






一、写在前面
29日上午有幸在现场听了汤富酬老师进行了题为“新一代单细胞组学测序技术“的精彩报告。其中涉及到一篇"Chromatin accessibility landscape of mouse early embryos revealed by single-cell NanoATAC-seq2"的文章。汤老师开发了使用长读长测序的单细胞ATAC-seq,以揭示小鼠着床发育前的染色质可及性图谱。NanoATAC-seq可以精准的捕捉重复元件的染色质状态,能够揭示转录因子在胚胎发育中的共表达网络与顺式调控功能。不得不感叹一句,十六年过去了,单细胞发明者依旧在发明单细胞!或许单细胞组学+三代测序会是一个新的行业增长点与风口。
感兴趣的同学可以阅读一下原文:
Li M, Jiang Z, Xu X, Wu X, Liu Y, Chen K, Liao Y, Li W, Wang X, Guo Y, Zhang B, Wen L, Kee K, Tang F. Chromatin accessibility landscape of mouse early embryos revealed by single-cell NanoATAC-seq2. Science. 2025 Mar 28;387(6741):eadp4319.
二、背景介绍
受精后,合子基因组保持转录静止,合子依靠母体因素进一步发育。两波合子基因组激活(ZGA)确保了母体向合子的成功过渡(MZT),这伴随着合子染色质的广泛表观遗传重编程。次要的ZGA开始于单细胞中期,转录活性相对较低,而主要的ZGA发生在两细胞中期至晚期,此时胚胎逐渐激活数千个基因和一组转座元件来控制随后的发育。参与ZGA起始的关键因素包括Obox家族、Zscan4基因簇和转座元件,如长穿插核元件-1 (LINE1)和小鼠内源性逆转录病毒- l (MERVL)。
