







Engineering Escherichia coli for Efficient De Novo Synthesis of Salidroside
将大肠杆菌工程化用于高效的红景天苷从头合成

摘要
红景天苷是一种具有多种生物活性的高价值植物来源糖苷,但目前主要的工业生产方式是从红景天植物中提取,这已不足以满足日益增长的市场需求。通过微生物发酵的生物合成路径是一种可持续且环保的替代方法。通过引入 ARO10 和 ADH6 基因,建立了前体酪醇的从头合成途径。经过系统的代谢工程改造,实现了 3.0 g/L 的酪醇产量,但酪醇的积累抑制了细胞的生长。通过适应性进化获得了一株进化菌株,其 OD600 提高了 10.0%,酪醇产量达到 3.3 g/L。随后,通过引入葡糖基转移酶 AtUGT85A1,以及过表达磷酸葡萄糖变位酶 pgm 和 UDP-葡萄糖焦磷酸化酶 galU,实现了红景天苷的从头合成。此外,对 AtUGT85A1 进行了半理性工程化改造,得到 A21G 突变体,将红景天苷的产量提高了 31.2%。最终,优化后的工程菌株在 5 L 生物反应器中实现了 16.8 g/L 的红景天苷产量,生产效率达到 0.4 g/(L·h)。本研究为未来红景天苷的工业化生产奠定了基础,并为其他酪醇衍生物的高效生物合成提供了重要指导。
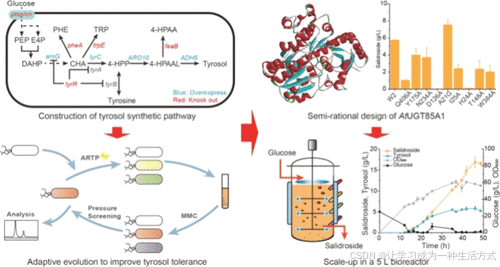
1. 引言
红景天苷(Salidroside, p-hydroxyphenylethyl-O-β-d-glucopyranoside)是一种来源于植物的酚丙烷类糖苷,由酪醇(tyrosol, 2-(4-hydroxyphenyl)ethanol)的羟基与尿苷二磷酸葡萄糖(UDPG)的半缩醛羟基缩合而成。(1,2) 红景天苷广泛存在于一些珍贵的药用植物中,如红景天(Rhodiola)、女贞子(Ligustrum lucidum)、肉苁蓉(Cistanche deserticola)和唐古特乌头(Aconitum tanguticum)。(3,4) 红景天苷具有多种有益健康的生物活性,包括抗疲劳、抗衰老、抗癌、抗肿瘤、抗氧化和抗炎作用,还对中枢神经系统和心血管系统具有保护作用。(4−6) 基于这些生物活性,红景天苷已广泛应用于食品、保健品、化妆品和药品中,因此市场需求正在快速增长。(7,8)
目前,工业生产红景天苷的方法有三种可能的途径,即传统植物提取、化学合成和生物合成。(9) 红景天植物材料的提取是当前的主要生产方式,但这一方法受到红景天苷含量极低以及红景天植物资源稀缺的限制,而后者是由于红景天植物生长缓慢且对生长环境要求苛刻。(10,11) 化学合成方法因其复杂的纯化过程以及有毒废物的产生,不适合可持续的工业生产。(12) 因此,通过微生物细胞工厂利用廉价或可再生生物质材料从头合成(de novo biosynthesis)红景天苷的可持续且环保的生物合成途径成为了一个重要的研究方向。目前已在模式生物宿主大肠杆菌(Escherichia coli)和酿酒酵母(Saccharomyces cerevisiae)中实施了多种策略。(13−15)
