






点击蓝字 关注我们
艰难梭菌感染的宿主免疫反应

iMeta主页:http://www.imeta.science
综 述
● 原文链接DOI: https://doi.org/10.1002/imt2.200
● 2024年5月11日,西北农林科技大学曹阳春团队在iMeta在线发表了题为 “Understanding host immune responses in Clostridioides difficile infection: Implications for pathogenesis and immunotherapy” 的文章。
● 本文综述了近期关于Clostridioides difficile感染(CDI)发病机制的理解,以及宿主免疫系统反应如何影响疾病的进展和严重程度,旨在增强我们开发基于免疫治疗的CDI治疗方法的能力。
● 第一作者:王腊梅、Javier A. Villafuerte Gálvez
● 通讯作者:曹阳春(caoyangchun@126.com)、 Xinhua Chen (xchen1@bidmc.harvard.edu)
● 合作作者:Christina Lee、武圣儒、Ciaran P. Kelly
● 主要单位:西北农林科技大学动物科技学院
亮 点

● Clostridioides difficile感染(CDI)受多种因素影响,包括抗生素使用、年龄、并发症、免疫基因多态性和抗体水平;
● 免疫反应在控制CDI的发展和严重程度方面起着至关重要的作用;
● 免疫治疗对于推动CDI的治疗和预防策略具有重要意义。治疗方法包括单克隆抗体、疫苗和粪便微生物移植(FMT)的应用。
摘 要
Clostridioides difficile(C. difficile)是全球住院病人腹泻的主要致病菌。感染C. difficile是由于分泌大量糖基化毒素蛋白,这可能导致易感宿主出现毒性巨结肠或死亡。C. difficile的一个关键生物学特征是其在人体宿主中持续无症状存在的能力。与有症状的患者相比,携带无症状菌群或经历C. difficile感染(CDI)一次而无复发的个体显示出更强的免疫反应。这些免疫反应起着重要作用,因为它们在CDI的发展、进展、预后和最终致病中起着至关重要的作用。尽管如此,我们目前对参与CDI的免疫反应的理解仍然有限。因此,进一步的研究以阐明它们的潜在机制是必要的。本文综述了近期关于CDI发病机制的理解,以及宿主免疫系统反应如何影响疾病的进展和严重程度,旨在增强我们开发基于免疫治疗的CDI治疗方法的能力。
视频解读
Bilibili:https://www.bilibili.com/video/BV12s421A77A/
Youtube:https://youtu.be/eom4BuNDglE
中文翻译、PPT、中/英文视频解读等扩展资料下载
请访问期刊官网:http://www.imeta.science/
全文解读
引 言
Clostridioides difficile (C. difficile) 是一种厌氧的革兰氏阳性杆菌,能够产生孢子。它在人体肠道中生长繁殖,尤其是抗生素的使用引起的结肠微生物群自然紊乱之后,更利于C. difficile生长。这种病原体可以引起人类严重的胃肠道感染,从轻度腹泻到急性结肠炎甚至死亡。在美国,C. difficile感染导致了巨额的医疗支出。C. difficile感染(CDI)造成了全球公共卫生的重大挑战,每年全球报告约有50万例病例和约2万例死亡。广谱抗生素的使用是CDI的主要危险因素之一,可导致肠道微生物群的组成和功能发生显著变化。在过去几十年中,CDI的发病率和严重程度显著增加,特别是在医疗机构内。像老人、曾长期使用抗生素或免疫系统功能减弱的住院患者等易受影响的人群更容易患上CDI。
CDI发生时,C. difficile在肠道内定植并释放毒素,损害肠道粘膜和肌动蛋白细胞骨架。CDI的发病机制涉及几个关键步骤。首先,通常在抗生素治疗期间,C. difficile定植发生在孢子被摄入的时候,它们可以在胃酸性环境下存活并到达结肠。在特定条件下,例如来自自然肠道微生物群的竞争减少,这些孢子在进入结肠后会发生萌发转变为营养细胞。随后,C. difficile释放两种主要毒素,分别称为肠毒素A(TcdA)和细胞毒素B(TcdB),它们与肠道细胞上的受体结合,破坏细胞功能并引发炎症反应和腹泻。TcdA和TcdB通过损害细胞功能的重要性突显了它们在CDI病理中的作用,并强调了在治疗干预中聚焦这些毒素的重要性。
肠道微生物组在保护免受CDI侵害方面发挥着至关重要的作用,通过与C. difficile竞争资源并产生抑制其生长的代谢产物。当广谱抗生素扰乱了这种平衡时,就会形成对C. difficile有利的环境,从而促进其增殖。此外,抗生素可能会清除有助于维持肠道健康的有益细菌,进一步加重CDI并促使其复发。由于肠道内持久的孢子和广谱抗生素引起的肠道微生物组紊乱等因素,复发性CDI构成了重大的临床挑战。因此,迫切需要研究新型的替代治疗方法来管理CDI。虽然现有的治疗方法显示出了有效性,但挑战仍然存在,特别是在处理复发性CDI方面。正在进行的研究正在探索替代的治疗途径,包括益生菌、噬菌体疗法、抗毒素疗法、抗微生物肽和免疫调节策略。此外,像基因编辑和其他生物技术方法这样的新兴技术也有望在彻底改革CDI治疗方面发挥作用。在这种情况下,我们的重点是探索免疫调节策略用于CDI治疗,旨在确定更有效且更安全的治疗选择,以应对这一临床挑战。
免疫反应在对抗CDI中发挥着至关重要的作用,因为肠道粘膜免疫系统是对抗这种感染的主要防御机制。在CDI期间,C. difficile产生的毒素会在肠道粘膜中引起炎症反应,从而激活宿主免疫细胞并释放免疫因子(图1)。巨噬细胞、淋巴细胞和树突状细胞(DCs)是参与检测和消除有害细菌和毒素以保护宿主的重要免疫细胞。值得注意的是,天然淋巴细胞(ILCs)的缺失在急性CDI阶段可以显著增加死亡率。研究表明,将CD90+和CD127+ ILCs转移到ILC缺乏的小鼠中,可以促进对CDI具有较高的易感性小鼠的恢复。尽管C. difficile持续增殖和TcdA和B的产生,这些小鼠仍能够增加体重恢复腹泻。此外,免疫系统和炎症的调节可能会影响CDI患者的临床表现,如疾病严重程度和复发的可能性。研究发现,复发性CDI患者往往在对TcdA和B的IgM水平较初次感染的患者更低。尽管存在这一观察结果,但C. difficile与免疫功能相互作用的精确机制仍不清楚。全面了解对C. difficile的免疫反应在非炎症(稳态)和炎症条件下的作用,将极大地增强我们有效管理和预防疾病进展的能力。