染色质状态在整个胚胎发生过程中动态变化。然而,由于早期胚胎的高度异质性和有限的细胞数量,在单细胞分辨率下研究染色质可及性提出了明显的挑战。候选顺式调控元件(cCREs)在谱系分离过程中被动态激活或沉默的时间和机制仍有待阐明。最近的一项研究利用外源性甲基转移酶检测染色质可及性来绘制小鼠早期胚胎的单细胞表观遗传图谱。然而,从转座酶可及染色质测序(ATAC-seq)技术分析中获得的单细胞分辨率染色质可及性图谱仍然缺乏,该图谱具有相对较高的信噪比,有利于大规模的单细胞研究。除了由DNA甲基化介导的典型基因组印迹外,小鼠植入前胚胎中的一部分基因表现出父系偏倚的染色质可及性,这可归因于母系遗传的抑制标记,组蛋白H3赖氨酸27三甲基化(H3K27me3),独立于DNA甲基化。尽管大多数由母体特异性H3K27me3标记的非典型印记在植入后丢失。然而,在正常发育的植入前胚胎的不同细胞系中驱动非典型印迹基因的机制尚不完全清楚。
单细胞ATAC-seq (scATAC-seq)已被广泛用于分析开放染色质可及性和cCREs。目前的scATAC-seq方法通常需要Tn5转位步骤的初始输入大约5000到100,000个细胞。在基于bulk的ATAC中,已经努力将每次转位反应的最小输入量从超过200减少到大约20个细胞。然而,在单细胞分辨率下研究哺乳动物着床前胚胎的染色质可及性仍然是一个巨大的挑战。作者此前开发了一种基于单分子测序(SMS,也称为第三代测序)平台的scATAC-seq方法,scNanoATAC-seq。在这项工作中,我们介绍了scNanoATACseq2这种基于长读序列的染色质可及性方法,用于分析含有单个细胞的珍贵和稀缺样品。作者将scNanoATAC-seq2应用于小鼠着床前胚胎,以表征早期胚胎发育过程中驱动ZGA和谱系分化的表观遗传调控机制。