通过引入来自酿酒酵母(S. cerevisiae)的苯丙酮酸脱羧酶 ARO10 和来自克氏红球藻(R. cretinii)的葡糖基转移酶 UGT73B6,采用在发酵不同阶段提供不同培养基的方法,可从葡萄糖生产 56.9 mg/L 的红景天苷。(16) 类似地,通过引入来自香芹(Petroselinum crispum)的芳香醛合酶和来自拟南芥(Arabidopsis thaliana)的 UGT85A1,可获得 288 mg/L 的红景天苷。(17) 通过两株工程化大肠杆菌的共培养,经过 129 小时发酵,可达到 6.03 g/L 的红景天苷产量;(18) 而通过合理的染色体工程构建的非诱导型工程菌株,在 120 小时内可达到 9.34 g/L 的红景天苷产量。(19) 此外,还建立了大肠杆菌和酿酒酵母的共培养体系,并基于 UDP-葡萄糖系统和实验室适应性进化策略,在摇瓶水平下获得了 3.8 g/L 的红景天苷。(20) 迄今为止,红景天苷的最高产量为 26.55 g/L,这是通过对酿酒酵母进行多模块工程改造并引入来自红景天(R. rosea)的 RrU8GT33 后,在 168 小时内实现的。(21) 然而,上述微生物发酵方法由于大肠杆菌共培养系统的低产量和复杂性,以及酿酒酵母所需的长发酵时间,均不适合红景天苷的工业化生产。
本研究中,选择大肠杆菌 BL21(DE3) 作为从头合成红景天苷的底盘菌株。首先,通过引入 ARO10 和乙醇脱氢酶 ADH6,并对酪醇合成途径进行系统代谢工程改造,构建了用于从头合成酪醇的工程菌株,以解决前体供应不足和共培养系统不稳定的问题。此外,通过适应性进化提高了工程菌株的酪醇耐受性。通过引入拟南芥(A. thaliana)的葡糖基转移酶 AtUGT85A1,并过表达内源性磷酸葡糖变位酶(pgm)和 UDP-葡萄糖焦磷酸化酶(galU),实现了红景天苷的从头合成。对 AtUGT85A1 酶进行了半理性设计,得到高效的突变体 AtUGT85A1A21G。最终,优化后的工程菌株在 5 L 发酵罐中,通过补料分批发酵生产出了 16.8 g/L 的红景天苷,生产效率为 0.4 g/(L·h)。
2. 材料与方法
2.1 菌株与质粒
使用的菌株和质粒列于补充信息中。质粒 pETDuet-1 和 pRSFDuet-1 用于相关基因的表达。大肠杆菌 JM109 用作克隆宿主,而大肠杆菌 BL21(DE3) 用作亲本菌株。
2.2 质粒与菌株构建
-
使用酿酒酵母基因组和质粒 pRSFduet-1 为模板,分别扩增 ARO10 片段、ADH6 片段以及 pRSFduet 骨架,使用引物 ARO10-F/ARO10-R、ADH6-F/ADH6-R 和 RSF2-F/RSF2-R(详见补充信息)。片段与载体之间具有 20 bp 的同源臂,并通过 Shine–Dalgarno(SD)序列连接;通过 Gibson 装配获得 pRSFduet-ARO10-ADH6。
-
使用 aroGfbr 片段、tyrC 片段和质粒 pETDuet-1 为模板,分别扩增 aroGfbr、tyrC 和 pETDuet 骨架,使用引物 aroGfbr-F/aroGfbr-R、tyrC-F/tyrC-R 和 ET1-F/ET1-R;通过 Gibson 装配获得 pETDuet-aroGfbr-tyrC。
-
