







摘要:人类大脑具有独特的自发思维模式,这使得每个大脑可以通过其神经活动被唯一识别。然而,深度全身麻醉会抑制主观体验,然而不可知它是否会抑制人脑的独特性。在此,本研究使用在全身麻醉剂七氟醚和丙泊酚作用下获得的功能磁共振成像(fMRI)扫描,来确定麻醉诱导的无意识状态是否会降低人类大脑的独特性,无论是在与其他个体的大脑相比,还是在与其他物种的大脑相比。
通过功能连接,研究报告显示,在麻醉状态下,个体大脑的自我相似性降低,彼此之间的可区分性也降低。独特性丧失是高度有组织的:它与原型的感官-联想轴共定位,并与人类与其他灵长类动物之间系统发育差异的遗传和形态学标记相关。这种效应在更深的麻醉深度下更为明显,且在七氟醚和丙泊酚之间具有可重复性,并在恢复后逆转。
作为一致性的证据,研究表明麻醉将人类大脑的功能连接更接近低维空间中猕猴大脑的功能连接。最后,麻醉降低了自发大脑活动与从Neurosynth 元分析引擎聚合的认知大脑模式之间的匹配度。
总体而言,本研究结果揭示,麻醉状态下的人类大脑不仅彼此之间的可区分性降低,而且与其他灵长类动物的大脑之间的可区分性也降低,尤其是那些在人类中扩展的特定区域受到麻醉的影响最大。
1. 引言
意识——在麻醉和无梦睡眠中丧失、并在苏醒时恢复的——本质上因人而异,正如“主观体验”和“第一人称体验”这两个近义词的使用所示。换句话说,每个人的意识都是独特的。这引发了一个引人入胜的问题:如果意识使我们每个人变得独特,那么在失去意识时,我们会变得更加相似吗?一种短暂的无意识状态可以通过麻醉剂诱导产生。麻醉的医学益处已得到充分证明,但将其作为一种研究大脑功能的工具的价值也在不断增加。与自发睡眠不同,麻醉引起的无意识状态(表现为行为反应的丧失)可以通过实验控制:它可以被可靠地诱导、维持和逆转。
在这篇文章中,我们结合了由深度麻醉(使用不同麻醉剂——七氟烷和丙泊酚)引起的意识的丧失和恢复,以及对人类大脑自发活动的功能性磁共振成像(fMRI)记录。我们提出以下问题:在无意识状态下,人脑是否会失去其独特性?我们从三个概念角度来探讨这一问题。
首先,我们比较了个体内部和个体之间的大脑活动。开创性研究表明,大脑区域之间的功能连接模式在个体间具有显著差异,这使得可以根据fMRI扫描结果对个体进行“大脑指纹识别”。因此,我们利用功能连接组指纹识别来评估在深度麻醉(假设会导致无意识,注意这与单纯的镇静不同,在镇静状态下参与者仍然具有反应性和意识,尽管反应较迟缓)下,个体是否变得不太容易区分。
其次,我们评估了每位受试者在不同麻醉水平下的大脑活动与认知操作的典型脑图之间的对应程度。这些典型脑图是通过整合超过14,000项神经成像实验得到的。尽管我们的研究涉及无任务的功能性磁共振成像,但我们推断,即使在没有任何任务的情况下,大脑仍可能自发地进入与各种认知操作相关联的状态。然而,这种情况在意识丧失时不应发生,因为即使是内在驱动的认知活动也应被消除。这一范式受到以下证据的启发:检测大脑对特定任务(如想象打网球或想象在家中导航)的反应能力,是即使在行为上无反应(由于意识障碍)的个体中,意识的有力标志。
最后,我们探讨了麻醉是否会使得人类大脑的功能连接变得与其他物种更为相似——换句话说,减少我们物种与其他灵长类动物相比的独特性。
2. 方法
数据集
七氟醚数据
此处包含的七氟醚数据之前已发表过,具体细节可参考原始文献。该研究获得了德国慕尼黑工业大学医学院伦理委员会的批准,并按照赫尔辛基宣言进行。研究开始前至少48小时,已从志愿者处获得书面知情同意。研究招募了20名20至36岁的健康成年男性(平均年龄26岁),通过校园公告和个人联系招募,并为他们的参与支付了报酬。在纳入研究前,详细向参与者介绍了研究方案和风险,并审查了病史,以评估是否存在既往神经系统或精神疾病。进行了重点体格检查,并记录了静息心电图。进一步的排除标准如下:美国麻醉医师协会(ASA)体格状态非I级,长期服用药物或毒品,听力障碍或耳聋,不会说流利的德语,已知或怀疑易患恶性高热,急性肝性卟啉病,氟烷肝炎病史,肥胖且体质指数(BMI)>30 kg/m²,有胃食管反流倾向的胃肠道疾病,已知或怀疑气道困难,以及体内有金属植入物。数据采集时间为2013年6月至12月。
七氟醚浓度的选择是让受试者能够耐受人工通气(达到2.0体积%),并且所有受试者都能达到爆发抑制(约为4.4体积%)。为了便于进行组间比较,还使用了中间浓度3.0体积%。在磁共振成像(MRI)扫描仪中,受试者闭眼静息700秒。由于在MRI扫描过程中同时采集了脑电图(EEG)数据(尽管在本研究中未进行分析),因此通过视觉在线检查EEG来确认受试者在麻醉前基线扫描期间没有入睡。通过一个紧贴面部的面罩,使用兼容fMRI的麻醉机(Dräger的Fabius Tiro)将七氟醚与氧气混合后输送。进行标准的美国麻醉医师协会监测:使用心肺监护仪(DatexaS;通用电气)监测七氟醚、氧气和二氧化碳的浓度。在给予0.4体积%的七氟醚呼气末浓度5分钟后,每3分钟将七氟醚浓度逐步增加0.2体积%,直到受试者失去对反复说出的指令“捏我的手”的反应,判断为意识丧失。
随后,七氟醚浓度被提高以达到大约3体积%的呼气末浓度。在临床需要时,由医生管理通气,并插入适合fMRI的喉罩(I-gel,Intersurgical)。然后将吸入氧浓度设定为0.8,并调整机械通气,以在爆发抑制期间维持呼气末二氧化碳浓度稳定在33±1.71 mmHg,在3体积%浓度时维持在34±1.12 mmHg,在2体积%浓度时维持在33±1.49 mmHg(本文中,数据以均值±标准差表示)。通过手背静脉中的静脉导管持续输注去甲肾上腺素(0.1±0.01 μg·kg⁻¹·min⁻¹),以维持平均动脉血压接近基线值(基线:96±9.36 mmHg;爆发抑制:88±7.55 mmHg;3体积%:88±8.4 mmHg;2体积%:89±9.37 mmHg;后续:98±9.41 mmHg)。在插入喉罩后,七氟醚浓度逐渐增加,直到脑电图显示爆发抑制,抑制期至少持续1000毫秒,且约50%的电活动被抑制(达到4.34±0.22体积%),这是深度麻醉的特征。此时,又记录了700秒的脑电图和fMRI结果。随后,在呼气末七氟醚浓度稳定在3体积%和2体积%时,分别又采集了700秒的数据(分别对应Ramsay评分6级,即最深的麻醉水平),每次采集前都有15分钟的平衡时间。最后一步,将呼气末七氟醚浓度降低到失去反应性时浓度的两倍。然而,在此条件下,大多数参与者出现移动或无法耐受喉罩,因此这一阶段的数据未纳入分析。随后停止七氟醚给药,将扫描床移出MRI扫描仪,监测麻醉后恢复情况。参与者在自主呼吸恢复前一直接受手动通气。一旦患者按指令张嘴,就移除喉罩。医生定期要求参与者捏手:一旦遵从指令,即记录恢复反应性的时间。在恢复反应性的时间起15分钟后,进行Brice访谈,以评估七氟醚暴露期间是否存在意识;次日通过电话重复访谈。在总共45分钟的恢复时间后,又进行了一次静息态fMRI-EEG联合扫描(闭眼,与基线扫描相同)。当参与者清醒、定向、合作且生理状态稳定时,由事先指定的家人或朋友将其带回家。
尽管原始研究同时采集了fMRI和EEG数据,但本研究仅考虑了fMRI数据。数据采集使用的是3.0T磁共振成像扫描仪(Philips Achieva Quasar Dual 3.0T 16CH),配备八通道相控阵头线圈。数据采集使用梯度回波平面成像序列(回波时间=30毫秒,重复时间(TR)=1.838秒;翻转角=75°;视野=220×220毫米²;矩阵=72×72;32层;层厚=3毫米;层间距=1毫米;采集时间=700秒)。在功能扫描之前,使用T1加权磁化准备快速梯度回波(MPRAGE)序列采集解剖扫描,覆盖整个大脑,体素为240×240×170(体素大小=1×1×1毫米)。共有16名志愿者完成了整个协议;由于运动过多,排除了1名参与者,最终纳入分析的样本量为n=15。
人类异丙酚数据
异丙酚数据于2014年5月至11月在加拿大安大略省伦敦市的西方大学罗巴茨研究所收集,并已发表。该研究获得了西方大学健康科学研究伦理委员会和心理学研究伦理委员会的批准。共招募了19名健康志愿者(年龄18-40岁;13名男性)。志愿者均为右撇子,以英语为母语,且无神经系统疾病史。根据相关伦理指南,每位志愿者均提供了书面知情同意,并因参与研究而获得了报酬。由于设备故障或扫描仪内麻醉的生理障碍,3名参与者(1名男性)的数据被排除在分析之外,最终纳入分析的样本量为n=16。
