







第一章 肌肉活动
第一节 细胞生物电现象
一、刺激、反应与兴奋
1、
刺激
泛指能够引起机体或细胞发生反应的环境变化。刺激可依据其性质不同分为物理性刺激、化学性刺激和生物性刺激。
2、机体或细胞受到刺激后所发生的功能活动的变化,称为反应。反应可分为两种:一种是由相对静止变为活动状态,或活动增强,称为兴奋;另一种是由活动变为相对静止状态,或活动减弱,称为抑制。
3、刺激引起反应的条件
①一定的刺激强度
②一定的持续时间
③一定的“强度-时间”变化率
刺激的这三个条件可以互相影响,其中一个或两个条件的数值改变时,其他条件的数值也会发生相应的改变。
当刺激的持续时间和强度变化率都固定时,引起组织发生反应的最小刺激强度称为
阈强度或
阈值。阈值或阈强度是评定神经肌肉兴奋性的最简易指标。
能够触发细胞兴奋产生动作电位的临界膜电位,称为
阈电位
阈强度的刺激称为
阈刺激,小于强度的刺激称为
阈下刺激,大于阈强度的刺激称为
阈上刺激。阈刺激和阈上刺激称为有效刺激。
细胞受到阈下刺激时,在细胞膜上产生的局部去极化,其电位变化不能向远处扩布,因此称为
局部反应。
兴奋
是生物体的器官、组织或细胞受到足够强的刺激后所产生的生理功能加强的反应,如神经冲动的发放、肌肉的收缩、腺体的分泌等。
神经、肌肉和内分泌腺细胞则能产生可传播的动作电位,这些细胞被称为
可兴奋细胞。
兴奋性是指 机体感受刺激后发生兴奋反应的能力或特性,它是在新陈代谢基础上产生 的,是机体生命活动的基本特征之一。
兴奋性是指 机体感受刺激后发生兴奋反应的能力或特性,它是在新陈代谢基础上产生 的,是机体生命活动的基本特征之一。
二、细胞生物电活动
(一)静息电位
1、
静息电位是指
细胞未受刺激时存在于细胞膜两侧的电位差(内负外正)。由于这一电位差存在于安静细胞膜的两侧,故又称为跨膜静息电位或膜电位。
静息电位表现为膜内比膜外电位低,即膜内带负电荷而膜外带正电荷。这种安静时存在于膜两侧的稳定的“内负外正”的状态,称为
极化(状态)。以极化(或静息电位)为基准,膜内负电位增大,称为
超极化;膜内负电位减小,称为
去极化或
除极化;细胞发生去极化后,膜电位又恢复到极化状态,称为
复极化。
2、静息电位的形成机制
静息电位主要是K⁺外流所形成的
电-化学平衡电位。细胞膜在安静时,对K⁺的通透性最大,对Na⁺和CL⁻的通透性很小,对A⁻(有机离子)几乎不通透。因此,K⁺便顺着浓度差向膜外扩散,使膜外具有较多的正电荷;
膜内的A⁻虽有随K⁺外流的倾向,但因不能透过膜而被阻留在膜的内侧面,使膜内具有较多的负电荷。这就造成膜外为正、膜内为负的
极化状态;
由K⁺外流造成的这种以膜为界的内负外正的电位差,将成为阻止K⁺外流的力量。随着K⁺外流的增加,阻止K⁺外流的电位差也增大。当促使K⁺外流的浓度差和阻止K⁺外流的电位差两种拮抗力量达到平衡时,将不再有K⁺的净移动。此时,膜两侧内负外正的电位差将稳定于某一数值不变,即
K⁺的平衡电位,也就是静息电位。
(二)动作电位
1、
动作电位:是指
细胞受到
有效
刺激而兴奋时,细胞膜在原来静息电位的基础上发生的一次迅速、短暂、可向周围扩布的电位波动。动作电位的产生是细胞兴奋的标志。
2、
动作电位的形成机制.