使用大肠杆菌基因组为模板,用引物 pgm-F/pgm-R 和 galU-F/galU-R 扩增 pgm 和 galU。
-
使用质粒 pUC-AtUGT85A1 为模板,用引物 UGT-F/UGT-R 扩增 AtUGT85A1 片段;用引物 RSF1-F/RSF1-R 线性化载体 pRSFdute-ARO10-ADH6;通过 Gibson 装配构建 pRSFduet-pgm-galU-ARO10-ADH6。
-
用引物 ET2-F/ET2-R 线性化载体 pETDuet-aroGfbr-tyrC,并将线性化载体与 AtUGT85A1 片段通过 Gibson 装配构建 pETDuet-aroGfbr-tyrC-AtUGT85A1。
-
糖基转移酶突变体的构建采用全质粒 PCR 方法,使用完整质粒 pETDuet-aroGfbr-tyrC-AtUGT85A1 作为模板,在选定的突变位点设计上下游引物,通过高保真 DNA 聚合酶完成 PCR 操作。PCR 产物转化至大肠杆菌 JM109,通过 PCR 检测引物筛选菌落,选取阳性菌落进行测序进一步验证。
2.3. 适应性进化结合ARTP诱变
通过大气压常温等离子体(ARTP)处理和自动高通量微生物液滴培养仪(MMC,源清天木生物科技有限公司,中国)进行适应性进化。取10 μL细胞悬液培养至对数生长期,并均匀涂布在无菌金属载玻片上;诱变腔室提前用紫外线照射30分钟,然后将金属载玻片依次放置于诱变腔室载体上。在不同的照射时间后,将金属载玻片自动投放到含有甘油(1 mL)的2.0 mL Eppendorf离心管中,震荡5分钟后取出液体。液体依次稀释至10-1、10-2、10-3和10-4倍,然后接种在Luria–Bertani(LB)琼脂平板上以确定细胞死亡率。根据测得的细胞死亡率,计算出最佳处理时间为60秒。将底盘菌株进行60秒ARTP诱变处理后,将甘油细胞悬液直接置于LB液体培养基中,培养至OD600达到0.6。根据MMC厂商的说明书开展适应性进化实验。酪醇浓度分别设置为0 g/L(第4号注射瓶)和10 g/L(第6号注射瓶)。传代方法设置为根据时间传代;初始传代参数设为8,随后根据测得的OD600随时调整。
2.4. AtUGT85A1突变体的预测与分析
使用ChemDraw 14.0软件绘制二维分子结构,并通过ChemDraw3D将二维结构转换为三维结构,保存为mol2文件。将三维结构导入后,使用Molecules模块中的“Prepare ligands”功能对小分子进行处理。将目标氨基酸序列输入UniProt进行BLAST分析,然后使用AutoDock软件对蛋白质进行预处理,包括删除水分子、添加质子和电荷,并提取结构中的原始配体,随后通过PyMOL可视化处理后的蛋白质。利用AutoDock软件在蛋白质中搜索潜在的结合位点,并导出对接盒文件。分子对接过程中,首先将准备好的大分子、配体和Vina放入一个文件夹中。接着,将准备好的配体化合物结构和大分子作为受体,保存为pdbqt格式,并使用Vina进行对接。利用PyMOL将对接结果组合成复合体,最后使用PyMOL和Discovery Studio 2019对对接相互作用进行分析和可视化。
2.5. 培养基和细胞培养条件