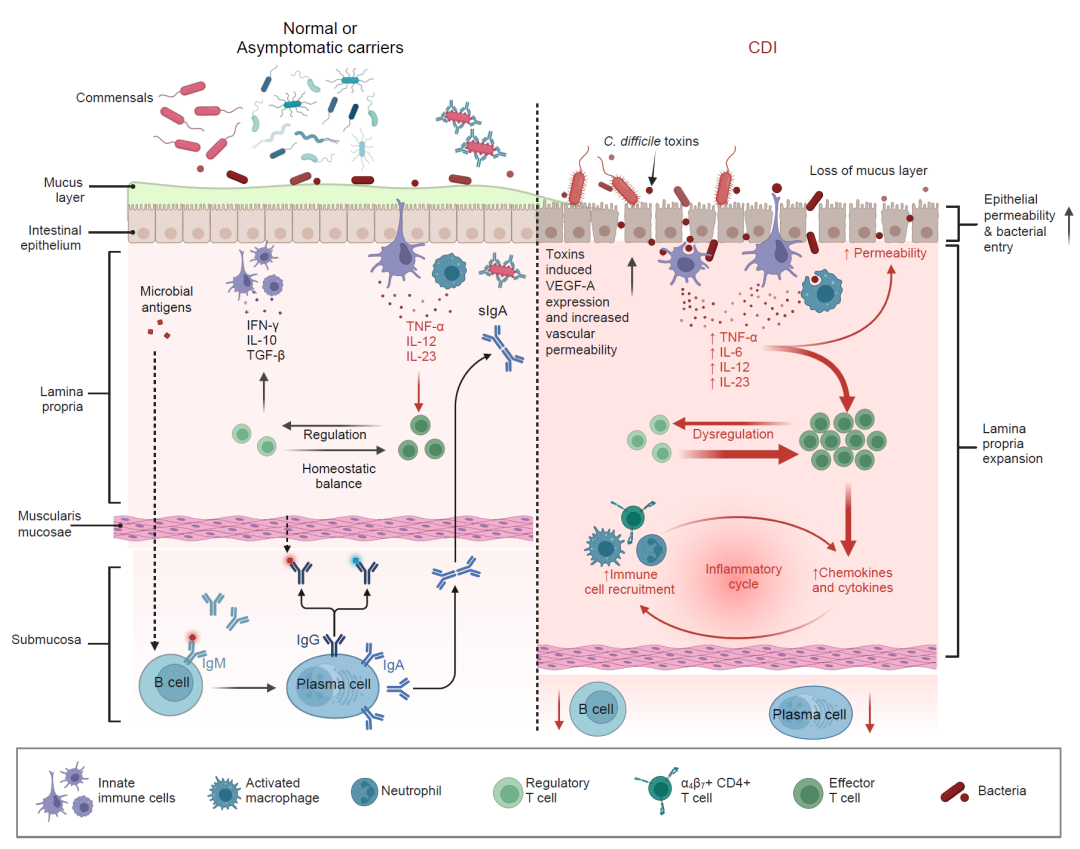
图1. 在健康状态和患有艰难梭菌感染(CDI)的个体之间的肠道免疫反应存在显著差异。
在健康个体中(左侧面板),粘膜下通常存在多种免疫细胞并分泌细胞因子。抗炎因子,如TGF-β和IL-10,在抑制免疫反应方面发挥着关键作用。此外,先天和适应性免疫细胞释放促炎因子,调节肠道微生物的进入,并防御病原体的侵入。健康个体的IgA、IgM和IgG水平通常比患有CDI的个体更高。在经历肠道炎症的患者中(右侧面板),各种因素导致细菌暴露增加。这些因素包括保护性黏液层的破坏,上皮细胞紧密连接的失调,肠道内壁的增透性增加,以及细菌对上皮细胞的附着增强。在CDI患者中,先天免疫细胞促进TNF-α、IL-6、IL-12、IL-23和趋化因子的水平增加。C. difficile, 艰难梭菌; CDI, 艰难梭菌 感染; TGF-β, 转化生长因子-β; IL-10, 白介素-10; IgA, 免疫球蛋白A; IgM, 免疫球蛋白M; TNF-α, 肿瘤坏死因子-α; sIgA, 分泌性 免疫球蛋白A; VEGF-A, 血管内皮 生长因子A; IFN-γ, 干扰素-γ。
CDI的发病机制
CDI的发病主要源于两种同源毒素TcdA和TcdB的作用,它们结合宿主细胞受体后被内吞。这两种毒素含有能够通过糖基化直接损害宿主的Rho/Rac GTP酶的结构域,导致细胞功能受损。这种损害破坏了结肠上皮的完整性,引起肌动蛋白细胞骨架的崩裂,并破坏了上皮细胞的屏障功能。随后,引起细胞凋亡和组织损伤,对宿主上皮细胞的损伤引发血管通透性增加,进而导致红细胞和血红素释放到胃肠道腔内,最终导致腹泻和结肠炎。此外,某些菌株的C. difficile产生二元毒素(CDT),加剧了CDI的严重程度,进一步损害宿主细胞。
肠道菌群的失调是CDI发病机制的核心。随着C. difficile毒素扰乱了胃肠道内共生菌的平衡,引发了炎症,并影响了多种代谢途径,包括宿主细胞对碳水化合物和氨基酸的吸收和利用。这种干扰创造了一个有利于C. difficile定植和增殖的环境。肠道菌群失调不仅有利于C. difficile的生存,而且显著影响了肠道免疫功能。
CDI患者和无症状携带者微生物组的比较分析揭示了明显的差异。无症状携带者的微生物组中富集了大肠杆菌属物种。此外,肠道微生物组学分析表明共生大肠杆菌携带的蔗糖降解途径的增加,以及可能参与淀粉和蔗糖降解的糖苷水解酶的增加。粪便微生物代谢组学分析进一步证实了这种碳水化合物降解模式,发现无症状携带者中的碳水化合物化合物丰富于CDI患者。此外,C. difficile毒素引起的菌群失调可能通过损害有益菌的保护作用并削弱对C. difficile等病原体的免疫抑制功能。这种干扰影响了肠道相关淋巴组织(GALT)和粘膜免疫系统,导致抗菌肽、免疫球蛋白(Ig)等保护因子的减少。因此,粘膜免疫防御力减弱,创造了一个更有利于CDI的环境。宿主免疫系统与肠道菌群之间的微妙平衡对于维持肠道内稳态和预防感染至关重要。C. difficile毒素破坏了这种平衡,不仅促成了初次感染,还促进了CDI的复发。
CDI的进展和发病率受到多种因素的影响,所有这些因素都可能显著影响感染的发展和严重程度,例如抗生素使用和肠道手术、年龄增长、导致功能障碍的并发症、免疫基因(如白细胞介素(IL)-8)的遗传变异,以及对毒素的抗体水平降低(图2)。免疫功能减弱个体的CDI患病率明显高于普通人群。例如,在血液学-肿瘤学领域,CDI患病率范围从6%到33%,而肺移植接受者的患病率高达23%。在艾滋病毒感染者中,患病率为7.1-8.3/1000人年。免疫功能低下个体CDI的复发也很常见,血液学-肿瘤学人群和实体器官移植接受者的复发率可达40%。在2019冠状病毒疾病(COVID-19)大流行期间,在人类免疫缺陷病毒感染患者中,世界卫生组织统计关于CDI的发病率范围为每1万人有2.8至15.8例。此外,研究表明,诊断为炎症性肠病(IBD)的个体比没有IBD的人更容易感染CDI。已确诊为严重急性呼吸综合症冠状病毒2(SARS-CoV-2)感染者和从COVID-19康复的个体被视为CDI的高风险患者。多种因素导致了这种增加的风险,包括在COVID-19相关并发症治疗期间频繁使用抗微生物药物以及对正常肠道菌群的潜在破坏。此外,免疫系统可能在SARS-CoV-2感染过程中或由于感染后并发症而受损,使个体更容易受到CDI等机会性感染的影响。因此,医疗保健提供者在具有COVID-19病史的患者中应密切注意CDI的征兆和症状,并实施适当的预防措施和治疗策略,以减轻该人群中CDI的风险。
理解这些因素对于制定有效的预防和治疗CDI策略至关重要,特别是对于易感人群,如免疫受损者和具有基础健康问题者。
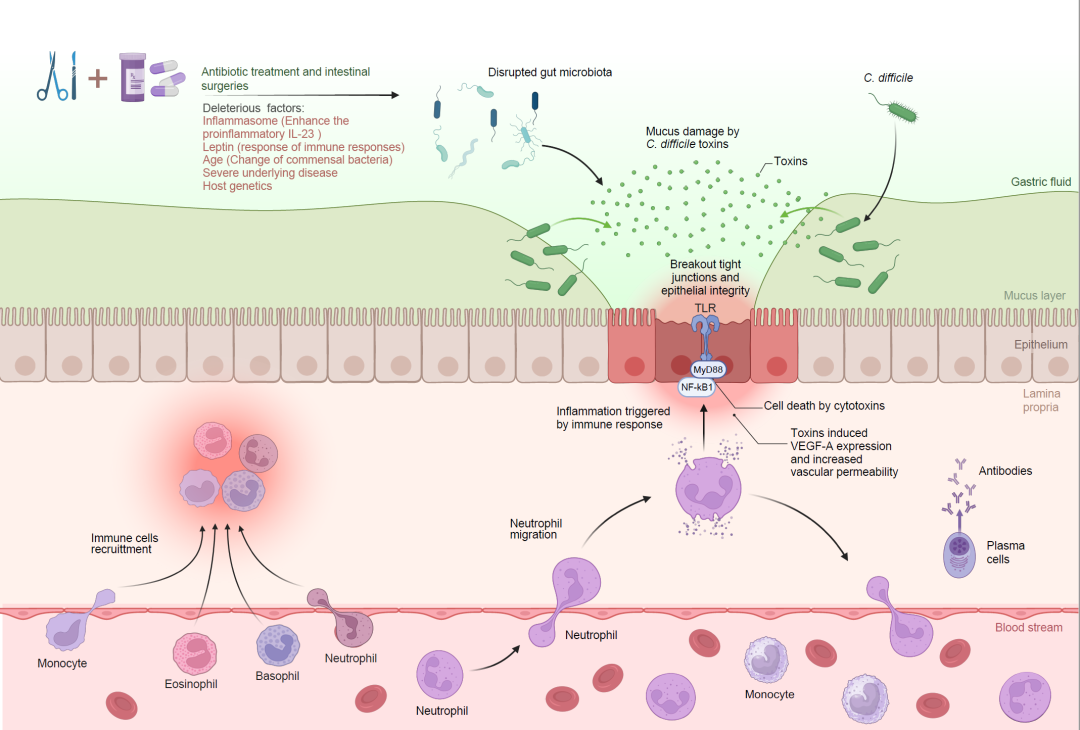
图2. CDI的发病机制
长期使用抗生素和肠道手术可能会扰乱肠道微生物群落,从而有可能导致CDI。与CDI进展相关的风险因素包括年龄相关的共生菌变化、潜在合并疾病以及宿主遗传学。NF-κB:核因子κB,TLR:Toll样受体。
宿主免疫反应对治疗C. difficile至关重要
免疫细胞在CDI的发展、进展和预防中发挥着至关重要的作用。它们产生各种细胞因子,包括干扰素(IFNs)、肿瘤坏死因子(TNF)和白细胞介素(ILs),调节炎症反应和免疫反应(图3)。这些因素通过协调免疫反应来影响CDI的严重程度和预后。它们招募和激活其他免疫细胞,促进细菌清除,并调节炎症过程。然而,免疫反应失调可能会促进CDI的发病机制。由于过度免疫反应导致的过度炎症可能会导致组织损伤,并加重疾病结果。因此,保持平衡的免疫反应对于控制CDI至关重要。
