








电荷分布是核心
分区区域:蛋白质IDR区域上的电荷分布
功能:选择性分区(action),在IDR区域凝聚物中——》导致gene活化《——这种选择性分区是由电荷分布模式所介导的
QA:
**Q:**转录调控机制涉及到很多蛋白质/DNA/RNA大分子组分,这些组分在相分离中通常分散在不同的、特定的凝聚物中,也就是分散到特定的相中,这种分散的特异性是由蛋白质的无序(区域)所介导的,问题就是蛋白质
how(如何)介导这种相分离的特异性(即什么分子聚集在一起)?
转录所需的多种成分是如何选择性地浓缩(形成凝集物),普遍认为弱多价相互作用驱动生物凝集物的组装——》但是组装的特异性,或者说这种生物大分子油相的特异性,是如何通过弱多价相互作用形成的?
蛋白质如何分选成不同凝集物的一般原理
A:
分区蛋白质的idr对于选择性区室化是必要和充分的,并需要带电氨基酸的交替块
重点在于IDR+可变电荷块
破坏这种电荷模式会阻止分离,而将这种模式添加到蛋白质中会促进分离,从而对基因激活产生功能性影响。具有相似图案电荷块的idr具有相似的分区和功能。
——》无序介导的相互作用可以选择性地从复杂的生物分子混合物中分离出特定的功能相关蛋白,从而调节生化途径
作用的核心是IDR,作用的机制是IDR上的可变电荷快:
选择性分配是由这些idr中的带电残基形成交替的电荷块所驱动的
操纵idr的电荷模式或交换idr可以重新定位cell中的分区,从而导致功能的变化
例子:
由MED1的内在无序区(IDR)组成的凝聚体选择性地分割RNA聚合酶II及其正变构调节因子,而排除负变构调节因子;
这种选择性区室化足以激活转录,并且是细胞状态转换期间基因激活所必需的;
虽然我们确定了转录途径的特定特征模式,但很可能idr在区室化中发挥作用的其他生化途径将进化出自己的一套特征模式,以促进蛋白质分选到各种细胞凝集物中
Extend:
通过IDR介导的弱多价相互作用,所形成/驱动的相分离/区室化,的其他生化途径(上面例子是讲Med1通过IDR的可变电荷块调控转录),
这些生化途径中可能观察到其他的IDR的弱多价互作模式,然后可以促进基因表达调控过程中的生物大分子选择性分选(选择性凝聚)
疏水性——》也有序列预测
一,result
1,result1:MED1(IDR)区域所介导(形成)的凝聚物能够选择性地将转录的正调节因子分选在一起(凝聚在一起)
目的:研究idr介导的凝集物的选择性分配
IDR用的也是预测的工具,我用fiDPNN一样
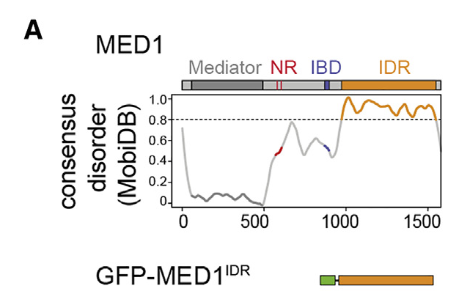

而且很奇怪的是它居然就只把这一大段IDR区域单独提取出来做研究了(当然,这一段IDR区域太大了)
——其实我也可以看一下哪些ZF的IDR非常大(看fiDPNN图)
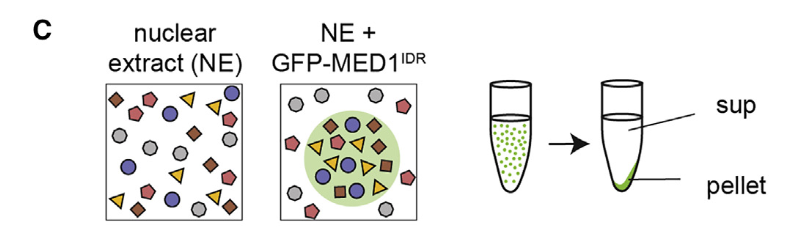
也就是说这个蛋白质(含有IDR为主导的多肽),就像1个海绵,扔进核提取液里就吸水,就会立刻分选招募吸收周围的一些蛋白质(当然是选择性的),然后立马就析出了(油水相分离)
然后就是对颗粒以及上清液两个相里的蛋白质进行了蛋白质组学分析(其实就是鉴定分别两相中有什么蛋白质),然后定义了一个分离比(partition ratio),就是分配在下面的相里的,比上分配在上面的相里的的蛋白质浓度比
By proteomic analysis of pellet and supernatant fractions, we calculated a partition ratio, defined as pellet over supernatant (pellet/sup) for every identified protein in the extract
然后就是用底部富集的蛋白做了一个富集分析,实际上是GO

看看是哪些蛋白被选择性地富集在底部:大都是RNA polII转录调控过程中富集到的正向调节因子
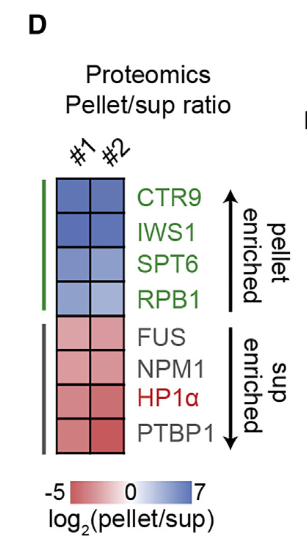
其实我感觉这种图本质上和DEG的火山图没什么区别,就是两个极端的富集方向,比如富集在上清中,或者富集在上调中,每种蛋白质或gene有富集的分配比分离比,对应gene的logFC;
然后正好两个方向上的蛋白质都可以做一个富集分析;
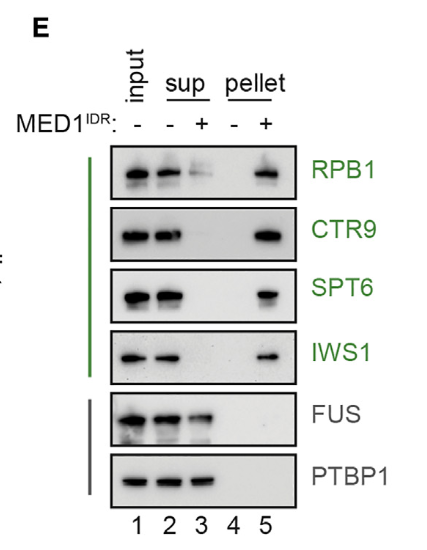
然后做了一个蛋白质的免疫印迹分析,主要是为了确保颗粒确实是MED1所引起的,就做了一个对照,主要是看看凝聚物离心分离前后的组分,在这个过程中进行对照:
就是没加这个IDR蛋白质的,然后看看在加没加前后,离心比对前后确实有区别,
就是因为加了IDR这段蛋白,所以右上角那4个绿色的蛋白都被凝聚到底部pellet了(也就是说体外的实验中,这些特定的转录/正向调节因子确实是被MED1-IDR选择性的分选到凝聚物中);
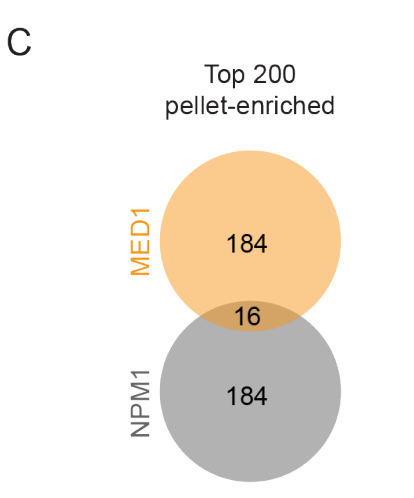
然后首先就是分析这些蛋白质是否是MED1-IDR所特异富集分选起来的,和另外前人所做的NPM1研究(也是蛋白质凝聚物分离实验)做了个对比,
发现是两个实验中所富集到的蛋白质基本上都是特殊的,就是特异性富集的,所以我们这里实验所富集到的蛋白质肯定是MED1-IDR所富集的,而且是特异性富集的(虽然说这个结论不是很可靠,理论上肯定要和其他蛋白质分选都做1个对比)
然后就是发现MED1-IDR凝聚物对RNA polII正向调节因子的的富集倾向是强于NELF等因子(之所以选择NELF等因子作为比对,是因为NELF蛋白调节因子作为前面所正向富集的蛋白质实际上与RNA polII的互作是互斥性的;
RNA Polⅱ与CTR9和SPT6或与NELF复合体的相互作用是互排性的,每种相互作用都提供了不同的构象,导致RNA Polⅱ处于活性或非活性状态。
