex chromosome systems, including XX/XO (dosage) systems where chromosome number changes between males and females, XX/XY systems where the heterogametic sex chromosome pair is found during spermatogenesis in males, and ZZ/ZW systems where the heterogametic pair is found in females during oogenesis.
在这些基础研究以及更多研究中,使用了许多不同的术语来描述我们现在所说的性染色体,这是当时研究人员提出的一个问题。“自从在某些昆虫中发现特殊修饰的染色体以来,人们已经为它们提出了各种各样的名称,其中大多数都具有相当不必要的长度。我自己早期的术语“异染色体”和“染色质核仁”很麻烦,而“副染色体”和“异向性染色体”在这方面同样有罪,而“特殊染色体”和“同色染色体”则不是不言自明的。1906年,威尔逊首次使用“性染色体”[41],到1909年,他使用“X”和“Y”来区分异形对[17]。关于“性染色体”一词及其代表的含义的混淆至今仍在继续(关于定义的讨论,参见[42])。通过撒下如此广泛的物种多样性网络,来自这些独立和互补研究中的每一项的科学家们都偶然发现了性染色体系统现代多样性的基础,包括男性和女性之间染色体数量变化的 XX/XO(剂量)系统,XX/XY 系统,其中在男性精子发生过程中发现异配子性染色体对, 和 ZZ/ZW 系统,其中异配子对在卵子发生过程中在雌性中发现。
3. The prismatic sex chromosomes of plants 3. 植物的棱柱形性染色体
Undoubtedly, Nettie Stevens’ research transformed animal genetics. The ground-breaking impact it had on plant genetics, however, is equally significant even if less obvious. Shortly after Studies in spermatogenesis was published, studies focusing on potential sex chromosome systems in plants burgeoned. Unlike animals, separate sexes, or dioecy, is rarer in angiosperms, occurring in approximately 6% of species with 34 clades—such as Diospyros (700 species) and Pandanus (600 species)—and sometimes entire families (e.g. Myristicaceae), accounting for roughly 43% of the dioecious angiosperms [43]. Even with its uncommonness, dioecy has evolved hundreds of independent times in angiosperms, while in other plant lineages, such as those of the bryophytes and gymnosperms, it appears to be more common [44] (figure 3). Despite being seemingly disadvantageous for a sessile organism, dioecy is still the dominant reproductive strategy for critical fruit (fig), nut (pistachio), vegetable (asparagus), ornamental (gingko) and special products crops (hops, hemp) among many other species valuable to forestry, conservation, and bioremediation efforts [47]. While botanists, farmers and horticulturalists had always been acutely aware of this trait, Stevens’ work was the first to provide a foundation for exploring its genetic basis in plants.
毫无疑问,妮蒂·史蒂文斯的研究改变了动物遗传学。然而,它对植物遗传学的突破性影响同样重要,即使不那么明显。在《精子发生研究》发表后不久,关注植物中潜在性染色体系统的研究如雨后春笋般涌现。与动物不同,被子植物中分离性别或雌雄异株的情况更为罕见,约6%的物种有34个分支,如Diospyros(700种)和Pandanus(600种),有时还有整个科(如肉豆蔻科),约占被子植物的43%[43]。尽管雌雄异株并不常见,但在被子植物中,雌雄异株已经独立进化了数百次,而在其他植物谱系中,如苔藓植物和裸子植物,它似乎更常见[44](图3)。尽管雌雄异株对无柄生物似乎不利,但雌雄异株仍然是关键水果(无花果)、坚果(开心果)、蔬菜(芦笋)、观赏作物(银杏)和特殊产品作物(啤酒花、大麻)以及许多其他对林业、保护和生物修复工作有价值的物种的主要繁殖策略[47]。虽然植物学家、农民和园艺家一直敏锐地意识到这一特性,但史蒂文斯的工作是第一个为探索植物遗传基础提供基础的工作。

Figure 3.
Dioecious angiosperm orders studied to date. The heatmap shows the number of species in log scale and is mapped onto the topology from Angiosperm Phylogeny Group IV [45] using ggtree v. 3.0.4 [46]. (a) dioecious species within each order [44], (b) species with heteromorphic sex chromosomes identified through cytological approaches, © dioecious species with at least one genome reference in the NCBI Assembly database (accessed 30 August 2021).
迄今为止研究的雌雄异株被子植物目。热图显示了对数尺度的物种数量,并使用ggtree v. 3.0.4 [46]映射到被子植物系统发育组IV [45]的拓扑上。(a)每个目内的雌雄异株物种[44],(b)通过细胞学方法鉴定出具有异形性染色体的物种,(c)在NCBI Assembly数据库中至少有一个基因组参考的雌雄异株物种(2021年8月30日访问)。
A rush of cytological studies emerged as botanists re-examined the karyotypes of dioecious species (electronic supplementary material, table S1). Some of the earliest records of this frenzy come from 1909, when Eduard Strasburger and Mary G. Sykes observed the absence of heteromorphic sex chromosomes in Mercurialis annua, Bryonia dioica and Spinacia oleracea [48–50]. It took until 1917, more than decade after Stevens’ discovery of sex chromosomes, for Charles E. Allen to confirm the presence of heteromorphic sex chromosomes in the liverwort Sphaerocarpos [51]. A slew of cytological studies followed (figure 3), suggesting the presence of heteromorphic sex chromosomes in 68 plants and their absence in 46 plants by 1940 [52]. In 1958, Mogens Westergaard proposed a standard to temper the continuous outpouring of scantly supported claims of heteromorphic sex chromosome pairs. He argued that observations of such pairs are only valid if the heteromorphic pair is observed in the meiotic cycle of the heterogametic sex and not the homogametic sex, and if the sex chromosomes are also observable in the somatic cells of both sexes [53], as was done in Stevens’ Studies in spermatogenesis [1]. Since this time, only 19 species have been confirmed to have heteromorphic sex chromosomes, such as Cannabis, Humulus, Silene, Trichosanthes and Rumex. Species are being added and removed from this list as the meaning of ‘heteromorphic’ continues to evolve in genomic literature [42]. In the decades following Westergaard’s review, the diversity of species studied on sex chromosomes in plants decreased as Silene, Spinacia and Asparagus spp. emerged as model systems.
随着植物学家重新检查雌雄异株物种的核型,出现了一大批细胞学研究(电子补充材料,表S1)。这种狂热的一些最早记录来自1909年,当时Eduard Strasburger和Mary G. Sykes观察到Mercurialis annua、Bryonia dioica和Spinacia oleracea中不存在异形性染色体[48\u201250]。直到1917年,也就是史蒂文斯发现性染色体十多年后,查尔斯·艾伦(Charles E. Allen)才证实在苔类Sphaerocarpos中存在异形性染色体[51]。随后进行了一系列细胞学研究(图 3),表明到1940年,68种植物存在异形性染色体,而46种植物中不存在异形性染色体[52]。1958年,莫根斯·韦斯特加德(Mogens Westergaard)提出了一个标准,以缓和对异形性染色体对的缺乏支持的主张的持续涌现。他认为,只有当异质对是在异配子性别的减数分裂周期中观察到的,而不是在同配子性别中观察到的,并且如果性染色体在两性的体细胞中也可以观察到[53],就像史蒂文斯的精子发生研究[1]一样,对这种对的观察才有效。自那时以来,只有 19 个物种被证实具有异形性染色体,例如大麻、葎草、Silene、Trichosanthes 和 Rumex。随着基因组文献中“异形性”的含义不断演变,物种正在从此列表中添加和删除[42]。在Westergaard的审查之后的几十年里,随着Silene,Spinacia和Asparagus spp.作为模型系统的出现,在植物性染色体上研究的物种多样性有所下降。
Sex chromosome research has always been limited by the capabilities of microscopic or genomic technologies. The absence of heteromorphic sex chromosomes in many dioecious species presented a distinctive challenge that would not be taken on until the advent of modern sequencing techniques in the early 2000s (figure 2). Yet, the cytologists of Stevens’ day did not lack an abundance of surprising and often bewildering observations of heteromorphic sex chromosomes. As with many animals, plants also exhibit a wide variety of karyotypes that do not follow the standard XY or ZW systems. Hitoshi Kihara and Tomowo Ono first described the XX/XY1Y2 system in Rumex acetosa in 1925 [54]. Soon after, Öjvind Winge elaborated on the polytypic qualities of Humulus species, which are well-known for their markedly variable cytotypes [55]. Unusual systems continued to intrigue researchers into the late 1900s, such as the sex-associated floating translocation complexes in Viscum, where four, six and sometimes eight chromosomes form multivalent rings at meiosis [56]. The UV systems are similarly variable, with many homomorphic and heteromorphic pairs found by C. E. Allen [57], as well as multiple systems like the U1U2/V found in Frullania dilatata [58,59]. Methods for the visualization of sex chromosomes have dramatically improved since the days of Stevens’ Carl Zeiss Jena 8261 compound monocular microscope (figure 1; Bryn Mawr College Special Collections). Today, modern technologies, such as PacBio HiFi sequencing, have opened the door to more robust assembly of repeat-rich sex chromosomes and made research on homomorphic sex chromosomes, sex-determining regions (SDRs) and pseudo-autosomal regions more accessible than ever before [60].
性染色体研究一直受到微观或基因组技术能力的限制。在许多雌雄异株物种中缺乏异形性染色体是一个独特的挑战,直到2000年代初现代测序技术的出现才开始应对这一挑战(图2)。然而,史蒂文斯时代的细胞学家并不缺乏对异形性染色体的大量令人惊讶且常常令人困惑的观察结果。与许多动物一样,植物也表现出各种各样的核型,这些核型不遵循标准的 XY 或 ZW 系统。Hitoshi Kihara和Tomowo Ono于1925年首次在Rumex acetosa中描述了XX/XY1Y2系统[54]。不久之后,Öjvind Winge详细阐述了葎草物种的多型性,这些葎草以其明显可变的细胞型而闻名[55]。直到 1900 年代后期,不寻常的系统继续吸引着研究人员,例如 Viscum 中与性别相关的漂浮易位复合物,其中四条、六条甚至有时八条染色体在减数分裂时形成多价环 [56]。UV系统具有类似的可变性,C. E. Allen[57]发现了许多同态和异态对,以及在Frullania dillatata[58\u2229]中发现的多个系统。自Stevens的Carl Zeiss Jena 8261复合单目显微镜时代以来,性染色体的可视化方法已经有了显着改进(图1;布林莫尔学院特别收藏)。如今,PacBio HiFi测序等现代技术为更稳健地组装重复丰富的性染色体打开了大门,并使得对同态性染色体、性别决定区(SDR)和假常染色体区域的研究比以往任何时候都更容易获得[60]。
In the last several decades, genomic approaches have shed light on many previously unexamined or unidentified plant sex chromosome pairs (figures (figures22 and and3).3). The first plant genome reference for the hermaphroditic species Arabidopsis thaliana was published in 2000 [61], and quickly several sex chromosome assemblies followed, including for papaya and the common liverwort [62,63], with whole-genome references eventually to follow [64,65]. Today over 100 dioecious angiosperm genome references, at various levels of contiguity, are available on NCBI (figure 3). Yet, only a fraction of these references have been used to examine the sex chromosomes.