三、主要结果
1、scNanoATAC-seq2方法的构建及其在小鼠着床前胚胎分析中的应用
作者开发了一种scATAC-seq方法,scNanoATAC-seq2,能够用于分析稀缺样品(甚至只有一个细胞的样本)。采用单管策略进行单细胞分离、细胞核提取、地黄苷渗透和Tn5转位等步骤,以尽量减少样品损失,并在Oxford Nanopore Technologies平台上进行长读测序,以产生染色质可及性数据(Fig.1A)。在人类和小鼠细胞系的测试结果显示出极小的交叉污染与细胞系区分能力。作者共获得包括受精卵、早期和晚期2细胞、4细胞、8细胞、16细胞的着床前scNanoATAC-seq2的数据,共包含3302个单细胞的染色质可及性图谱、fragment的中位数为23160个,片段中位长度为5486个碱基对。值得注意的是,该数据的线粒体DNA片段仅占比0.65%。该数据里共包括11种不同的细胞类型与3种细胞系:547个受精卵、278个早期二细胞期细胞、205个晚期二细胞期细胞、193个四细胞期细胞、387个八细胞期细胞、265个桑葚期细胞、303个ICM细胞、325个早期TE细胞、221个EPI细胞、178个PE细胞和263个晚期TE细胞。数据里共检测到240,494个染色质可及性峰,其中启动子峰占总峰的8~24%,随着发育进程,启动子峰的数量有所增加,表面活性染色质从远端向近端转移。轨迹分析结合UMAP与tSNE反映了这些细胞类型的发育特征(Fig.1C&D),拟时间分布与实际的胚胎发育阶段相对应(Fig.1E&F)。
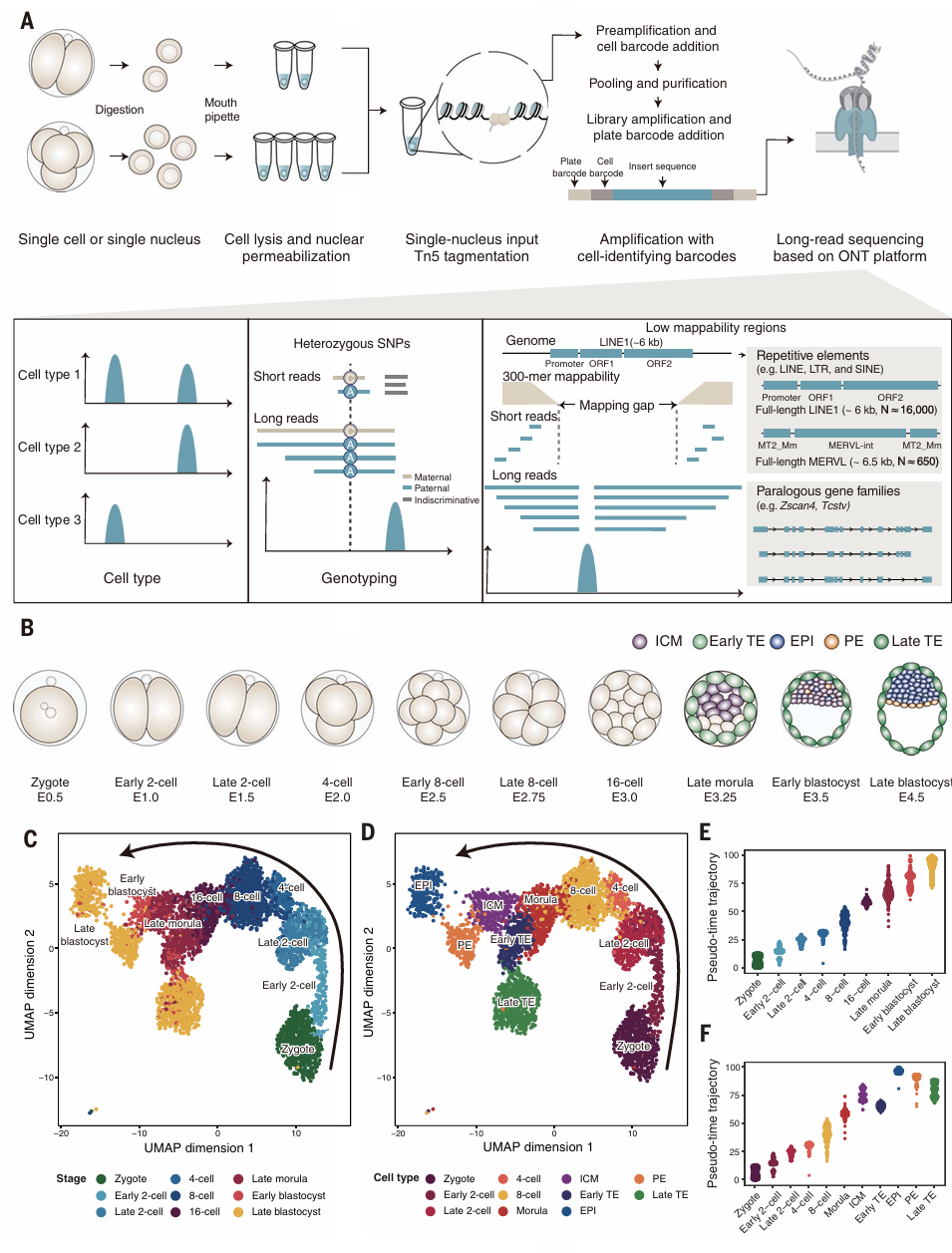
Figure 1
2、在minor ZGA与major ZGA中染色质可及性的变化