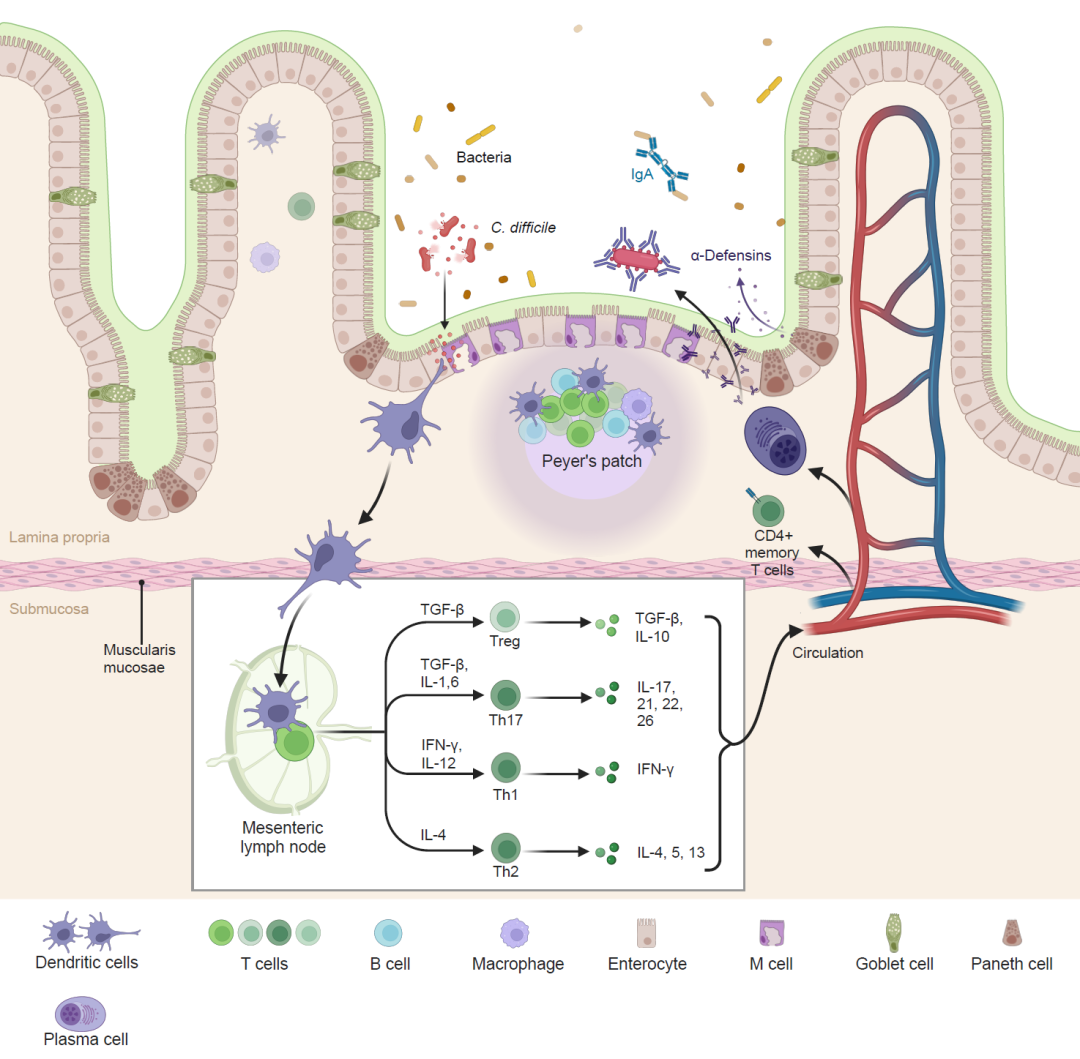
图3. 肠道免疫系统
在健康状态下,杯状细胞分泌黏液,保护肠道上皮细胞免受细菌的侵袭。此外,潘氏细胞释放抗菌肽,包括α-防御素,而IgA被合成以增强对肠腔微生物的防御。次生淋巴器官中的树突状细胞,如佩耶尔氏斑块和肠系膜淋巴结,向未经过刺激的CD4+ T细胞呈递抗原。CD4+ T细胞亚群的分化,包括调节性T细胞(例如,Treg)和Th细胞(例如,Th1、Th2和Th17),受到抗原呈递细胞表型和细胞因子环境(例如,TGF-β和IL-10)等因素的影响。此外,肠道特异性分子被诱导以促进淋巴细胞从全身循环到肠道的归巢。一旦被激活,CD4+ T细胞迁移到肠道粘膜下层执行效应功能。DCs, 树突状细胞; Th, T辅助细胞。
免疫细胞和免疫因子
先天免疫系统包括多种具有进化古老的造血细胞类型,包括树突状细胞(DCs)、单核细胞、巨噬细胞和粒细胞。例如,巨噬细胞将内分泌或旁分泌信号与来自吞噬细胞、巨噬细胞的大泡和细胞外基质中的分子的信号整合在一起。此外,巨噬细胞具有直接与其他组织驻留细胞群、受伤时招募的免疫细胞和细胞外蛋白的表面受体相互作用的能力。
巨噬细胞与粒细胞巨噬细胞集落刺激因子(GM-CSF)和树突状细胞一起,在免疫应答中对CDI起着至关重要的作用。它们捕获和处理C. difficile抗原,迁移至淋巴结,在CDI引起的中性粒细胞招募中发挥作用,并激活特定的T细胞。在淋巴结内,未经刺激的T细胞遇到抗原提呈细胞呈现的加工抗原,导致它们的激活并分化成各种效应T细胞亚群,包括CD4+ T辅助(Th)细胞和CD8+ 细胞毒性T细胞。一旦激活,这些T细胞迁移到CDI部位并执行效应功能来对抗感染。这种涉及不同细胞类型的协调免疫反应对于有效处理CDI至关重要。
在肠道内,类型3固有淋巴细胞(ILC)在感染后恢复肠道完整性中扮演关键角色。具体而言:ILC1s:这些细胞产生干扰素γ(IFN-γ)和肿瘤坏死因子(TNF),有效地抑制细菌复制并帮助控制感染。ILC2s:通过释放细胞因子,包括白细胞介素-4(IL-4)、IL-5、IL-9和IL-33,为免疫防御做出贡献。ILC3s:产生白细胞介素-17A(IL-17A)、IL-17F、IL-22、粒细胞巨噬细胞集落刺激因子(GM-CSF)和TNF等细胞因子,支持抗菌免疫、炎症或组织修复。这些不同ILC亚群的细胞因子谱共同保护免受CDI感染,增强抗菌免疫力,促进组织修复,并抑制肠道内细菌的复制。
此外,其他免疫细胞如γδ T细胞在防御CDI中也起着至关重要的作用:γδ T细胞产生IL-17A:IL-17A促进免疫细胞的招募,增强抗微生物肽的产生,并帮助维持肠道屏障功能,这些都有助于抵抗CDI。ILC缺失导致CDI易感性增加:缺乏ILC会增加对CDI的敏感性,例如缺乏IL-17A或γδ T细胞的小鼠显示出增加组织炎症和死亡率。IL-33的作用:IL-33通过激活结肠ILC2来预防C. difficile相关的死亡和上皮破坏。这种激活作用导致在CDI期间减少促炎细胞因子和增加抗炎细胞因子,从而有助于宿主防御机制。总的来说,这些发现突显了在抵御CDI和维持肠道稳态中,不同免疫细胞亚群和细胞因子之间的复杂相互作用。
自然杀伤(NK)细胞是起源于骨髓的淋巴细胞的一个独特亚群,在对抗病毒和细菌感染以及癌细胞增殖方面发挥着关键作用。它们主要以其细胞毒作用而闻名,针对并消灭感染、转化或受压力的细胞。激活后,NK细胞通过释放含有穿孔素和粒酶的细胞毒颗粒等多种机制诱导靶细胞死亡。此外,NK细胞在激活时可以产生多种细胞因子,包括干扰素γ(IFN-γ)、肿瘤坏死因子α(TNF-α)和各种白细胞介素。这些细胞因子通过影响其他免疫细胞的活动并塑造整体免疫环境,在调节免疫应答中发挥着至关重要的作用。它们可以调节炎症、促进细胞分化、增强抗原呈递,并促进不同免疫系统组分之间的交流,从而有助于对抗病原体和肿瘤的有效免疫应答。
例如,干扰素γ(IFN-γ)刺激抗原呈递细胞上的主要组织相容性复合物II(MHC II)分子的上调,激活巨噬细胞,并增强T细胞反应的初级阶段。此外,NK细胞释放趋化因子,吸引并激活关键的抗原呈递细胞,如树突状细胞(DCs),到组织损伤或感染部位。这一过程通过招募额外的免疫细胞和促进抗原呈递来扩大免疫应答,从而加强对入侵病原体的免疫应答。此外,肿瘤坏死因子α(TNF-α)不仅触发炎症反应,而且直接抑制了C. difficile的增殖,从而减轻了感染。相反,白细胞介素-10(IL-10)由各种免疫细胞亚型产生,包括NK细胞。值得注意的是,研究表明,在CDI后,IL-10缺陷小鼠的结肠炎加剧 [32]。
免疫细胞产生的细胞因子,包括干扰素、肿瘤坏死因子和白细胞介素,在CDI中发挥着双重作用。与IL-6和IL-23等细胞因子相关的17型免疫被认为与严重的CDI有关。来自Th17细胞的IL-17可以增加与CDI相关的死亡率,实验证明,将Th17细胞单独转移至原始小鼠就足以增加死亡率。然而,在小鼠模型中,人体共生真菌白色念珠菌的存在导致了对CDI的易感性降低,因为它增加了IL-17A表达水平。了解免疫细胞、细胞因子与CDI之间的复杂相互作用对于有效地开发调节免疫应答的干预手段至关重要。针对特定免疫通路或使用免疫调节疗法可能有望改善CDI的预防和治疗。
免疫调节因子
在CDI的背景下,Toll样受体(TLRs)在识别特定C. difficile成分的先天免疫细胞中发挥关键作用。这些受体检测细菌表面的病原相关分子模式(PAMPs),启动信号传导途径,激活核因子-κB(NF-κB)并刺激促炎细胞因子的产生。在激活后,CD4+T细胞分化为各种效应子亚型,每种分泌不同的细胞因子,并组织多样化的免疫反应。例如,C. difficile的鞭毛蛋白通过TLR5激活NF-κB和p38丝裂原活化蛋白激酶(MAPK)途径,导致IL-8的产生。CDT和TLR2之间的相互作用在CDI期间对宿主免疫具有重要意义。此外,在CDI期间,小鼠中完整的TLR2/6信号传导上调基因途径,包括NF-κB和MAPK。此外,FMT调节TLR4信号传导途径以治疗CDI。这凸显了微生物病原体、宿主免疫受体和治疗干预之间错综复杂的相互作用在CDI管理中的重要性。NF-κB作为中心转录因子,在调节一系列先天和适应性免疫过程方面发挥作用,并在炎症级联中充当关键介质。在遇到CDI时,NF-κB被激活,促使关键基因的表达,这些基因对炎症、抗微生物防御和组织再生至关重要。这说明了NF-κB在组织宿主对CDI的反应中的重要性,并突显了它作为CDI管理中潜在治疗靶点的作用。