在过去的几十年里,基因组学方法揭示了许多以前未经检查或未确定的植物性染色体对(图 (图2 2 和 和 3)。 3).2000年发表了雌雄同体物种拟南芥的第一个植物基因组参考文献[61],随后很快出现了几种性染色体组装,包括木瓜和普通苔类[62,63],最终也出现了全基因组参考文献[64,65]。如今,NCBI上有超过100个雌雄异株被子植物基因组参考,它们处于不同的连续性水平(图3)。然而,这些参考资料中只有一小部分被用于检查性染色体。
Genomic analyses of plant sex chromosomes have addressed many theories developed for this kingdom. Because of the thousands of independent origins of plant sex chromosomes and few heteromorphic pairs identified cytologically (figure 3), the age of most evolutions was thought to be recent. The expectation is that heteromorphic pairs have had sufficient time for degeneration, or gene loss, to have occurred on the sex-specific chromosome to suggest older origins [66]. Such is the case in the older, cytologically heteromorphic pairs of Humulus lupulus [67], Phoenix dactylifera [68] and Silene latifolia [69]. In some cases, given enough time, the Y (or W) can be completely lost, transitioning to the XO (or ZO) system seen in studies of the ‘accessory chromosomes’ [42]. Though, curiously, to our knowledge, no plant species has been reliably identified as having a dosage (e.g. XO) system. Instead, homomorphic sex chromosomes are expected to have more recent origins, with little to no gene loss on the Y (or W). Consistent with this, many species studied have recent origins of sex chromosomes, within the last 5 million years, such as in Asparagus officinalis [70], M. annua [71] and S. oleracea [72], and so few (critical) genes have been lost from the Y that individuals with a YY karyotype remain viable [73–76]. However, some plant sex chromosomes defy these expectations. The moss UV sex chromosomes evolved hundreds of millions of years ago but are homomorphic in Ceratodon purpureus [77] and in Cannabis sativa the sex chromosomes share an origin with H. lupulus [67]; however, they are instead homomorphic.
植物性染色体的基因组分析已经解决了为这个王国开发的许多理论。由于植物性染色体有数千个独立起源,并且细胞学上鉴定出的异态对很少(图 3),因此大多数进化的年龄被认为是最近的。人们的期望是,异形对有足够的时间在性别特异性染色体上发生退化或基因丢失,从而提示更古老的起源[66]。在较老的细胞学异态对中,Humulus lupulus [67]、Phoenix dactylifera [68]和Silene latifolia [69]就是这种情况。在某些情况下,如果有足够的时间,Y(或W)可能会完全丢失,过渡到XO(或ZO)系统,这在“辅助染色体”的研究中可见[42]。然而,奇怪的是,据我们所知,还没有一种植物物种被可靠地确定为具有剂量(例如XO)系统。取而代之的是,同态性染色体预计具有更近的起源,Y(或W)上几乎没有基因丢失。与此相符的是,许多研究的物种在过去500万年内都有最近的性染色体起源,例如芦笋[70]、M. annua[71]和S. oleracea[72],并且从Y中丢失的(关键)基因如此之少,以至于具有YY核型的个体仍然存活[73\u201276]。然而,一些植物性染色体违背了这些期望。苔藓紫外线性染色体在数亿年前进化而来,但在Ceratodon purpureus[77]和Cannabis sativa中,性染色体与H. lupulus[67]有同形性;然而,它们是同态的。
The size of the non-recombining region also does not correlate with age in species studied to date [43], which may relate to haploid gene expression. Unlike animals, the plant life cycle consists of two separate generations, one haploid and the other diploid (i.e. alternation of generations), which has consequences for sex-specific development and sex chromosome evolution [76]. Because plants express genes in pollen or other haploid gametophyte stages, the non-recombining region of the sex chromosomes is expected to degenerate slower than is seen in animals [78,79]. Indeed, Mank suggests haploid expression in plants may represent the biggest difference known for sex chromosomes between these two kingdoms [80]. The S. latifolia sex chromosomes evolved over 10 million years, and while some genes have been lost on the SDR, the rate of loss is 60% lower than that of animals with a similar time since suppressed recombination occurred [81,82]. Estimates of divergence between XY genes in Rumex hastatulus suggest a minimum age of 9 million years [83], and while some genes have also been lost on the Y [84], pollen-expressed genes are significantly less likely to be lost than those expressed in diploid tissues [83,85]. The haploid C. purpureus UV sex chromosomes contain over 3400 genes each, half of which were shown to be expressed in the gametophytes [77]. In addition to haploid gene expression, the lack of degeneration could be due to the small size of the SDR seen in many plants, as degeneration is predicted to be faster when many genes are under selection [86,87].
在迄今为止研究的物种中,非重组区域的大小也与年龄无关[43],这可能与单倍体基因表达有关。与动物不同,植物的生命周期由两个独立的世代组成,一个是单倍体,另一个是二倍体(即世代交替),这对性别特异性发育和性染色体进化有影响[76]。由于植物在花粉或其他单倍体配子体阶段表达基因,因此性染色体的非重组区域预计退化速度比在动物中看到的要慢[78,79]。事实上,Mank认为植物中的单倍体表达可能代表了这两个王国之间已知的性染色体的最大差异[80]。S. latifolia性染色体进化了1000万年,虽然一些基因在SDR上丢失了,但自抑制重组发生以来,丢失率比具有相似时间的动物低60%[81,82]。对Rumex hastatulus中XY基因之间分化的估计表明,Rumex hastatulus的最小年龄为900万年[83],虽然一些基因也在Y上丢失[84],但花粉表达的基因丢失的可能性明显低于在二倍体组织中表达的基因[83,85]。单倍体C. purpureus UV性染色体每个包含超过3400个基因,其中一半被证明在配子体中表达[77]。除了单倍体基因表达外,缺乏退化可能是由于在许多植物中看到的SDR尺寸较小,因为当许多基因处于选择状态时,预计退化速度更快[86,87]。
Plant sex chromosomes are not without consequences from suppressed recombination. A consistent pattern found is an enrichment of transposable elements (TEs) and other repeats [77,88–90], which often accumulate in regions of low recombination [91]. In fact, in several species TE expansions have instead driven the Y chromosome to be larger than the X, such as in Coccinia grandis [92] and S. latifolia [93]. This pattern is counter to the smaller-Y heteromorphy found in the insects studied in Stevens’ day. While many Y chromosomes in animals are also riddled with repeats, in well-studied species most of the genes have been lost [94–97]. Moving beyond analyses of single species, comparisons between sister species with a shared evolution of sex chromosomes, such as in Coccinia [98], will provide greater insight into these degenerative processes.
植物性染色体并非没有受到抑制重组的后果。发现的一致模式是转座元件(transposable element, TEs)和其他重复序列的富集[77,88–90],这些重复序列通常在低重组区域积累[91]。事实上,在一些物种中,TE的扩增反而使Y染色体大于X染色体,例如在Coccinia grandis [92]和S. latifolia [93]中。这种模式与史蒂文斯时代研究的昆虫中发现的较小的Y异形性相反。虽然动物中的许多Y染色体也充满了重复染色体,但在经过充分研究的物种中,大多数基因已经丢失[94–97]。除了对单一物种的分析之外,对具有共同性染色体进化的姐妹物种之间的比较,例如Coccinia[98],将为这些退化过程提供更深入的见解。
The genes underlying the transition to dioecy, and subsequently the evolution of sex chromosomes, are also an area of interest in plants. Given many dioecious species are economically important, or closely related to ones that are, uncovering the genes that control reproductive structures is useful to breeding programmes. Additionally, these genes amass critical insight into how sex chromosomes evolve. In theory, the transition from hermaphroditic flowers to dioecy can occur through two mutations: one affecting female fertility, or carpel development, and another affecting male fertility, or stamen development [53,99]. Recent evidence in several plant species supports this two-gene model, such as in Actinidia deliciosa [100], A. officinalis [101] and P. dactylifera [102]. Contrastingly, a few systems have strong evidence of a single gene initiating female versus male development, as shown in persimmons and poplars [103,104]. Complementary to the many independent evolutions of dioecy, in each of these species examined, different genes have been identified as sex-determining and they function at varying parts of floral development (see reviews in [43,105]). Studies of additional independent origins of sex chromosomes in plants may indeed identify more novel genes involved in carpel and stamen pathways. Undisputedly, there is a veritable array of sex chromosomes found in plants (figure 3 and electronic supplementary material, table S1) and every species examined garners new insight on these fascinating parts of the genome.
向雌雄异株过渡的基因,以及随后的性染色体进化,也是植物感兴趣的一个领域。鉴于许多雌雄异株物种在经济上具有重要意义,或者与雌雄异株物种密切相关,因此揭示控制生殖结构的基因对育种计划很有帮助。此外,这些基因积累了对性染色体如何进化的重要见解。从理论上讲,从雌雄同体花到雌雄异株的转变可以通过两种突变发生:一种影响雌性生育能力或心皮发育,另一种影响男性生育能力或雄蕊发育[53,99]。最近在几种植物物种中的证据支持这种双基因模型,如Actinidia deliciosa [100]、A. officinalis [101]和P. dactylifera [102]。与此相反,一些系统有强有力的证据表明,雌性发育与雄性发育的基因是单一的,如柿子和杨树所显示的[103,104]。作为对雌雄异株的许多独立进化的补充,在所研究的每个物种中,不同的基因已被确定为性别决定因素,并且它们在花发育的不同部分发挥作用(参见[43,105]中的综述)。对植物中性染色体的其他独立起源的研究确实可能确定参与心皮和雄蕊途径的更多新基因。毋庸置疑,在植物中发现了一系列名副其实的性染色体(图3和电子补充材料,表S1),每个被检查的物种都对基因组的这些迷人部分有了新的认识。
4. The future of sex chromosome studies is through a multi-kingdom lens 4. 性染色体研究的未来是通过多王国的视角
Across the species Nettie Stevens studied, she found many that contained what she expected to find after their first discovery in mealworms: a heteromorphic XY pair. As we can see in the plants described, many also fitted the theoretical mould, but there are always exceptions that make us question the ‘rules’ at play for sex chromosomes [106]. In Stevens’ 1911 manuscript she writes, ‘At present, the all-important questions seem to me to be: What is the meaning of the differentiation of heterochromosomes in one form and not in others closely related? What has been the history of such differentiation where we have an unpaired heterochromosome or an unequal pair of heterochromosomes?’ She adds ‘… But in no case are we able to say when or how or why certain spermatogonial chromosomes became specially differentiated as heterochromosomes.’ [35].
在妮蒂·史蒂文斯(Nettie Stevens)研究的物种中,她发现了许多包含了她在首次发现黄粉虫后期望发现的东西:一对异形XY。正如我们在所描述的植物中看到的那样,许多植物也符合理论模式,但总有一些例外让我们质疑性染色体的“规则”[106]。在史蒂文斯1911年的手稿中,她写道:“目前,在我看来,最重要的问题是:异染色体以一种形式分化而在其他密切相关的形式中没有分化的意义是什么?当我们有一条不成对的异染色体或一对不相等的异染色体时,这种分化的历史是什么?她补充说:“…但在任何情况下,我们都无法说出某些精原染色体何时、如何或为什么被特别分化为异染色体。[35].