Luria–Bertani(LB)培养基中含有10 g/L蛋白胨、5 g/L酵母提取物和10 g/L氯化钠,用于种子培养,根据需要加入琼脂(20 g/L)、氨苄青霉素(100 μg/mL)和卡那霉素(50 μg/mL)。发酵培养基包括25 g/L葡萄糖、10 g/L甘油、7 g/L酵母提取物、7.5 g/L硫酸铵((NH4)2SO4)、3 g/L磷酸氢二钾三水合物(K2HPO4·3H2O)、2 g/L磷酸二氢钾(KH2PO4)、2 g/L七水硫酸镁(MgSO4·7H2O)、10 g/L碳酸钙(CaCO3)、1 g/L枸橼酸单水合物、0.1 g/L硫胺素,以及1 mL/L微量元素溶液。(22) 根据需要加入氨苄青霉素(100 μg/mL)和卡那霉素(50 μg/mL)。
摇瓶发酵在250 mL摇瓶中进行,提前在LB培养基(3 mL)中加入所需抗生素。挑取单菌落接种,在37 °C、220 rpm条件下于摇管中培养过夜12小时。将种子液加入预先含抗生素的发酵培养基(100 μg/mL氨苄青霉素和50 μg/mL卡那霉素);接种量为2%(500 μL种子液加入25 mL发酵培养基)。在220 rpm、37 °C条件下于摇瓶中培养3小时后,加入终浓度为0.1 mM的异丙基-β-d-1-硫代半乳糖吡喃糖苷(IPTG),将发酵温度调整为30 °C。分别在发酵24、48和72小时后取样。
工程菌的放大培养在5 L生物反应器中进行。菌株在含所需抗生素(100 μg/mL氨苄青霉素和50 μg/mL卡那霉素)的LB琼脂平板上活化,并在37 °C下培养过夜。单菌落转移至含LB培养基(3 mL)和所需抗生素的摇管中培养8小时。以2%体积比接种量将菌液加入250 mL摇瓶中,含LB培养基,在37 °C条件下培养6小时。生物反应器的培养温度为37 °C,通气量为2 L/min,初始搅拌速度为300 rpm,当溶解氧(DO)下降时,通过调整搅拌速度将DO恒定控制在30%或40%。搅拌速度范围设置为300–900 rpm。pH值设为7.0 ± 0.1,使用4 M NaOH控制pH值。种子液接种量为6%体积比。当OD600达到30时,将反应器冷却至30 °C,加入终浓度为0.1 mM的IPTG进行诱导,并补充葡萄糖(500 g/L)。通过调整进料流速,将残糖量控制在10 g/L以内。
2.6. 分析方法
发酵培养基中酪醇和红景天苷的含量通过高效液相色谱(HPLC,岛津LC-20A,日本京都)测定。样品用甲醇稀释20倍,并在40 °C使用反相C18柱进行定量分析。流动相为(A)水/0.1%三氟乙酸(TFA)和(B)乙腈/0.1% TFA,流速为1.0 mL/min,紫外检测器设置为274 nm。梯度洗脱程序如下:0–5分钟10% B,10分钟60% B,12分钟90% B,16分钟2% B,16–20分钟0% B。(22)
2.7. 统计分析
所有实验均以三次重复进行,结果以平均值 ± 标准差(SD)表示。使用Origin 2019b进行统计分析。
结果与讨论
3.1. 高效酪醇前体的从头合成途径构建
高效从头合成红景天苷需要充分供应其前体酪醇,因此构建了一个用于合成酪醇的生物合成途径(图1A)。通过在大肠杆菌BL21(DE3)中过表达来源于酿酒酵母的ARO10和ADH6,获得了一株能够从头合成酪醇的菌株LC1。具体而言,LC1积累了26.78 mg/L的酪醇(图1B)。