在卵裂阶段,胚胎迅速重编程并启动ZGA,Obox6在此期间被激活(Fig.2A&B)。在早期2细胞阶段,Obox6启动子附近的cCREs开放,这表明未检测到Obox6潜在增强子。接下来作者评估了ZGA相关转录因子的活性,包括Obox5、Obox6、Zscan4集群基因等。这些转录因子的染色质可及性模式与其结合位点从受精卵到两细胞晚期的可及性相关(Fig.2B)。这意味着它们在ZGA基因表达的初始阶段中重塑染色质可及性的潜在作用。
染色质可及性与转录组通常变化一致,但由于母体RNA的降解,受精卵到2细胞期的RNA表达模式下调。通过将scNanoATACseq2数据与已发表的卓创前胚胎数据集相结合,表明染色质开放和RNA表达之间的时间滞后很小,比相应基因的RNA表达变化早几小时发生。在ZGA中,Kdm5b、Obo6、Craos、Testu3展现出不同的时间滞后模式(Fig.2C-D)。z转录抑制与正常的晚期2细胞胚胎形成鲜明对比,胚胎表现出与受精卵相似的染色质可及性模式,并在5269个基因位点上染色质可及性较低(Fig.2F)。
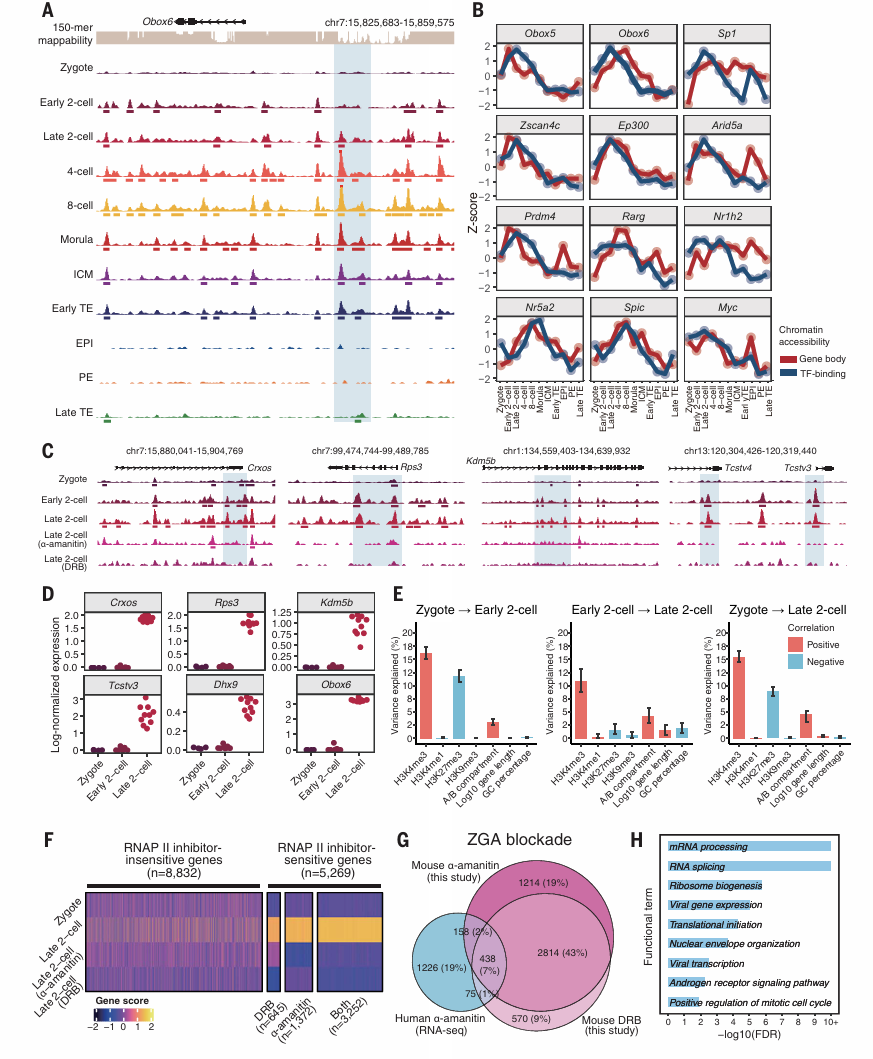
Figure 2
3、细胞系分化中染色质可及性的调控
为验证scNanoATACseq2捕获的染色质可及性的细胞类型特异性差异,作者确定了受精卵到囊胚阶段的染色质可及性和表达变化差异一致的基因,其中Sox2、Lgr4、Gdf3和Bmp4在ICM中特异性上调。相反,在TE中,Oct4、Oboz6、Klf2、Klf4、Wdr5、Sp110、Spic和aeal的染色质可及性降低。Jakl、Krt8、Krt18、Tfap2a、Klf6及Gata2在TE中表现出染色质可及性和RNA表达的增加(Fig.3A)。