Today these questions remain at the heart of most studies on sex chromosomes. What drives gene gain and loss from the SDR, and what is the tempo at which these processes tick? The insights from plant sex chromosomes have highlighted differences that exist between them and animal systems, though there are ample similarities [80]. Yet, there are many independent evolutions across plants from which we can uncover more. Future studies could focus on the many existing genome references where the sex chromosomes have not been closely studied (figure 3). Nearly half of the dioecious orders do not have even a single dioecious genome reference, let alone ones at the genus or species level, highlighting the need for more genomic efforts focused on dioecious species (figure 3). Attention on more animal species is just as pressing, as well the other kingdoms that we have not focused on here, such as protists and fungi. Most critically, to answer these ongoing questions that have been posed for nearly as long as sex chromosomes have been known, we need to take a note from Stevens’ brilliant career and examine many isolates and many species across kingdoms.
今天,这些问题仍然是大多数性染色体研究的核心。是什么驱动了SDR的基因增益和损失,这些过程的运行速度如何?来自植物性染色体的见解突出了它们与动物系统之间存在的差异,尽管存在大量的相似之处[80]。然而,植物之间存在许多独立的进化,我们可以从中发现更多。未来的研究可以集中在许多现有的基因组参考上,在这些基因组参考中,性染色体尚未得到仔细研究(图3)。近一半的雌雄异株目甚至没有一个单一的雌雄异株基因组参考,更不用说属或物种水平的基因组参考了,这凸显了需要更多地关注雌雄异株物种的基因组工作(图3)。对更多动物物种的关注同样紧迫,以及我们在这里没有关注的其他王国,例如原生生物和真菌。最关键的是,为了回答这些几乎在性染色体已知以来就一直存在的问题,我们需要从史蒂文斯的辉煌职业生涯中吸取教训,并检查各个王国的许多分离株和许多物种。
There appears to be so little uniformity as to the presence of the heterochromosomes, even in insects, and in their behavior when present, that further discussion of their probable function must be deferred until the spermatogenesis of many more forms has been carefully worked out. [1]
关于异染色体的存在,甚至在昆虫中,以及它们在存在时的行为似乎都缺乏一致性,因此必须推迟对其可能功能的进一步讨论,直到仔细研究出更多形式的精子发生。[1]
Acknowledgements 确认
We thank Susanne Renner, Niels Müller and an anonymous reviewer for their helpful feedback on this manuscript. We also thank Marianne Weldon and Allison Mills for their assistance in searching the Bryn Mawr College Special Collections.
我们感谢 Susanne Renner、Niels Müller 和一位匿名审稿人对本文的有益反馈。我们还要感谢 Marianne Weldon 和 Allison Mills 在搜索 Bryn Mawr College 特别馆藏时提供的帮助。
Data accessibility 数据可访问性
The R script and materials to generate figure 3 can be found at https://github.com/sarahcarey/angiosperm_dioecy.
可以在 https://github.com/sarahcarey/angiosperm_dioecy 中找到用于生成图 3 的 R 脚本和材料。
Authors’ contributions 作者的贡献
S.B.C.: conceptualization, data curation, formal analysis, visualization, writing—original draft, writing—review and editing; L.A.: conceptualization, data curation, formal analysis, methodology, visualization, writing—original draft, writing—review and editing; A.H.: conceptualization, writing—original draft, writing—review and editing.
S.B.C.:概念化、数据管理、形式分析、可视化、写作——原始草稿、写作——审查和编辑;洛杉矶:概念化、数据管理、形式分析、方法论、可视化、写作——原始草稿、写作——审查和编辑;A.H.:构思、写作——初稿、写作——审查和编辑。
All authors gave final approval for publication.
所有作者均最终批准发表。
Competing interests 利益争夺
The authors declare no competing interests.
提交人声明没有竞争利益。
Funding 资金
This work was supported by NSF-IOS no. 2128196 to A.H.
这项工作得到了 NSF-IOS 第 2128196 至 A.H. 号的支持。
References 引用
- Stevens NM. 1905. Studies in spermatogenesis. Washington, DC: Carnegie Institution of Washington. See https://play.google.com/store/books/details?id=pP0CAAAAIAAJ.
- Ogilvie MB, Choquette CJ. 1981. Nettie Maria Stevens (1861–1912): her life and contributions to cytogenetics. Proc. Am. Phil. Soc. 125, 292-311. [PubMed]
- Stevens NM. 1901. Studies on ciliate infusoria. Proc. Calif. Acad. Sci. 3, 1-42.
- Mendel G. 1866. Versuche über Pflanzen-Hybriden. Verh. Naturforsch. Ver. Brunn 4, 3-47. (10.5962/bhl.title.61004) [In German.]
- Correns CFJE. 1900. Mendel’s Regel über das Verhalten der Nachkommenschaft der Rassenbastarde. Ber. Dt. Bot. Ges. 18, 158-167. [In German.]
- Tschermak E. 1900. Über künstliche Kreuzung bei Pisum sativum. Ber. Dt. Bot. Ges. 18, 232-239. [In German.]
- de Vries HM. 1900. Das Spaltungsgesetz der Bastarde. Ber. Dt. Bot. Ges. 18, 83-90. [In German.]
- Sutton WS. 1903. The chromosomes in heredity. Biol. Bull. 4, 231-250. (10.2307/1535741)
- Boveri T. 1904. Ergebnisse über die Konstitution der chromatischen Substanz des Zellkerns. Jena: G. Fischer. [In German.] See https://play.google.com/store/books/details?id=yopFsbbpihAC.
- Wilson EB. 1896. The cell in development and inheritance. New York, NY: Macmillan. See https://play.google.com/store/books/details?id=zHfjDY5o1GsC.
- Stevens NM. 1903. Further studies on the ciliate infusoria, Licnophora and Boveria. Arch. Protistenk. 3, 1-43.
- Brush SG. 1978. Nettie M. Stevens and the discovery of sex determination by chromosomes. Isis 69, 163-172. (10.1086/352001)
- Stevens NM. 1908. The chromosomes in Diabrotica vittata, Diabrotica soror and Diabrotica 12-punctata. A contribution to the literature on heterochromosomes and sex determination. J. Exp. Zool. 5, 453-470. (10.1002/jez.1400050402)
- Jones RN. 1995. B chromosomes in plants. New Phytol. 131, 411-434. (10.1111/j.1469-8137.1995.tb03079.x)
- Stevens NM. 1905. A study of the germ cells of Aphis rosae and Aphis oenotherae. J. Exp. Zool. 2, 313-333. (10.1002/jez.1400020302)
- Cattell JM. 1921. The biographical directory of American men of science. Science 53, 118. (10.1126/science.53.1362.118)
- Wilson EB. 1909. Recent researches on the determination and heredity of sex. Science 29, 53-70. (10.1126/science.29.732.53)
- Bachtrog D, et al… 2014. Sex determination: why so many ways of doing it? PLoS Biol. 12, e1001899. (10.1371/journal.pbio.1001899)
- Holleley CE, O’Meally D, Sarre SD, Marshall Graves JA, Ezaz T, Matsubara K, Azad B, Zhang X, Georges A. 2015. Sex reversal triggers the rapid transition from genetic to temperature-dependent sex. Nature 523, 79-82. (10.1038/nature14574)
- von Henking H. 1891. Untersuchungen uber die ersten Entwicklungsv-organge in den Eiern der Insekten. H. Uber Spermatogense und deren Beziehung zur Eientwicklung bei Pyrrhocoris apertus L. Z. Wiss. Zool. 51, 685-736. [In German.]
- McClung CE. 1899. A peculiar nuclear element in the male reproductive cells of insects. Zool. Bull. 2, 187-197. (10.2307/1535425)
- McClung CE. 1902. The accessory chromosome—sex determinant? Biol. Bull. 3, 43-84. (10.2307/1535527)
- Wallace LB. 1900. The accessory chromosome in the spider. Anat. Anz. 18, 327-329.
- Paulmier FC. 1899. The spermatogenesis of Anasa tristis. J. Morphol. 15, 224-272.
- Montgomery TH. 1904. Some observations and considerations upon the maturation phenomena of the germ cells. Biol. Bull. 6, 137-158. (10.2307/1535575)
- Stevens NM. 1906. Studies in spermatogenesis: a comparative study of the heterochromosomes in certain species of Coleoptera, Hemiptera and Lepidoptera, with especial reference to sex determination. Washington, DC: Carnegie Institution of Washington. See https://play.google.com/store/books/details?id=22W1AAAAIAAJ.
- Farmer JB, Moore JES. 1905. Memoirs: on the maiotic phase (reduction divisions) in animals and plants. J. Cell Sci. s2–48, 489-558. (10.1242/jcs.s2-48.192.489)
- Wilson EB. 1905. Studies on chromosomes. I. The behavior of the idiochromosomes in Hemiptera. J. Exp. Zool. 2, 371-405. (10.1002/jez.1400020305)
- Rheinberger HJ. 2000. Mendelian inheritance in Germany between 1900 and 1910. The case of Carl Correns (1864–1933). C. R. Acad. Sci. (Ser. III) 323, 1089-1096. (10.1016/s0764-4469(00)01267-1)
- Stevens NM. 1908. A study of the germ cells of certain Diptera, with reference to the heterochromosomes and the phenomena of synapsis. J. Exp. Zool. 5, 359-374. (10.1002/jez.1400050304)
- Stevens NM. 1912. The chromosomes in Drosophila ampelophila. In Proc. 7th Int. Zool. Congr., Boston**, 19–24 August 1907, pp. 380-381. Cambridge, MA: University Press.
- Stevens NM. 1910. An unequal pair of heterochromosomes in Forficula. J. Exp. Zool. 8, 228-241.
- Stevens NM. 1911. Heterochromosomes in the guinea-pig. Biol. Bull. 21, 155-167. (10.2307/1535884)
- Stevens NM. 1909. An unpaired heterochromosome in the aphids. J. Exp. Zool. 6, 115-123. (10.1002/jez.1400060105)
- Stevens NM. 1911. Further studies on heterochromosomes in mosquitoes. Biol. Bull. 20, 109-120. (10.2307/1536039)
- Boring AM. 1907. A study of the spermatogenesis of twenty-two species of the Membracidæ, Jassidæ, Cercopidæ and Fulgoridæ. J. Exp. Zool. 4, 469.
- Boring AM, Pearl R. 1914. The odd chromosome in the spermatogenesis. J. Exp. Zool. 16, 53-83. (10.1002/jez.1400160103)
- Boring AM. 1923. Notes by N. M. Stevens on chromosomes of the domestic chicken. Science 58, 73-74. (10.1126/science.58.1491.73)
- Guyer MF. 1916. Studies on the chromosomes of the common fowl as seen in testes and in embryos. Biol. Bull. 31, 221. (10.2307/1536235)
- Montgomery TH Jr. 1906. Chromosomes in the spermatogenesis of the Hemiptera Heteroptera. Trans. Am. Phil. Soc. 21, 97-173. (10.2307/1005443)
- Wilson EB. 1906. Studies on chromosomes. III. The sexual differences of the chromosomegroups in Hemiptera, with some considerations on the determination and inheritance of sex. J. Exp. Zool. 2, 507-545.