因为是用MED1的IDR区域去捕获的,所以后面需要分析MED1蛋白和所富集的蛋白质的关系,而且反正都要做体内试验,也就是内源性试验,所以需要分析,ED1是否与细胞中这些选择性分选的蛋白质所共定位;
然后就做了一个co-IP,验证了在内源性蛋白浓度下,MED1焦点与RPB1、CTR9和SPT6共定位;
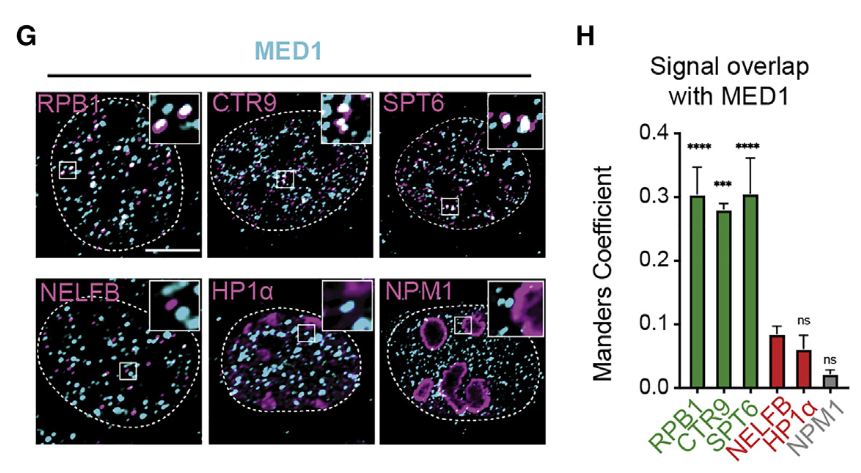
总体而言,我们是能够提出这么一个由MED1-IDR所介导的相分离的凝聚物的(凝聚物的分布图、组分图)——》这个组分中大部分是pol II的正向调节因子
2,result2:MED1在增强子上的高水平占用与RPB1、CTR9、SPT6和相关基因的转录增加相关
其实这个结论不用看都知道,光从结论1上面讲,如果MED1能介导与正向转录调节相关的蛋白质的凝聚体,那么在增强子上发现高水平的MED1蛋白,也基本上就能将逻辑引导到高水平的这些蛋白质当中去,以及正向转录调节随之而来的就是正向转录(转录增加);
应该是对增强子的一个增强转录机制提供一个解释,一个相分离的解释(就是why enhancer enhance,因为聚集招募了高水平的MED1,然后借由MED1的IDR招募了高水平的正向调控因子所组装富集而成的凝聚体,然后就可以解释why 增强转录了)。
分析了已经发表的公共数据库中的ChIP-seq数据,发现MED1占用率(结合率)最大的增强子(其实就是SE)附近的gene,其被那些正向调节蛋白结合比例很高;
因为我们做ROSE,基本上就是将SE邻近的gene作为靶基因,所以上面的意思就是:
越是SE,MED1结合越富集,它所调节的靶基因也就会富集更高水平的这些正向调节蛋白。
下面的这些图意思就很简单:
我有这4种蛋白质的ChIP-seq数据,然后我自然会得到每种蛋白质的结合peak;
对于这些peak,我手头上又有SE的注释数据(应该就是bed文件,其实就是MED1占据信号强弱的一个注释bed);
那我就可以将增强子分为两类SE以及TE(其实就是依据的MED1 信号),然后我分别取寻找这两类E的靶基因,
那就是对应着高MED1占据的SE的靶基因,以及低MED1占据的TE的靶基因,然后有了生物学意义的分组之后,我就可以去看一下这个分组中两个不同的靶gene集各自的这4种蛋白质的ChIP-seq信号
——》就说明MED1占据水平有差异的E,其所调控的靶基因,在这4种蛋白质占据水平上也有差异;
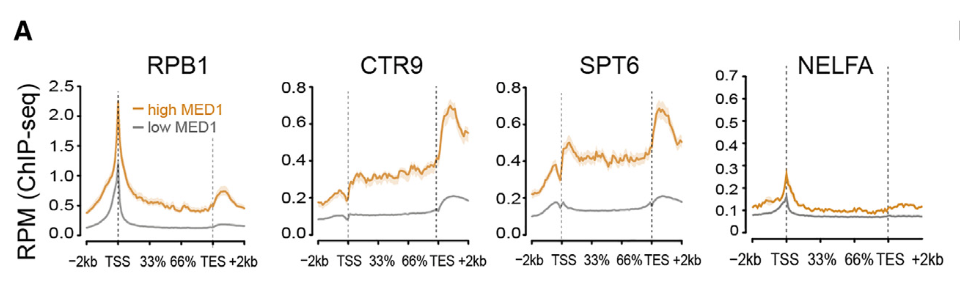
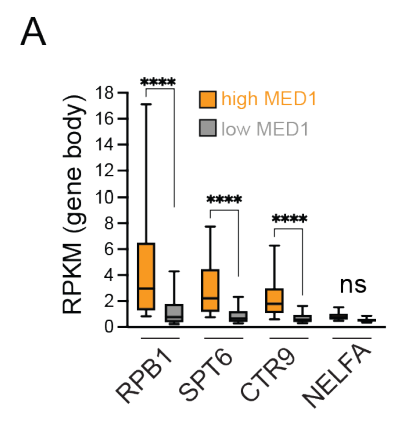
做了loess曲线拟合:
在增强子中 MED1 结合度最高的基因,其基因体中的 RPB1、CTR9 和 SPT6 结合率也最高,存在一个明显的 MED1 阈值
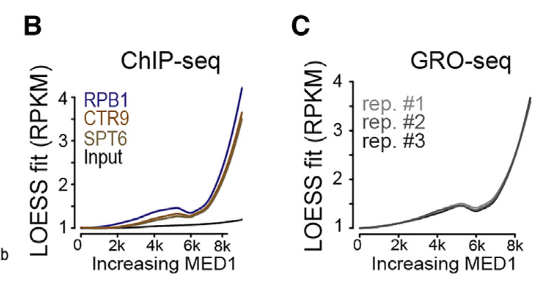
然后结论就是
MED1占用率最高的基因组位点(比如说那些enhancer)被(鉴定为MED1-IDR成分的分区蛋白)所凝集;
说白话就是超级增强子所调控的靶gene,富集那些被MED1-IDR招募的相中的正向调控蛋白,而TE所调控的靶gene,富集就少了;
接下去就是探究为什么这些地方会富集正向调控因子,
转录正调节因子占用率的增加是否与RNA Polⅱ活性和转录率的增加相关?
我们要研究的作用因子var是RNA pol ii活性,而该活性与其S5或S2位点的磷酸化相关:

参考文献:https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkae1240/7932184?login=false
这里对于RNA polii的活性有个CTD code(也就是特定位点的PTM等修饰,与该酶的特定活性相关联对应)

所以SE调控的靶gene上,去捕获的RNA polii的ChIP-seq,上面的S5或S2磷酸化水平就比TE高(B~D);然后活性方面除了RNA polii特定位点的磷酸化之外,还有一个事件——就是启动子近端暂停(promoter-proximal pausing),可以通过一个指标进行评估:traveling ratio,该指标越低就表明RNA polii越不容易在近端暂停(大概意思就是转录活性会高一点,转录会继续延伸之类)——》图E
所以两个指标,一个是特定位点的磷酸化code,另外一个是判断RNA polii是否在启动子近端暂停,来评估SE以及TE调控的gene上的RNA polii活性;
然后SE调控的gene上的RNA polii活性高,TE的反而低
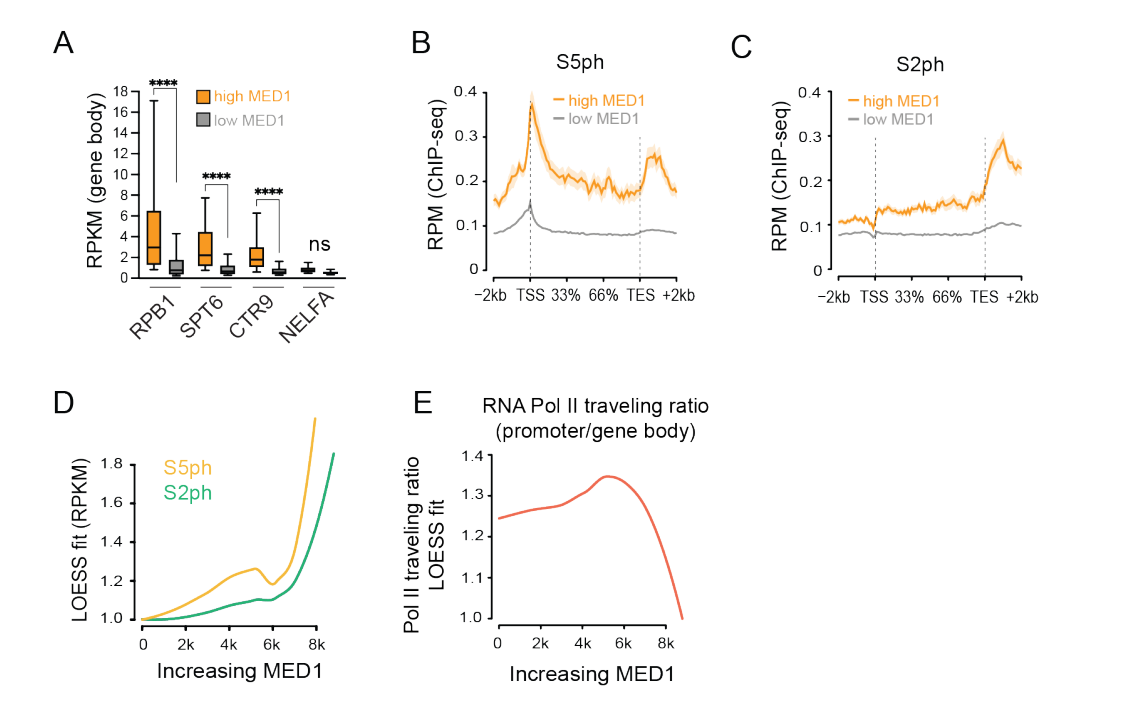
总结来说就是正调节因子占用率高、RNA polii低暂停
与正调节子占用率较高和暂停较低一致,增强子MED1占用率最高的基因具有较高的转录率(见GRO-seq)
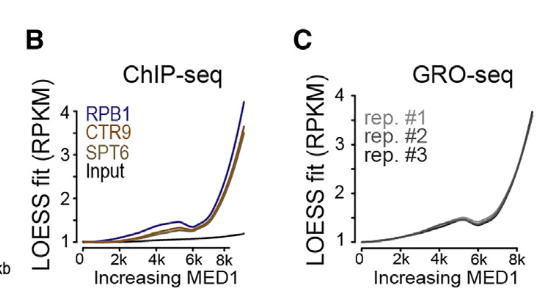
然后结论就是高局部浓度的MED1将RNA Polⅱ及其正变构调节因子分选(在一起),导致具有延伸能力的RNA Polⅱ的组装频率较高,因此转录速率较高——》还是从相分离之后,同一相中的组分能够促进RNA polii的活性来说的
3,result3:高局部浓度的MED1-IDR足以在细胞中分选正调节因子并激活基因转录
光看标题上,是和上一个结论相关联的,上一个劲结论我们知道了高局部浓度的MED1-IDR,与RNA polii+正变构调节因子的分选相关(这里是相关关系),然后RNA polii活性和转录是因果关系
——》但是我们不知道MED1-IDR与这个分选的因果关系
因为前面都是在体外的核提取物里做的,我们需要知道体内/cell 内,在基因组位点上结论如何。
实际上就是充分性验证:
充分性:MED1 IDR会选择性吸收正调控因子吗?