在不同异丙酚水平下采集了静息态fMRI数据:无镇静(清醒)、深度麻醉(对应Ramsay评分为5)以及麻醉后恢复期。如之前报道,对于每种条件,fMRI采集开始前,两名麻醉师和一名麻醉护士在扫描室独立评估了Ramsay水平。由于麻醉师和麻醉护士的部分职责是确定参与者的麻醉水平,因此他们无法对实验条件进行盲处理。需要注意的是,Ramsay评分是为重症监护患者设计的;因此,在异丙酚给药前的清醒状态下,参与者并未获得评分。相反,他们被要求保持完全清醒、警觉并进行适当沟通。为了进一步独立评估参与者的反应性水平,要求他们完成两项任务:一项言语记忆回忆测试和一项基于计算机的听觉目标检测任务。此外,通过放置在扫描仪内的红外摄像机监测清醒状态。通过AS50自动注射泵(Baxter Healthcare)静脉注射异丙酚。结合计算机控制的注射泵,使用效应部位/血浆导向算法实现逐步镇静增量,随后根据需要手动调整,以根据TIVA Trainer(欧洲静脉麻醉学会)药代动力学模拟程序达到所需的异丙酚目标浓度。该软件还指定了根据Marsh三室模型的异丙酚血药浓度,这些浓度被用作提供目标控制输注的药代动力学模型的目标。在初始异丙酚目标效应部位浓度为0.6 μg/ml后,浓度逐渐以0.3 μg/ml的增量增加,并在每次增量后评估Ramsay评分。如果Ramsay评分<5,则进行进一步增量。通过TIVA Trainer注射泵提供的药代动力学模型保持异丙酚的平均估计效应部位和血浆浓度稳定。当参与者停止对口头指令做出反应、无法进行对话且仅对物理刺激有反应时,达到Ramsay 5级。当两名麻醉师和麻醉护士一致认为达到Ramsay 5级镇静水平且参与者对两项任务均无反应时,开始数据采集。异丙酚的平均估计效应部位浓度为2.48(1.82-3.14)μg/ml,平均估计血浆异丙酚浓度为2.68(1.92-3.44)μg/ml。平均给药总量为486.58(373.30-599.86)mg。这些变异性值是异丙酚药代动力学和药效学的典型表现。通过调节氧气以维持SpO₂高于96%。在Ramsay 5级时,参与者能够维持自主心血管功能和通气。然而,由于该镇静程序并非在医院环境中进行,因此无法在扫描期间使用插管来确保气道安全。因此,尽管有两名麻醉师密切监测每位参与者,但扫描时间被尽量缩短,以确保在深度镇静后恢复正常呼吸。在本研究中,纳入的参与者在深度镇静扫描期间均未出现状态变化或运动。在深度麻醉扫描后停止异丙酚给药,参与者大约在11分钟后达到Ramsay 2级,表现为对口头指令的清晰且迅速反应,这对应于恢复期。如之前报道,一旦进入扫描仪,参与者被指示闭眼放松,但不要入睡。在没有任何任务的情况下,为每位参与者采集了8分钟的静息态fMRI结果。此外,还通过耳机播放一个情节驱动的故事(持续5分钟)进行听觉呈现,并采集了扫描数据。参与者被指示在保持眼睛闭合的情况下聆听。本分析仅重点关注静息态数据。
用3T西门子Tim Trio扫描仪(32通道线圈)进行MRI扫描,从每位参与者处收集了256个体积的功能图像(回波平面成像,EPI),参数如下:切片数=33;层间距=25%;分辨率=3 mm各向同性;重复时间(TR)=2000 ms;回波时间(TE)=30 ms;翻转角=75度;矩阵尺寸=64×64。采集顺序为交错式,从底部向上。还进行了解剖扫描,使用3D MPRAGE序列获取高分辨率T1加权体积(32通道线圈;1 mm各向同性体素尺寸),参数如下:采集时间(TA)=5分钟,TE=4.25 ms;矩阵尺寸=240×256;翻转角=9°。
功能磁共振成像预处理和去噪
我们按照之前的麻醉数据研究,使用CONN工具箱(版本17f;http://www.nitrc.org/projects/conn)在MATLAB 2016a中实现了标准的预处理流程。流程包括以下步骤:移除前10秒以达到稳态磁化;运动校正;切片时间校正;通过质量保证/伪影检测软件ART(http://www.nitrc.org/)识别用于后续剔除的异常体积;并将数据归一化到蒙特利尔神经研究所(MNI-152)标准空间(2 mm各向同性重采样分辨率),使用每位参与者的T1加权解剖图像分割的灰质图像以及先验的灰质模板。
去噪也使用CONN工具箱完成,方法与之前药理磁共振成像数据集的研究一致。药理学药物可能会引起生理参数(心率、呼吸频率和运动)或神经血管耦合的改变。解剖成分基础噪声校正(aCompCor)方法通过从不太可能被神经活动调节的区域提取主成分来去除生理波动,这些成分随后被作为干扰回归因子。按照这种方法,从白质和脑脊液信号中提取了五个主成分(使用从T1加权结构MRI图像获得的个体组织掩模),并将其与六个个体特定的重定位参数(三个平移和三个旋转)以及它们的一阶时间导数一起从功能数据中回归。随后,使用普通最小二乘回归剔除ART识别的异常值。最后,去噪后的BOLD信号时间序列被线性去趋势,并进行带通滤波以消除低频漂移效应和高频噪声,保留0.008至0.090 Hz之间的频率。全球信号回归作为一种去噪方法在文献中受到了广泛关注-。然而,最近的研究表明,全球信号包含与行为相关的信息,并且在药理学和病理学干扰下关于意识状态的信息至关重要。因此,与我们和其他人的先前研究一致,这里我们避免了全球信号回归,转而采用aCompCor去噪程序,这是推荐的方法之一。最后,去噪后的BOLD信号被划分为200个皮质感兴趣区域(ROIs),来自Schaefer图谱。我们还使用68个ROI的解剖学Desikan–Killiany皮质划分以及包含200个皮质ROI的Schaefer图谱和额外32个皮下ROI的联合皮质–皮下图谱(来自Tian等人的皮下图谱)复制了我们的结果,如先前推荐。为了与猕猴数据进行比较,使用了由Kötter和Wanke的82个ROI皮质划分的人类适应版本,如参考文献所述。对于每个个体和每个条件,功能连接(FC)被估计为去噪和划分后的BOLD时间序列对之间的皮尔逊相关性。
来自PRIME-DE的清醒猕猴fMRI数据
第一个非人灵长类动物MRI数据集是作为灵长类动物神经影像数据交换(PRIME-DE)猴MRI数据共享计划的一部分提供的——这是一个最近推出的非人灵长类动物成像开放资源。
数据预处理和去噪遵循之前出版物中的相同程序。我们使用了在纽卡斯尔大学扫描的恒河猴(Macaca mulatta)的fMRI数据。样本包括14个样本(12只雄性和2只雌性),年龄分布在3.90至13.14岁之间,体重分布在7.2至18.0公斤之间。
猕猴MRI 数据采集
猕猴在垂直的Bruker 4.7T 灵长类动物专用扫描仪中进行扫描,使用单通道或四至八通道并行成像线圈。扫描过程中未使用对比剂。在数据采集前,通过 Bruker 的二阶匀场和自定义扫描序列优化对磁场进行优化。猕猴在扫描过程中保持直立,使用 MRI 兼容的头部支架或非侵入性头部固定装置,并在执行任务或处于静息状态时进行扫描(本研究仅包括静息态扫描)。在扫描过程中,使用眼动追踪、视频和音频监测。静息态扫描持续时间为 21.6 分钟,参数如下:重复时间(TR)= 2600 毫秒;回波时间(TE)= 17 毫秒;有效回波间隔 = 0.63 毫秒;体素大小 = 1.22×1.22×1.24 毫米³;相位编码方向 = 按列编码。结构扫描包括 T1 加权的 MDEFT 序列,参数如下:TE = 6 毫秒;TR = 750 毫秒;反转延迟 = 700 毫秒;切片数 = 22;成像视野 = 12.8×9.6 厘米²;网格中的体素数 = 256×192;体素分辨率 = 0.5×0.5×2.0 毫米³;分段数 = 8。
猕猴MRI 数据使用最近开发的非人灵长类动物 MRI 分析流程 Pypreclin 进行预处理,该流程针对猴子研究的特定需求进行了优化。该流程的详细描述见相关出版物。简而言之,它包括以下步骤:(1)切片时间校正;(2)纠正由运动引起的、随时间变化的 B0 不均匀性;(3)将采集位置重新定向到模板(本研究使用了最近开发的国家心理健康猕猴模板——这是一个由 31 只恒河猴(Macaca mulatta)的体内 MRI 生成的高分辨率平均猕猴大脑模板);(4)使用 FSL 的 MCFLIRT 功能对数据进行重新对齐到中间体积;(5)使用 Joe 的图像程序(JIP-align)例程进行归一化和掩模处理(该例程专为临床前研究设计,归一化步骤通过仿射变换和非线性变形场对齐将解剖数据对齐到通用模板空间);(6)进行 B1 场校正,以纠正数据中的低频强度不均匀性;(7)使用 JIP-align 对功能图像和解剖图像进行共配准,通过应用刚性变换将平均功能图像(移动图像)对齐到解剖图像(固定图像)。通过在归一化步骤中计算的变形场,将模板脑掩模变形以获得解剖脑掩模。然后,通过组合归一化和共配准的空间变换,将功能图像对齐到模板空间。
去噪
为了确保与人类数据分析流程的一致性,使用CONN 工具箱中实现的 aCompCor 去噪方法对猕猴 fMRI 数据进行去噪。从高分辨率国家心理健康猕猴模板的相应概率组织图中获取白质和脑脊液掩模(掩模被侵蚀了 1 个体素),并将它们的前五个主成分从功能数据中回归出去。同时,还回归了线性趋势和六个运动参数(三个平移和三个旋转)及其一阶导数。为了使人类和猕猴数据具有可比性,猕猴数据也在与人类数据相同的 0.008–0.090 Hz 范围内进行了带通滤波。