在EPI和PE的谱系分离过程中,Sox2、Fgf4、Nanog、Fgfr1、Epcam在PE中表达下调,而在EPI中表达稳定。相比之下,Gata6和Pdgfra在EPI中下调,而在PE中稳定(Fig.3B)。Nanog是EPI发育最早激活的转录因子之一,与PE相比,EPI中的染色质更开放(Fig.3C),其启动子附近的几个峰为Nanog裂解阶段的cCREs。作为TE标记基因,Gata3在桑葚胚和TE早期、晚期表现出开放的染色质特征,并揭示其下游的几个cCREs。TE谱系中GATA3下游靶位点的染色质可及性也特异性增加(Fig.3E)。
作者进一步揭示了每个TF基因位点的染色质可及性及其结合位点活性在细胞类型之间的同步。EPI中包括SOX2、NANOG和KLF2;PE中的SOX17、HNFIB、GATA4、GATA6、TEAD4、GATA3、CDX2和ELF3在TE中的表达(Fig.3E)。Nanog的基因score升高在在TF结合score之前,即表观中TF的结合比下游靶基因位点染色质打开更早。
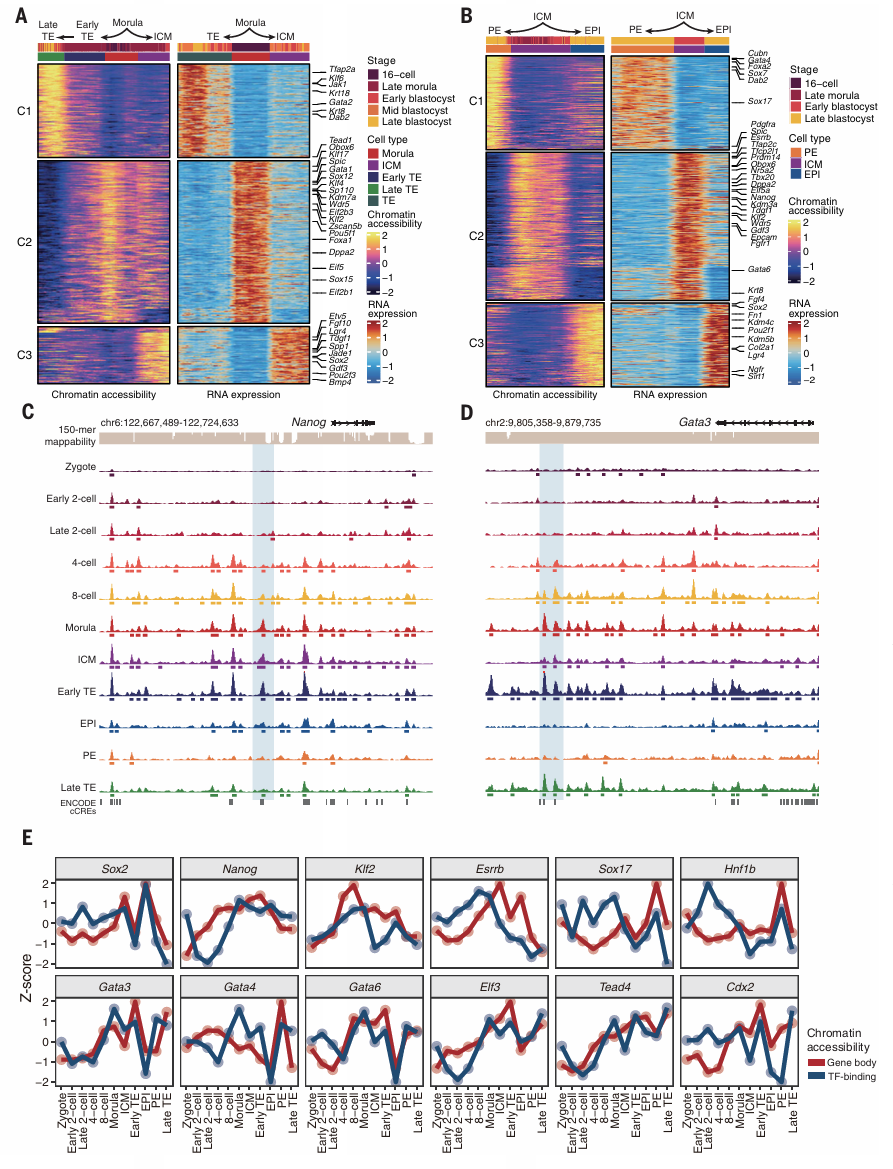
Figure 3
4、非典型印记技能的等位基因特异性染色质可及性
为了阐明早期胚胎中母本和父本等位基因印迹的不对称调控,作者鉴定了C57xDBA杂交不同发育阶段的等位基因特异性和可及基因(ASGs)。通过比较每个细胞类型的等位基因之间的染色质可及性来确定ASGs。与桑葚胚和早期相比,在囊胚阶段,包括ICM、早期TE、EPI、PE和晚期TE,所有三个谱系的ASG数量都明显减少(Fig.4A)。在卵裂阶段,副本特异性ASGs的数量远超母本特异性ASGs(Fig.4B)。与此前的DNA-seq研究一致。