弄了一个人工定位体系(LacO-Lacl),
首先LacO和Lacl的互作是特异性的,然后LacO阵列是已经集成到U2OS 2-6-3这个细胞中了;
LacI与lacO的结合是为了招募CFP-LacI融合蛋白到特定的DNA位点,确保该融合蛋白能够在细胞中形成荧光焦点。当CFP蛋白接近激发光源并被激发时,会发光;
所以我们只是为了将IDR融合蛋白招募到细胞基因组中特定位点罢了,(lacl和laco是特异性定位用的,不用管;CFP是为了可视化的,也不用管)
整个实验其实就一个有MED1-IDR,一个没有,作为对照看看这个CFP发荧光的地方,如果用下面这些正、反(NELFB)去做IF实验,会不会得到聚集效果;
——》其实就是模拟的MED1-IDR在体内定位的情况下,是否能够观察到选择性蛋白富集的效果
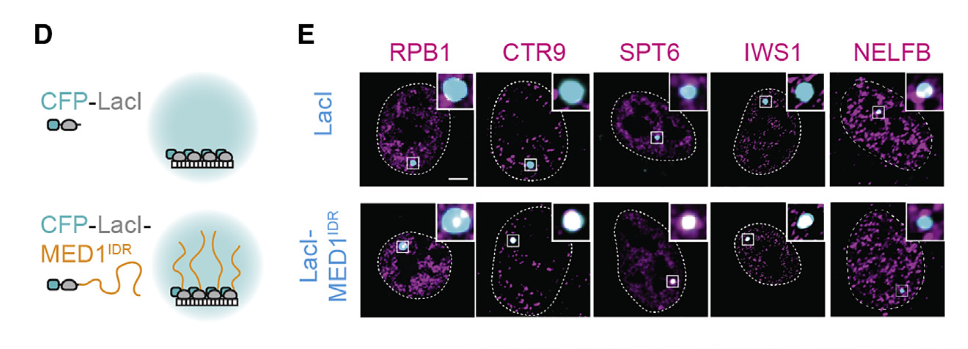
在该实验中,CFP(氰色荧光蛋白)主要作为荧光标记,用于可视化融合蛋白的位置。具体来说,研究人员将CFP与LacI或LacI-MED1IDR融合,转染到含有集成LacO阵列的U2OS 2-6-3细胞中。通过荧光显微镜,CFP发出的荧光信号能够明确显示出融合蛋白在细胞内的定位,从而帮助研究人员观察和比较不同处理组中的蛋白聚集情况。
在这些实验中通常会有两类荧光信号:
-
用于定位融合蛋白(如CFP-LacI或CFP-LacI-MED1⁽ᴵᴰᴿ⁾)的荧光,这一信号用来识别融合蛋白在细胞核内富集的位置;
-
针对感兴趣因子(例如转录因子、组蛋白修饰等)的免疫荧光(IF)信号,用不同的荧光染色来检测它们在所定位区域的富集程度 .
在对照组(CFP-LacI)中,LacI只是将CFP锚定到LacO阵列上。你会看到一个CFP的焦点,但并不会显著富集转录正调控因子或活化染色质标记,主要用来对比观察:
• 如果没有MED1 IDR的存在,目标因子是否也会自然聚集在此?
• 由此可排除非特异性结合或其他实验过程导致的假阳性 .
在实验组(CFP-LacI-MED1 IDR)中,除了CFP-LacI用于锚定之外,MED1的无序区(IDR)会将特定的转录正调控因子富集到该位点上,相应的免疫荧光染色会在同一位置出现加强信号:
• 若看到明显的转录因子(如RPB1、CTR9、SPT6等)或活化标记(如H3K27ac、RNAP II磷酸化修饰)信号增强,说明MED1 IDR能够招募并激活该基因组区域 .
• 若看到NELFB等负调控因子信号未富集或相对降低,则进一步证明了这种选择性聚集效应 .
因此,通过比较这两类荧光信号(锚定蛋白的CFP和目标因子的免疫荧光),可以判断MED1 IDR对于凝聚和招募转录正调控因子的贡献。对照实验的逻辑在于排除其他干扰因素,而实验组的观测则验证高浓度MED1 IDR是否足以在指定基因组位点富集相关因子并激活转录 .
总而言之:
在上述实验设计中,lacO(乳糖操纵子)和LacI(乳糖操纵子结合蛋白)起到了关键的工具作用,用于特定基因座的定向蛋白招募和功能研究。以下是对实验设计思路及这两者作用的详细理解:
- 建立特定基因座(Lac array cells):
- U2OS 2-6-3细胞系中整合了一个LacO阵列(Lac array),即在细胞基因组中引入多个乳糖操纵子序列。这些LacO序列作为特定的结合位点,用于招募融合蛋白,从而在细胞核内形成可视化的荧光焦点。
- 融合蛋白的构建与表达:
- 实验中,研究人员转染细胞表达两种融合蛋白:CFP-LacI 和 CFP-LacI-MED1IDR。
- CFP-LacI:将荧光蛋白(CFP)与LacI蛋白融合,用于作为对照,确保LacI能够特异性结合LacO序列并在荧光显微镜下形成可见的荧光焦点。
- CFP-LacI-MED1IDR:在LacI基础上进一步融合MED1的IDR(低复杂度富集区)域,目的是研究MED1IDR在特定基因座上对转录正调控因子的选择性分隔作用。
- 实验中,研究人员转染细胞表达两种融合蛋白:CFP-LacI 和 CFP-LacI-MED1IDR。
- 实验分析与结果观察:
- 免疫荧光(IF)检测:通过免疫荧光技术,检测转录正调控因子在CFP-LacI或CFP-LacI-MED1IDR焦点处的富集情况。
- 结果显示,在表达CFP-LacI-MED1IDR的细胞中,转录正调控因子在荧光焦点处显著富集,而对照组(仅CFP-LacI)则缺乏这种富集。
- 特别是,只有在CFP-LacI-MED1IDR焦点最亮的情况下,转录正调控因子才被观察到富集,表明MED1IDR在高浓度下具有选择性分隔的能力,且存在一个阈值效应。
- 免疫荧光(IF)检测:通过免疫荧光技术,检测转录正调控因子在CFP-LacI或CFP-LacI-MED1IDR焦点处的富集情况。
- lacO与LacI的作用总结:
- lacO提供了一个可被特定蛋白(如LacI或LacI融合蛋白)结合的DNA序列,作为定点招募平台。