数据集说明
在纽卡斯尔样本的14 只动物中,有 10 只有可用的清醒静息态 fMRI 数据;在这些 10 只动物中,除了第一只动物外,其余动物都有两个扫描会话可用。因此,总共有 19 个不同的扫描会话,涉及 10 只猕猴。
来自虚拟大脑项目的麻醉猕猴fMRI 数据
虚拟大脑项目提供了一个包含9 只成年雄性猕猴(8 只恒河猴和 1 只食蟹猴;年龄在 4 到 8 岁之间)的预处理猕猴 fMRI 数据集。这些数据是在轻度异氟醚麻醉下采集的。这是之前一些作者在另一篇论文中使用过的相同数据集;为了报告的一致性,我们使用了相同的措辞。关于数据采集和处理的完整描述见参考文献。所有手术和实验程序均获得了加拿大安大略省西部大学动物护理委员会的动物使用小组的批准,并符合加拿大动物护理委员会的指导方针。
简而言之,在扫描前,动物被轻度麻醉,并使用1.0–1.5% 的异氟醚维持麻醉。扫描在 7T Siemens MAGNETOM 头部扫描仪上进行,参数如下:结构 MRI:序列 = 3D MPRAGE T1 加权;切片数 = 128;体素大小 = 0.5 毫米各向同性;10 分钟(600 个体积)静息态 fMRI:序列 = 2D 多带 EPI;TR = 1000 毫秒;切片数 = 42;体素大小 = 1.0×1.0×1.1 毫米³。如原始出版物所述,使用 FSL 的 FEAT 工具箱对 fMRI 数据进行预处理,包括运动校正、高通滤波、配准、归一化和空间平滑(半高全宽 = 2 毫米)。fMRI 数据中的运动很小,所有动物和所有扫描的平均帧间位移为 0.015 毫米(范围 = 0.011–0.019 毫米)。使用“功能性神经影像分析”3dDeconvolve 函数对全局白质和脑脊液信号进行线性回归。全局平均信号未进行回归。然后,从每个静息态 fMRI 扫描的区域图划分中提取每个感兴趣区的 fMRI 信号。
来自人类连接组计划的人类结构连接组
我们使用了人类连接组计划(HCP)900 名参与者数据发布的 100 名无关参与者的弥散 MRI 数据(54 名女性和 46 名男性;平均年龄 = 29.1±3.7 岁)。所有 HCP 扫描协议均获得了圣路易斯华盛顿大学当地机构审查委员会的批准。弥散加权成像(DWI)采集协议的详细内容在其他地方有详细描述。弥散 MRI 扫描在 Siemens 3T Skyra 扫描仪上进行,使用 2D 自旋回波单次多带 EPI 序列,多带因子为 3,采用单极梯度脉冲。空间分辨率为 1.25 毫米各向同性(TR = 5500 毫秒;TE = 89.50 毫秒)。b 值为 1000、2000 和 3000 s/mm²。每个壳层的弥散采样方向总数为 90、90 和 90,外加 6 个 B0 图像。我们使用了在 DSI Studio 兼容格式下提供的数据版本。
我们采用先前报道的程序从DWI 数据重建人类连接组。对 HCP 数据进行了最小预处理,以纠正涡流和易感性伪影。然后,使用 DSI Studio(https://dsi-studio.labsolver.org/)中实现的 q 空间微分重建(QSDR)对 DWI 数据进行重建。QSDR 在标准空间中计算扩散水密度的方向分布,以保持可扩散自旋并保留纤维几何的连续性,用于纤维追踪。QSDR 首先在原生空间重建 DWI 图像,并在每个体素中计算定量各向异性。这些定量各向异性值用于将大脑变形到蒙特利尔神经学研究所空间中的模板定量各向异性体积,使用统计参数映射软件中实现的非线性配准算法。使用了 2.5 的弥散采样长度比,输出分辨率为 1 毫米。然后,使用修改后的 FACT 算法对重建数据进行确定性纤维追踪,参数如下:角度截止 = 55°;步长 = 1.0 毫米;最小长度 = 10 毫米;最大长度 = 400 毫米;自旋密度函数平滑 = 0;以及由脑脊液中的 DWI 信号确定的定量各向异性阈值。生成的每条纤维自动筛选其终止位置。通过将 DSI Studio 的默认各向异性阈值(0.6 Otsu 阈值)应用于自旋分布函数的各向异性值,创建了白质掩模。该掩模用于消除在白质区域内提前终止的纤维。对每个个体进行确定性纤维追踪,直到重建了 1000000 条纤维。
对于每个个体,通过在Schaefer 皮层图谱中每对区域 i 和 j 之间画一条边来重建其结构连接组,如果存在连接相应大脑区域的白质纤维,则画一条边。边权重被量化为连接每对区域的纤维数量,按 ROI 距离和大小归一化。
然后,通过Betzel 及其同事的距离依赖程序在参与者之间获得一个群体共识矩阵 A,以减轻对个体参与者结构连接组重建不一致性的担忧。这种方法旨在保留个体参与者连接组的边密度以及跨半球和内半球边长分布的普遍性和长度分布,并旨在产生一个具有代表性的连接组。该程序产生一个二进制共识网络,指示要保留哪些边。最终边密度为 27%。然后,每个非零边的权重被计算为跨参与者相应非零边的平均值。
典型脑图
为了将麻醉诱导的可识别性区域模式变化置于上下文中,我们使用neuromaps(https://netneurolab.github.io/neuromaps/)从文献中获取相关脑图。我们获取并划分了来自参考文献的感官 - 关联原型轴图、猕猴与人类之间皮层扩张的图以及个体间功能连接变异性的图。
人类加速基因
人类加速基因是指与人类基因组中所谓的“人类加速区域”相关的基因。这些区域是通过将人类基因组与黑猩猩(Pan troglodytes)基因组进行比较而识别出的一组位点,这些位点在人类谱系中显示出加速的分化。在这些人类加速基因中,与大脑功能和发育相关的基因被称为 HAR-大脑基因。HAR-大脑基因的平均区域表达图谱是通过以下步骤获得的:首先,从参考文献中获取 415 个 HAR-大脑基因的列表。然后,使用 abagen 工具箱(https://abagen.readthedocs.io/),按照 abagen 的默认处理流程,并在同源皮层区域之间镜像数据,以确保覆盖左右半球(左侧数据来自 6 名供体,右侧数据来自 2 名供体),获取 Schaefer 图谱中 200 个皮层区域的区域基因表达。在皮层表面上,使用 2 毫米的距离阈值评估样本之间的距离。使用抗异常值的 S 形归一化方法对基因表达数据进行皮层归一化。在最终的 15,633 个基因中,有 392 个属于 Wei 等人列出的 HAR-大脑基因。最后,通过计算这 392 个基因的区域平均归一化基因表达,获得了 HAR-大脑基因的区域平均表达图。
人类fMRI 的信噪比图
为了量化人类大脑中fMRI 信号的区域信噪比,我们使用了参考文献 36 提供的原始图谱。简而言之,Shafiei 等人计算了 HCP 中 n=201 名受试者的每个区域 BOLD 时间序列的均值与其标准差的比值,并随后对个体进行平均,以获得人类大脑的代表性信噪比图。更多细节请参考文献 36。
大脑指纹识别
大脑指纹识别是指使用大脑衍生的指标(此处为静息态fMRI 获得的功能连接,FC)来区分个体,类似于指纹上的纹路可用于识别身份。这要求大脑指纹(就像传统指纹一样)在不同人之间有所不同(以避免混淆不同个体),但在同一个人内保持一致(以追踪身份)。
设A 为可识别性矩阵(即个体测试和重测扫描之间的相似性矩阵,为非对称的方阵),其大小为 S×S(S 为数据集中个体的数量)。A 的每个条目是通过计算相应个体的分区 FC 的向量化矩阵之间的相关性获得的。设 Iself = 〈Aii〉表示 A 的主对角线元素的平均值,这些元素是相同个体扫描之间的皮尔逊相关值。从现在开始,我们将这个量称为自我可识别性或 Iself。同样,设 Iothers = 〈Aij〉定义为矩阵 A 的非对角线元素的平均值(即不同个体 i 和 j 的扫描之间的相关性)。然后,我们将样本的差异可识别性(Idiff)定义为这两个项之间的差异:Idiff = (Iself - Iothers),它量化了参与者内平均 FC 相似性与参与者间平均 FC 相似性之间的差异。Idiff 的值越高,个体指纹在整个群体中的总体可识别性就越高。
我们还可以通过使用ICC(组内相关系数)来量化个体的边缘可识别性。ICC 是统计学中广泛使用的度量,最常用于评估不同组(或评分者或评判者)之间的单位(或评分或得分)的一致性百分比。它描述了同一组中的单位彼此之间的相似程度。评分之间的一致性越强,其 ICC 值就越高(与皮尔逊相关性一样,ICC 的范围在 -1 到 +1 之间)。我们使用 ICC 来量化一个边缘(两个脑区之间的 FC 值)的连接值在参与者内和参与者间分离的程度。换句话说,ICC 越高,连接边缘的可识别性就越高。我们使用 GitHub 上提供的代码(https://github.com/eamico/MEG_fingerprints)实现了 ICC 分析,如参考文献 10 所述。实际上,ICC 是通过样本均方的差异来估计的:ICC = (MSB - MSW) / (MSB + (k - 1)MSW),其中 MSB 是组均值与总均值的变异性,MSW 是个体得分与其各自组均值的变异性,k 是样本大小。其原理是,如果组成员身份无关紧要,则组内变异性应与组间变异性相同(即 MSB = MSW,ICC 等于 0)。然而,如果组内变异性大于组间变异性,则会观察到负 ICC。在我们的主要分析中,我们包含了所有 ICC 值。然而,我们还通过仅使用置信区间不包括零的 ICC 值来复制我们的结果。