- Charlesworth D. 2022 Some thoughts about the words we use for thinking about sex chromosome evolution. Phil. Trans. R. Soc. B 377, 20210486. (10.1098/rstb.2021.0486)
- Renner SS, Müller NA. 2021. Plant sex chromosomes defy evolutionary models of expanding recombination suppression and genetic degeneration. Nat. Plants 7, 392-402. (10.1038/s41477-021-00884-3)
- Renner SS. 2014. The relative and absolute frequencies of angiosperm sexual systems: dioecy, monoecy, gynodioecy, and an updated online database. Am. J. Bot. 101, 1588-1596. (10.3732/ajb.1400196)
- Chase MW, et al. 2016. An update of the angiosperm phylogeny group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 181, 1-20. (10.1111/boj.12385)
- Yu G, Smith DK, Zhu H, Guan Y, Lam TT. 2017. ggtree: an R package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 8, 28-36. (10.1111/2041-210X.12628)
- Testolin R, Pilkington SM, Akagi T. 2021. Editorial: dioecy in fruit crops: the gender rise and decline and its agronomic impact. Front. Plant Sci. 12, 719588. (10.3389/fpls.2021.719588)
- Strasburger E. 1909. Zeitpunkt der Bestimmung des Geschlechts: Apogamie, Parthenogenesis und Reduktionsteilung. Jena, Germany: G. Fischer. [In German.]
- Strasburger E. 1909. Das weitere Schicksal meiner isolierten weiblichen Mercurialis annua-Pflanzen. Z. Bot. 1, 507-525. [In German.]
- Sykes MG. 1909. Note on the nuclei of some unisexual plants. Ann. Bot. 23, 341-341. (10.1093/oxfordjournals.aob.a089223)
- Allen CE. 1917. A chromosome difference correlated with sex differences in Sphaerocarpos. Science 46, 466-467. (10.1126/science.46.1193.466)
- Allen CE. 1940. The genotypic basis of sex-expression in angiosperms. Bot. Rev. 6, 227-300. (10.1007/BF02882399)
- Westergaard M. 1958. The mechanism of sex determination in dioecious flowering plants. Adv. Genet. 9, 217-281. (10.1016/s0065-2660(08)60163-7)
- Kihara H, Ono T. 1925. The sex-chromosomes of Rumex acetosa. Z. Indukt. Abstamm. Vererbungsl. 39, 1-7. (10.1007/bf01961517)
- Winge Ö. 1929. On the nature of the sex chromosomes in Humulus. Hereditas 12, 53-63. (10.1111/j.1601-5223.1929.tb02497.x)
- Wiens D, Barlow BA. 1979. Translocation heterozygosity and the origin of dioecy in Viscum. Heredity 42, 201-222. (10.1038/hdy.1979.23)
- Allen CE. 1945. The genetics of bryophytes. II. Bot. Rev. 11, 260-287. (10.1007/BF02861195)
- Renner SS, Heinrichs J, Sousa A. 2017. The sex chromosomes of bryophytes: recent insights, open questions, and reinvestigations of Frullania dilatata and Plagiochila asplenioides. J. Syst. Evol. 55, 333-339. (10.1111/jse.12266)
- Sousa A, Schubert V, Renner SS. 2020. Centromere organization and UU/V sex chromosome behavior in a liverwort. Plant J. 106, 133-141. (10.1111/tpj.15150)
- Wenger AM, et al. 2019. Accurate circular consensus long-read sequencing improves variant detection and assembly of a human genome. Nat. Biotechnol. 37, 1155-1162. (10.1038/s41587-019-0217-9)
- Arabidopsis Genome Initiative. 2000. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408, 796-815. (10.1038/35048692)
- Liu Z et al . 2004. A primitive Y chromosome in papaya marks incipient sex chromosome evolution. Nature 427, 348-352. (10.1038/nature02228)
- Yamato KT, et al… 2007. Gene organization of the liverwort Y chromosome reveals distinct sex chromosome evolution in a haploid system. Proc. Natl Acad. Sci. USA 104, 6472-6477. (10.1073/pnas.0609054104)
- Yu Q et al . 2009. A physical map of the papaya genome with integrated genetic map and genome sequence. BMC Genom. 10, 371. (10.1186/1471-2164-10-371)
- Bowman JL et al . 2017. Insights into land plant evolution garnered from the Marchantia polymorpha genome. Cell 171, 287-304. (10.1016/j.cell.2017.09.030)
- Charlesworth D. 2016. Plant sex chromosomes. Annu. Rev. Plant Biol. 67, 397-420. (10.1146/annurev-arplant-043015-111911)
- Prentout D, Stajner N, Cerenak A, Tricou T, Brochier-Armanet C, Jakse J, Käfer J, Marais GAB. 2021. Plant genera Cannabis and Humulus share the same pair of well-differentiated sex chromosomes. New Phytol. 231, 1599-1611. (10.1111/nph.17456)
- Cherif E, Zehdi-Azouzi S, Crabos A, Castillo K, Chabrillange N, Pintaud J-C, Salhi-Hannachi A, Glémin S, Aberlenc-Bertossi F. 2016. Evolution of sex chromosomes prior to speciation in the dioecious Phoenix species. J. Evol. Biol. 29, 1513-1522. (10.1111/jeb.12887)
- Krasovec M, Chester M, Ridout K, Filatov DA. 2018. The mutation rate and the age of the sex chromosomes in Silene latifolia. Curr. Biol. 28, 1832-1838. (10.1016/j.cub.2018.04.069)
- Norup MF, Petersen G, Burrows S, Bouchenak-Khelladi Y, Leebens-Mack J, Pires JC, Linder HP, Seberg O. 2015. Evolution of Asparagus L. (Asparagaceae): out-of-South-Africa and multiple origins of sexual dimorphism. Mol. Phylogenet. Evol. 92, 25-44. (10.1016/j.ympev.2015.06.002)
- Veltsos P et al . 2019. Early sex-chromosome evolution in the diploid dioecious plant Mercurialis annua. Genetics 212, 815-835. (10.1534/genetics.119.302045)
- Okazaki Y, Takahata S, Hirakawa H, Suzuki Y, Onodera Y. 2019. Molecular evidence for recent divergence of X- and Y-linked gene pairs in Spinacia oleracea L. PLoS ONE 14, e0214949. (10.1371/journal.pone.0214949)
- Sneep J. 1953. The significance of andromonoecism for the breeding of Asparagus officinalis L. II. Euphytica 2, 224-228. (10.1007/bf00053730)
- Wadlington WH, Ming R. 2018. Development of an X-specific marker and identification of YY individuals in spinach. Zücht. Genet. Breed. Res. 131, 1987-1994. (10.1007/s00122-018-3127-1)
- Li X, Veltsos P, Cossard GG, Gerchen J, Pannell JR. 2019. YY males of the dioecious plant Mercurialis annua are fully viable but produce largely infertile pollen. New Phytol. 224, 1394-1404. (10.1111/nph.16016)
- Cronk Q. 2022. Some sexual consequences of being a plant. Phil. Trans. R. Soc. B 377, 20210213. (10.1098/rstb.2021.0213)
- Carey SB, et al. 2021. Gene-rich UV sex chromosomes harbor conserved regulators of sexual development. Sci. Adv. 7, abh2488. (10.1126/sciadv.abh2488)