- LacI作为结合剂,通过与lacO特异性结合,将融合蛋白(如CFP-LacI-MED1IDR)定向至该基因座,进而在特定位置高浓度表达目标蛋白域(MED1IDR),用于研究其在转录调控中的作用机制。
通过这种设计,能够在细胞内精确控制和观察MED1IDR在特定基因座上的功能,验证其在选择性分隔转录正调控因子中的作用
然后结论就是高浓度的MED1-IDR在一个确定的基因组位点再现了其在核提取物中观察到的选择性分配(因为确实是有选择性的,正向的IF荧光高,负向的NELFB就不高)
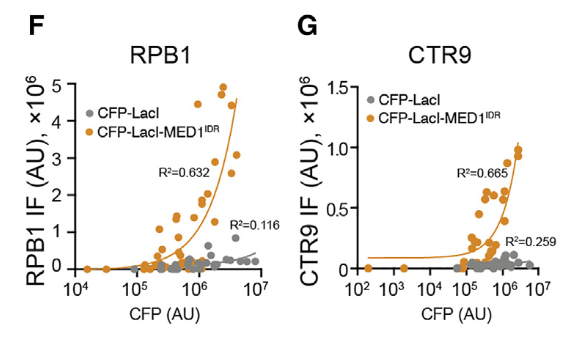
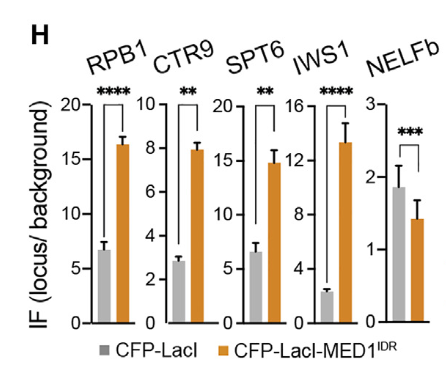
紧接着就是研究了局部高浓度的MED1IDR募集是否足以改变染色质格局并激活转录,其实就是研究在异染色质区域如果引进MED1-IDR,将会发生什么现象?
左边两个标记是异染色质标记,在lacl对照中是富集的,说明是异染色质,加入MED1-IDR之后就不再异染色质了;
右边3个(从H3K27ac开始)是活性染色质标记(以及RNA polii转录不暂停的活性信号),在lacl对照中是不富集的,但是加入MED1-IDR之后这个区域就活化了。
还有一个报告gene的可视化
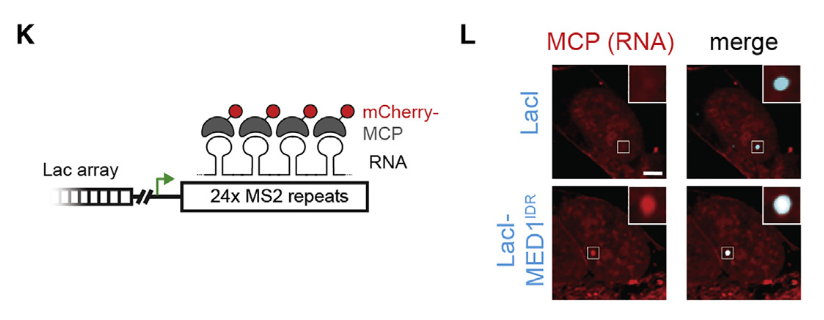
结论是:在特定的基因组位点上,高浓度的MED1IDR足以创造一个活跃的染色质环境。
4,result4:MED1IDR对于选择性分区是必需的
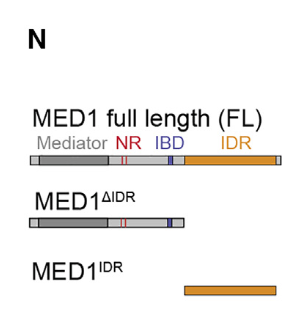
所以很自然的,对于我们的主要研究对象MED1-IDR,针对IDR要不要保留制作了3种变体:
全长、去掉IDR、只有IDR
少了IDR,分选效果基本上和对照组没有显著性差异,比其他2种差很多,基本上不能分选CTR9/SPT6蛋白
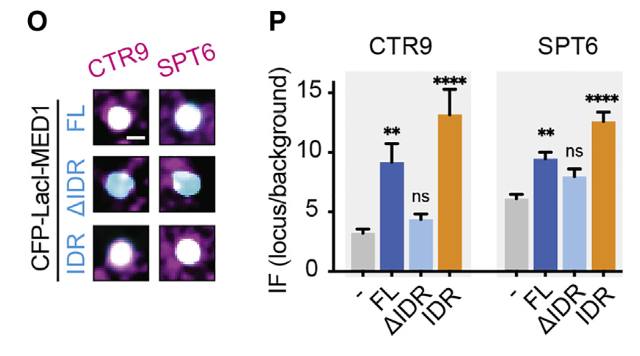
总之综合来看结论是:
高局部浓度的MED1-IDR能够选择性分选/富集RNA Polⅱ及其正调节因子,排除负调节因子,改变染色质环境,并激活基因转录
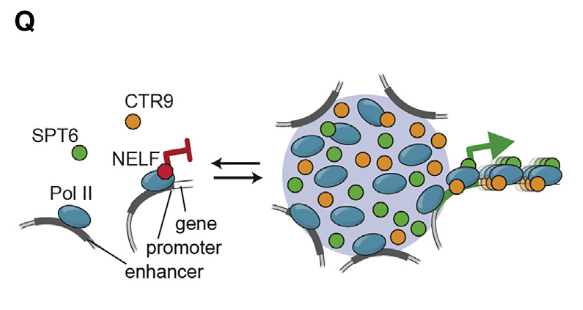
5,result5:MED1-IDR及其分区蛋白是细胞状态转换过程中基因激活所必需的
这一步其实就是验证MED1-IDR招募的相中的蛋白质的重要性,找一条生物体细胞内需要MED1的生物学通路来验证——这个result单纯就是验证MED1-IDR相分离在该通路中的作用,就是验证,可以跳过
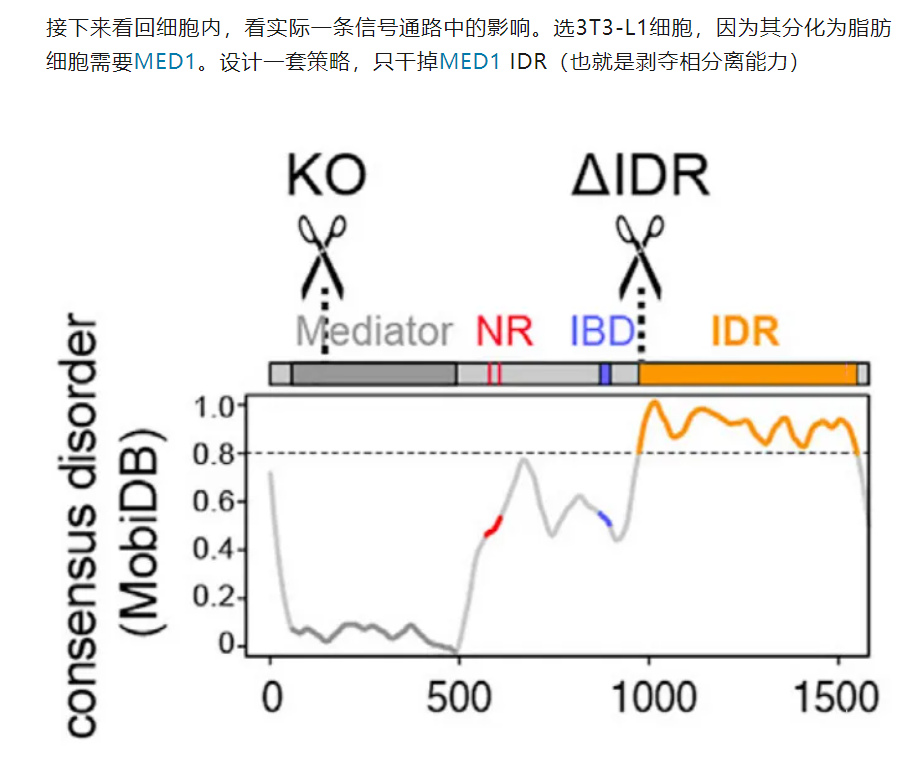
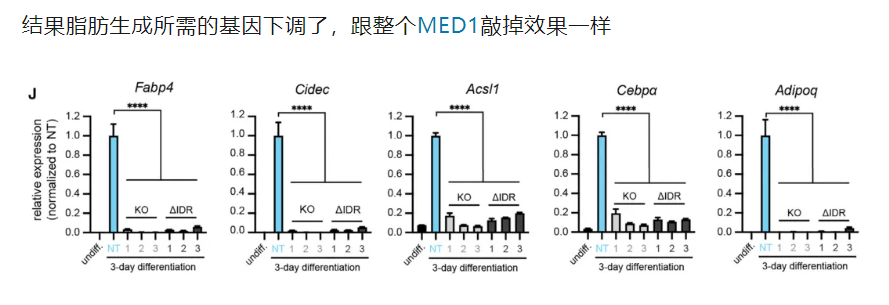
6,result6:IDR对于MED1这类分选因子而言是很重要的,也是实现相分离(选择性分区)充分且必要的
我们先来看看在MED1这个相中被招募过来的蛋白质:大部分都是有IDR的
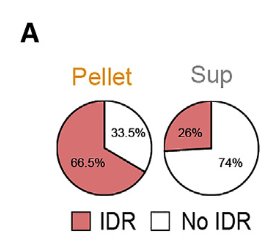
这促使我们研究IDR在选择性区室化到MED1IDR凝集物的充分性。
然后就是正负样本选择:
正样本为CTR9、SPT6
负样本选择NELFA、HP1α、FUS
纯化的部分是IDR:
纯化了CTR9和SPT6的末端IDRs,并将其作为重组mCherry融合蛋白(总之就是红色荧光蛋白标记那一套)
作为阴性对照,纯化了先前研究的NELFA、HP1a和FUS的IDRs(这些IDRs已知参与核凝聚体的形成,但在MED1IDR凝聚体中表现为中性或被排斥)
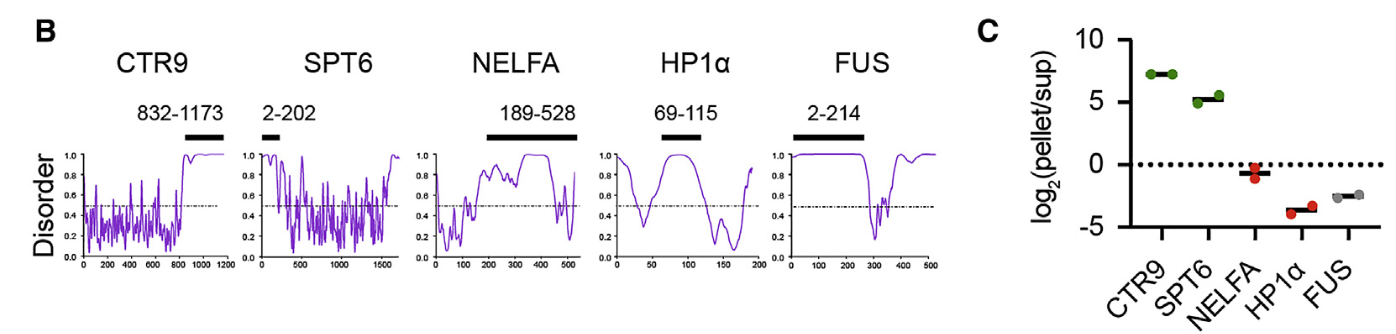
将上述mCherry-IDRs在体外测试其是否能分隔到GFP-MED1IDR液滴中,说白了就是测试仅凭客人的IDR能否被主人MED1邀请到到这个油相当中,结果与蛋白质组学分析中富集到的结果一致,只有正向的CTR9以及SPT6被招募进去了,然后在油相中发光被我们观察到了
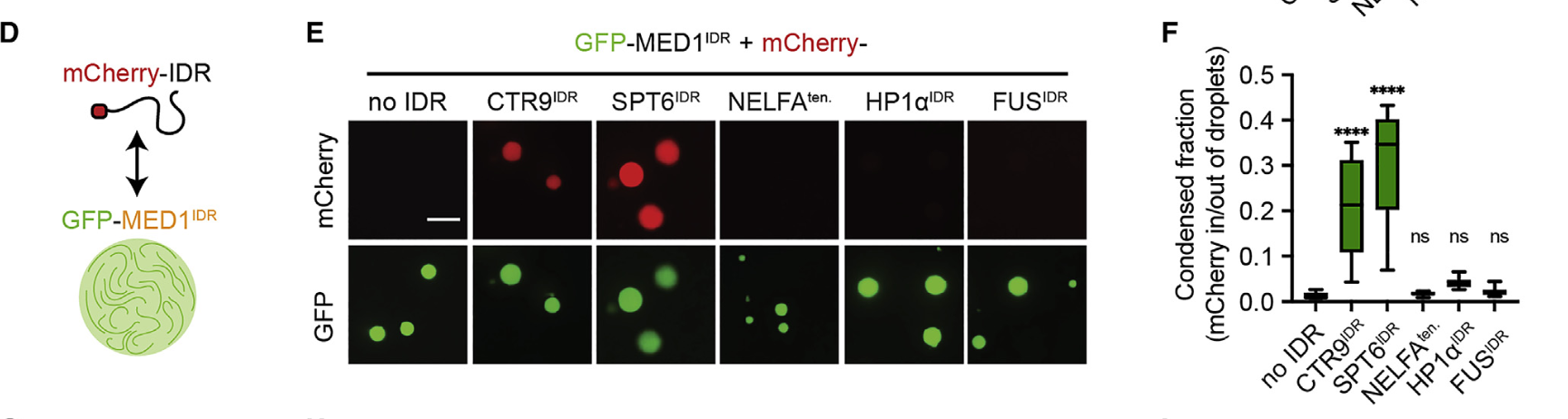
然后没有这个主人MED1的话,客人(IDR)也不会自主形成油相

结论就是:CTR9IDR和SPT6IDR足以选择性地分离到MED1IDR凝聚体中,而不需要任何其他蛋白或注释的相互作用域,也就是说客人只要来了IDR就可以了(甚至不需要全长序列来),这些客人/蛋白质也会被主人MED1邀请到油相中;
然后同样的,前面是体外数据,我们自然要接着做体内数据,所以要使用前面说的LacO阵列;
结论逻辑和上面体外的是一致的:
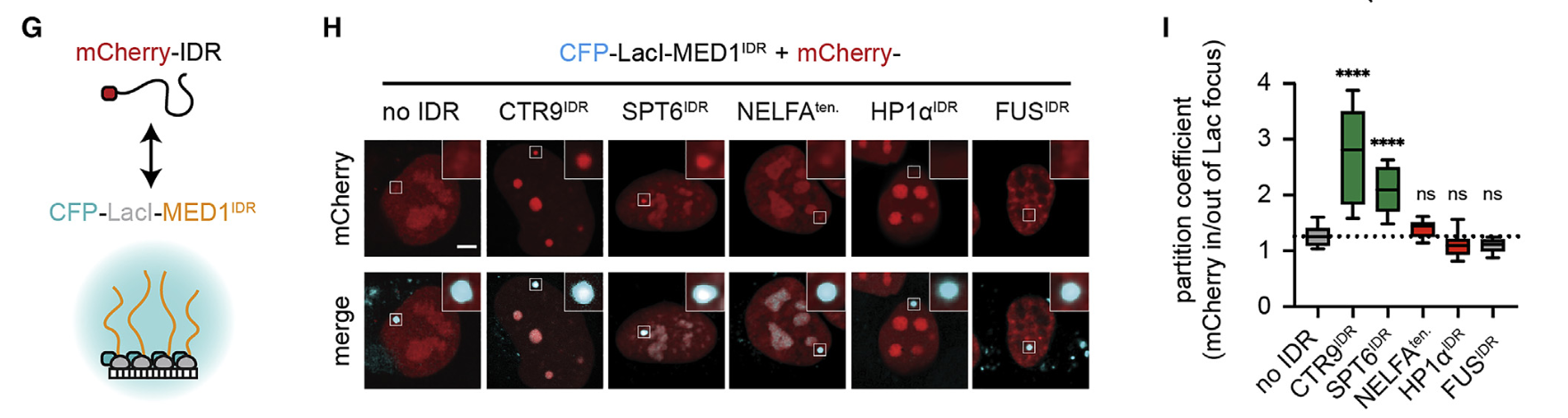
然后做了一个反转实验,效果相似;
同样没有主人MED1-IDR的话,客人就无法被邀请进去
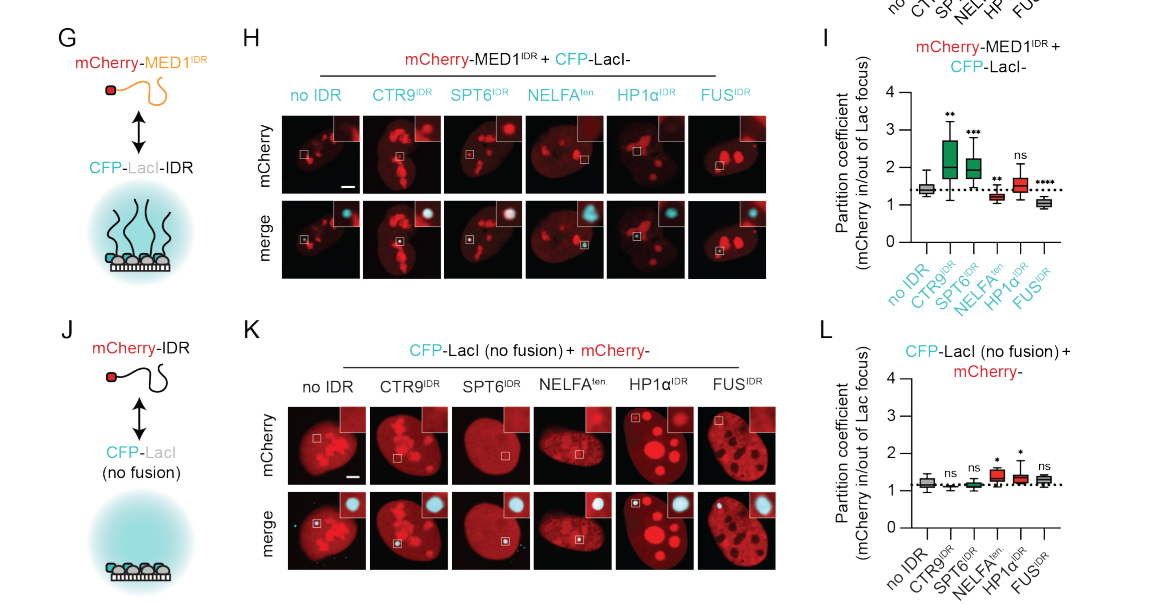
同样的充分性证明了:有IDR就足以被分选进去,那必要性呢,没了IDR呢?