神经合成的元分析认知匹配
从神经合成(Neurosynth)获得的连续测量值反映了体素与认知类别的关联。神经合成是一种自动化的基于术语的元分析工具,通过搜索在论文中与 fMRI 体素坐标一起系统提到的高频关键词(如“疼痛”和“注意力”等术语),综合了超过 14,000 篇已发表的 fMRI 研究的结果。这种关联强度的度量是,如果在给定体素处观察到激活,则在功能神经影像学研究中报告给定术语的倾向。需要注意的是,神经合成不区分与感兴趣术语相关的激活或去激活区域,也不区分激活的程度,而只是某些脑区是否经常与某些词汇一起被提及。尽管神经合成引擎中记录了 1,000 多个术语,但为了专注于认知功能,我们将术语限制为认知和行为术语。为了避免引入选择偏差,我们选择以数据驱动的方式选择术语,而不是手动选择术语。因此,我们从认知图谱(Cognitive Atlas)中选择术语,这是一个包含神经认知术语的公共本体。这种方法总共涉及 123 个术语,范围从宽泛的术语(如注意力和情绪)到具体的认知过程(如视觉注意力和情景记忆)、行为(如饮食和睡眠)以及情绪状态(如恐惧和焦虑)(注意,神经合成的 123 个基于术语的元分析图并不明确排除患者研究)。认知图谱的细分之前已与神经合成结合使用过,因此我们选择相同的方法,以使我们的结果与先前的报告具有可比性。本分析中包含的术语完整列表见补充表 1。神经合成报告的概率度量可以被解释为区域活动波动与心理过程相关的定量表示。与静息态 BOLD 数据一样,体素化的神经合成图被划分为 200 个皮层区域,根据 Schaefer 图谱(或在使用 Desikan-Killiany 图谱进行复制时为 68 个区域,在包含皮层下区域的复制中为 232 个皮层和皮层下区域)。进行认知匹配的代码可在 https://github.com/netneurolab/luppi-cognitive-matching 获取。
对于每个个体,他们在每个时间点的分区BOLD 信号与每个神经合成图进行了空间相关性分析,每个神经合成图每个 BOLD 体积产生一个相关性值。我们将此操作称为认知匹配。对于每个体积,认知匹配的质量被量化为所有地图中的最高(正)相关性值。这些值随后在所有体积上进行平均,以获得每个条件每个参与者的单一值。作为替代,除了使用最高的正相关性外,我们还考虑了所有地图的相关性幅度的平均值(无论符号),并如上所述在体积上进行平均。
我们还分别对与参考文献34中的典型轴呈正空间相关性的Neurosynth脑图(即主要在跨模态联合皮层中呈现正值的脑图)和与典型轴呈负相关的脑图(主要包含单模态[感觉]皮层中的激活)进行了认知匹配。
从BrainMap获取的替代性元分析匹配。与Neurosynth作为自动化工具不同,BrainMap是一个专家策划的数据库。它包含了数千项已发表的神经影像学研究中显著激活的大脑坐标49,50。因此,Neurosynth术语和BrainMap行为领域存在显著差异。在此,我们使用了与66个独特行为领域相关的Desikan–Killiany解剖图谱中的脑图,这些脑图来源于8703项实验。本次分析中包含的BrainMap术语完整列表见补充表2。排除了在非健康受试者中进行的实验,以及没有明确定义行为领域的实验。
使用PCA进行低维表示
我们使用PCA(主成分分析)来获得人类和猕猴功能连接(FC)在一个共同空间中的低维表示。PCA通过将数据重新表示为线性不相关(正交)的变量(称为主成分),这些变量是从数据本身中提取的,作为最大变化的轴125。因此,PCA被广泛用于降维和高维数据的可视化,因为它在保留尽可能多的原始数据变化的同时,提供了数据的低维表示。
为了获得联合PCA空间,我们首先将人类数据集中每个个体的每次扫描(清醒、恢复以及不同麻醉水平)的功能连接矩阵的上三角部分向量化。对于清醒和麻醉状态下的猕猴数据集,我们也进行了相同的处理。然后,我们将这些向量化的功能连接模式连接起来,形成矩阵M的单独列,矩阵M的行对应于功能连接的边。然后,我们将PCA算法应用于矩阵M,以提取最大变化的线性正交主成分(按解释方差的降序排列)。该算法还提供了将矩阵M的每一列与每个提取的主成分相关联的权重。我们使用这些权重将矩阵M的每一列(对应于功能连接模式)表示为一个点,位于主成分空间中。在我们的主要分析中,我们使用了前两个主成分来定义低维空间,但我们也使用前三个主成分复制了我们的结果。
统计分析
通过置换t检验(配对样本和双侧)来确定不同条件(在此为麻醉水平)之间的差异的统计显著性,置换次数为10,000次。使用置换检验避免了假设数据分布的正态性(这并未正式检验)。所有检验均为双侧,α值为0.05。效应量使用Hedge的标准化平均差异量度g来估计,其解释方式与Cohen的d相同,但更适合小样本量。使用MATLAB的效应量工具箱(https://github.com/hhentschke/measures-of-effect-size-toolbox)。将麻醉条件分别与清醒和恢复状态进行比较,并使用假发现率校正分别对这两种情况进行多重比较的假阳性率控制。通过Spearman相关系数量化脑图之间的空间相关性,并通过与保留空间自相关的空地图的空分布进行非参数比较来评估其统计显著性。
优势分析。为了综合考虑所有区域相关性,并评估它们各自的贡献,我们以四个典型脑图为预测变量,以麻醉诱导的ICC变化的区域图为目标,进行了优势分析。优势分析旨在确定多个线性回归模型中每个独立变量的相对贡献(即每个预测变量对整体拟合优度[调整后的R²]的主导性)。这是通过在每种预测变量组合上拟合相同的回归模型(对于有p个预测变量的模型,有2^p - 1个子模型)来实现的。总优势定义为在所有2^p - 1个子模型中,添加一个感兴趣的单预测变量时相对增加的R²的平均值。所有输入变量的优势之和等于完整模型的总调整R²,这使得相对重要性的百分比成为一种直观的方法,可以将总效应量分配给预测变量。因此,与其他评估预测变量重要性的方法(如基于回归系数或单变量相关性的方法)不同,优势分析考虑了预测变量之间的相互作用,并且是可解释的。我们使用非参数置换检验(单侧)来确定优势分析模型的统计显著性,通过将经验解释的方差与从重复多次回归中获得的R²的空分布进行比较,这些回归使用了保留空间自相关的空地图124,129。
3. 结果
本文中,我们考虑了从n = 15 名健康志愿者在静息状态下获得的功能性磁共振成像(fMRI)数据,分别在基线(清醒状态)和吸入麻醉剂七氟烷(sevoflurane)诱导的行为反应丧失后进行记录。具体记录阶段包括脑电图(EEG)爆发抑制阶段、3%和 2%七氟烷浓度,以及术后麻醉恢复阶段。我们还在一个独立的数据集中验证了我们的结果,该数据集包含n = 16 名健康志愿者在静息状态下接受 fMRI 扫描的数据,分别在清醒状态、丙泊酚(propofol)诱导的行为反应丧失期间以及术后恢复阶段。
麻醉状态下大脑识别度降低
首先,我们测试了以下假设:麻醉会消除每个个体特异性的自发神经活动模式,使得个体间的功能连接(FC)模式更难区分。具体而言,我们将每位个体在清醒状态下的 FC 与其在术后恢复或麻醉状态下的 FC 进行相关性分析。这生成了一个识别度矩阵,其中矩阵的行和列代表个体,每个矩阵元素代表两个个体之间的功能连接相似性(相关性)(图 1a)。
成功的脑指纹识别需要满足两个条件。第一是持续性:个体的FC 需要随着时间保持一致,才能用于识别个体。第二是多样性:不同个体的 FC 需要彼此不同,以避免混淆。如果所有 FC 模式都相同,即使 FC 具有持续性,识别度也将较低。相反,如果 FC 随时间变化较大,即使不同个体的 FC 配置具有多样性,识别度也将较低。我们通过同一被试两次扫描之间的自我-自我相关性来量化持续性(即第一次和第二次扫描时的 FC 相关性)。我们通过平均自我-他人相关性来量化多样性(即某被试第一次扫描的 FC 与所有其他被试第二次扫描的 FC 之间的平均相关性)。最后,差异识别度定义为自我-自我相关性(持续性)与平均自我-他人相关性(缺乏多样性)之间的差异。