炎性小体代表了复杂的多蛋白复合物,可以检测微生物成分或细胞损伤,最终导致促炎细胞因子的激活。这种激活不仅促使IL-1β和IL-18的释放,还诱导一种称为细胞焦亡的炎症性细胞死亡状态。炎性小体的传感器,包括核苷酸寡聚化结构域(NOD)样受体家族,含有吡啶结构域1(NLRP1)、NLRP3和吡啶,不直接结合配体,而是对由PAMPs或危险相关分子模式触发的特定细胞事件做出反应。最近的研究强调了炎性小体的重要性,特别是吡啶炎性小体,在宿主对CDI的免疫应答中的作用。C. difficile释放的毒素激活了吡啶炎性小体,启动IL-1β的加工和释放,从而增强了炎症反应。
宿主免疫反应的多样性
毒性C. difficile菌株的患病率差异很大,研究表明,高达71%的婴儿和15%的健康成年人可能无症状地携带这些菌株。研究表明,无症状携带者和经历单次CDI但没有复发的个体往往表现出比有症状更强的抗毒素免疫应答能力。目前的研究旨在阐明这些宿主免疫反应差异的机制,以期开发更有效的预防和治疗策略。例如,一项研究表明,含有高浓度IgA的母乳喂养婴儿对毒素具有保护作用。对于轻度C. difficile相关疾病患者,与持续或严重腹泻患者相比,血清IgG抗体水平和粪便IgA抗体水平均较高。观察到抗毒素抗体水平在感染和未感染个体中都可能显著变化,这可以解释尽管携带C. difficile但有症状感染的罕见发生。
免疫疗法的潜在应用
宿主对于C. difficile的防御主要包括三条主要防线:上皮屏障、先天免疫反应和适应性免疫。最初,上皮屏障起着主要的防御作用,但它可能会被毒素破坏,导致后续的非特异性反应。C. difficile通过其毒力因子引发强烈的促炎反应,触发适应性免疫反应,以有效管理感染。GALT和基膜下层的B细胞负责产生IgA,它有效中和C. difficile TcdA,从而防止其对宿主组织的有害影响。这些抗体具有直接中和毒素或通过增强吞噬作用帮助其从体内清除的能力。体液免疫应答涉及各种组分,如T细胞、B细胞和巨噬细胞,整合了先天和适应性免疫机制来对抗CDI。这种协调的免疫反应对于有效对抗C. difficile并降低感染风险至关重要。
艰难梭菌毒素的被动免疫治疗方法
对于C. difficile毒素的被动免疫疗法涉及给予特定抗体,例如单克隆抗体,能够中和细菌产生的毒素。这种方法旨在通过直接针对毒素提供即时保护和减轻CDI的严重程度。因此,它为治疗和预防C. difficile相关疾病提供了一种有前景的策略。Actotoxumab和bezlotoxumab(BEZ)是人源单克隆抗体,能够分别结合并对抗TcdA和TcdB的作用。特别是,BEZ靶向TcdB,从而中和其对人类细胞的有害影响。一项于2015年至2019年进行的研究比较了BEZ与标准治疗(通常使用万古霉素或费达霉素)缓解CDI症状的疗效。该研究共招募了107名参与者,其中BEZ组54名,标准治疗组53名。结果显示,BEZ组在90天内复发CDI的发生率显著低于标准治疗组(11% vs 43%,p < 0.001)。此外,接受BEZ治疗的患者90天内的重返院率也显著低于标准治疗组(40% vs 64%,p = 0.011)。值得注意的是,BEZ治疗显示出显著减少CDI发生率的能力,而且没有发现任何安全问题。与这一发现一致的是全球III期试验MODIFY(MOnoclonal antibodies for C. DIFfficile therapY)I和MODIFY II显示,与接受抗生素治疗CDI的人群相比,BEZ有效地降低了CDI复发率。值得注意的是,使用actoxumab单独使用无效,当与BEZ联合使用时也未增强其有效性。
传统免疫疗法利用抗体对抗感染,而基因疗法代表了一种创新的范式,通过操纵遗传物质增强对CDI的免疫防御。最近,基因疗法已被用于实现长期抗体表达。在一项研究中,研究人员设计了一种新型的三重突变腺病毒(AAV)载体,命名为AAV6.2FF,用于在小鼠和仓鼠中产生actotoxumab或BEZ。在血清中,这两种抗体的检测水平均超过90 μg/mL,并且在两种动物模型的粘膜表面也观察到它们。重要的是,在所有接受AAV6治疗的小鼠中观察到了抗体的成功表达。这些发现为开发更有效的CDI治疗方法提供了一个有前景的途径。通过将诸如BEZ之类的单克隆抗体与创新的基因疗法技术相结合,研究人员有可能增强CDI的管理和预防。这种综合方法有望通过提供长期和有针对性的抗体表达,从而提供更好的保护,改善患者预后,并减轻疾病的负担。
Kelly等人在2020年进行的研究为了解自然产生的抗体对抗复发性C. difficile感染的毒素,特别是TcdB。他们的研究发现,对抗C. difficile毒素B的抗体水平较高的个体更不容易出现复发性CDI。这一发现与第三期MODIFY试验中观察到的结果相一致,该试验评估了BEZ和actoxumab在预防复发性CDI中的有效性。此外,MODIFY试验观察到,在CDI期间给予BEZ可以预防全身性疾病和胸腺萎缩,而不影响肠道损伤,这一发现也很值得注意。这表明BEZ可能在肠道内部毒素中性化之外发挥着全身性的效应。了解这些全身性效应对于阐明BEZ和其他CDI治疗干预的完整作用机制至关重要。
人源化单克隆抗体针对C. difficile产生的特定毒素是管理CDI的一种有前景的方法。分别针对TcdA和TcdB的PA-50和PA-41抗体在结合其各自毒素的特定区域方面表现出很高的效力。在仓鼠CDI模型进行的临床前研究中,PA-50和PA-41抗体的联合治疗显示出比标准抗生素治疗显著更高的长期存活率。此外,Koon等人的研究进一步阐明了这些抗体的作用机制。他们发现,利用人源单克隆抗体(TcdA为MK3415,TcdB为MK6072)中和TcdA和TcdB有效阻断了这些毒素在人结肠黏膜和人外周血单核细胞中引发的先天免疫反应。这些发现强调了利用单克隆抗体靶向C. difficile毒素的治疗潜力,以减轻CDI的有害影响。通过中和TcdA和TcdB,这些抗体可能防止与CDI相关的炎症反应和组织损伤的激活,最终改善患者的临床症状。