为验证这一发现,作者进行了DBAxC57、C57xCAST的反向杂交,在三种遗传背景下,共鉴定出325个非典型印迹基因,包括317个父系ASG和8个母系ASG(Fig.4 C&E)。这些ASG展现出父系优势模式。这些例子展示了印迹强度和等位基因失衡的统计意义。等位基因标记率中位数为96.8%(Fig.4D)。这些结果证明了scNanoATAC-seq2在检测ASGs中的有效性。
Xm和Xp之间的染色质可及性的偏好性反映了iXCI,并从四细胞期开始逐渐增加。到桑葚阶段,这一印迹在EPI与ICM相比降低(Fig.4H),晚期囊胚EPI中iXCL的清除仍不完全,iXCI强度仍高于四细胞胚胎(Fig.4H)。
从受精卵到8小阶段,雌性胚胎在Xist结构域表现出父本特异性的染色质可及性,而Tsix结构与在这两个亲本等位基因之间表现出平衡的染色质可及性(Fig.4J)。在谱系分离到ICM和早期TE时,Xist结构域的等位基因不平衡减少,而Tsix结构域开始表现出母体特有的开放染色质特征(Fig.4J)。
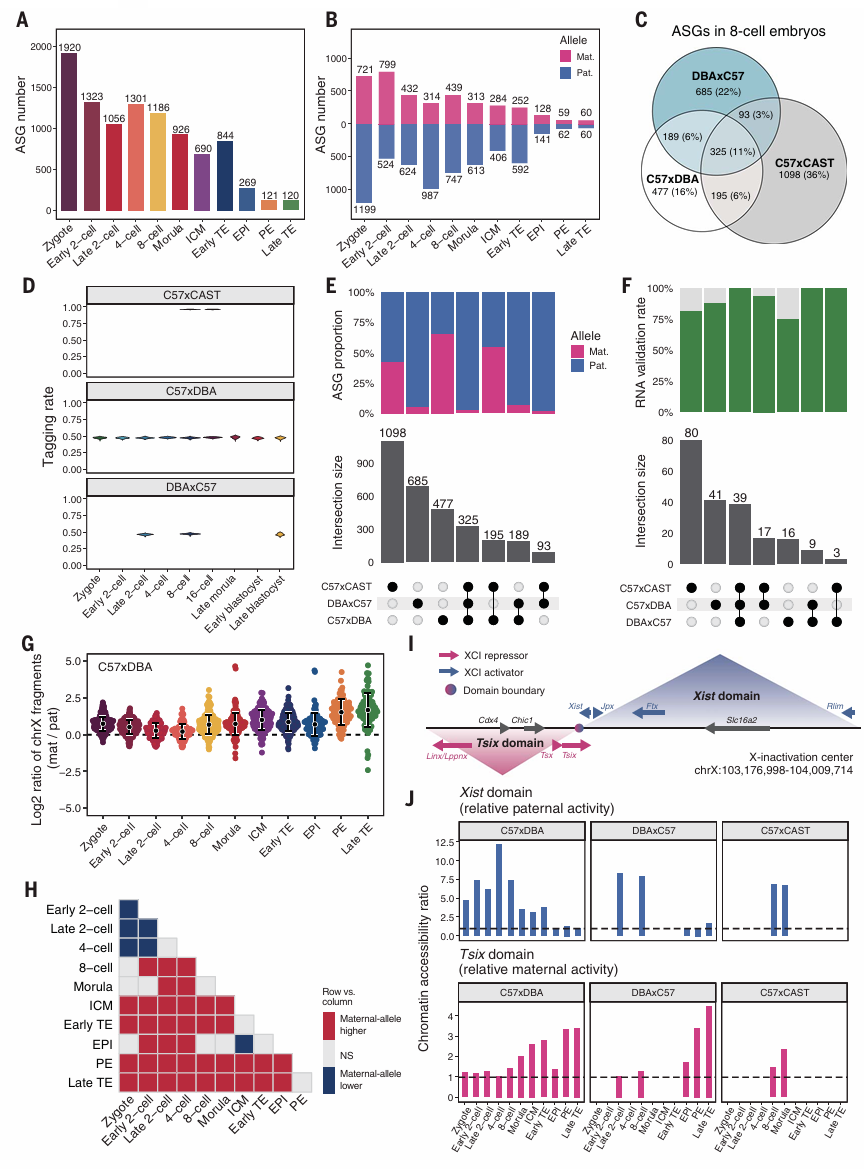
Figure 4
5、小鼠着床前发育过程中重复元件染色质的可及性动态
重复DNA序列约占小鼠基因组的45%,最近的研究表明,它们通过顺式和跨调节机制在ZGA和细胞命运中发挥关键作用。但短读长测序方法在识别重复元件的单个拷贝方面存在困难,长读长测序方法一定程度上克服了这一困难,能够捕获重复元素的单个拷贝及它们明显可比对的侧翼区域信息(Fig. 1A)。长时间分散的核元件(LINEs)与长末端重复序列发挥独特的染色质可及性动力学特征。LINE1元件的染色质可及性从受精卵到双细胞的数量急剧下降,而在桑葚期的EPI、PE、TE中逐渐增加(Fig.5A)。在当前研究中,LINE1元件被表观遗传抑制,只有一小部分被显著激活(Fig.5B)。在八个细胞阶段的数据中,展现出了异质性的染色质可及性(Fig.5E)。