为了进一步提高酪醇的积累量,实施了一系列优化措施,包括敲除tyrR以限制通量分流至芳香族氨基酸合成途径,过表达aroGfbr和tyrC,敲除参与磷酸转移系统的ptsG和crr,以及敲除竞争途径中的pheA和trpE。构建了六株工程菌株,分别命名为LC2–LC7(图1B)。
最终,菌株LC7在72小时内实现了最高的酪醇产量,达到3.0 g/L,但酪醇的高浓度显著抑制了菌株LC7的细胞生长(图1C)。
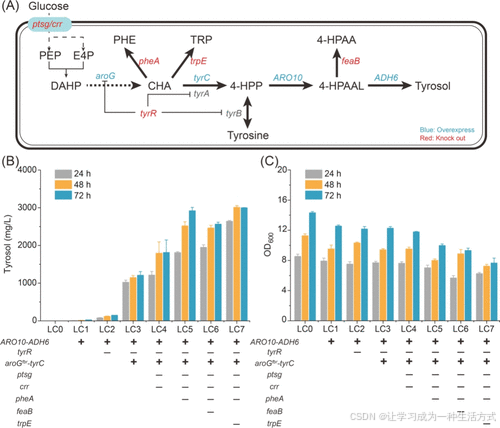
图1. 在大肠杆菌中构建从头合成酪醇的生物合成途径 (A) 通过代谢工程构建高效酪醇途径的示意图。(B) 不同工程菌株在摇瓶中生产酪醇的能力。(C) 不同菌株在摇瓶中的OD600(细胞生长)。PEP: 磷酸烯醇式丙酮酸,E4P: 赤藓糖-4-磷酸,DAHP: 3-脱氧-D-阿拉伯庚酮糖-7-磷酸,CHA: 莽草酸,PHE: 苯丙氨酸,TRP: 色氨酸,4-HPP: 4-羟基苯丙酮酸,4-HPAAL: 4-羟基苯乙醛,4-HPAA: 4-羟基苯乙酸。“+”表示引入相关基因,“–”表示敲除相关基因。
关于酪醇生物合成的先前研究中,已报道使用大肠杆菌或酿酒酵母进行的生物转化或从头合成方法。(21,23,24) 报道的酪醇不同生物合成途径包括:(1) 通过脱羧和脱氢将由莽草酸合成的4-羟基苯丙酮酸(4-HPP)转化为酪醇;(25) (2) 通过转氨、脱羧和脱氢将酪氨酸转化为酪醇,或生成4-羟基苯乙醛(4-HPAAL)后再脱氢;(27) 以及(3) 通过香豆酸代谢途径生物合成。(28) 为提高大肠杆菌中酪醇的产量,采用了多种策略,包括关键酶的过表达和半理性设计、缓解抑制性调控、消除竞争代谢途径以及优化系统发酵条件。(24,25,29) 本研究选择了一条相对较短的4-HPP途径。通过协同工程改造获得了酪醇高产菌株LC7,但也出现了细胞生长受抑的现象。这一现象可能由两个因素引起:过多的基因敲除和/或积累的酪醇。
3.2. 通过自适应进化提高工程菌株的酪醇耐受性
为缓解细胞生长抑制问题,采用自适应进化进行酪醇压力筛选,并同步对大肠杆菌BL21(DE3)ΔtyrRΔptsGΔcrrΔpheAΔtrpE菌株进行ARTP诱变(图2A)。在评估ARTP致死率后,选择60秒处理时间(98.0%致死率),遵循致死率大于90%时诱变效果更高的原则(图2B)。对ARTP处理获得的突变株通过MMC进行自适应进化,调整传代参数以逐步增加酪醇浓度,维持连续传代。当调整参数后菌株难以生长时,终止自适应进化过程,此时酪醇浓度达到4.0 g/L(图2C)。
选择了七株进化菌株,并通过引入pRSFDuet-ARO10-ADH6和pETDuet-aroGfbr-tyrC获得了七株对应的进化工程菌株,分别命名为LC8–LC14。摇瓶发酵测试显示,其中六株菌的酪醇产量高于LC7,产量最高的菌株为LC10,达到3.3 g/L(图2D)。对高酪醇产量的两株进化菌(LC10和LC12)在3.0 g/L酪醇存在下的生长进行了测定。总体来看,LC10的生长(OD600)比对照菌株LC7高出10.0%(图2E)。
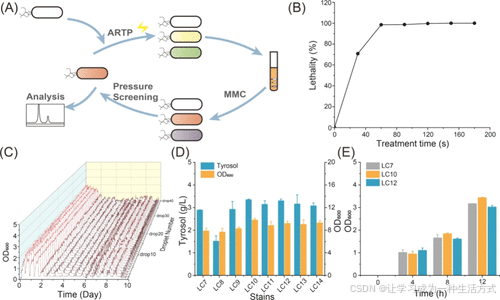