我们观察到,当比较同一被试两次清醒状态下的扫描时,很容易区分自我与他人。这种识别度体现在矩阵对角线上清晰的自我-自我相似性模式,与非对角线上的自我-他人相似性模式形成鲜明对比(图 1a)。这与之前使用测试-重复扫描的功能连接组指纹识别研究一致10。相比之下,当清醒状态下的扫描与麻醉状态下的扫描进行比较时,对角线几乎无法辨认,表明个体识别度较低(图 1b)。确实,清醒与麻醉大脑之间的自我-自我相关性显著低于清醒与恢复期的自我相似性(图 1c)。同样,差异识别度(自我-自我相关性与个体在时间 tx 的扫描与其他所有个体在时间 ty 的扫描的平均相关性之间的差异)在考虑麻醉大脑时显著降低(图 1d)。这些结果共同表明,麻醉诱导的行为反应丧失表现为个体功能连接组独特性的降低。在七氟烷麻醉的不同深度下也观察到了类似的结果(扩展数据图 1)。在本小节的其余部分,我们进一步探讨了功能连接的解剖组织如何贡献于这种独特性变化。
我们使用类内相关系数(ICC)来量化边的可识别性,该系数描述了同一组内元素之间的相似程度。在这种情况下,我们为每条边(两个大脑区域之间的功能连接值)获得了一个ICC值,这表明该边的权重在区分个体内部和个体之间方面的效果如何。因此,边的ICC值越高,其可识别性就越高。
清醒-恢复状态与清醒-麻醉状态之间边的可识别性差异表明,麻醉对每条功能边的可识别性有多大影响。换句话说,ICC差异矩阵反映了每条边在意识状态下获得的可识别性增益(即使用恢复扫描而不是麻醉扫描时,区分个体的额外能力有多大)。
我们发现,七氟醚麻醉几乎降低了所有边对可识别性的贡献。这种模式既不是均匀的,也不是随机的。相反,麻醉诱导的每条功能连接的可识别性变化与其在清醒状态下对可识别性的贡献成比例。此外,在麻醉状态下,能够可靠识别个体的FC边(即其置信区间不包含零)的比例大幅减少。
值得注意的是,受麻醉影响最大的功能连接是连接两个跨模态皮层区域的那些。这是因为跨模态皮层区域在清醒静息大脑中对可识别性的贡献最大。换句话说,一条功能边在清醒个体中对可识别性的贡献越大,它受麻醉的影响就越大。跨模态皮层内的功能连接特别容易受到这种干扰的影响,失去其独特性,变得更加相似。
接下来,我们定位了区域性的可识别性变化,量化为每个区域边的边可识别性变化的平均值。可识别性下降最大的区域是默认模式网络和前顶网络以及更广泛的跨模态皮层,而单一模态(体感运动和视觉)皮层受影响最小。这种单一模态-跨模态的区别通过与大脑原型感觉-联想轴的空间相关性得到了证实。
鉴于跨模态皮层的FC在清醒状态下表现出最大的个体间变异性,我们试图测试麻醉诱导的可识别性下降是否优先针对这些区域。事实上,可识别性下降的模式与Mueller及其同事开发的个体间变异性的地图相关。
在观察到区域性可识别性变化在个体间FC差异最大的地方更为显著后,我们进一步研究了一种更普遍的现象。麻醉诱导的区域性可识别性变化是否不仅反映了个体之间的独特性,更普遍地反映了物种之间的独特性?虽然我们在后续部分更详细地探讨了这一点,但这里我们表明,区域性可识别性变化与人类和非人灵长类动物之间系统发育皮层分化的分子和形态标记相关。具体来说,麻醉诱导的区域性可识别性变化与猕猴和人类之间进化扩张的皮层地图在空间上相关,进化上较新的区域观察到的独特性变化更大。同样,我们观察到区域性可识别性变化与所谓人类加速区域(HAR)的人类基因组中与大脑功能和发育相关的基因的区域平均表达之间存在显著的空间相关性。这些基因与在人类谱系中与黑猩猩相比表现出加速分化的位点相关,因此表明相应区域的进化相关变化。换句话说,独特性变化较大的大脑区域也表现出更多的人类加速基因的表达。总的来说,麻醉选择性地降低了在个体之间以及人类和非人灵长类动物之间最具独特性的大脑区域的可识别性。
相关分析不能确定区域性可识别性变化的因果决定因素。然而,多变量分析可以帮助提供超出多个单独相关性所能提供的见解。具体来说,我们可以使用优势分析来评估不同的典型大脑地图在预测区域性可识别性变化分布中的相对重要性。优势分析将模型的拟合度分配给预测变量,以便可以评估和比较每个预测变量的贡献,反映所有预测变量共同解释的方差中可以归因于每个预测变量的比例。
个体间变异性、原型轴、进化扩张和HAR-大脑基因表达的地图共同解释了七氟醚诱导的区域性可识别性变化地图中51%的方差,这显著多于保留空间自相关的空地图所解释的方差。原型轴和个体间变异性是最重要的预测变量,分别解释了46.5%和36.0%的方差,而HAR-大脑基因表达和进化扩张分别解释了12.4%和5.0%的方差。
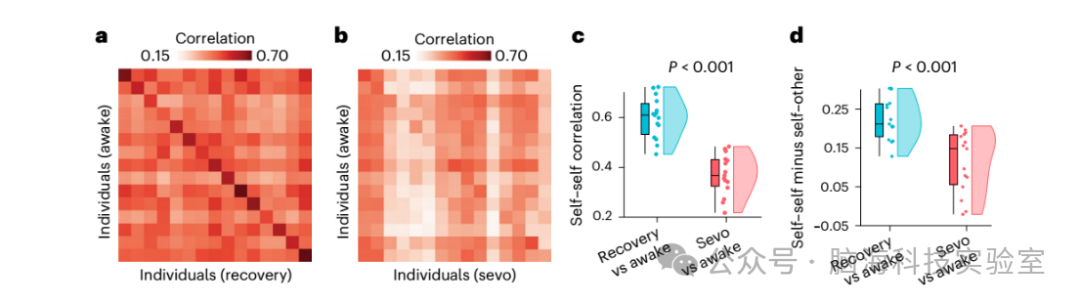
图 1 | 七氟醚麻醉下个体功能连接组的可识别性降低
a、清醒与麻醉后恢复状态之间的可识别性矩阵。成功识别率为 93%。
b、清醒与七氟醚(sevo)麻醉状态之间的可识别性矩阵。
在a 和 b 中,对角线元素表示自身-自身相似性(功能连接模式的相关性),而非对角线元素表示自身-他人相似性。
c、两种意识状态之间的自身-自身相似性显著高于清醒与麻醉状态之间(清醒与恢复状态:均值 = 0.60,标准差 = 0.08;清醒与麻醉状态:均值 = 0.37,标准差 = 0.08;t(14) = 8.36;P < 0.001;效应量(Hedge’s g)= 2.71;置信区间(CI)= [2.22, 3.69];双尾 t 检验)。
d、差异可识别性(每个个体自身-自身相关性与平均自身-他人相关性之间的差异)在两种意识状态之间显著高于清醒与麻醉状态之间(清醒与恢复状态:均值 = 0.19,标准差 = 0.06;清醒-麻醉状态:均值 = 0.07,标准差 = 0.09;t(14) = 4.79;P < 0.001;效应量(Hedge’s g)= 1.57;置信区间(CI)= [1.08, 2.35];双尾 t 检验)。对于 c 和 d 中的箱线图,中间线表示中位数值,箱子的边界表示第 25 和第 75 百分位数,须表示 1.5 倍的四分位数范围,极端值以单独的圆圈显示(n = 15 名人类志愿者)。提供原始数据。
与典型认知图谱对应度降低
一种研究意识病理或药理学干扰的有力方法是确定是否可以从神经活动中算法推断(或解码)认知过程。例如,患有意识障碍的患者可能会被要求在扫描仪中想象打网球,以确定运动相关区域是否能可靠地响应指令而激活,尽管患者可能无法表现出明显的外显行为响应23,24。虽然这种方法通常在存在明确任务或刺激(例如观看电影)的情况下进行,但在这里,我们试图通过基于数千项神经影像实验的元分析图谱15,40的综合性评估,更普遍地探究是否可以从自发神经活动中辨别出潜在的认知过程。
具体而言,在不同麻醉水平下,我们评估了每位个体在每个时间点的神经活动图谱与从>14,000项神经影像实验中通过元分析整合得到的123个典型脑图谱之间的对应程度(图3a;方法部分详细说明了如何选择Neurosynth脑图谱,补充表1列出了所有包含的术语)。为了简化表述,以下我们将基于活动的反向推断方法称为认知匹配(cognitive matching)。通过对扫描持续时间的平均处理,我们为每个个体和每个条件提供了一个整体的认知匹配质量指标。
我们发现,随着麻醉深度增加(七氟烷浓度升高),认知匹配的质量下降:在更深度的麻醉水平下,大脑活动与Neurosynth元分析脑图谱之间的最佳空间相关性(在整个扫描持续时间范围内取平均)较低(图3b)。这一趋势在恢复行为反应能力后得到逆转(图3b)。麻醉引起的认知匹配质量下降在那些加载到大脑典型轴更高阶(跨模式/关联)端的Neurosynth图谱(例如认知控制或情绪调节)中比在加载到单模态/感觉端的图谱中更为显著。换句话说,麻醉减弱了自发脑活动反映文献中认知模式的程度,尤其是更高阶的认知操作,这可能解释了为什么个体独特性会因麻醉而受到抑制。
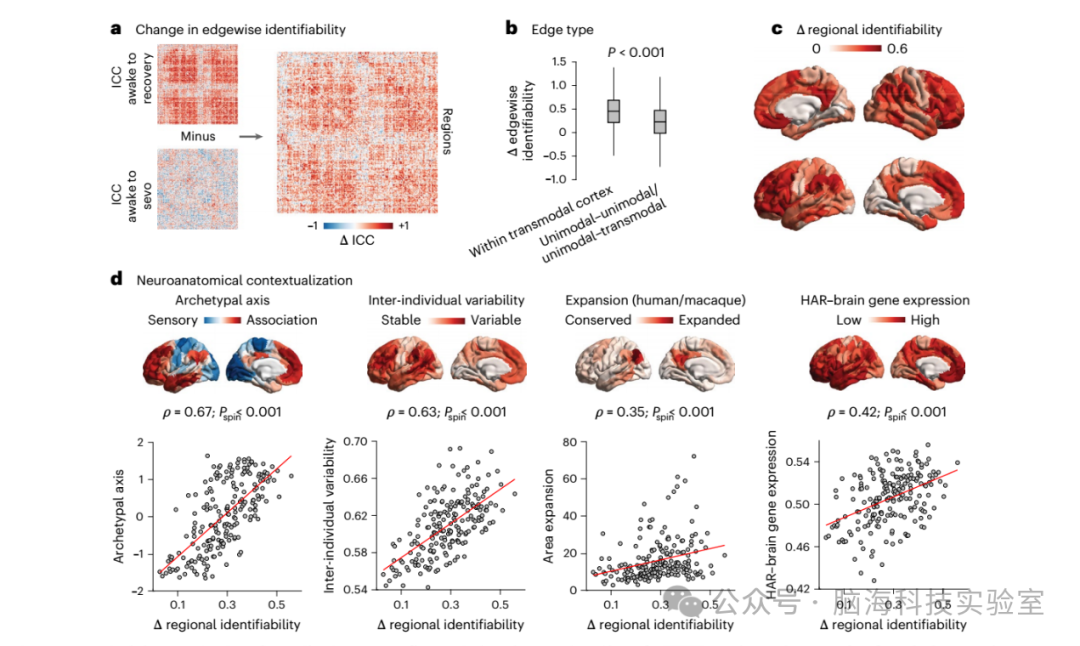
图2 | 七氟醚诱导的功能可识别性丧失的解剖学特征。
a、清醒与恢复状态以及清醒与七氟醚状态之间的边 ICC 差异。
b、麻醉诱导的跨模态皮层内功能连接的 ICC 丧失显著大于涉及单一模态区域的那些功能连接。对于跨模态皮层内的边,均值 = 0.44,标准差 = 0.34;对于其他边,均值 = 0.22,标准差 = 0.35;t(39,998) = 55.93;P < 0.001;效应量(Hedge’s g)= 0.63;置信区间(CI)= [0.60, 0.65];双尾 t 检验)。ICC 的范围在 -1 到 +1 之间。n = 5,580 条跨模态皮层内的边和 n = 34,420 条单一模态 - 跨模态和单一模态 - 单一模态边。在箱线图中,中间线表示中位数值,箱子的边界表示第 25 和第 75 百分位数,须表示 1.5 倍的四分位数范围。
c、麻醉诱导的 ICC 丧失的区域分布,投影到皮层表面。