孢子化过程的被动免疫疗法
CDI的复发受到C. difficile孢子萌发的显著影响,这些孢子是疾病传播的主要载体。C. difficile可以通过使用蛋白酶和鞭毛附着于粘液层并侵入肠细胞。这种由营养匮乏引发的孢子形成过程发生在停滞阶段。在小肠中,孢子萌发的最佳条件包括较高水平的胆酸盐和较低水平的胆酸醇,导致产生营养细胞。C. difficile孢子与肠黏膜的相互作用在穿过肠道屏障过程中至关重要,是疾病复发的关键因素。研究人员已经确定了特定的宿主分子、细胞受体以及对孢子进入肠上皮细胞(IECs)至关重要的孢子表面配体。值得注意的是,位于孢子外壳层的三种胶原样蛋白(BclA1、BclA2和BclA3)发挥着重要作用。特别是BclA3,被确定为抗原表位,在大肠杆菌中过度表达,并在小鼠中用作免疫原。结果显示,BclA3诱导了特异性IgG产生,并在接触C. difficile孢子后部分缓解了CDI症状。这些发现表明BclA3具有作为CDI疫苗免疫原的潜力。
在C57BL/6小鼠中给予抗孢子IgY,在CDI发生前和期间已经显示出显著效果,表明具有潜在的治疗益处。首先,腹泻发作延迟1.5天暗示着疾病进展的潜在减缓,为干预和管理提供了途径。其次,观察到结肠黏膜孢子粘附的显著减少90%,表明C. difficile的定植和侵袭减少,这是CDI发展的关键步骤。此外,在复发模型中,抗孢子IgY与万古霉素的联合给药导致复发性腹泻延迟平均2天。这表明抗孢子IgY可能对初次CDI和复发CDI具有保护作用,无论是单独使用还是与标准抗生素治疗结合使用。此外,利用枯草芽孢杆菌PXN21孢子在小鼠模型中的研究显示了令人鼓舞的结果。这些孢子通过上调TLR2的表达激活先天免疫,TLR2在识别微生物成分并启动免疫反应中起着关键作用。TLR2的激活导致促炎细胞因子如IL-6和TNF-α的释放,可以帮助抑制CDI症状,预防感染的初次发生和复发。然而,值得注意的是,尽管这些发现令人鼓舞,但仍需要进一步的研究,包括在人类受试者中进行临床试验,以验证抗孢子IgY和枯草芽孢杆菌PXN21孢子作为CDI潜在治疗干预手段的有效性和安全性。
抗鞭毛免疫治疗
最近的研究表明,C. difficile的鞭毛在引发免疫反应方面起着重要作用,进一步强调了免疫应答在对抗由C. difficile引起的定植和相关疾病中的重要性。CDI的发展在很大程度上依赖于C. difficile对宿主的定植,其中鞭毛在各种过程中起着关键作用,如定植、附着、生物膜形成和毒素产生,可能增强某些菌株的毒力。作为肠道病原体,C. difficile利用其表面的鞭毛蛋白增强运动性、定植和附着于宿主肠道,从而有助于其致病性。包括FliC等粘附蛋白和鞭毛帽蛋白FliD在内的鞭毛蛋白与TLR5等蛋白酶相互作用,促进病原体附着于粘膜。这种相互作用触发了NF-κB和MAPK等信号通路的激活,导致促炎细胞因子的产生和炎症反应的启动。研究表明,利用基于鞭毛的蛋白作为免疫疗法的抗原在对抗CDI方面显示出潜力。小鼠实验的研究表明,免疫接种鞭毛蛋白后接种CDI会产生更强的毒素免疫力,导致病原体在粪便中显著减少。此外,Pechine等人的研究观察到,CDI患者的活跃期抗体水平低于对照组,暗示了这些抗体在对抗CDI中的潜在作用。
对抗CDI的疫苗疗法
以往的研究突显了在开发疫苗以对抗CDI方面的重大努力,这是由抗生素耐药性和复发性感染所带来的挑战。赛诺菲巴斯德和其他研究机构正在积极研究针对CDI风险人群的疫苗候选物,如住院患者或长期使用抗生素的患者。赛诺菲巴斯德的疫苗旨在预防原发性CDI,目前正在进行II期临床试验(NCT00772343和NCT01230957),在减少易感人群中CDI负担方面表现出了潜能。此外,C. difficile毒素疫苗(批号05C02)在I期试验中表现出安全性和有效性,支持其进一步开发用于预防目的。在日本,井上等人进行的I期试验发现C. difficile疫苗在日本健康老年人群中安全且耐受良好。尽管辉瑞最近基于毒素的疫苗在III期临床试验中失败,但重组毒素基肽和与细胞表面相关的抗原仍然是未来CDI预防工作中至关重要的疫苗候选物。
此外,当前的研究正强调黏膜疫苗的重要性,这对于成人和儿童来说都可能具有优势,考虑到黏膜免疫在对抗CDI中的关键作用。最近的研究表明,开发一种活性的、非毒性的疫苗是预防CDI的可行策略。为此,作为孢子抗原的CdeM蛋白已被选择作为黏膜免疫疫苗开发中的载体。已证实口服给予这种抗原可在小鼠中诱导免疫应答以对抗CDI。在英国,Bradshaw等人发现脂蛋白CD0873的特定特征具有作为针对C. difficile的疫苗的潜力。给予重组脂蛋白CD0873的小鼠显示出由于发展出强烈的分泌性IgA免疫应答而长期预防了C. difficile在肠道中的定植。这些发现对于开发针对C. difficile的疫苗具有重要意义,并可能推进CDI的治疗和预防策略。
肠道微生物群介导的CDI免疫反应
肠道微生物群在维持肠道平衡和调节免疫反应方面发挥着至关重要的作用。肠道中的某些有益菌可以激活免疫系统,刺激抗微生物肽的产生,并增强病原体监测,从而提高整体免疫功能。旨在恢复健康的肠道微生物群的干预措施,如益生菌、FMT或益生元,在改善CDI患者的治疗结果、加强宿主免疫力以及减少复发感染风险方面显示出潜力。
肠道固有免疫系统协调着各种防御机制来对抗CDI。这种复杂的防御涉及多个组分之间的复杂相互作用,旨在保持肠道完整性和稳态(图4)。关键成分包括肠上皮细胞(IECs)、巨噬细胞、单核细胞、肥大细胞、ILCs和DCs。IECs覆盖着肠道黏膜,形成对抗病原体和毒素的物理屏障。它们包括负责营养吸收和屏障功能的吸收上皮细胞、产生保护性黏液的杯状细胞、分泌抗微生物肽的泌乳细胞,以及产生5-羟色胺并影响肠道蠕动和免疫反应的肠色素细胞。参与肠道固有免疫的免疫细胞包括巨噬细胞,它们是吞噬病原体和细胞碎片的吞噬细胞,循环的单核细胞,能够分化为巨噬细胞或DCs,释放组胺和其他介质以应对病原体或过敏原的肥大细胞,以及ILC1s、ILC2s和ILC3s等ILCs,它们有助于免疫调节、组织修复和抗感染。此外,DCs作为抗原呈递细胞调节适应性免疫反应。