图2. 通过自适应进化筛选高酪醇耐受菌株 (A) 自适应进化联合ARTP诱变处理大肠杆菌BL21(DE3)ΔtyrRΔptsGΔcrrΔpheAΔtrpE的示意图。(B) ARTP诱变处理的致死率曲线。(C) 自适应进化(MMC)的瀑布图。(D) LC7和进化菌株在72小时摇瓶发酵中的酪醇产量和OD600。(E) 在3.0 g/L酪醇存在下,进化菌株LC10和LC12与对照菌株LC7(对照组)的OD600(细胞生长)对比。
酪醇具有显著的抗菌特性,因此酪醇及其衍生物能够抑制细胞生长,并影响细胞形态和生理状态。(30,31) 在本研究中,酪醇对高酪醇生产菌株的抑制作用似乎是限制细胞生长的主要因素,因为无论是本研究还是以往的报道中,菌体生物量都相对较低。(25,32) 之前研究中,使用群体感应动态调控技术缓解了酪醇衍生物4-羟基苯乙酸对大肠杆菌的毒性。(33) 在本研究中,通过自适应进化技术改善了微生物的功能,例如耐受恶劣环境压力、加速生长或代谢速率以及激活潜在的生物合成途径。(34,35) 此外,自适应进化与ARTP联合使用提高了效率。同样的联合方法已经成功获得了一株高1,3-丙二醇耐受性的小克螺菌菌株。(36)
3.3. 红景天苷的从头合成
为了构建红景天苷的从头合成途径,分别引入了来自拟南芥的糖基转移酶AtUGT85A1以及内源的磷酸葡萄糖变位酶(pgm)*和*UDP-葡萄糖焦磷酸化酶(galU)。相关质粒被转入对照菌株大肠杆菌BL21(DE3)ΔtyrRΔptsGΔcrrΔpheAΔtrpE和进化菌株Lce03,分别获得菌株W0和W1。在摇瓶中培养72小时后,菌株W0产生了0.2 g/L的红景天苷,而菌株W1产生了0.4 g/L的红景天苷(图3A)。发酵液中累积的产物通过LC-MS分析确认为红景天苷(见支持信息)。然而,在W0和W1中,大部分酪醇(>2.8 g/L)未被糖基化。
为了进一步增加UDPG的供应,对pgm和galU进行了过表达,获得了菌株W2。在摇瓶中培养72小时后,菌株W2生产了4.7 g/L的红景天苷,但仍有2.4 g/L的酪醇未被糖基化(图3B),这表明糖基化过程可以进一步加速。
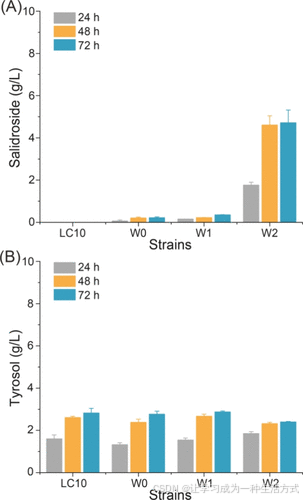
图3. 在大肠杆菌中构建红景天苷的从头合成途径 (A) 红景天苷产量。(B) 不同工程菌株在摇瓶发酵中的酪醇残留浓度。
酪醇的糖基化是通过微生物发酵合成红景天苷的限速步骤。除了构建酪醇生物合成途径外,挖掘更高效的糖基转移酶是实现红景天苷高效生物合成的另一种策略。(37) 已鉴定出多种植物来源的糖基转移酶能够将酪醇糖基化为红景天苷,例如来源于红景天属植物的RsUGT73B6、RsUGT72B14 和 RrU8GT33,来源于拟南芥的AtUGT85A1 和 AtUGT73C5,以及来源于芝麻的SiUGT1。(17,21,38) 一些微生物中的功能性糖基转移酶也被发现,如来源于地衣芽孢杆菌的UGTBL1 和 YjiC。(13,39) 基于这些糖基转移酶的研究,已经建立了一种使用全细胞UDP-葡萄糖转移酶生物催化剂将酪醇转化为红景天苷的工艺。(12) 充足的UDPG(UDP-葡萄糖)供应也是提高植物来源天然产物糖基化效率的重要因素。(40) 本研究通过过表达pgm 和 galU 增加UDPG的供应,显著提高了红景天苷的产量。