- Haldane JBS. 1933. The part played by recurrent mutation in evolution. Am. Nat. 67, 5-19. (10.1086/280465)
- Bull JJ. 1978. Sex chromosomes in haploid dioecy: a unique contrast to Muller’s theory for diploid dioecy. Am. Nat. 112, 245-250. (10.1086/283267)
- Mank JE. 2022 Are plant and animal sex chromosomes really all that different? Phil. Trans. R. Soc. B 377, 20210218. (10.1098/rstb.2021.0218)
- Chibalina MV, Filatov DA. 2011. Plant Y chromosome degeneration is retarded by haploid purifying selection. Curr. Biol. 21, 1475-1479. (10.1016/j.cub.2011.07.045)
- Bergero R, Charlesworth D. 2011. Preservation of the Y transcriptome in a 10-million-year-old plant sex chromosome system. Curr. Biol. 21, 1470-1474. (10.1016/j.cub.2011.07.032)
- Crowson D, Barrett SCH, Wright SI. 2017. Purifying and positive selection influence patterns of gene loss and gene expression in the evolution of a plant sex chromosome system. Mol. Biol. Evol. 34, 1140-1154. (10.1093/molbev/msx064)
- Hough J, Hollister JD, Wang W, Barrett SCH, Wright SI. 2014. Genetic degeneration of old and young Y chromosomes in the flowering plant Rumex hastatulus. Proc. Natl Acad. Sci. USA 111, 7713-7718. (10.1073/pnas.1319227111)
- Sandler G, Beaudry FEG, Barrett SCH, Wright SI. 2018. The effects of haploid selection on Y chromosome evolution in two closely related dioecious plants. Evol. Lett. 2, 368-377. (10.1002/evl3.60)
- Bachtrog D. 2008. The temporal dynamics of processes underlying Y chromosome degeneration. Genetics 179, 1513-1525. (10.1534/genetics.107.084012)
- Kaiser VB, Charlesworth B. 2009. The effects of deleterious mutations on evolution in non-recombining genomes. Trends Genet. 25, 9-12. (10.1016/j.tig.2008.10.009)
- Jesionek W, Bodláková M, Kubát Z, Čegan R, Vyskot B, Vrána J, Šafář J, Puterova J, Hobza R. 2021. Fundamentally different repetitive element composition of sex chromosomes in Rumex acetosa. Ann. Bot. 127, 33-47. (10.1093/aob/mcaa160)
- Zhou R, et al. 2020. A willow sex chromosome reveals convergent evolution of complex palindromic repeats. Genome Biol. 21, 38. (10.1186/s13059-020-1952-4)
- Montgomery SA, et al. 2020. Chromatin organization in early land plants reveals an ancestral association between H3K27me3, transposons, and constitutive heterochromatin. Curr. Biol. 30, 573-588. (10.1016/j.cub.2019.12.015)
- Kent TV, Uzunović J, Wright SI. 2017. Coevolution between transposable elements and recombination. Phil. Trans. R. Soc. B 372, 20160458. (10.1098/rstb.2016.0458)
- Sousa A, Fuchs J, Renner SS. 2017. Cytogenetic comparison of heteromorphic and homomorphic sex chromosomes in Coccinia (Cucurbitaceae) points to sex chromosome turnover. Chromosome Res. 25, 191-200. (10.1007/s10577-017-9555-y)
- Kubat Z, Hobza R, Vyskot B, Kejnovsky E. 2008. Microsatellite accumulation on the Y chromosome in Silene latifolia. Genome 51, 350-356. (10.1139/G08-024)
- Skaletsky H, et al. 2003. The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes. Nature 423, 825-837. (10.1038/nature01722)
- Wilson SMA, Makova KD. 2011. Genome analyses substantiate male mutation bias in many species. Bioessays 33, 938-945. (10.1002/bies.201100091)
- Bachtrog D. 2013. Y-chromosome evolution: emerging insights into processes of Y-chromosome degeneration. Nat. Rev. Genet. 14, 113-124. (10.1038/nrg3366)
- Peichel CL, et al. 2020. Assembly of the threespine stickleback Y chromosome reveals convergent signatures of sex chromosome evolution. Genome Biol. 21, 177. (10.1186/s13059-020-02097-x)
- Janousek B, Gogela R, Bacovský V, Renner SS. 2022 The evolution of huge Y chromosomes in Coccinia grandis and its sister Coccinia schimperi. Phil. Trans. R. Soc. B 377, 20210294. (10.1098/rstb.2021.0294)
- Charlesworth B, Charlesworth D. 1978. A model for the evolution of dioecy and gynodioecy. Am. Nat. 112, 975-978. (10.1086/283342)
- Akagi T, et al. 2019. Two Y-chromosome-encoded genes determine sex in kiwifruit. Nat. Plants 5, 801-809. (10.1038/s41477-019-0489-6)
- Harkess A, Huang K, van der Hulst R, Tissen B, Caplan JL, Koppula A, Batish M, Meyers BC, Leebens-Mack J… 2020. Sex determination by two Y-linked genes in garden asparagus. Plant Cell 32, 1790-1796. (10.1105/tpc.19.00859)
- Torres MF, et al. 2018. Genus-wide sequencing supports a two-locus model for sex-determination in Phoenix. Nat. Commun. 9, 3969. (10.1038/s41467-018-06375-y)
- Akagi T, Henry IM, Tao R, Comai L. 2014. A Y-chromosome-encoded small RNA acts as a sex determinant in persimmons. Science 346, 646-650. (10.1126/science.1257225)
- Müller NA, et al. 2020. A single gene underlies the dynamic evolution of poplar sex determination. Nat. Plants 6, 630-637. (10.1038/s41477-020-0672-9)
- Carey S, Yu Q, Harkess A. 2021. The diversity of plant sex chromosomes highlighted through advances in genome sequencing. Genes 12, 381. (10.3390/genes12030381)
- Furman BLS, Metzger DCH, Darolti I, Wright AE, Sandkam BA, Almeida P, Shu JJ, Mank JE. 2020. Sex chromosome evolution: so many exceptions to the rules. Genome Biol. Evol. 12, 750-763. (10.1093/gbe/evaa081)
via:
-
The contributions of Nettie Stevens to the field of sex chromosome biology - PMC
她在百年前破解性别之谜,却因性别而无缘荣誉
原创 科研圈
工作、攒钱、回归校园,43 岁拿到博士学位,大器晚成的内蒂・史蒂文斯(Nettie Stevens) 在短暂的科研生涯中取得了辉煌成果。她发现了 X/Y 性染色体,终结了上千年来人们对性别决定的种种臆测,却因为没有 Y 染色体而被人遗忘。
撰文 Brian Resnick/Vox
翻译 王可
审校 张士超

20 世纪初,生物学家 Stevens 解决一个困扰了人类千年的问题,这个问题说起来十分简单,但又万分难解:为什么有男女之分?她在布林茅尔学院 (Bryn Mawr)进行了开拓性的研究,终于发现了决定差异的原因 —— 性染色体。
蒙昧年代
多亏了 Stevens 的工作,以及在其基础上的后续研究,我们现在知道了性别是遗传所得,父亲的精子决定了后代的性别。但在人类之前漫长的历史中,这一问题显得十分神秘,并且催生出很多有意思的理论。
亚里士多德相信孩子的性别是由性行为时父亲的体温决定的。教科书《发育生物学》(Developmental Biology)中写道:“他劝年长的男性,如果想要生儿子,就在夏天准备这事。”
在 19 世纪的欧洲,人们广泛相信营养决定性别,营养差的父母生男孩,营养好的生女孩。几个世纪中,很多别的奇怪的理论也层出不穷。
十八世纪的法国解剖学家 Michel Procope-Couteau【《生男孩的艺术》(The Art of Having Boys)一书的作者】相信两个睾丸 / 卵巢分别对应两种性别。他 “建议控制后代性别的最佳方法就是移除属于另一种性别的一个睾丸或者卵巢;虽然还有一种不那么激烈的方法 — 女性侧躺(让正确性别的那侧在下),然后靠重力达到目的。” 这段话出自生物学家 Leo W. Beukeboom 和 Nicolas Perrin 所著《性别决定的演化》(The Evolution of Sex Determination)一书。
现在我们知道,以上理论都是无稽之谈。这都要感谢 Stevens 的研究。