紧接着做实验,还是3种序列变种,然后依然是体内做的
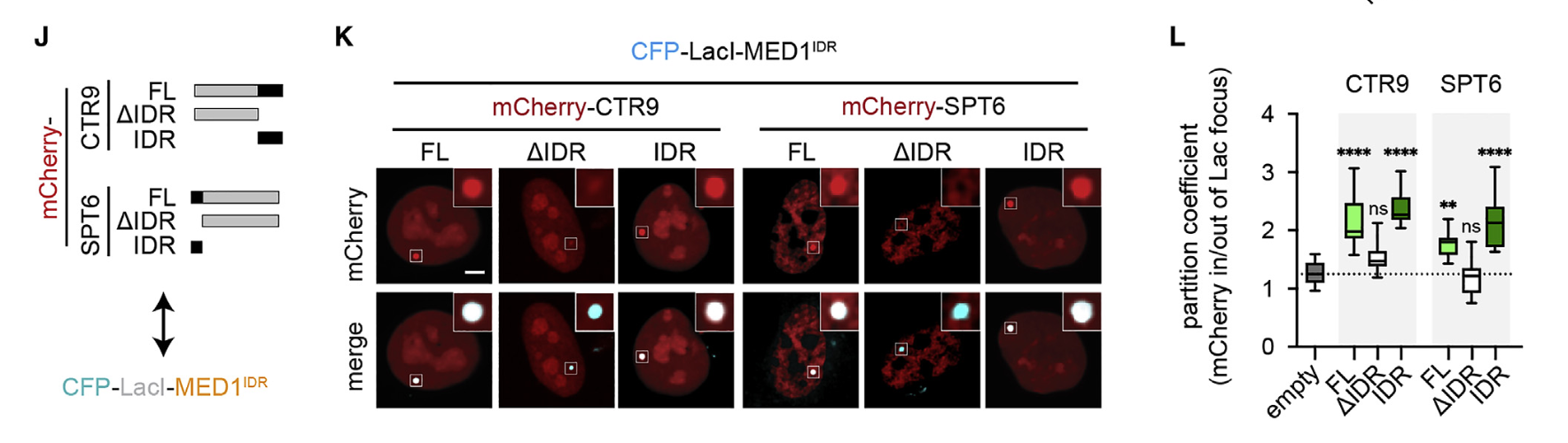
总之证明IDR对于分区是必要的。
接下来测试了来自两个类别的另外7个IDR:
(1)来自我们蛋白质组学数据中其他高度分割蛋白的idr (IWS1、LEO1和PAF1),
以及(2)与核凝聚物形成相关的其他IDR (DDX4、NELFE、EWSR1和p300)
这些数据其实是额外的:
有趣的是,EWSR1IDR和p300IDR都参与转录的正向调节,但实际上被排除,这表明转录凝集物之间存在专一性。这些结果表明,即使在紊乱的区域之间,也存在广泛的分区
说白了就是,主人还是那个主人MED1,但是用其他的客人的IDR,尽管也是IDR也是促进转录的,但不一定是被邀请进这个房间,可能是其他房间;
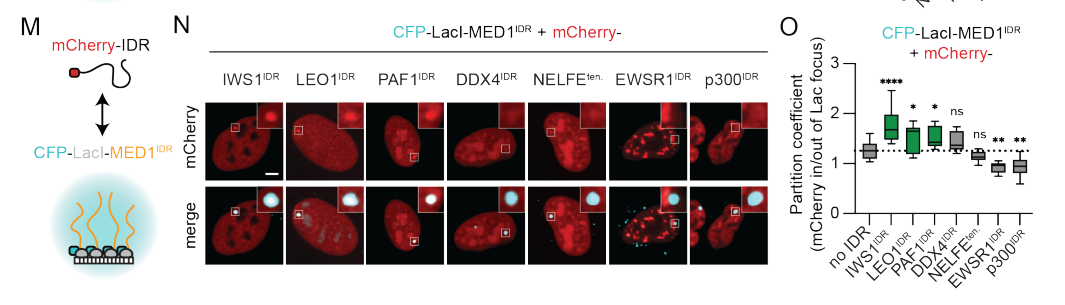
7,result7:选择性分割的IDR富集正氨基酸和负氨基酸的区块
IDR这么重要,是否有什么关键的序列特征?首先发现是带电的,而且这个电荷还呈现了模块化的分布模式
首先是依据IDR的分类,对这4种分类蛋白质做了一些统计分析:
为了了解导致观察到的选择性的IDR序列特征,我们比较了来自分区最多的蛋白质的IDR(分区IDR)、来自分区最少的蛋白质的IDR(排除IDR)、蛋白质组中所有IDR(所有IDR)(基于IDR的共识预测),以及来自我们颗粒试验中使用的核提取物中鉴定出的所有蛋白质的IDR(提取IDR)的序列参数。
正如预期的那样,所有组均有相似程度的促进disorder无序的氨基酸。
虽然芳香残基在相分离中发挥了众所周知的作用,但各组的芳香残基分数分布或模式相似。
与未分区IDR相比,分区IDR的带电氨基酸(E、D、R和K)比例显著较高,而A、G、Q和P比例显著较低。
与这些结果一致,分区IDR的带电残基比例(FCR)、酸性残基比例和碱性残基比例显著较高,但分区idr作为一个组,每个残基的净电荷(NCPR)没有差异。
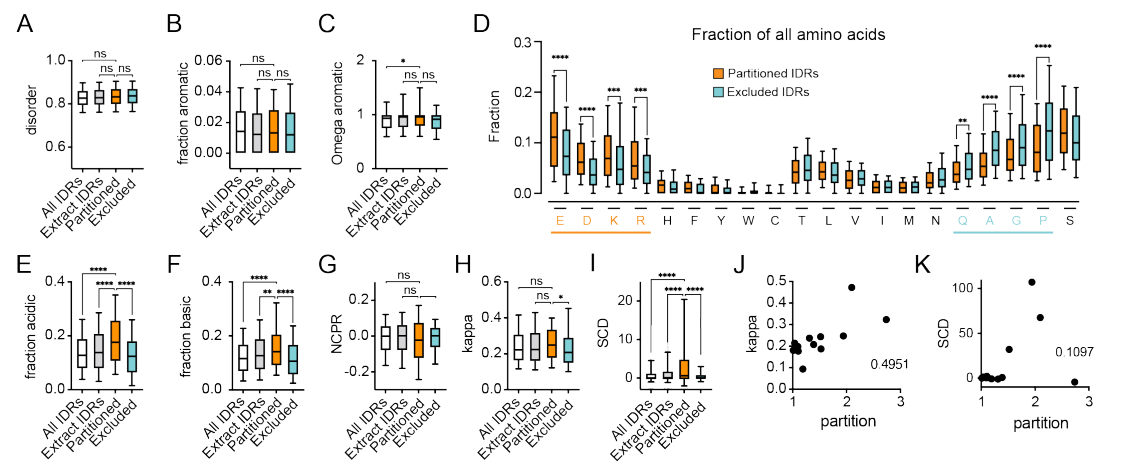
总之比较了各种值,还是选择一个最简单的方式来查看电荷分布的block/区域特征;
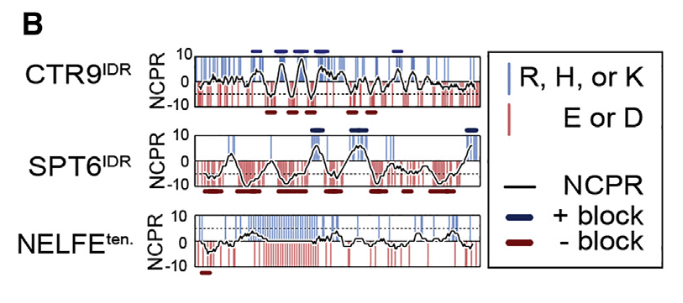
方法其实很原始,就是滑动窗口

pattern of alternating charge blocks
blocky pattern
块状图纹
NCPR block definition
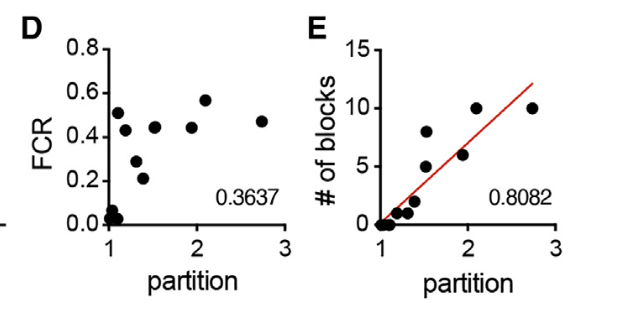
总之评估了很多参数和分选系数的相关性,只有block的数目线性拟合最优:
这些数据引导我们假设多价电荷块负责优先分配到MED1IDR凝聚体:multivalent charge blocks