作为一类内源性逆转录病毒(erv)的重复元件,LTR元件在发育过程中表现出染色质可及性的异质性和动态变化(Fig.5A)。MERV元件在2细胞胚胎中作为zg相关基因的顺式调控激活因子被强烈激活。MERVL元件的平均可及性在2细胞后期达到峰值,逐渐降低,最终在囊胚后期小时。利用长独长测序,作者发现在晚期2细胞胚胎中,完整MERVL转座子两端的MT2-Mm LTR元件显示出明显的开放染色质特征,而内部序列显示出低染色质可及性。MT2-Mm元素上开放染色质的富集在短读长ATAC-seq数据中不太明显(Fig.5F)。
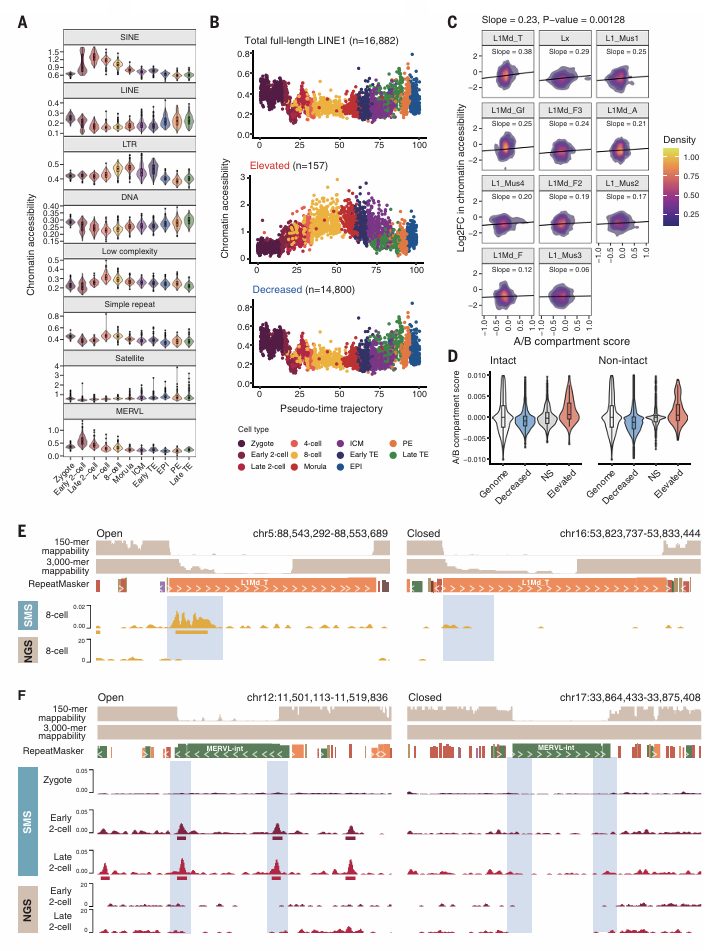
Figure 5
6、2细胞胚胎晚期的同源基因染色质可及性特征
在人类与小鼠的ZGA时期RNA表达量会上升,而Fam172a、Kifc3、Neo1这类接近MERVL的基因没有上调。Zscan4同源基因是2细胞胚胎和2细胞胚胎样ESCs的自我更新标记,在ZGA中起到关键作用。作者对Zscan4相似的全长基因进行整理并聚类(Fig.6A),展示了它们在顺式调控中逐渐变化的模式与表达水平(Fig.6B)。在2细胞晚期,与ZGA相对应,scNanoATAC-seq2清除的描述了每个Zscan4平行序列的内含子区域中关键CRE的激活(Fig.6C)。而短读长ATAC-seq数据未检出这一激活(Fig.6D)。对于Zscan4b和Zscan4e下游LTR元件中检测到的染色质开放特征在短读长ATAC-seq中很大程度上缺失。再次展现出了长读长测序的优势。
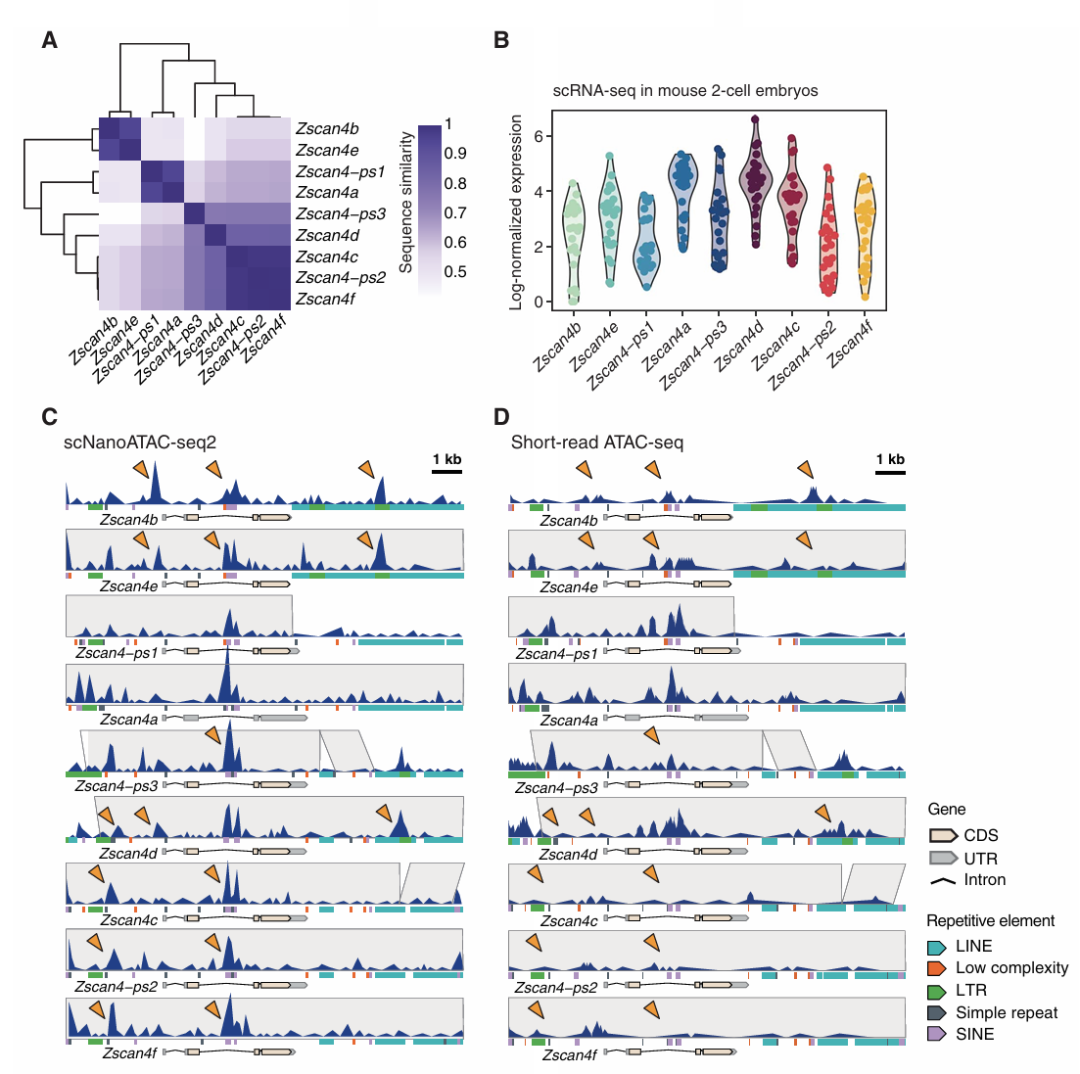
Figure 6
四、讨论