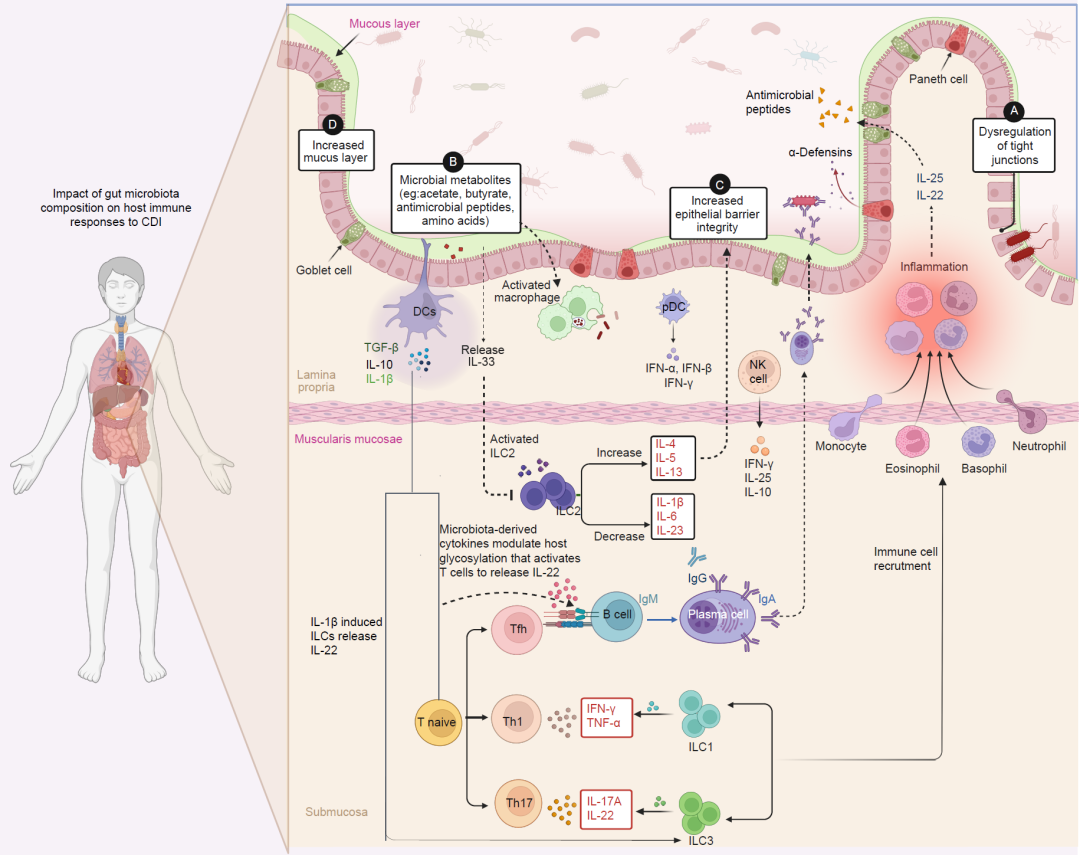
图4. 肠道微生物群的组成对宿主对CDI的免疫反应产生了深远的影响
在艰难梭菌感染(CDI)的患者中。肠道完整性受影响促进了微生物穿过肠道屏障的易位,导致系统性免疫失调。微生物代谢产物通过树突状细胞和巨噬细胞激活免疫应答,促进炎症T细胞亚型(如Th1、Th17和Tfh细胞)的分化。B细胞不仅产生抗体,还协调对抗CDI的免疫应答。此外,B细胞还可以被细胞因子IL‐22直接激活,诱导它们分化为产生保护性分泌型IgA(sIgA)的浆细胞,以及潜在的病理性自身抗体。肠道菌群通过产生代谢产物如短链脂肪酸(SCFAs)、抗菌肽和氨基酸来调节艰难梭菌的生长,发挥了对CDI的保护作用。这些代谢产物可以通过IL‐33调节ILCs。IL‐33有助于抑制促炎细胞因子(如IL‐1β、IL‐6和IL‐23),同时促进抗炎细胞因子(如IL‐4、IL‐5和IL‐13)的产生。因此,ILC2的激活强化了上皮屏障紧密连接,而ILC1通过分泌IFN‐γ和肿瘤坏死因子TNF‐α增强了局部免疫应答来对抗感染。在IL‐1β的影响下,ILC3和Th17细胞都可以诱导产生IL17A和IL‐22。泛上皮细胞和腺细胞分泌的α‐防御素作为释放到肠腔的抗菌肽的重要来源,有助于调节肠腔细菌的组成。Tfh:T细胞辅助型;SCFAs:短链脂肪酸;ILCs:固有淋巴细胞;NK细胞:自然杀伤细胞;pDC:专业树突状细胞。
ILCs在恢复感染后的肠道完整性中扮演着关键角色。微生物群对ILCs的刺激促使它们释放细胞因子,招募宿主免疫系统来对抗感染。已经证明,微生物群产生的乙酸通过协同调节中性粒细胞和ILC3s,增强了对C. difficile的固有反应。这种调节促进了中性粒细胞中的炎症小体激活,并增加了ILC3s中IL-1受体的表达,最终在IL-1β刺激下增加IL-22的分泌,以抵消CDI。
微生物群产生的细胞因子在保护粘膜屏障免受CDI方面发挥着关键作用。在小肠黏膜基质中由γδ T细胞分泌的IL-17和IL-22受到肠道微生物群的刺激。通过肠道微生物群的定殖,粘膜IL-22信号的激活调节了宿主N-连接的糖基化,促进了消耗琥珀酸的细菌(如澳洲考拉杆菌属)的增长。这种代谢改变阻止了C. difficile在肠道中的增殖。FMT已被证明是一种成功的治疗策略,用于复发性和严重的CDI。发现FMT恢复了肠道微生物群的组成,导致细胞因子和抗体水平增加。成功的FMT治疗与TcdB特异性Th17细胞的比例增加相关,这些细胞对抗C. difficile毒素至关重要。此外,FMT后观察到针对TcdA和TcdB的IgG和IgA抗体水平增加,对于中和C. difficile毒素至关重要。研究表明,FMT可使结肠组织中IL-25水平升高,这是一种由微生物群产生的细胞因子。IL-25通过招募嗜酸粒细胞并选择性降低有害的IL-23来减轻CDI相关的死亡和组织损伤,从而调节炎症反应。IL-33治疗通过降低与CDI相关的炎症反应,减少例如IL-1β、IL-6和IL-23等促炎细胞因子的产生,同时增加抗炎细胞因子的产生,例如IL-4、IL-5和IL-13。IL-33的保护作用依赖于ILC2s,这些细胞产生IL-13。IL-13对于招募或将这些细胞转化为巨噬细胞至关重要。中和假性受体IL-13Rα2可以保护免受CDI,突显了IL-13信号在防御感染中的重要性。
人体肠道微生物群在合成对人类健康产生深远影响的必需代谢产物方面发挥着至关重要的作用。包括醋酸、丙酸和丁酸在内的短链脂肪酸(SCFAs)是研究最为充分的代谢产物,以其在宿主免疫稳态和健康中的作用而闻名。SCFAs通过肠道细菌对膳食纤维和复杂碳水化合物的发酵产生,并提供多种有益效果。这些效果包括刺激肠道IgA反应,促进宿主与微生物组之间的相互沟通。此外,它们调节结肠pH以维持免疫稳态,并调节哺乳动物结肠上皮细胞的增殖、分化和基因表达。其中,醋酸是SCFAs之一,进入系统循环并对单核细胞、T细胞和中性粒细胞等不同免疫细胞发挥免疫调节作用。它影响抗氧化剂和氧化剂之间的平衡,并影响这些免疫细胞中的细胞因子产生。另一方面,丁酸通过抑制促炎细胞因子和减少白细胞募集在炎症部位而表现出抗炎特性。此外,它增加了调节性T细胞的数量,促进耐受性CD103+ DCs的发育,从而减轻肠炎症,并增强肠道屏障功能,这些结果已经在CDI小鼠实验中得到证实。
除了SCFAs之外,肠道微生物群还有助于合成氨基酸、维生素和次生胆酸,这些都在宿主生理和免疫功能中发挥着至关重要的作用。例如,某些肠道细菌产生维生素,如维生素K和B族维生素,对于各种代谢过程和免疫功能至关重要。次生胆酸是由肠道细菌对原始胆酸进行生物转化而产生的,被认为参与调节宿主代谢和免疫应答。
结 论
宿主免疫反应在对抗CDI(一种致病菌引起的胃肠感染)中发挥着至关重要的作用。通过诸如产生抗菌肽、吞噬和消除病原体、调节炎症反应等一系列机制,免疫系统能够识别、清除和预防由该致病菌引发的感染。此外,免疫系统还促进有益肠道细菌的增殖,以维持肠道内稳态,减轻CDI的发生或复发的可能性。因此,加强宿主免疫反应是预防和治疗CDI的基石。通过理解和促进免疫系统与肠道微生物群之间的相互作用,我们可以制定更有效的预防和控制策略。
引文格式:
Lamei Wang, Javier A. Villafuerte Gálvez, Christina Lee, Shengru Wu, Ciaran P. Kelly, Xinhua Chen, Yangchun Cao. 2024. Understanding host immune responses in Clostridioides difficile infection: Implications for pathogenesis and immunotherapy. iMeta 3, e200. DOI: 10.1002/imt2.200
作者简介