Nettie Steven 用的显微镜。图片来源:Wikimedia Commons
大器晚成
Stevens 于 1861 年生于美国佛蒙特州,她在 35 岁 “高龄” 才攒到了足够的资金之后,进入加州的一所初创不久的大学 —— 斯坦福大学,在这里她开始学习科学并不断成长,于 1990 年拿到了本科和硕士学位。从斯坦福毕业后,Stevens 申请攻读宾州布林茅尔学院的博士学位 —— 这一教育水平对于当时女性来说是非常罕见的。
20 世纪初时,染色体携带遗传信息仍是一个新理论。孟德尔工作的内容在 1900 年才刚刚被人们 “重新发现”(他活着的时候没人听他的理论),科学界当时也在试图解答包括性别在内的各种性状如何在世代间传递。
Stevens 想要弄清楚性别到底如何(或者说是否)通过基因遗传。她在用一台显微镜观察黄粉虫(Tenebrio molitor)的染色体时,发现了几千年来逃过人们视线的秘密。
她发现雌性黄粉虫的细胞有 20 个大型染色体,而雄性虽然也有 20 个,但却有一个明显比另外 19 个小很多。Stevens 在总结报告中写道:“这看起来确实是染色体决定性别的现象。”
她推断,这样的差异能够追溯到黄粉虫的精子。的确,她发现黄粉虫精子的染色体有两种版本:有一条染色体或大或小。“含有一条小型染色体的精子(产生的后代)是雄性,” 她写道,“而那些 10 条染色体大小相似的精子产生的后代是雌性。”
“这是人类两千多年来对于动物植物和人类性别形成的思索和实验积累的成果”,历史学家 Stephen Brush 在他的《科学学会历史》(The History of Science Society)中解释道,“同时它为最近重新发现的孟德尔基因理论提供了一个重要的验证,后者已成为现代生物学的中心理论。”
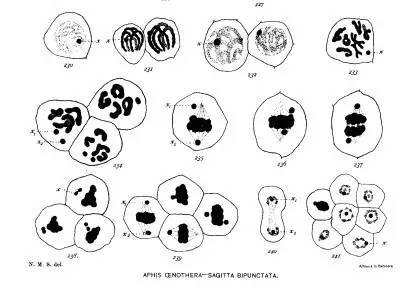
Steven 笔下的细胞分裂。 来源:Studies in Spermatogenesis
无缘荣耀
Stevens 的同事及前导师 ——E.B. Wilson,本身是一个传奇的生物学家,他常常被认作性染色体的发现者。
而导致这一情况的原因很简单:性别歧视。
Wilson 当时和 Stevens 一样,也在研究这个问题,并且在同一时期发表了相似的成果。然而他研究的物种中,雄性比雌性缺失一条染色体,这种情况在自然界中不那么常见。而 Steven 的 XY 染色体模型才是人类性别决定理论的基础。另外,Steven 的模型更好地支持了孟德尔的基因理论 —— 一部分基因起主导作用,而与它们相对应的等位基因的作用会被遮蔽。
“大家总是说 E. B. Wilson 和 Stevens 在同时观察到了同样的结果,”Brush 在书中这样写道,但 “很可能 Wilson 直到看到 Stevens 的结果,才得出性别决定的结论… 因为本来 Wilson 在其他领域已经有了很多成就,所以他总是被给予最多的荣誉。“
Wilson 文章比 Stevens 发得早,而且学术声望也更高,所以他一直被认为是发现性别决定的人。然而虽然这两篇文章相似,Stevens 的结论明显更加正确,且证据更充足有力。Wilson 仍然相信环境因素会对性别决定产生影响,而 Stevens 认为这完全取决于染色体。这两种观点中在当时都无法完全验证,但时间证明 Stevens 是正确的(译注:至少对于人类等哺乳动物性别由染色体决定,但确实有部分物种性别由环境决定),他们应该被认作共同发现者。尽管如此,只有 Wilson 一人获得了承认和荣誉。
这是一个 “玛蒂尔达现象” 的典型案例,这个名词来源于废奴主义者 Matilda Gage,指女性所取得的成就往往被归功于她们的男性同事 —— 他们成为共同作者、被夸大功劳、甚至完全掩盖她们所做的贡献。Stevens 绝不只是唯一一个有如此遭遇的女科学家:比如 20 世纪的 Rosalind Franklin,她的工作对于 DNA 的发现起到了决定性的作用,却被同样遗忘。
Stevens 在 1912 年死于乳腺癌。纽约时报为此发表了一篇讣告,这样总结她的成就:“她是极少数杰出女性科学家之一,也是当代生物学领域中的佼佼者。”
这真的是一个保守的评价。
原文链接:
https://www.vox.com/2016/7/7/12105830/nettie-stevens-genetics-gender-sex-chromosomes
via:
-
她在百年前破解性别之谜,却因性别而无缘荣誉_生物探索 原创 科研圈 环球科学科研圈 2017 年 07 月 20 日 18:20 北京
篇外 :性别的起源、流动、进化
不只是雌雄:性别的起源和流动
浙江大学生命演化研究中心 2023 年 08 月 06 日
01 丰富的性别定义:生理性别(Sex)与社会性别(Gender)
在传统意义上,人们对于性别的区分只有两种,即生理性别(Sex)为男或女。在当今社会,性别已经成为一个更加复杂和多样化的概念。人们对于性别的理解不止停留在生物性征,进而衍生出社会性别(Gender)这一更为弹性与复杂的社会学建构。2014 年,社交媒体平台 Facebook 通过性别研究组织 GLAAD 制定了 56 种性别可供选择。截止至 2023 年,全球范围已经出现了 107 种社会性别分类,其中不仅包括人们熟知的 LGBT、酷儿、无性别(Agender),还包括两性人(Androgyne)、顺性人(Cis)、性别流动(Gender Fluid)、泛性别(Pangender)等非传统性别 [1]。

32 种社会性别符号,图片来源 caaloba-d81ds6u
02 动植物中的性别
让我们回到传统定义的生理性别,自然界中的生物并不仅仅局限于固定不变的雌雄二分,而是存在着更为多样和复杂的性别类型。在哺乳动物中,性别一般是由 XX(♀)和 XY(♂)性染色体决定的,而鸟类性别一般由 ZW(♀)和 ZZ(♂)性染色体决定,这两套遗传性别决定系统同样广泛存在于其他的动植物中。我们较为熟知的动物大都为雌雄异体(dioecy or gonochorism),雌雄生殖器官分别出现在不同的个体中,每个个体在其一生中仅以一种性繁殖角色存在 [2]。且雌雄个体会显示出明显的性二态性(sexual dimorphism)[3-4](即同一物种不同性别之间的差别),比如雌雄之间的体型大小差异,器官形态差异以及羽毛颜色的差异等等。

动物性二态性示例,图片来源 Adams Lab
然而,动物中同样存在 “无性别者”,即一个个体内同时拥有雄性和雌性的生殖器官的雌雄同体。事实上,雌雄异体的昆虫在动物中占有很大比例,而除去昆虫后,大约三分之一的动物属于雌雄同体 [5]。雌雄同体在无脊椎动物中很常见,例如刺胞动物门的珊瑚、海葵、水螅,软体动物门的蜗牛以及多种蠕虫物种。

雌雄同体刺胞动物门物种以及蜗牛示例
相比之下,大部分植物为雌雄同株,比如一朵花中既有雄蕊也有雌蕊(两性花),或是一棵植株上同时存在雄花和雌花。仅有 10% 的陆地植物是雌雄异株,而在被子植物(开花植物)中,仅有 6% 是雌雄异株 [6]。

慈姑包含雌雄异株和雌雄同株的不同种群,图片来源 Barrett Lab
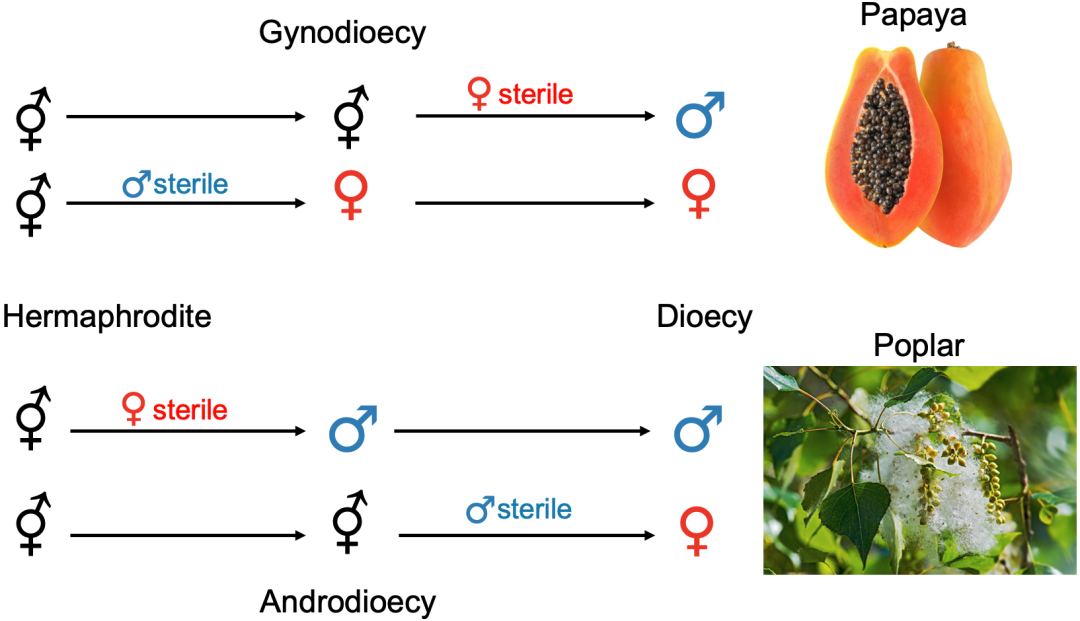
雄性雌雄同体(androdioecy,在植物中称为雄全异株)是指雄性和雌雄同体的个体共存在同一物种种群中,例如我们所熟悉的秀丽隐杆线虫,不过这种性别类型在植物和动物中都很少见。同样,雌性雌雄同体(gynodioecy,在植物中称为雌全异株)是指雌性和雌雄同体个体共存在同一物种种群中,在植物中较为常见。

上为秀丽隐杆线虫,种群中存在雄性和雌雄同体个体;
下为白玉草,种群中存在雌株(雌花植株)以及雌雄同株(两性花植株)
03 性别的起源
性别的起源是生物进化过程中的一个重要事件。相较于无性生殖,有性生殖更复杂,难度更高,但有性生殖通过不同个体间的精卵结合,进行基因交换,使得后代获得更高的遗传多样性,在环境中具有更强的适应能力与生存能力。植物和动物一些分支群中的雌雄同体物种既可以自体受精,也可以选择与其他个体进行异体受精进行有性生殖。雌雄独立性别的从头演化主要基于开花植物中的研究 [7-8],因为大部分植物是雌雄同体,并且已有研究估计在被子植物中雌雄异体已经独立发生了数百次演化 [9-10]。雌雄同体的独立性别演化至少需要两个突变,首先通过初始突变产生雌性 / 雄性个体,之后通过第二个突变产生雄性 / 雌性个体。通过雌全异株的中间状态,在性别类型转换过程中雌性能够完全避免近亲繁殖的压力。相较之下,若雄全异株作为中间状态,雄性个体需要与雌雄同体竞争向胚珠中传递花粉,作为生殖过渡成本更高。这些理论依据有助于解释在植物中观察到的雌全异株的普遍现象。
植物中雌雄同体到雌雄异体的演化 “两步” 途径,以木瓜和杨树示例
04 性别的流动
性别的演化方向是可逆的,例如在植物中,物种可以从雌雄异体演化为雌雄同体,也可以从雌雄同体演化为雌雄异体。如果在一个种群中,所有的物种都是雌雄同体,那么雌雄同体个体中的雄性和雌性功能应该是平均分配以获得同等的适应度。但如果种群中出现了雌性个体(与雌雄同体相比,其雌性功能占比更高),雌雄同体会通过增加对雄性功能的分配与其他雌雄同体竞争,以将花粉传播至雌性的胚珠;如果资源有限,接着就会降低对雌性功能的分配;进而建立两性群体。在某些情况下,雌雄异体种群中的雌性或者雄性个体都有可能被选择成为雌雄同体个体。例如,当雌雄异体种群迁移至新的生境时,雄性频率的改变(不稳定性)会对种群产生极大的影响。因此,对于雄性个体数量少的种群来说,雌性个体即使只获得少部分的雄性功能,都会产生自体繁殖的优势。在之后的演化过程中若雄性个体完全消失,雌雄同体将分配更多资源在雄性功能上 [4]。
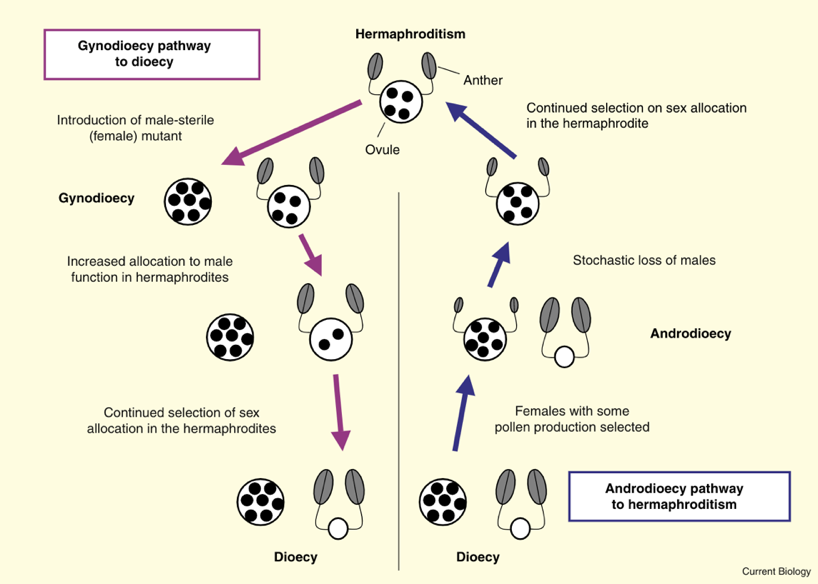
植物中雌雄同体和雌雄异体性别类型转换的常见途径 [4]
相较于植物,雌性雌雄同体在动物中极为罕见,到目前为止,只有极少数雌性雌雄同体的动物被报道过(例如脊椎动物盲鳗以及环节动物中的一个物种)[11]。雄性雌雄同体在植物和动物中都很少见,并且通常代表从雌雄异体到雌雄同体的过渡状态(如雄性雌雄同体 Caenorhabditis elegans)[11-12]。目前关于动物的性别类型转换案例很少,不同性别类型之间转换的分子机制仍鲜为人知。不过与植物类似,动物从雌雄异体祖先演化为雌雄同体的过程似乎仅限于在雌性个体中进行雌性功能的重新分配;例如雌雄同体取代雌性,与雄性共存。然而动物从雌雄同体转换为雌雄异体的途径中,更倾向于通过雄性雌雄同体作为中间状态,这与植物中的转换途径形成鲜明对比 [11]。在动物界中,从雌雄同体祖先到雌雄异体的转变可能涉及性二态性中体积大小的选择。举例来说,几乎所有扁形虫都是雌雄同体的,但血吸虫科中的寄生虫 —— 血吸虫却是个例外。血吸虫不是传统意义上的雌雄异体,雌性血吸虫需要与雄性血吸虫 “抱对” 后性器官才能发育成熟,并进行交配受精。雄性血吸虫个体体积较大,肌肉更丰富,这增加了它们 “抱对” 时的紧握和运动能力。而雌性个体则相对较小,受精后能够在血管内准确移动并高效地产卵 [13-14]。
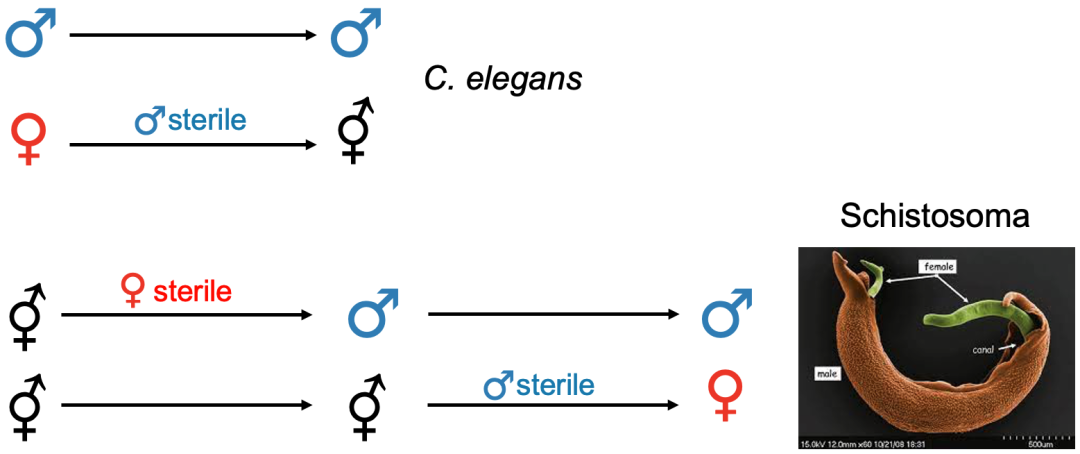
血吸虫,图片来源 New Frontiers of Molecular Epidemiology of Infectious Diseases
后记
在自然界中,各个生物种群都会选择适合自己的方式,繁衍后代。这些性别类型的存在丰富了生物界的多样性,为不同物种的适应和进化提供了不同的策略和机制,也等待着我们的进一步探索和深入理解。
参考网址以及文献
https://en.wikipedia.org/wiki/Sex–gender_distinction
BAWA K S. Evolution of dioecy in flowering plants [J]. Annual review of ecology and systematics, 1980, 11 (1): 15-39.
HEDRICK A V, TEMELES E J. The evolution of sexual dimorphism in animals: hypotheses and tests [J]. Trends Ecol Evol, 1989, 4 (5): 136-8.
DELPH L F. Sex allocation: evolution to and from dioecy [J]. Current Biology, 2009, 19 (6): R249-R51.
JARNE P, AULD J R. Animals mix it up too: The distribution of self-fertilization among hermaphroditic animals [J]. Evolution, 2006, 60 (9): 1816-24.
MING R, BENDAHMANE A, RENNER S S. Sex chromosomes in land plants [J]. Annual review of plant biology, 2011, 62: 485-514.
RENNER S S, MÜLLER N A. Plant sex chromosomes defy evolutionary models of expanding recombination suppression and genetic degeneration [J]. Nat Plants, 2021, 7 (4): 392-402.
HENRY I M, AKAGI T, TAO R, et al. One hundred ways to invent the sexes: theoretical and observed paths to dioecy in plants [J]. Annu Rev Plant Biol, 2018, 69: 553-75.