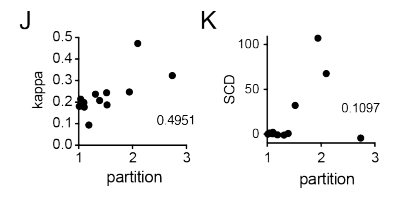
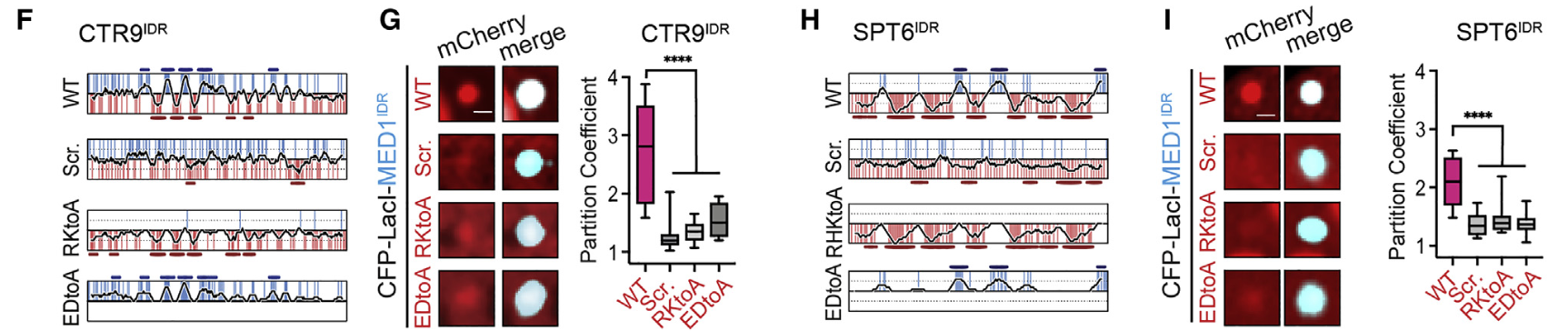
8,result8:CTR9IDR和SPT6IDR上电荷氨基酸的图案和局部密度是选择性分配所必需的
很自然的,有这种分布图案模式,我们就要打乱去看看没有这种分布规律会怎么样:
为了确定分区是否需要电荷模式,我们修改了CTR9IDR和SPT6IDR中的特定序列特征,并测试了它们对分区的影响;
怎么修改或者说是打乱这种序列模式呢,其实很简单,
如果单纯是打乱的话,我就用随机序列生成来替代这个电荷块,或者说将所有的酸碱性电荷块全部用A取代
scr是打乱,然后第3行是把RK变成A,最后一行是把DE变成A
至于为什么变成A用A取代,大概也许是A最小,不带电,没影响;
确实打乱之后分选现象就弱了
接着花样很多:还是scramble打乱序列
(1)只有带电残基的乱序(“带电乱序”)或(2)只有非带电残基的乱序(“非带电乱序”)
打乱带电分区有影响,打乱边上的序列影响基本没有
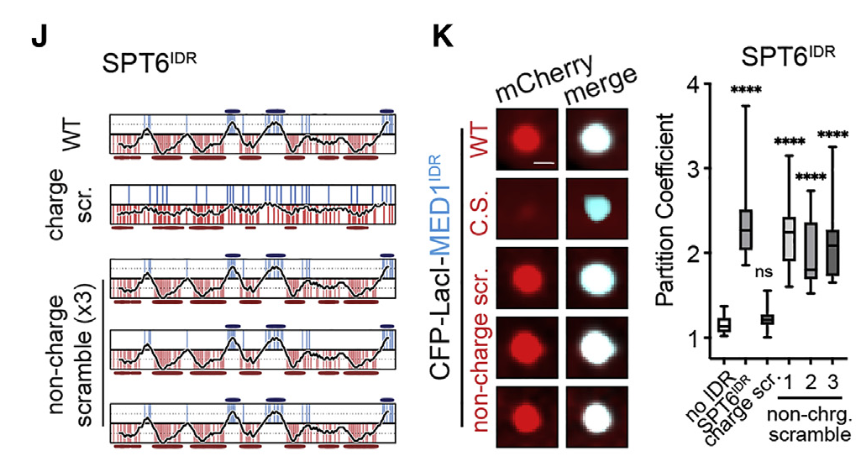
然后就是在SPT6中最显著交互的中间那一块(也就是ABABA那一块),这一块其实打乱之后就足以影响分选了
——》看实验荧光信号,或者看统计显著性
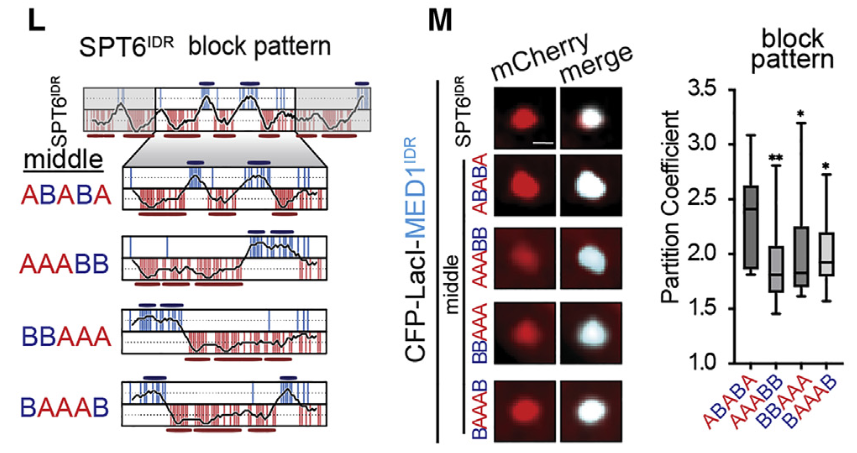
结论就是:
模式和局部电荷残基密度对idr介导的分配是重要的
这里的patterning and local density of charged residues,注意是对于残基而言的
所以计算block是patterning
density则是单位(区域内)的block数目——》个人理解
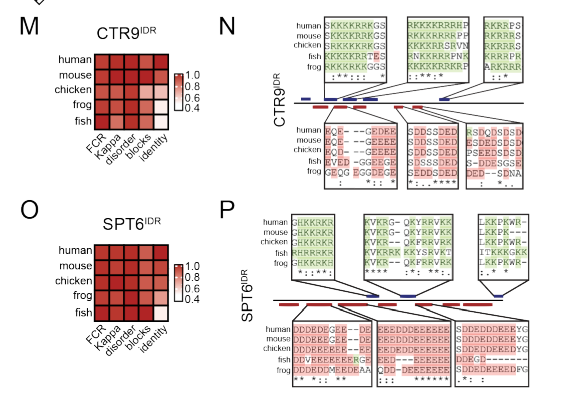
接着做这些block区域的保守性分析:
CTR9IDR和SPT6IDR内的电荷区是保守的,即使在分析的物种之间位置序列的一致性不同,挺保守的
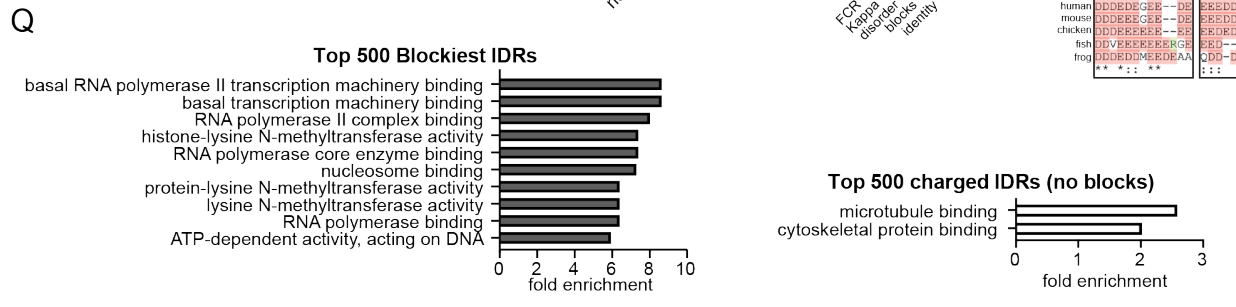
为了了解人类蛋白质组中哪些类型的蛋白质含有带电荷块的IDR:
有最多block数目的IDR被富集用于RNA Pol II转录的正向调节,而无block的电荷富集IDR被富集于细胞骨架蛋白结合
9,result9:工程化设计的IDR(包含交替电荷块)(能够)选择性地分选
在证明了模式电荷块是选择性分区所必需的之后,我们下一步将这些特征改造到蛋白质中,以实现功能获得性分区
——》这个没啥好说的了,就是纯粹工程性质应用了
其实就像基因编辑一样了,直接通过人为操纵电荷块的分布模式,让客人能够进入主人的房间