d、麻醉诱导的区域 ICC 丧失与大脑皮层组织的原型感觉 - 联想轴(Spearman’s ρ = 0.67;pspin < 0.001;n = 200 个区域)、FC 的个体间变异性(Spearman’s ρ = 0.63;pspin < 0.001;n = 200 个区域)、猕猴和人脑之间的皮层扩张(Spearman’s ρ = 0.35;pspin < 0.001;n = 200 个区域)以及与大脑功能和发育相关的人类加速区域(即 HAR - 大脑基因)的区域表达(Spearman’s ρ = 0.42;pspin < 0.001;n = 200 个区域)显著空间一致。对于每个大脑图,颜色条的值范围显示在散点图的 y 轴上。提供原始数据。
麻醉使人类功能连接(FC)更接近猕猴功能连接
最后,我们研究了麻醉是否改变了人类大脑功能连接(FC)与猕猴大脑功能连接之间的相似性。我们使用了来自10只清醒状态下的猕猴的fMRI数据,这些数据的处理方式与人类数据相同。此外,我们还使用了来自The Virtual Brain项目的独立处理的fMRI数据,这些数据来自9只用1.0%至1.5%异氟醚轻度麻醉的成年猕猴。为了能够在两个物种之间进行比较,我们根据Kötter和Wanke的区域映射划分对猕猴和人类数据进行了划分,这种划分方法旨在促进跨物种比较,并且最近已被翻译用于猕猴和人类大脑之间,使得每个皮层区域都能在两个物种之间找到其解剖学上的同源区域。
随后,我们使用主成分分析(PCA)将人类和猕猴的所有连接模式投影到一个共同的低维空间中。PCA广泛用于降维和高维数据的可视化,因为它在保留尽可能多的原始数据变异性的同时,提供了数据的低维表示。这种方法使我们能够将每个个体的功能连接重新表示为二维平面上的一个点,其中每个维度对应于数据中的一个主要变异轴。我们随后可以追踪个体功能连接在这个低维空间中的位置如何随麻醉而变化。
我们清楚地观察到,每种状态(清醒、恢复以及不同水平的七氟醚麻醉)倾向于占据由前两个主成分所张成的空间中的不同区域(图4a)。由于主成分1(PC1)似乎主要反映了我们的一组猕猴数据与其他所有数据之间的差异,我们主要关注主成分2(PC2),它捕捉了人类状态之间的差异。很明显,随着七氟醚剂量的增加,我们的人类受试者在麻醉前清醒状态下的初始位置(图4a、b中的红圈)沿着PC2逐渐远离——然后在麻醉后恢复清醒时又回到最接近初始清醒位置(图4a、b中的紫圈)。我们正式通过计算PC2上的欧几里得距离来量化这种偏移:我们发现,与清醒人类数据在PC2上距离最小的人类状态是恢复状态,而更深水平的七氟醚麻醉则对应于沿着PC2远离清醒状态的更远距离(图4c)。与此同时,我们观察到,随着人类麻醉功能连接模式从清醒状态移开,它们也减少了与两个猕猴功能连接数据集在PC2上的距离(图4d、e)——其中人类麻醉的最深水平(爆发抑制)既是最远离清醒人类功能连接的(图4c),也是最接近猕猴功能连接的(图4d、e)。在同时考虑前两个主成分的空间或使用余弦距离代替欧几里得距离时,也观察到类似的结果。
总的来说,这种低维表示突出了麻醉如何使人类大脑的功能连接从清醒状态移开,而更接近非人灵长类动物大脑的功能连接:麻醉状态下人类功能连接与猕猴功能连接之间的距离小于清醒状态下人类功能连接与猕猴功能连接之间的距离。这种现象在麻醉后恢复时被逆转,此时人类功能连接回到接近基线时所占据的原始位置。这些结果补充了我们之前的观察,即麻醉诱导的区域可识别性降低在人类大脑中最具人类特异性的区域最为明显(图2d)。
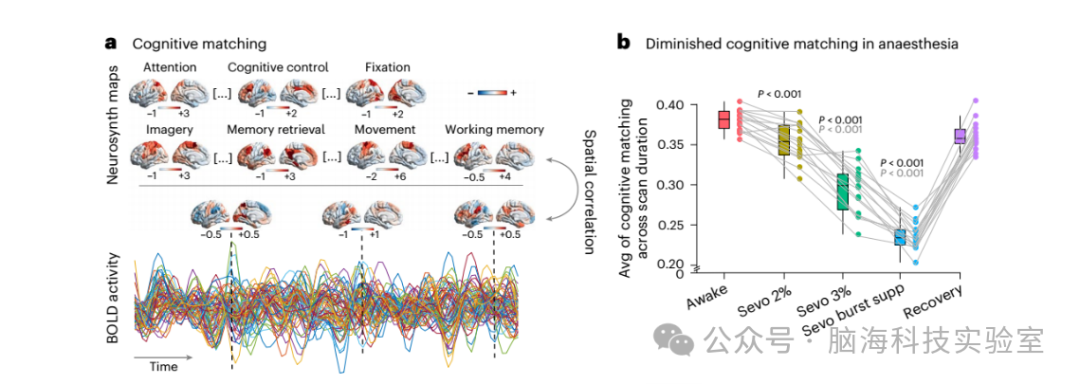
图3 | 麻醉下大脑活动与典型元分析模式的认知匹配降低,并在恢复时恢复。
a、在每个时间点,认知匹配得分被计算为大脑活动与 123 个 Neurosynth 元分析图谱之间的最佳空间相关性。对于每个参与者,通过在每个条件下对整个扫描期间的认知匹配得分进行平均,获得了一个认知匹配质量的总体指标。
b、y 轴表示认知匹配得分的时间平均值(n = 15 名人类志愿者)。在箱线图中,中间线表示中位数值,箱子的边界表示第 25 和第 75 百分位数,须表示 1.5 倍的四分位数范围,极端值以单独的圆圈显示。P 值通过重复测量 t 检验(双侧)获得,并通过假发现率校正多重比较。与清醒数据的比较显著性值以黑色显示,与恢复数据的比较显著性值以灰色显示。完整的统计报告见补充数据 1。提供原始数据。Avg,平均值;supp,抑制。
复制性、稳健性和敏感性
麻醉降低了人类功能连接组在相同状态下的可识别度。我们发现,在清醒状态与麻醉状态之间的可识别度显著低于清醒状态与恢复状态之间的可识别度(图1d)。这意味着,给定一名清醒个体的静息状态fMRI扫描,与该个体的另一清醒扫描相比,区分其麻醉扫描与其他个体的麻醉扫描会更加困难。这些结果是否仅反映了麻醉状态与清醒状态之间的差异大于恢复状态与清醒状态之间的差异?从逻辑上讲,基线(清醒状态)和恢复状态应比基线和麻醉状态更为相似,因为基线和恢复在某种意义上是同一种大脑状态(即清醒),而基线与麻醉则是完全不同的状态。事实上,正如我们对自我-自我相关性的分析所示,清醒与麻醉之间的自我相似性显著低于清醒与恢复之间的自我相似性(图1c)。
然而,自我-自我相关性降低本身并不必然保证可识别度降低。理论上,如果自我-他人相关性降低的程度与自我-自我相关性降低的程度相同甚至更大,可识别度可能会保持不变甚至增加。
为了实证证明这些结果并非仅仅由于在同一状态下比较相关性(即在清醒或麻醉状态之间进行比较),我们利用了七氟烷数据中包含多个麻醉状态下扫描的优势。这使我们能够比较两种清醒扫描(基线和恢复)与两种麻醉扫描(例如,2%和3%七氟烷浓度,或3%七氟烷浓度与爆发抑制状态)的相关性。通过这种方法,我们避免了仅比较同一扫描 session 的两半数据,因为后者涉及将同一名被试的扫描与其他被试的扫描进行比较,从而混淆了个体身份和扫描身份。
在比较清醒-恢复相似性与2%和3%七氟烷浓度相似性或3%七氟烷浓度与爆发抑制相似性时,我们发现与主要分析完全相同的结果模式。自我-自我相似性不仅在清醒与麻醉之间显著降低(如前所述),也在两种麻醉扫描之间显著降低。同样,在麻醉状态下,可识别度也降低,并呈现出清晰的单模态-跨模态皮层模式。具体而言,我们发现随着麻醉深度增加,自我相似性和自我-自我与自我-他人相关性之间的差异(即可识别度)逐渐降低,且两种分布的重叠程度不断增加(扩展数据图2)。这一模式与Colenbier等人最近的研究结果相反,他们发现严格控制的认知任务会增加自我-他人相似性,但自我-自我相似性的增加更为显著,从而导致整体可识别度的增加。
总体而言,由于这些结果是从同一状态下进行的比较(清醒-恢复(均为清醒状态)与麻醉-麻醉),我们可以排除可识别度的变化仅仅是大脑状态差异的反映的可能性。
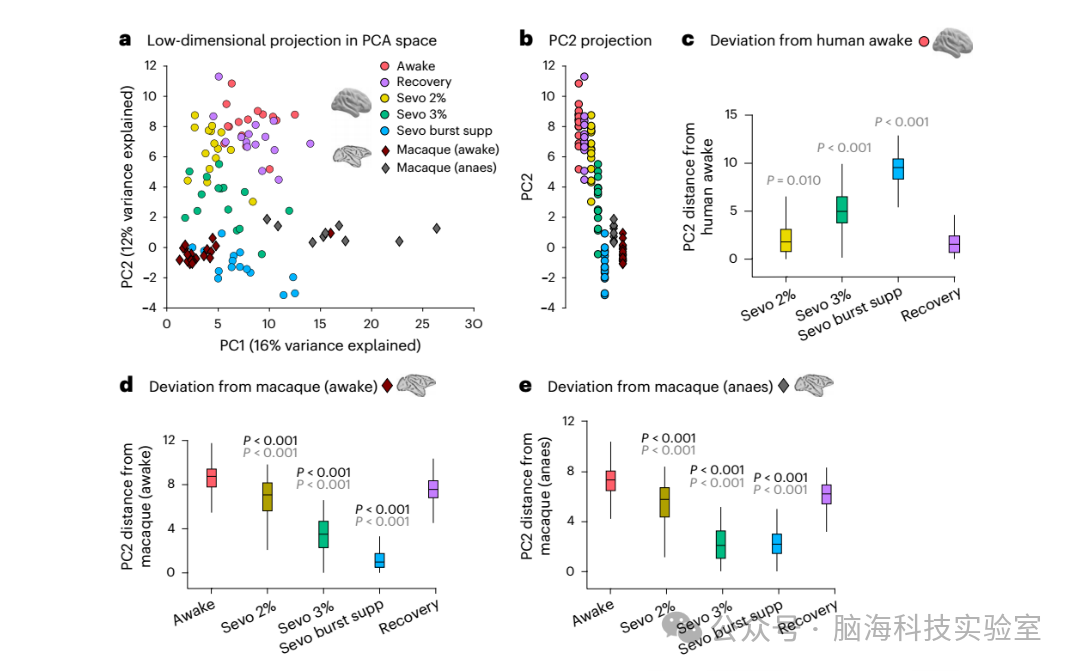
图4 | 麻醉使人类功能连接(FC)从清醒状态向猕猴的功能连接靠近。
a,人类和猕猴功能连接在主成分分析(PCA)的前两个主轴变化空间中的低维投影。每个圆圈代表一个人类的功能连接,颜色反映其状态(清醒、恢复或不同剂量的七氟醚)。每个菱形代表一只猕猴的功能连接,颜色代表数据集(清醒或麻醉(anaes))。b,将a中的数据投影到主成分2(PC2)上。c,人类清醒状态下功能连接模式在人类数据集中沿PC2的欧几里得距离分布,如b所示。每个箱线图中有225(15×15)对数据点。d,人类数据与清醒猕猴功能连接模式沿PC2的欧几里得距离分布。每个箱线图中有285(15×19)对数据点。e,人类数据与麻醉猕猴功能连接模式沿PC2的欧几里得距离分布。每个箱线图中有135(15×9)对数据点。在c-e中,显著性值与人类恢复组(灰色)或人类清醒组(黑色)对比。对于c-e中的箱线图,中间的线表示中位数,箱子的边界表示第25和第75百分位数,须表示1.5倍的四分位间距。P值是通过重复测量t检验(双侧)获得的,并经过多重比较的假发现率校正。完整的统计报告见补充数据2。提供原始数据。
扫描时长的稳健性
除了表明麻醉降低了不同麻醉状态下大脑的功能连接可识别度外,我们还证明了麻醉引起的自我-自我相似性和可识别度的变化并非由扫描时长的限制所致。首先,我们的每次扫描时长相同(见方法部分),因此无需担心扫描时长差异可能带来的混杂效应。其次,这一时长(约10分钟)显然足以实现优异的脑指纹识别,因为在清醒-恢复数据中,所有但有一位个体(93%)在清醒状态下均能被正确识别。这一结果与Van De Ville等人的研究完全一致,他们指出仅需1分钟左右的静息状态fMRI即可通过脑指纹识别实现90%以上的成功识别。第三,我们发现,即使将三种七氟烷条件(2%、3%和爆发抑制)下获取的所有血氧水平依赖(BOLD)信号进行合并(在计算区域间相关性前,将时间序列按顺序拼接),麻醉状态下的可识别度和自我-自我相似性仍显著降低。因此,与脑指纹识别文献一致,我们的结果表明,麻醉引起的自我-自我相似性和可识别度的变化无法归因于扫描时长不足以实现指纹识别。相反,即使通过人为增加用于功能连接估算的时间点数量(将麻醉数据的优势三倍化),这些结果仍然稳健存在。