王腊梅(第一作者)
● 西北农林科技大学博士,哈佛大学博士后。
● 主要研究病原菌对宿主消化道微生物及机体免疫的影响,应用动物模型结合多组学(宏基因组、蛋白质组、代谢组等)关联分析技术系统阐明梭菌及其毒素的致病机理,解析梭菌调控微生物区系及其机体免疫的机制与网络。相关学术成果已发表于Gastroenterology、PNAS、Clinical Infectious Diseases、Microbiology Spectrum等期刊。

曹阳春(通讯作者)
● 西北农林科技大学教授,博士生导师。
● 研究方向为消化道微生物与营养物质高效利用,已在Gastroenterology、Microbiome、Microbiology Spectrum、iMeta等期刊发表学术论文50余篇,主持国家自然科学基金项目、国家重点研发子课题等30余项,以第一完成人授权国家专利11项,以第一完成人发布团体标准和陕西省地方标准各2项,获陕西省科技进步一等奖和甘肃省科技进步二等奖各1项。获聘哈佛医学院营养中心专家组成员,入选中国畜牧兽医学会第三届“青年拔尖人才”、2022年仲英青年学者、陕西省杰出青年科学基金。兼任全国动物营养指导委员会肉牛营养分会委员,中国畜牧兽医学会动物营养学分会副秘书长,动物微生态与生物饲料专业委员会陕西省分会副主任,陕西省添加剂预混料“四主体一联合”技术工程中心主任,陕西省秦创原畜禽饲料添加剂研发与应用团队首席科学家等。
更多推荐
(▼ 点击跳转)
iMeta | 引用7000+,海普洛斯陈实富发布新版fastp,更快更好地处理FASTQ数据
iMeta | 德国国家肿瘤中心顾祖光发表复杂热图(ComplexHeatmap)可视化方法