3.4. 糖基转移酶AtUGT85A1的半理性设计
为了提高糖基化效率,以AlphaFold 2数据库中预测的AtUGT85A1三级结构为基础,对其进行了半理性设计。在模拟模型中,大多数区域的预测局部距离差异测试(pLDDT)得分高于90,仅有少数与预测活性中心无关的区域得分较低。该预测结构具有较高精度,可用于分子对接分析。利用Discovery Studio对AtUGT85A1与糖基供体UDP-葡萄糖和糖基受体酪醇进行了分子对接(图4A, B)。
对活性中心5 Å范围内的氨基酸进行了丙氨酸扫描,将这些氨基酸突变为丙氨酸,或者如果该位置本身是丙氨酸,则突变为甘氨酸。筛选出六个位点(Q406, Y157, N234, D126, A21 和 I25)进行突变(详见支持信息)。此外,通过分子对接分析发现与配体相互作用的三个位点(H24, T148 和 W384)也进行了突变,共获得了九种突变体,分别命名为W3–W11。
突变体W7(A21G)累积了7.5 g/L的红景天苷,比野生型高31.2%,同时酪醇的积累显著减少。其余突变体对红景天苷生物合成未表现出积极作用,而D126A和H24A显著降低了红景天苷的产量,表明这两个氨基酸是关键活性位点残基(图5A, B)。
对A21、D126和H24这三个位点进行了NNK饱和突变筛选,但未发现其他有效突变体。然而,对突变体A21G的进一步建模显示,将丙氨酸突变为甘氨酸改善了活性中心附近一段无规卷曲结构的柔性,从而降低了底物进入活性中心结合口袋的空间位阻(图5C–E)。
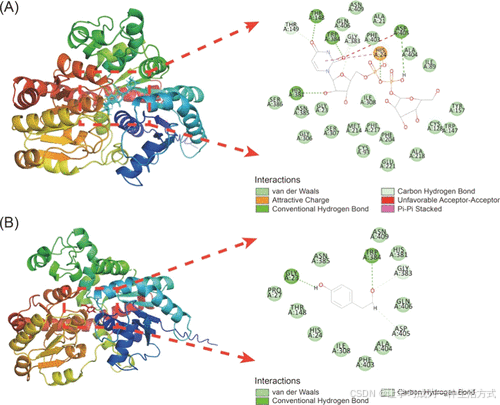
图4. UGT85A1的同源建模和分子对接 (A) 糖基转移酶UGT85A1与UDPG的分子对接。(B) 糖基转移酶UGT85A1与酪醇的分子对接。
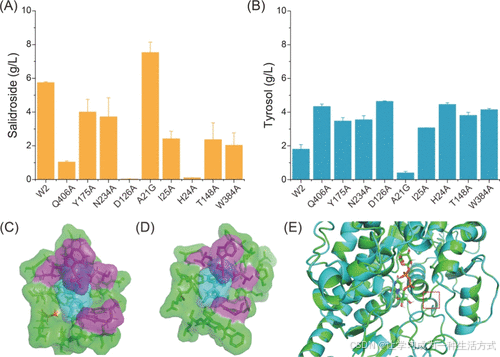
图5. 筛选最优UGT85A1变体 (A) 不同UGT85A1变体在72小时摇瓶培养中的红景天苷产量(以表达野生型UGT85A1的菌株W2为对照)。 (B) 不同UGT85A1变体在72小时摇瓶培养中的酪醇积累量。 (C) 野生型UGT85A1的建模结构。 (D) 变体UGT85A1A21G的建模结构。 (E) 野生型UGT85A1与其变体UGT85A1A21G的结构对比,红色方框标记了A21的位置。
计算机辅助定向进化已发展成为一种有效的酶改造方法,用于引入期望的特性,例如更高的活性、稳定性以及更高或更低的选择性。(41,42) 半理性和理性设计可以帮助识别潜在的有益突变位点,从而解决天然酶的功能不足问题。与传统的随机诱变相比,(半)理性设计不仅显著减少了实际工作量,还提高了获得有益突变的概率。(43,44) 已有多种糖基转移酶被改造以提高其对天然产物的糖基化效率,例如用于人参皂苷生物合成的UGT1和YjiC,(45,46) 以及用于酚类糖苷生物合成的UGTBL1。(47) 近年来,计算机辅助的“学习与设计”工具被开发出来,包括蛋白质结构预测、分子对接和分子动力学模拟,从而使预筛选过程更高效。(48) 在本研究中,尽管AtUGT85A1的晶体结构尚未报道,但通过预测结构成功筛选出了潜在的突变位点,其中一个突变位点(AtUGT85A1A21G)被证实是一个积极的变体。