丙泊酚麻醉的复制性
我们在另一独立数据集中使用静脉麻醉剂丙泊酚复制了主要结果。尽管该数据集的麻醉深度未达到七氟烷实验中所用的深度,但结果与七氟烷麻醉下的观察结果总体一致。自我-自我相似性在丙泊酚诱导的行为反应丧失期间降低(清醒-恢复:平均=0.68,标准差=0.07;清醒-麻醉:平均=0.48,标准差=0.13;t(15)=6.81;P<0.001;效应量(Hedge's g)=1.82;置信区间=[1.32, 2.74]),差异性可识别度同样降低(清醒-恢复:平均=0.20,标准差=0.06;清醒-麻醉:平均=0.08,标准差=0.13;t(15)=4.35;P<0.001;效应量(Hedge's g)=1.14;置信区间=[0.71, 1.77];扩展数据图3)。此外,丙泊酚诱导的可识别度区域性变化在默认模式网络和前额顶叶网络中更为显著,而在躯体感觉运动皮层和视觉皮层中则相对较低,这一变化与感觉-关联轴(Spearman's ρ=0.46;p_spin<0.001;n=200个区域)相关(扩展数据图4)。与七氟烷类似,这一结果在考虑每条边的组内一致性系数(ICC)以及仅包括置信区间不包含零的ICC值时均成立。我们还复制了丙泊酚诱导的区域性可识别度变化与典型个体间变异图谱(Spearman's ρ=0.43;p_spin<0.001;n=200个区域)、进化皮层扩展(Spearman's ρ=0.28;p_spin=0.001;n=200个区域)以及人类加速区域(HAR)脑基因表达(Spearman's ρ=0.30;p_spin<0.001;n=200个区域)之间的相关性(扩展数据图4)。优势分析表明,四个典型图谱解释了丙泊酚诱导的区域性可识别度变化图谱中27%的方差,这一比例显著高于随机空间自相关保留图的预期(P<0.001),其中典型轴和个体间变异性再次是最关键的预测因子。
Neurosynth认知匹配的稳健性
基于Neurosynth元分析引擎的脑活动解码质量结果未显示出丙泊酚麻醉与清醒状态或术后恢复状态之间在大脑活动与元分析图谱之间最大相关性方面的显著差异(扩展数据图5和补充数据1)。然而,如果我们不只考虑最佳相关性,而是考虑大脑活动与所有Neurosynth图谱之间相关性的平均幅度,则在七氟烷和丙泊酚数据集中均发现基线与麻醉之间的显著差异(扩展数据图6和补充数据1)。这一后一种分析可解释为元分析模式再现自发脑活动模式的整体能力。我们还发现,尽管丙泊酚麻醉深度低于七氟烷实验中的主要分析,但它仍沿主成分分析(PCA)获得的低维空间中的PC2将人类功能连接从清醒状态推向猕猴的功能连接。
基于BrainMap的认知匹配复制
我们使用另一种元分析数据库BrainMap49,50中获得的66个独特的行为领域,成功复制了基于Neurosynth元分析引擎的脑活动解码质量结果。与Neurosynth采用数据驱动的自下而上分类方法并通过自动化流程识别特定认知和行为术语相关研究的脑坐标统计关联不同,BrainMap是专家整理的数据库。尽管两个数据库存在差异(例如,BrainMap明确排除了患者研究),但我们仍发现,随着麻醉深度增加,解码质量显著下降,并在恢复行为反应能力后恢复。这一成功复制表明,我们的认知匹配程序不仅对所选术语的具体选择稳健,而且对元分析数据库的选择更为广泛地稳健。
对不同分区方法的稳健性
本研究结果使用了基于清醒状态下fMRI数据的Schaefer功能图谱52。据我们所知,尚未有研究表明Schaefer(或其他任何)功能图谱的分区适宜性会因个体意识状态而变化。事实上,我们和其他研究者已成功将Schaefer图谱应用于先前涉及麻醉、致幻剂和病理意识干扰的研究53,54,包括在意识状态改变下的脑指纹识别47,55。然而,为了证明我们的结果并非严重依赖基于清醒个体的功能分区方法,我们使用另一种基于解剖标志的脑皮层分区方法——Desikan–Killiany图谱56复制了我们的结果。由于其基于解剖标志,Desikan–Killiany图谱的分区适宜性在麻醉状态下不应预期发生变化。类似地,在包含Tian图谱定义的32个皮下区域时也获得了类似结果。具体而言,在皮下结构中,我们观察到双侧苍白球对麻醉诱导的区域性可识别度变化具有特别高的区域贡献度。我们还发现,Neurosynth认知匹配结果对分区方法和皮下区域的纳入具有稳健性。丙泊酚结果也可通过解剖学Desikan–Killiany图谱复制。总体而言,我们清楚地证明,我们的当前结果对分区规模(从68个区域到200个区域)和分区类型(功能或解剖)均具有稳健性。
对头部运动的稳健性
在认知匹配分析中,我们报告了每组对比与对应均帧位移(Framewise Displacement, FD)差异的相关性(补充数据1)。尽管在丙泊酚数据集中观察到相关性,但在主要的七氟烷数据集中,未发现认知匹配结果与头部运动之间的显著相关性。对于指纹识别分析,我们发现结果并非仅由高运动的参与者驱动。为了证明这一点,我们在应用严格标准(排除在任何条件下均帧位移>0.3的参与者)后重复了分析,导致n=3名参与者被排除,剩余n=12名参与者用于分析。结果本质上未发生改变,麻醉显著降低了自我-自我相似性和差异性可识别度。在排除相同三名高运动个体后,功能连接在PCA空间中的低维投影结果也未发生改变。因此,我们的结果未因扫描仪中的头部运动而受到不当影响。
4. 讨论
在这项研究中,我们利用异氟醚和丙泊酚的药理磁共振成像(pharmacological MRI),以确定麻醉诱导的无意识状态是否会降低人脑的独特性,无论是在与其他个体的大脑相比,还是与另一种物种的大脑相比。我们发现,在深度麻醉下,个体的大脑变得不那么具有自我相似性,并且在功能连接(FC)方面也不那么可识别。从空间上看,这种效应是由跨模态联合皮层的可识别性降低所驱动的。
具体来说,我们发现对可识别性贡献最大的功能连接在麻醉下受到的影响最大,这些功能连接也是在基线时对可识别性贡献最大的,同时也是连接跨模态区域的功能连接(图2b)。这些结果与跨模态皮层(如默认网络和前额顶叶控制网络)特别容易受到麻醉和更广泛的意识丧失影响的观点一致。此外,联合皮层表现出最大的个体间变异率。然而,这种变异性并非仅仅是噪声,因为前额顶叶和默认网络在意识个体中始终对可识别性贡献最大,这表明它们的变异性是特定于个体的。这可能归因于跨模态联合皮层在人脑中具有最长的成熟时间,以及最高的突触可塑性和周转率。此外,它们还表现出最低水平的皮层内髓鞘化,而髓鞘化已知会在机械和化学上抑制可塑性。因此,跨模态皮层相对不受潜在的微观结构模式和解剖连接的约束-,因此它们能够在每个个体的生命周期中根据环境需求发生变化和适应,这可以解释它们在功能连接中编码特定于个体的信息的能力。这种功能相互作用中的个体特定信息随后被麻醉暂时(且可逆地)抑制,正如本研究结果所表明的那样。实际上,这一解释与最近的证据一致,即前额区域的功能连接和灰质体积的个体差异可以预测个体对丙泊酚镇静行为效应的易感性。
确实,我们推测麻醉诱导的功能连接(FC)中个体特异性差异的抑制可能是由于麻醉对意识的抑制作用。默认网络尤其以参与对个人过去和未来的反思而闻名,这些反思按定义是每个人独特的。通过抑制即使在静息状态下也具有特征性的人类大脑的自发思维模式,本研究指出,麻醉诱导的无意识状态降低了这些模式在大脑的宏观活动和连接性中的编码程度。事实上,我们发现随着麻醉加深,自发大脑活动在认知操作的元分析模式方面(无论是自动定义的还是专家策划的)越来越难以被描述。这种效应在恢复时被逆转,尽管麻醉药物仍在血液中存在。
综合来看,认知匹配减少和由于自我相似性丧失导致的可识别性降低的结果表明了以下初步解释。在清醒状态下,大脑活动由自发的生理过程以及每个个体独特的意识流共同驱动,大脑活动必须反映这种独特性。当意识被麻醉抑制时,生理过程受到干扰,但最重要的是,使每个人独特的核心驱动力消失了,导致自我相似性降低,从而降低了可识别性,这种状态在恢复意识时得以恢复。
我们的结果不仅仅是确认麻醉会在个体内部引起功能连接的变化,还揭示了麻醉改变了不同大脑之间的关系。具体来说,麻醉下的大脑不仅与其清醒时的自己变得不那么相似,而且与其麻醉状态下的自己也变得不那么相似,总体上也不那么容易与其他大脑区分开来。相比之下,Deng等人发现,根据参与者对丙泊酚镇静(以反应时间衡量)的高或低易感性将他们分组时,与基线相比,镇静期间两组之间的功能连接差异被放大了。然而,他们的结果并不与我们自己的结果相矛盾——不仅因为Deng等人使用了不同的分析方法(比较两组与个体大脑指纹识别),而且,至关重要的是,他们除三名参与者外,其余所有参与者仍然保持清醒(Ramsay镇静量表的3级),这从他们能够执行行为任务的能力中得到了明确证明。相比之下,我们数据集中的参与者达到了Ramsay镇静量表的5级(对于丙泊酚)和6级(对于异氟醚)(即最深的水平,对应于完全失去行为反应能力和可能的意识丧失),异氟醚麻醉达到了手术深度。