1卷1期

1卷2期

1卷3期

1卷4期

2卷1期

2卷2期

2卷3期

2卷4期

3卷1期

2卷2期封底

2卷4期封底

3卷2期
期刊简介
“iMeta” 是由威立、肠菌分会和本领域数百位华人科学家合作出版的开放获取期刊,主编由中科院微生物所刘双江研究员和荷兰格罗宁根大学傅静远教授担任。目的是发表原创研究、方法和综述以促进宏基因组学、微生物组和生物信息学发展。目标是发表前10%(IF > 20)的高影响力论文。期刊特色包括视频投稿、可重复分析、图片打磨、青年编委、50万用户的社交媒体宣传等。2022年2月正式创刊发行!目前期刊已经被ESCI、Scopus等数据库收录。
联系我们
iMeta主页:
http://www.imeta.science
姊妹刊iMetaOmics主页:
http://www.imeta.science/imetaomics/
出版社iMeta主页:
https://onlinelibrary.wiley.com/journal/2770596x
出版社iMetaOmics主页:
https://onlinelibrary.wiley.com/journal/29969514
iMeta投稿:
https://wiley.atyponrex.com/journal/IMT2
iMetaOmics投稿:
https://wiley.atyponrex.com/journal/IMO2
邮箱:
office@imeta.science


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