——》电荷分布模式可以决定互溶互斥性
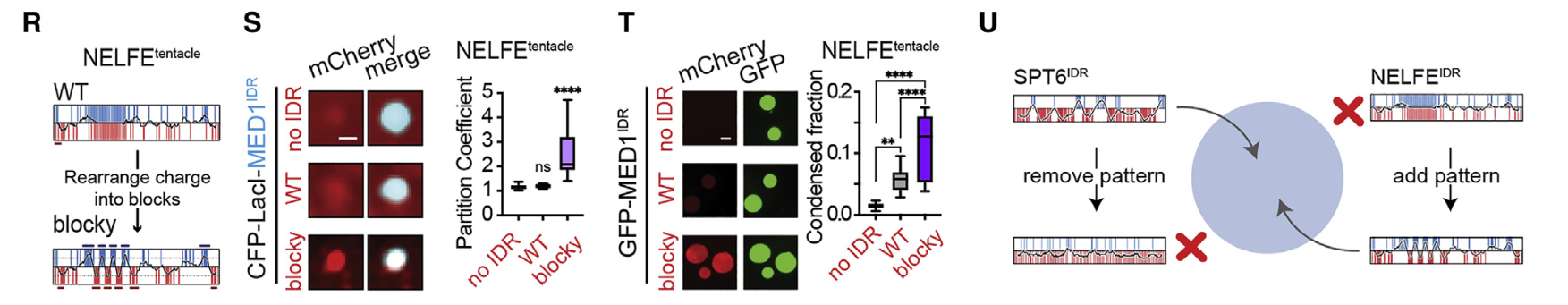
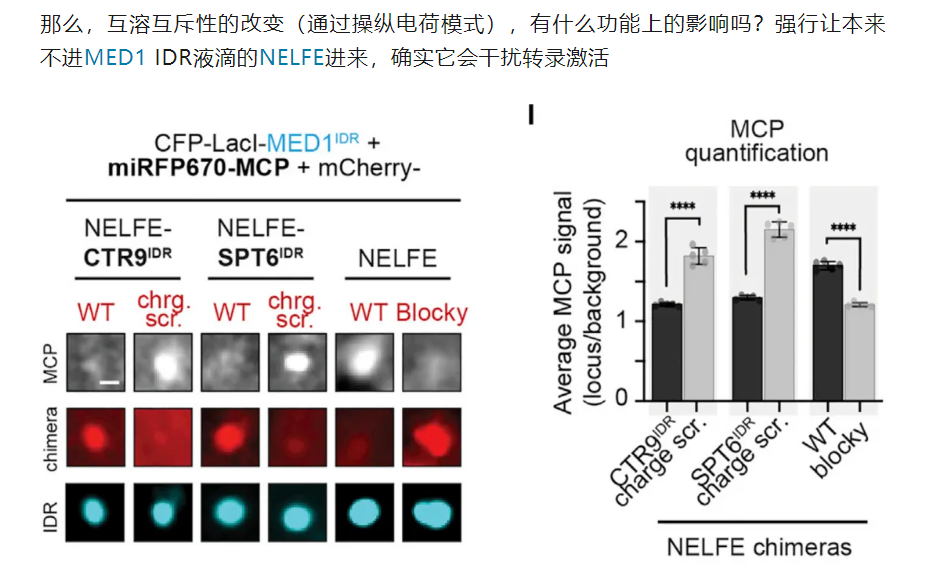
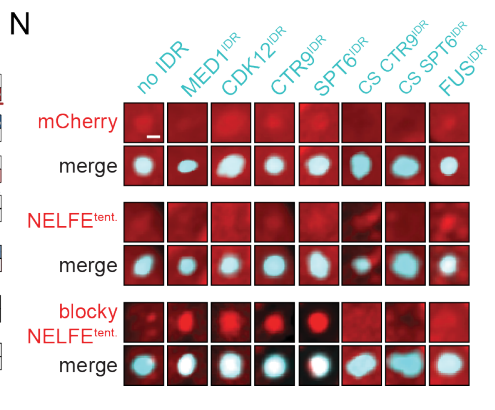
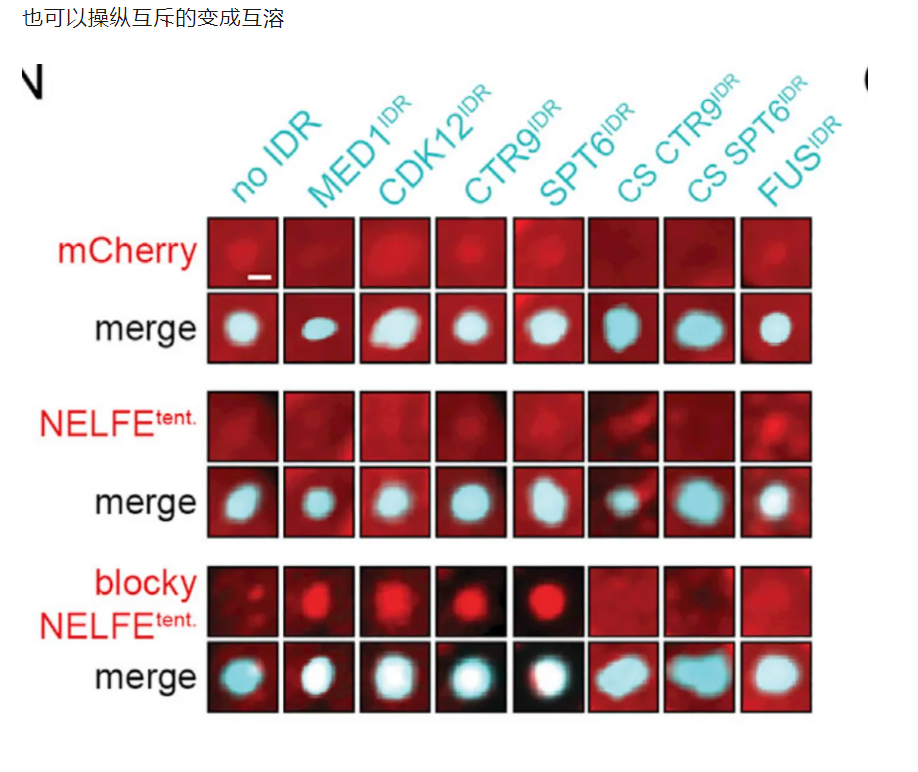
10,result10:在NELFE中引入具有交替电荷块的idr导致了NELFE复合体的分选和基因激活的下降
很自然的,有模式的要打乱模式,看能否再次被分选;
没有模式的要强加给它,看是否会被异常分选进入凝聚物中
当然就是负样本的NELFE了
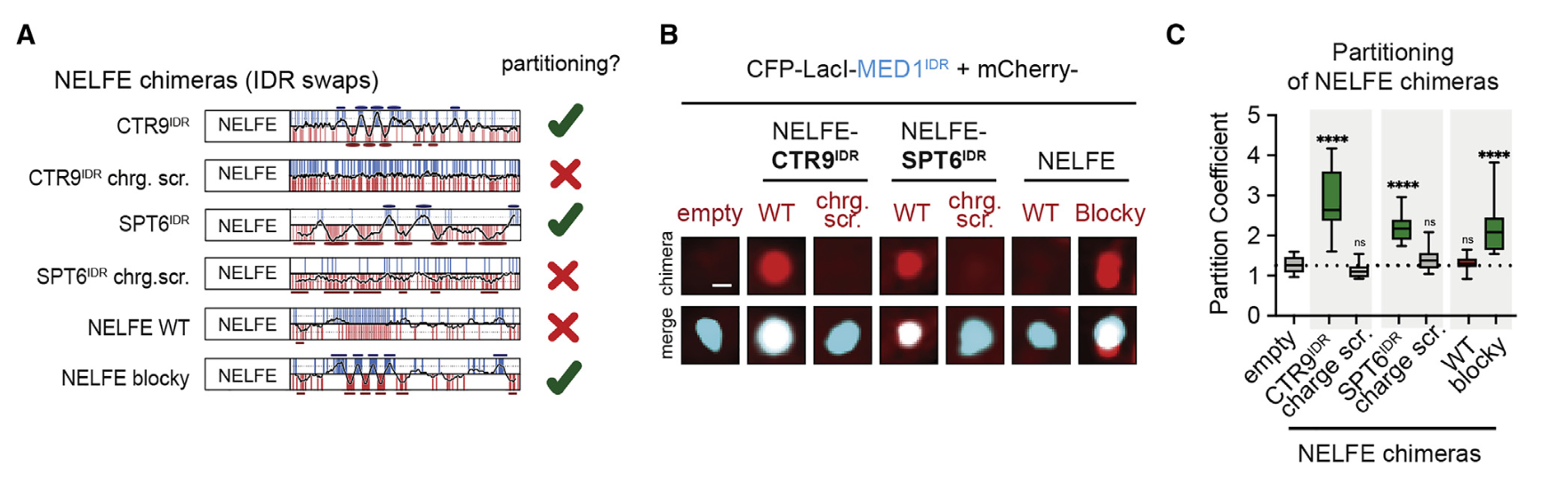
至于这个模式,就直接用了CTR9或者SPT6蛋白的IDR了
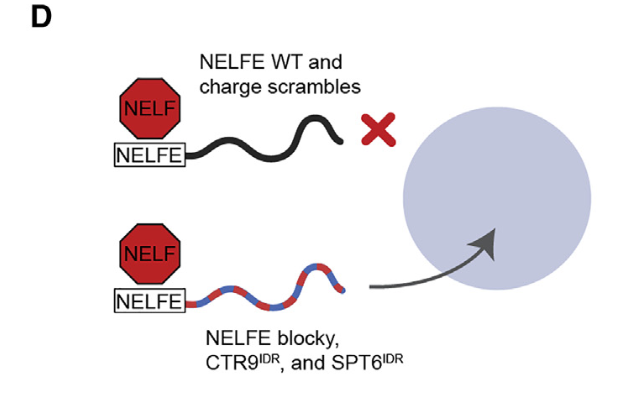
结论——》蛋白质复合物可以通过其idr中的序列特征进行空间组织,从而对基因活性产生功能性影响
11,result11:MED1IDR上带电氨基酸的模式和局部密度对于选择性分配、报告基因激活和细胞状态转换是必需的
很自然,不能光看招募过来的客人的IDR长啥样,主人MED1的IDR也要知道长啥样
scrambled and charge-to-alanine substitution mutants
同样主人的IDR变了,这个房间就不存在了
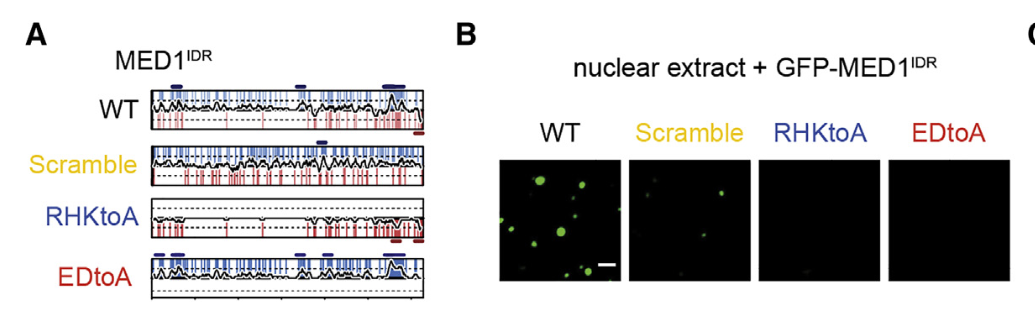
体外做了再体内,MED1-IDR的模式还是很重要
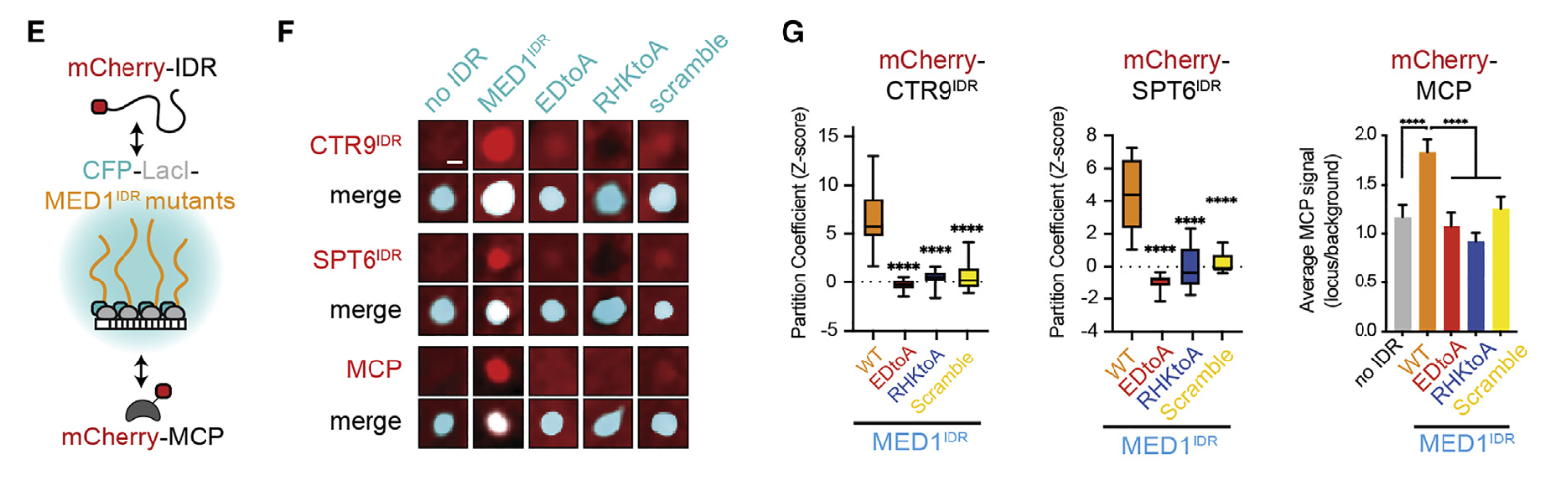
总之别看MED1-IDR长得丑,还没规律,但是这种模式就是很重要,破坏了就影响很大;
后续做了功能研究,都表明:MED1IDR上带电氨基酸的模式和局部密度对于选择性分配、报告基因激活和细胞状态转换是必需的
12,result12:具有相似块模式的idr具有相似的分区和功能
为了测试其他带有电荷块的IDR是否具有与MED1IDR相似的功能,
我们测试了三个包含交替电荷块的IDR (CDK12IDR、CTR9IDR和SPT6IDR)、
两个带电残基被打乱的对照IDR (CTR9IDR和SPT6IDR的电荷乱序)
和一个耗尽电荷块的IDR (FUSIDR)
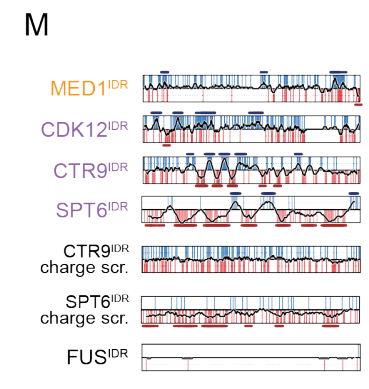
结果就是有相似的block模式,即使不是MED1,也能分选CTR9和SPT6蛋白,然后也同样拒绝了NELFE
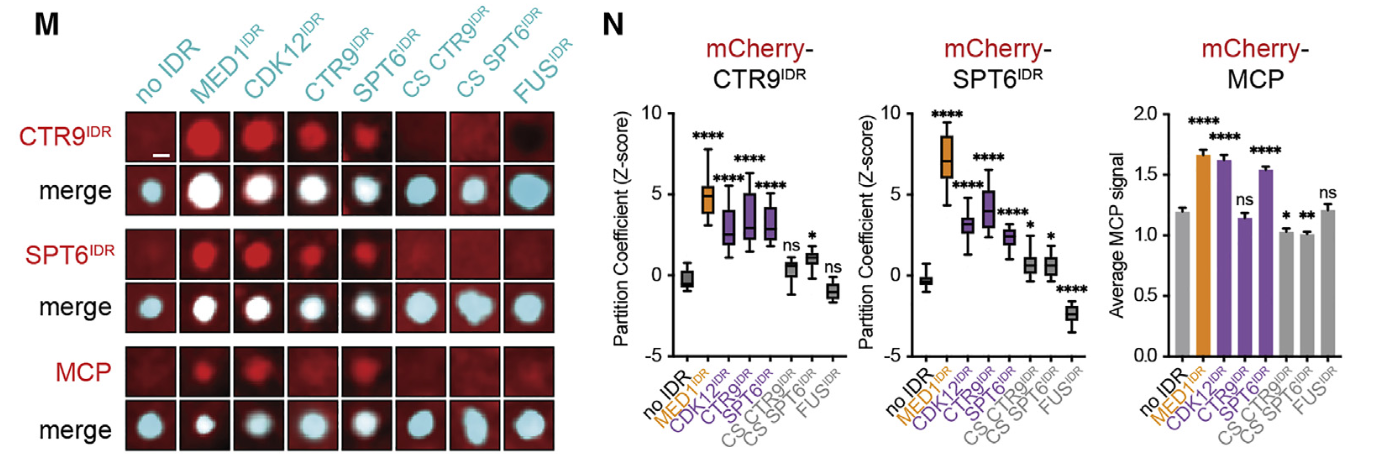
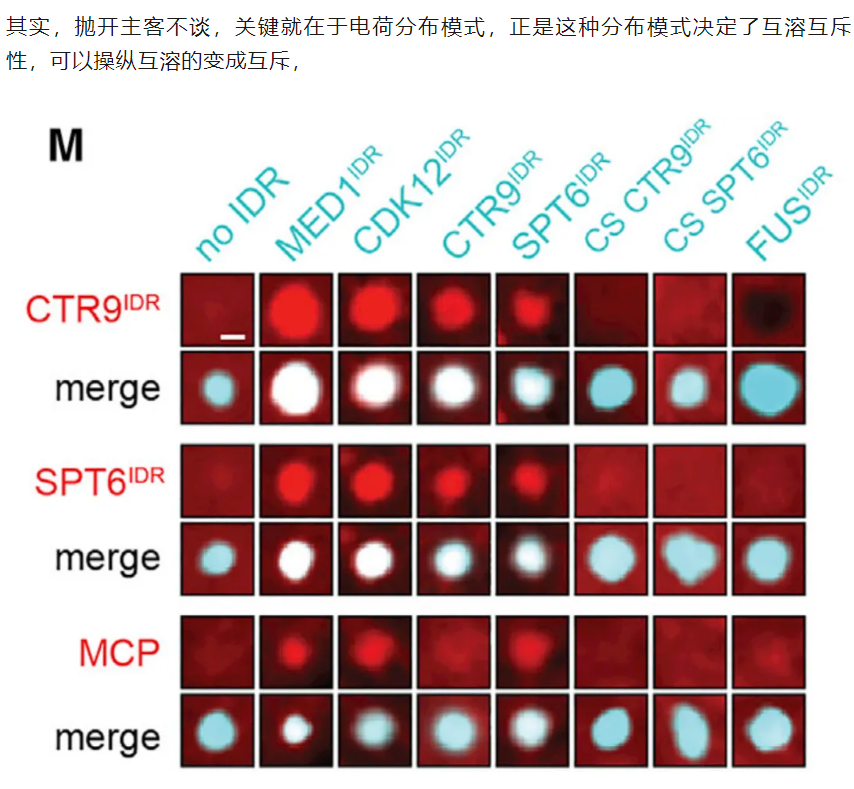