3.5. 红景天苷在5 L生物反应器中的补料分批发酵
为了进一步评估工程菌株W7(表达AtUGT85A1A21G)的潜力,在5 L生物反应器中对其进行了放大培养,并以表达野生型UGT85A1的菌株W2为对照。放大过程中出现了一个问题,这些菌株的生长表现较差,显然是因为trpE的敲除导致了营养缺陷。为了解决这一问题,通过将起始密码子ATG替换为GTG,在W2和W7中原位重新表达了trpE,分别生成菌株W12和W13,从而恢复了下降的细胞生长(图6A, B)。
菌株W12生产了4.5 g/L的红景天苷,同时残留了13.4 g/L的酪醇,而菌株W13生产了11.8 g/L的红景天苷,残留酪醇为7.0 g/L,验证了通过表达变体UGT85A1A21G显著改善了红景天苷的生物合成。然而,过量的酪醇积累仍然抑制了细胞生长,尤其是在菌株W12的发酵过程中。为了进一步提高红景天苷的产量,在W13的发酵过程中将溶解氧(DO)从30%提高到40%。与30% DO相比,细胞生长和葡萄糖消耗均提高了40%,最终OD600达到类似的最大值(约60)。红景天苷产量提高了42.4%,在46小时达到16.8 g/L,生产率提高至0.4 g/(L·h)(图6C)。
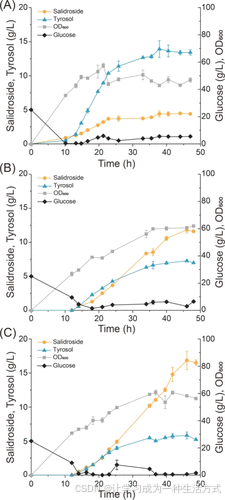
图6. 在5 L生物反应器中放大红景天苷生产的时间过程 (A) 在30%溶解氧(DO)下,菌株W12(表达野生型UGT85A1)的红景天苷生产过程。 (B) 在30% DO下,菌株W13(表达变体UGT85A1A21G)的红景天苷生产过程。 (C) 在40% DO下,菌株W13(表达变体UGT85A1A21G)的红景天苷生产过程。
本研究中获得的红景天苷生产率显著高于此前的报道,且红景天苷产量也是大肠杆菌中报道的最高水平。然而,即使在最佳发酵条件下,仍然残留大量酪醇,表明糖基化效率仍有不足。未来研究应致力于进一步提高糖基化效率,例如通过解析AtUGT85A1的晶体结构实现更有效的理性改造,或基于日益丰富的植物组学数据挖掘潜在更高效的糖基转移酶。(49,50)
此外,5 L生物反应器的生产效率与摇瓶发酵的结果不匹配,表明在发酵条件方面仍有较大优化空间,例如pH控制、补料模式以及通过动力学参数分析进行改进。(21,51) 在发酵后期,红景天苷的产量下降,这表明大肠杆菌中存在内源糖苷水解酶,其可以脱糖红景天苷。如果能够识别并敲除该酶,则有可能进一步提高产量和生产率。
4. 结论
本研究通过系统改造获得了一株高效的红景天苷从头合成工程大肠杆菌菌株。这些改造包括构建可积累高浓度前体酪醇的平台菌株、实施自适应进化、引入外源糖基转移酶AtUGT85A1并过表达内源pgm和galU以实现红景天苷生物合成,以及通过半理性设计提高AtUGT85A1的催化效率。最终优化的工程菌株在5 L生物反应器中生产了16.8 g/L红景天苷,生产率达到0.4 g/L/h,为未来工业化生产奠定了基础。这些研究结果还为其他酪醇衍生物的高效生物合成提供了指导。

















 5905
5905

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