因此,完全有可能在镇静期间(个体虽然困倦但仍然保持清醒和反应能力)个体在功能连接上的差异得以保留,甚至与个体在行为反应上的差异相关的功能连接差异被放大,而这些差异在完全失去意识时被消除,此时行为差异也被消除。
关于镇静与完全深度麻醉之间的相同注意事项也适用于之前的一项大脑指纹识别研究,该研究报告称,个体在右美托咪定镇静下仍然是可识别的(尽管也观察到个体全脑功能连接模式之间的差异有所减少)。与丙泊酚和异氟醚(本研究中使用的两种麻醉剂)不同,右美托咪定诱导的生理状态类似于非快速眼动睡眠第3阶段,保留了个体对外部刺激迅速恢复定向反应的能力。右美托咪定与异氟醚和丙泊酚在行为、生理和大脑功能连接上的不同效应可能归因于它们各自不同的分子作用机制:右美托咪定是一种α2肾上腺素能激动剂,而丙泊酚和异氟醚主要作用于GABA-A受体。本研究的一个优势是我们能够在两个使用不同麻醉剂的独立数据集中复制我们的结果,证明我们的发现并非特定于某种药物。
除了这些麻醉剂之间的分子差异外,Liu等人研究中的参与者处于Ramsay量表的3-4级,仍然清醒,这从他们仍然能够对指令做出反应这一事实中可以看出。因此,我们的结果(在两个使用不同麻醉剂的独立数据集中复制)并不与Liu等人的结果相矛盾;相反,这两项研究共同表明,功能大脑指纹相对能够抵御大脑状态的变化,只有在高剂量下才会出现显著的干扰,这些高剂量也会干扰反应能力并可能抑制意识。
有趣的是,跨模态联合皮层不仅在个体之间最具有异质性,而且在物种之间也表现出最大的进化扩张和大脑相关人类加速基因的最高表达。我们发现,区域对麻醉诱导的可识别性丧失的贡献在空间上与进化皮层扩张和人类加速基因的区域平均表达相关。此外,我们发现随着麻醉加深,它将人类功能连接组的位置在联合低维空间中更接近猕猴功能连接组,恢复时又回到初始位置附近。
更广泛地说,本研究在深度麻醉下人类与猕猴之间的偏差减少的结果与之前的研究一致,这些研究显示在麻醉下结构与功能之间的偏差减少。之前的研究表明,在不同物种中,麻醉大脑的时间变化功能连接模式变得更类似于其潜在的结构连接,但也有报告称在丙泊酚麻醉下局部结构-功能耦合降低。有趣的是,最近发现,像LSD和裸盖菇素这样的致幻剂(它们会引发幻觉和非常奇特的主观体验)对结构-功能耦合有相反的影响,使大脑活动和连接性不再受潜在结构连接组的限制。与麻醉和致幻剂对结构-功能关系的相反效应一致,最近的一份报告表明,裸盖菇素增加了功能连接的特异性,导致更大的差异可识别性,这与我们在不同麻醉剂中发现的结果相反。值得注意的是,相反地,最近另一项关于致幻剂ayahuasca的研究报告称,在仪式性使用致幻剂的人群中(如Santo Daime宗教团体的成员),功能连接的特异性降低。这一结果表明,致幻剂可能能够以双向方式调节功能连接的特异性,增加陌生人之间的独特性,但增加个体之间的相似性,对于那些将致幻体验作为共享、仪式化文化体验一部分的个体来说,这可能会在个体之间诱导出一种共同的心理状态。实际上,Colenbier等人和Finn等人已经表明,大脑可识别性可以通过不同的认知任务来调节。因此,尽管我们的主要结果是麻醉诱导的意识丧失会降低大脑的可识别性,但麻醉显然不是降低功能连接组独特性的唯一方式,正如Liu等人所展示的那样,可识别性相对能够抵御麻醉暴露,只有在可能抑制意识本身的高剂量下才会降低。
本研究结果表明,麻醉状态下的人类大脑更类似于其他灵长类动物的大脑,特别是人类进化扩张的区域受麻醉影响最大。未来的研究可能会调查致幻剂是否对人类大脑有相反的影响,导致人类和猕猴之间的差异更大,尤其是因为经典致幻剂的主要分子靶点——5HT-2A受体——在进化扩张的跨模态皮层中特别丰富。
本研究存在一些局限性。首先,我们遵循文献中的常见做法,使用行为反应能力的丧失作为意识丧失的标志。尽管异氟醚(sevoflurane)和丙泊酚(propofol)偶尔被报道会引发梦境,但这种情况较为罕见,因此可以推测我们研究中的大多数参与者确实处于无意识状态,尤其是考虑到所使用的麻醉深度,异氟醚数据集中的麻醉深度甚至达到了引起爆发抑制的程度。尽管如此,未来的研究可以通过使用额外的方法来评估无意识状态,而不仅仅是基于行为反应能力的丧失,例如通过EEG的慢波活动饱和度,或者基于EEG和经颅磁刺激结合的扰动复杂性指数。
从EEG文献来看,麻醉诱导所谓的EEG alpha振荡(8-12 Hz)分布的前移(anteriorization)是众所周知的,alpha功率的峰值从枕叶电极转移到额叶电极。尽管我们发现麻醉降低了fMRI连接组的整体可识别性,但个体的fMRI功能连接(FC)模式在麻醉下变得不那么相似,这似乎与EEG地形图在个体间预期的类似重新配置相矛盾。然而,重要的是要认识到,两个对象A和B各自变得更类似于第三个对象C,同时彼此之间的相似性降低,并不矛盾。此外,fMRI和EEG反映了不同的神经生物学过程,并且在不同的空间和时间尺度上运作。不同频率带的脑磁图(MEG)和EEG共波动模式彼此之间以及与fMRI之间可能非常不同,并且为指纹识别提供了不同的信息。特别是,EEG alpha前移现象发生在与本研究中研究的fMRI BOLD信号波动相隔几个数量级的时间尺度上(8-12 Hz对比0.008-0.090 Hz)。最后,前移涉及区域的行为,而指纹识别则基于不同区域之间的相互作用。在补充代码2中,我们提供了一个示例,两个系统各自经历了每个元素活动幅度空间模式的相同变化,同时在边缘水平上降低了它们的相关性。总的来说,这些因素中的任何一个都可以解释我们在fMRI结果中观察到的个体间和物种间独特性的降低,与麻醉诱导的EEG前移现象共存。通过专门研究在麻醉下进行的MEG和EEG大脑指纹识别,将这些因素区分开来,是未来研究的一个有前景的方向。
本研究的另一个明显局限性是样本量较小,这是由于在扫描仪中进行麻醉的技术和伦理挑战所导致的。确实,我们承认该领域需要更大的样本量。然而,我们在一个独立的数据集中复制了我们的结果,证明了跨数据集和药物的普遍性。我们还确保了我们的结果对分割选择(解剖或功能)的稳健性,以及对潜在混杂因素(如头动)的稳健性。此外,令人鼓舞的是,即使在时间上相隔最远的两次扫描之间(基线清醒和恢复),可识别性和自我相似性也更高,而其他条件相同的情况下,扫描之间的时间间隔越长,可识别性越低。此外,异氟醚数据集完全由男性参与者组成,丙泊酚数据集中的大多数参与者也是男性。我们期待随着该领域的扩展,在性别平衡的数据集中进行未来的复制。我们还承认,Neurosynth中功能激活与心理学术语的映射并未区分激活与去激活。然而,我们认为我们使用BrainMap50定义的元分析图进行的复制提供了对我们方法有效性的保证。尽管如此,我们注意到,Neurosynth中的认知匹配效应明显依赖于麻醉深度(图3),并且仅在达到更深麻醉水平的异氟醚数据集中观察到。因此,确定这一结果是否可以在其他数据集中复制将特别令人感兴趣。特别是,这种方法可能在意识障碍患者的数据库中具有价值,其中基于任务指令的大脑反应性解码(例如,“想象打网球”)已经使得在行为无反应的患者中识别出隐匿意识成为可能。然而,这种范式要求患者能够理解指令、将其保留在工作记忆中并执行它们——对于遭受严重脑损伤的个体来说,这是一个非平凡的要求。为了减轻这一负担,研究人员也开始使用对引人入胜的叙述(例如,电影《Taken》的片段)的自发大脑反应。然而,这种方法仍然需要语言理解和工作记忆来跟踪事件。基于元分析图与自发大脑活动之间的匹配进行解码,而不依赖于刺激,可能会进一步推进这一研究方向。
总的来说,本研究结果表明,无论使用何种特定的麻醉剂,麻醉状态下的人类大脑在个体间甚至跨物种间都变得不那么具有独特性,而那些在个体间和跨物种间最不均匀的区域受到的影响尤为显著。
参考文献
Luppi, A. I., Golkowski, D., Ranft, A., Ilg, R., Jordan, D., Bzdok, D., Owen, A. M., Naci, L., Stamatakis, E. A., Amico, E., & Misic, B. (2025). General anaesthesia decreases the uniqueness of brain functional connectivity across individuals and species. Nature Human Behaviour, Article 202502121. https://doi.org/10.1038/s41562-025-02121-9

















 394
394

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