总之结论就是:没有共享位置序列身份的idr由于其共享电荷模式的性质而表现出相似的分区和功能,并表明idr上的模式和局部电荷密度通常是选择性分区、报告基因激活和细胞状态转换所需要的
二,讨论:
这里描述的紊乱区域之间的多价相互作用,与位点特异性结构介导的相互作用(例如,DNA/ RNA结合域,特定蛋白-蛋白相互作用域等)一起,通过在特定位点创建功能特化的局部蛋白质组来调节转录。
Multivalent interactions among disordered regions, described here, work together with site-specific structure-mediated interactions (e.g., DNA/ RNA-binding domains, specific protein-protein interaction domains, etc.) to regulate transcription by creating functionally specialized local proteomes at specific loci。
其实上面讨论的IDR比较特殊,是蛋白质-蛋白质之间的相聚集;
如果要讨论DNA、RNA那就是motif了,而且蛋白质和DNA、RNA之间那就不是简单的IDR上的多价电荷块的作用了,
因为蛋白-蛋白都有多价电荷块,但是蛋白-核酸就难说了,所以后面CTCF那篇文献就只说了NCR,为什么不谈论相分离呢,不好说,至少电荷-电荷互作这一条路,大概只能对于蛋白质对蛋白质。

我们的研究支持了一种模型,即不同的凝聚体部分通过利用idr之间特定的模式和类型的相互作用来分配生物分子。在转录方面,一些研究表明idr足以将蛋白质靶向到特定的基因组位点。在形成凝集物的IDR序列中,成分的复杂性有很大的差异,这表明存在分割机制尚未被发现。
当我们关注idr时,应该注意到凝集物的形成不需要蛋白质紊乱。
相分离有很多其他的机制:

由idr介导的多价相互作用可以选择性地从复杂的蛋白质混合物中分离功能相关的蛋白质。这种选择性所需的序列特征对转录活性也是必需的。
1,虽然许多由MED1IDR凝聚物划分的蛋白质含有具有多个图型电荷块的idr,但有些idr不具有这些特征,这表明蛋白质可以通过其他方式进行划分。
2,虽然我们证明了必要性和充分性,但我们没有揭示交替电荷块如何使特异性。
Sources:
https://pmc.ncbi.nlm.nih.gov/articles/PMC9910284/#SD5
其实发现就算是cell,也是可以正大光明地使用其他人的GEO数据的·


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