可能很多同学和我一样,硬着头皮看到这里并不是对胚胎发育感兴趣,而是被scNanoATAC-seq2所吸引。scRNA-seq的问世已经十余年,整个行业仍面临巨大挑战。高精尖的技术能否走向转化?单细胞组学的热度是否会冷却?这都是产学研各界人士的担忧。在这一背景下,汤老师执着于三代测序+单细胞组学的应用,解决了重复序列、等位基因检测、低检出度基因检测等问题。
当前,scRNA-Seq的测序过程主要依托于二代测序(Next generation sequencing,NGS),这相较于一代的桑格测序大大的提升了通量,让"组学"的测序成为了可能(不熟悉高通量测序的同学可以参考这一篇:测序技术的发展与应用)。但是,读长短也一直是NGS的硬伤。一方面,真核生物的一个基因通常编码8~9个mRNA、生成3~4种蛋白;虽然NGS能够通过拼接完成剪切体的预测,但检测能力与说服力仍有不足。另一方面,对于人类样本来说,NGS也无法充分考虑到基因组拥有两个拷贝的情况。并且,NGS过度依赖PCR扩增检出信号,扩增的bias也是NGS的痛点。近年来,三代测序的出现为这些NGS的短处带来了改观。作为scRNA-Seq的开创者,汤老师目前正在利用三代测序开展单细胞测序的相关工作,并获得了大量成果。经验证,基于三代测序的单细胞测序在基因捕获数量(2~300 genes/cell → 8000 genes/cell)pseudo gene的检出(~1400个)、BCR与TCR的VDJ重排、scATAC-Seq质量(3~5w个reads/cell、200 cells/细胞类型即可满足分析要求)、重复序列检测、环形DNA检出(用于单细胞基因组,100~200 cells即可完成基因组拼接)、序列染色体定位等方面具有较强的优势。也让我们更加地期待单细胞组学与测序技术的未来!


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