CHARLESWORTH D. Distribution of dioecy and self-incompatibility in angiosperms [M]//GREENWOOD P J, SLATKIN M. Evolution – Essays in Honour of John Maynard Smith. Cambridge; Cambridge University Press. 1985: 237-69.
RENNER S S. The relative and absolute frequencies of angiosperm sexual systems: dioecy, monoecy, gynodioecy, and an updated online database [J]. Am J Bot, 2014, 101 (10): 1588-96.
WEEKS S C. The role of androdioecy and gynodioecy in mediating evolutionary transitions between dioecy and hermaphroditism in the Animalia [J]. Evolution, 2012, 66 (12): 3670-86.
CHARLESWORTH D. Androdioecy and the evolution of dioecy [J]. Biol J Linn Soc, 1984, 22 (4): 333-48.
PLATT T R, BROOKS D R. Evolution of the schistosomes (Digenea: Schistosomatoidea): the origin of dioecy and colonization of the venous system [J]. J Parasitol, 1997, 83 (6): 1035-44.
WANG Y, GASSER R B, CHARLESWORTH D, et al. Evolution of sexual systems, sex chromosomes and sex-linked gene transcription in flatworms and roundworms [J]. Nature Communications, 2022, 13 (1): 3239.
via:
-
不只是雌雄:性别的起源和流动丨浙江大学生命演化研究中心 2023 年 08 月 06 日
性别,究竟能有几种:2 种?上万种?
原创 环球科学 2022 年 05 月 24 日

图片来源:Unsplash
性别远比看起来复杂。现在有越来越多的证据表明,性别不是简单的二元法。而且,追溯性别起源的科学家也在真菌身上发现了令人震惊的事实 —— 有些真菌不仅不只有 2 种交配型,它们甚至还演化出了上千、上万种交配型。但这些交配型等同于性别吗?
撰文 | 王怡博
审校 | clefable
不知道你有没有遇到过,在一些网站填写个人信息时,在性别(社会性别,gender)那一栏的下拉菜单里除了男和女之外,还有第三个选项:其他。事实上,与社会性别类似,生理性别(sex)也是一个非常复杂的问题。
那么,是什么决定了人的性别(以下通指生理性别)?我们对此通常会不假思索地说:“性染色体。”
目前已知人类拥有 2 种性染色体:X 和 Y。通常而言,女性有 2 条大小、形态完全相同的 X 染色体(XX),男性的性染色体则是 X 和 Y(XY),而且 Y 比 X 染色体小得多。1990 年,科学家发现 Y 染色上有一个叫做 *SRY*的基因能够决定雄性性别。 研究发现,如果 SRY 的转录出现异常,或移除 SRY ,可能会导致性别翻转,也就是说,可以让性染色体为 XY 的个体发育为雌性;而如果给一只性染色体为 XX 的老鼠导入 SRY ,则可能使其拥有雄性特征。
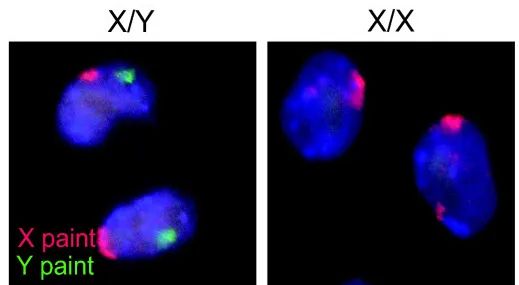
通常而言,男性的性染色体为 XY,女性的为 XX。图片来源:wikimedia commons, CC BY 2.0
自SRY发现以来,它就一直被认为只含有 1 个外显子(包含合成蛋白质所需的核心信息)。但是在 2020 年 10 月,一项发表于《科学》的研究表示,成年小鼠 Y 染色体上的SRY其实还有第二外显子。而且,包含 2 个 *SRY*外显子信息的转录本(被命名为 SRY-T)可能才是真正的雄性决定因素。他们发现,缺少 SRY-T 的雄性小鼠会发育为雌性,而携带 SRY-T 的雌性小鼠则能转变为雄性。更重要的是,实验成功率几乎为 100%,而此前性别翻转实验的成功率则在约 25%~75% 之间。
除了 Y 染色体上的性别决定基因外,科学家还在 2000 年左右发现,雌性也并非总是 “被动” 发育出来的,例如WNT4*基因能够促进卵巢发育,同时抑制睾丸发育。另外,一种叫做RSPO1*的卵巢基因也可能会导致性腺发育异常,例如导致性染色体为 XX 的个体发育出卵睾(ovotestis)—— 一种同时存在睾丸和卵巢组织的性腺。
这些结果都表明,性别远比看起来复杂。关于人的性别决定因素的研究仍在继续,科学家也依然在尝试进一步了解是否存在额外决定性别的基因,以及其他生物学因素,如性激素。
与此同时,还有一些科学家则着眼于真菌 —— 人类远亲的 “性别”。其中一个关键问题就在于,它们是否也有性别?
真菌有上万种性别?
正如美国哈佛大学医学院 Andrew Murray 教授所说:“人类会发生性关系,真菌也是如此。”
或者说,真菌也能发生有性生殖,而且它们的生殖周期也包括细胞核融合、减数分裂等过程。不过相比于人类,真菌没有完整的性染色体,它们的交配是由基因组上特定的小区域控制的,相关区域被称为交配型(MAT)位点。
而且,既然要发生关系、进行交配,我们就暂且不讨论自交的情况,而只关注异宗配合(heterothallism,指单个菌株不能完成有性生殖)的有性生殖方式。
事实上,真菌拥有 2 种异宗配合的有性生殖方式:双极交配和四极交配。它们分别是指真菌在减数分裂后能产生 2 种和 4 种交配型,且分别由 1 个和 2 个 MAT 位点控制。例如,绝大多数子囊菌(Ascomycota)属于双极交配,而对于担子菌(Basidiomycota)而言,既有一部分属于双极交配,也有一部分属于四极交配。其中,有研究表明,伞菌纲(担子菌门)中有 65% 的真菌属于四极交配,而且值得一提的是,伞菌纲还有一个特点:它们 2 个 MAT 位点上分别有多个等位基因,因此有些伞菌纲甚至拥有成百上千种交配型。
例如,一项在 2022 年 3 月发表于PLOS Genetics的研究利用长读测序的方法,确定附毛菌属(Trichaptum,担子菌门)MATA 和 MATB 位点上分别有大约 270 个和 65 个等位基因。此前已有研究证实,对于受 2 个 MAT 位点控制的真菌来说,只有双方在这 2 个位点互相完全不一致时才有可能发生交配。因此,那些主要生长在较冷地区树干上的附毛菌属或许能拥有 17 550 种交配型。这意味着它们有高达 98%(269×64/270/65=98%)的可能性与同类发生关系,从而进行交配,挪威奥斯陆大学的遗传学家 David Peris(这项研究的第一作者)说道,而且如此多的遗传多样性很有可能是为适应环境而演化出来的。

一种支架真菌,附毛菌属。图片来源:Keith Weller/wikimedia commons, public domain
伦敦大学学院演化生物学教授 Nick Lane 曾在《能量、性、死亡:线粒体与我们的生命》(Power, Sex, Suicide: Mitochondria and the Meaning of Life)一书中表示,如果种群中出现了一个突变体,能够与已有的 2 种交配型发生关系,那么这种突变体必然会迅速繁衍,因为它能选择的性伴侣增加了 1 倍。
但问题在于:此时的交配型是否等于性别?或者说,这是否意味着真菌拥有令人难以置信的上千、甚至上万种性别?
性别与交配型
Lane 也提到,有一种叫做裂褶菌(Schizophyllum commune)的担子菌拥有约 28 000 种交配型。其中一个 MAT 位点上有 300 多个等位基因,而另一个则具有 90 多个等位基因。同样的,当双方 2 个 MAT 位点的基因都各不相同时,便能自由配对,从而使它们能与种群中 99% 的个体自愿发生关系。但它们如何知道哪一个是自己的母亲,哪一个是父亲呢?
结果发现,这些真菌之间其实是无爱的关系 —— 它们只是让双方的遗传物质结合,然后就 “分开” 了,Lane 解释道。即使我们可以说它们拥有 28 000 种性别,倒不如说它们其实根本就没有性别,用交配型来表示 “性别” 就足以。
Joseph Heitman 也持同样的观点。Heitman 是美国杜克大学分子遗传学与微生物学学院的教授,他和同事在《真菌王国》(the Fungal Kingdom)一书中写道,事实上,大多数真菌都属于同配生殖(isogamous),也就是说,它们有性生殖进行交配的两个配子具有相同的形态、大小和结构,因此性别分化不明显,而常常用正负号表示。而在动物或植物进行有性生殖时,则往往产生较小的雄配子(如精子)和较大的雌配子(如卵细胞)。
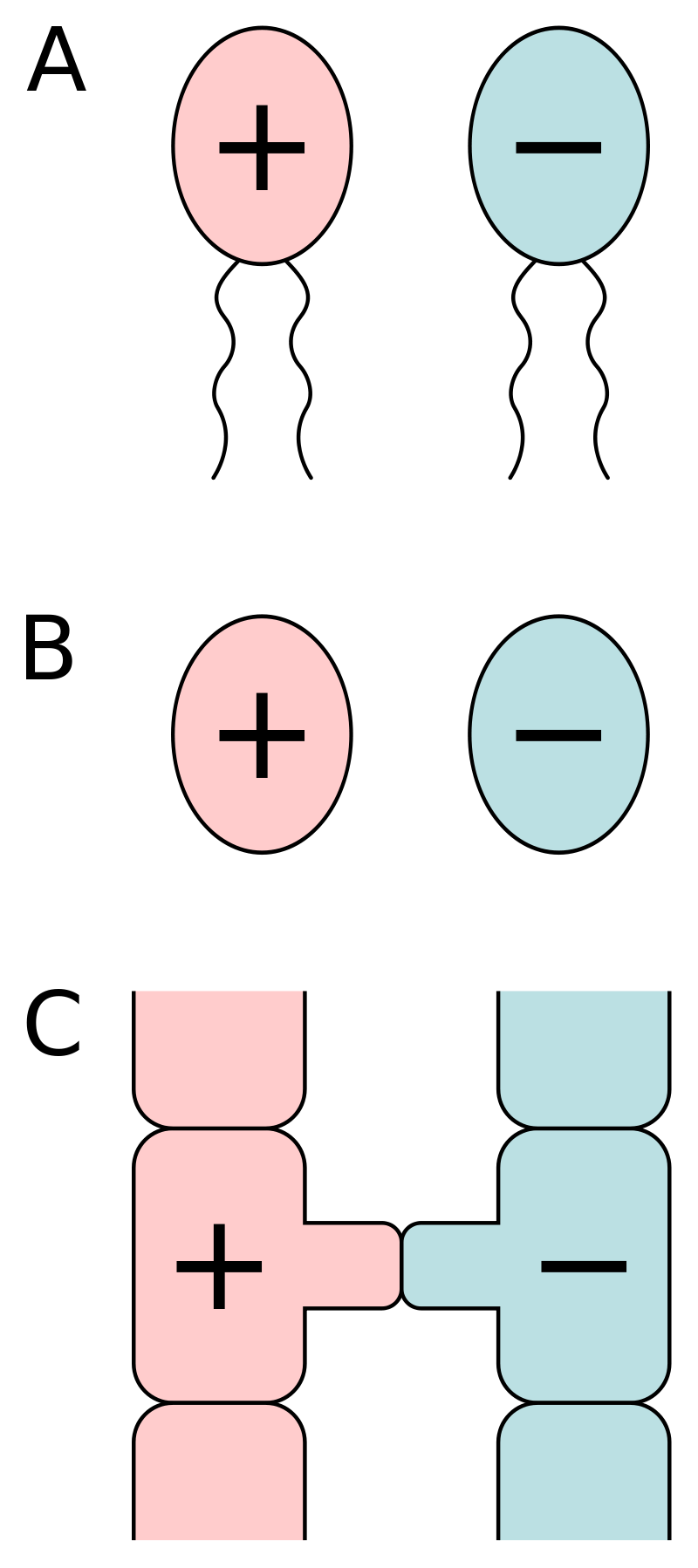
同配生殖时,两个配子具有相同的形态、大小和结构,因此性别分化不明显,而常常用正负号表示。图片来源:wikimedia commons, public domain
不过,真菌界是否也有特例呢?
子囊菌进行有性生殖时的确比较特别,因为它们能发育出性器官:产囊体(ascogonium)和雄器(antheridium)。例如,粗糙脉孢菌(Neurospora crassa)有 2 种交配型 A 和 a,假设 A 的分生孢子所释放的信息素特别吸引 a 的受精丝(trichogyne,一种菌丝),那么二者就会互相接触并实现细胞 - 菌丝的融合,此时 A 的细胞核就会经受精丝迁移到 a 的生殖结构 —— 产囊体内。这类似于精子细胞核进入卵细胞的过程。随后,A 和 a 的细胞核就会在产囊体内依次进行有丝分裂、配对,以及减数分裂等复杂过程,并最终形成子囊孢子,从而完成一次有性生殖。我们可以看到,A 在这里或许可以当作提供精子的雄性,而 a 则相当于拥有生殖结构及卵细胞的雌性。
对于科学家来说,探究真菌的性别和交配关系的目的之一是了解性别的起源,但这条路并不那么容易。有科学家发现,即使是古老的真核生物之间也存在性行为,但雄性和雌性的概念其实是很晚才演化出来的。加拿大不列颠哥伦比亚大学的动物学家 Sarah Otto 表示,了解生物拥有如此多样的繁殖方式,会让我们意识到,我们普遍持有的男女或雄雌的二元性别观念很有可能只是冰山一角。
封面来源:Unsplash
参考链接:
https://journals.plos.org/plosgenetics/article?id=10.1371/journal.pgen.1010097#abstract0
https://www.the-scientist.com/news-opinion/this-fungus-has-more-than-17-000-sexes-69930
https://doi.org/10.1128/9781555819583.ch7
https://www.nature.com/articles/nature06453
https://journals.asm.org/doi/10.1128/EC.5.3.544-554.2006
https://www.ibiology.org/archive/yeast-life-cycle/
https://en.wikipedia.org/wiki/Sex
https://en.wikipedia.org/wiki/Mating_in_fungi
via:
-
性别,究竟能有几种:2 种?上万种?原创 环球科学 2022 年 05 月 24 日 21:20 北京
Nature:性别进化的古老线索
来源:生物通 2008 年 01 月 11 日
真菌并没有精确区分开雌雄,但是确实存在性别差异,来自杜克大学医学院分子遗传与微生物学系,密苏里大学堪萨斯分校(University of Missouri, Kansas City,生物通注)生命科学学院的研究人员近期发现了一些真菌早期的进化形式,为高等动物 —— 包括真菌的远亲:人类 —— 提供了新线索。这一研究成果公布在《Nature》杂志封面上。
生物通报道:真菌并没有精确区分开雌雄,但是确实存在性别差异,来自杜克大学医学院分子遗传与微生物学系,密苏里大学堪萨斯分校(University of Missouri, Kansas City,生物通注)生命科学学院的研究人员近期发现了一些真菌早期的进化形式,为高等动物 —— 包括真菌的远亲:人类 —— 提供了新线索。这一研究成果公布在《Nature》杂志封面上。
这一研究由 Joseph Heitman 领导,主要研究手段是通过从一种最古老的真菌类型:Phycomyces blakesleeanus(布拉克须霉,生物通注)分离得到了性别决定基因。
布拉克须霉作为感光模型最早由诺贝尔奖获得者 Max Delbrück 采用 ——Max Delbrück、Alfred D. Hershey 和 Salvador E. Luria 组成的小组进行了著名的噬菌体实验,证明 DNA 就是遗传信息的物质载体。这一杰出的实验成就直接导致了 DNA 双螺旋结构的发现,并因此奠定了分子遗传学乃至整个分子生物学的基础。
在这项新研究中,布拉克须霉作为了另外一种不同领域的研究模型,即性别的决定过程。与其它真菌一样,布拉克须霉没有完整的性染色体,就像人类的 X 或 Y 染色体,它的性别决定是由基因组的一个小区域控制的,这个区域称为交配型位点(Mating type locus)。在许多进化水平上更高级的真菌物种中都发现过交配型位点,并且表现出不寻常的差异种类数量,甚至在相似的真菌物种中都存在这些差异,这让科学家们感到奇怪:这是如何进化的?
Heitman 研究小组认为真菌这一最早形式的性别决定方式也许揭示了交配型位点的一种古老结构,就像是一种分子化石。Heitman 表示,“真菌是研究人类性别差异进化的适合模式,这是因为真菌决定性别的遗传序列比人类性别决定的染色体要小。”
为了识别出布拉克须霉的交配型位点,研究人员利用计算机比较已知的其它真菌种系基因组种的交配型位点,然后绘制遗传图谱,“我们通过一种 usual-suspects 方法,比较了不同类型真菌蛋白,然后识别出了一种在所有种系都相关的‘候选人 '”。
利用这一 DNA,研究人员分离得到了调控配对的一种基因的两个版本,即 sexM (sex minus) 和 sexP (sex plus),具有这两种类型中的一种的真菌可以相互配对。
而且这两个版本的基因都能编码一种称为高迁移率蛋白质(high-mobility group protein,生物通注)位点蛋白,这种蛋白能通过一种未知途径调控性别差异。这种蛋白与人类 Y 染色体上一个主要调控因子:SRY 蛋白十分相似,因此研究人员认为这种相似性说明 HMG 位点蛋白也许标记了真菌和人类性别决定的进化开端。
(生物通:张迪)
原文摘要:
Nature 451, 193-196 (10 January 2008) |
doi:10.1038/nature06453; Received 13 August 2007; Accepted 5 November 2007
Identification of the sex genes in an early diverged fungus
附:
从物理学家到生物学家
1952 年,德尔布吕克、美国遗传学家赫尔希 (Alfred D. Hershey) 和意大利生物学家卢里亚 (Salvador E. Luria) 组成的小组进行了著名的噬菌体实验,证明 DNA 就是遗传信息的物质载体。这一杰出的实验成就直接导致了 DNA 双螺旋结构的发现,并因此奠定了分子遗传学乃至整个分子生物学的基础。因为这一成就,德尔布吕克等三人荣获诺贝尔生理学和医学奖。
德尔布吕克在斯德哥尔摩接受诺贝尔奖牌时为 1969 年,然而早在三十年前德尔布吕克就已经发表了划时代的科研成果:
1935 年,德尔布吕克与前苏联遗传学家梯莫菲也夫 - 雷索夫斯基 (Timofeeff-Ressovsky ) 和物理学家齐默尔 (K. G. Zimmer) 合作,应用物理学概念研究果蝇的 X 射线诱变现象,建立了一个突变的量子模型。他们三人共同署名的论文题为《关于基因突变和基因结构的性质》,刊登在德国哥廷根的科学协会通讯上,此文建立了遗传基因原子物理学的模型,并倡导 “遗传基因的高分子学说”,给理论遗传学打上了物理学的烙印。这是世界上第一个基因模型的提出,比沃森 (James Watson ) 和克里克 (Francis Crick ) 提出著名的 DNA 双螺旋结构模型整整早了 18 年。
生命的谜题
麦克斯・德尔布吕克 (Max Delbrück) 1906 年 9 月 4 日出生于德国柏林,其父为柏林大学历史学教授,母亲是著名化学家李比希 (J. Liebig) 的孙女。德尔布吕克从童年起就对科学有着浓厚兴趣。他早期的兴趣在天文学,后来转而从事量子力学。1930 年他在格丁根大学麦克斯・玻恩 (Max Born) 的指导下获理论物理学博士学位。然而他很快便对他的专业失去了热情:主流的思想和大的发现都出现于 20 世纪中期。1932 年,玻尔在哥本哈根举行的国际光疗会议上发表了《光和生命》的著名演讲,应用物理学的概念来解释生命现象。在当时,人们很难理解玻尔这些科学思想的意义,一些听讲的生物学家甚至不知所云。然而,玻尔以一种天才的直觉能力,借助于量子力学的范例,预感到在生物学中将有某些新的发现。这无疑给人们一种深刻的启示,并向当时的物理学家和生物学家提出了挑战。这引发了年轻的德尔布吕克向生物学以及生命的谜题探索的兴趣。从此德尔布吕克将一向彼此分离的物理学与生物学联结起来。他将物理学的模型和方法应用于活体生物研究。德尔布吕克关于噬菌体试验的论文发表于 1939 年。在序言中指出:“过去有种错误观点认为,细菌病毒是一种化学分子而非生物。我们的工作则表明,某些大蛋白分子 ( 病毒) 具有在物体内增殖的特性。” 这一试验被公认为 “现代” 噬菌体遗传学之开端。
为了实验的需要,德尔布吕克邀请意大利细菌学家卢里亚到纽约冷泉港实验室,共同研究噬菌体遗传现象。1943 年 1 月,卢里亚设计了另一项著名的 “波动试验”,结果表明:细菌的耐噬菌体突变的产生是随机的。这一结果表明细胞变种的随机自发的选择性,成为细菌遗传学的一项突破,动摇了拉马克主义关于细菌只有获得性遗传而无突变性的偏见。1 月底,赫尔希加入了他们的行列,“噬菌体学派” 的核心形成了。
麦克斯・德尔布吕克分子医学中心
德尔布吕克的专业贡献在科学界的影响力是毋庸置疑的,很多的科学研究机构都是以德尔布吕克命名:例如 1922 年组建的坐落于柏林布赫的麦克斯・德尔布吕克分子医学中心 (MDC)。与其他的科研机构一道,每年 MDC 授予卓越的研究者们麦克斯・德尔布吕克奖章。
麦克斯・德尔布吕克与许多的德裔科学家一样,在纳粹时代移居美国,更好的科研条件也是一个有力因素,战后他也没有再回德国。然而他仍情系故土,支持德国科学界的重建:他赞助创建科隆大学遗传学研究所并且在 1961 至 1963 年间担任所长。他也极力支持康斯坦茨大学生物系的创建。
德尔布吕克开辟了现代分子生物学的先河。他的研究工作对于后代的生物学研究者们仍具有重大科研意义。关于 “生命的谜题”,遗传分子的基础问题,德尔布吕克没有予以解答,这一任务势必落在他的学生詹姆斯 (James D. Watson) 肩头。
via:
-
Nature 头条:性别的古老传说 - 生物通 2008

















 538
538

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