①
上升支:当细胞受到刺激而兴奋时,Na⁺通道大量开放,膜对Na⁺的通透性突然增大并超过对K⁺的通透性,于是细胞外的Na⁺便顺着浓度差和电位差迅速内流,导致膜内电位急剧上升,即膜内负电位快速消失并转为正电位。当膜内正电位增大到足以阻止由电位差所推动的Na⁺内流时,Na⁺的净内流停止。此时膜两侧的电位差即为Na⁺的平衡电位,其电位值与动作电位的峰值基本一致。可见,动作电位的上升支主要是细胞外Na⁺快速内流造成的。
②
下降支:当膜去极化到达峰值时,Na⁺通道迅速失活关闭,此时,膜对K⁺的通透性增大,于是膜内的K⁻顺浓度差和电位差外向扩散,使膜内电位迅速下降,直至膜复极化到静息电位水平。可见,动作电位的下降支主要是细胞内K⁺外流造成的。
③
复极化:此时,膜对K⁺的通透性恢复正常,Na⁺通道失活状态解除,并恢复到可激活状态。钠泵激活,将进入膜内的Na⁻泵出细胞,同时把扩散到膜外的K⁻泵入细胞,从而恢复静息时细胞内外的离子分布,以维持细胞的正常兴奋性。
3、动作电位的特点
①有“全或无”现象:单一神经或肌细胞动作电位的一个重要特点是若刺激达不到阈值不会产生动作电位,刺激一旦达到阈值就会爆发动作电位,动作电位一旦产生,其大小和形状不再随着刺激的强弱和传导距离的远近而改变。
②有绝对不应期:由于绝对不应期的存在,动作电位不可能发生融合。
4、动作电位的传导
在膜上任何一处产生的动作电位都将沿着整个细胞膜扩布,即传导。沿着神经纤维传导的动作电位呈脉冲式的锋电位,称
神经冲动。
神经纤维传导兴奋的特征(神经纤维上传导动作电位的特点)
①
生理完整性:神经传导首先要求神经纤维在结构和生理功能上都是完整的。由于一些原因致使神经纤维局部结构或机能发生改变,神经的传导则中断。
②
双向传导:刺激神经纤维上任何一点,所产生的神经冲动可沿纤维向两侧方向传导,这是因为局部电流可向两侧传导的缘故。
③
不衰减和相对不疲劳性:在传导过程中,锋电位的幅度和传导速度不因传导距离增大而减弱,也不因刺激作用时间延长而改变。这是因为神经传导的能量来源于兴奋神经本身。
④
绝缘性:神经干内含有许多神经纤维,神经传导各行其道互不干扰;绝缘性主要由于髓鞘的存在。
第二节 肌肉收缩原理
一、肌肉的微细结构
肌肉是由成束排列的肌细胞组成,肌细胞外形呈长圆柱形状,又称
肌纤维,
是肌肉结构和功能的基本单位。
每条肌纤维外面被一层薄膜所包裹,这层薄膜称
肌膜
,肌膜相当于细胞膜,肌膜内有肌浆和多个细胞核。
(一)肌原纤维的结构
1、肌原纤维呈长纤维状,纵贯于肌纤维全长,直径约1~2μm。在显微镜下可见每条肌原纤维全长都呈现有规则的明暗交替,分别称
明带和暗带。
2、在平行排列的各肌原纤维之间,明带和暗带又分布在同一水平上,这就使肌纤维呈现横纹,
故骨骼肌被称为横纹肌。
3、在肌原纤维上,暗带长度比较固定,其中间有一个较透明的区域,为H区,H区中间有一横向暗线称
M线;明带长度可变,其中央有一条横向的暗线称
Z线。
4、两相邻Z线间的一段肌原纤维称为
肌小节,它包括中间的暗带和两侧各1/2的明带。肌小节是由更微细的平行排列的粗肌丝和细肌丝组成的。肌丝及其支持结构是肌原纤维的结构基础。
5、肌球蛋白分子分成两束,每束肌球蛋白分子的长杆部朝向M线而横向聚合,形成粗肌丝主干,而分子的球状头部,则有规则地突出在M线两侧的粗肌丝主干表面,形成所谓
横桥。
(二)肌管系统的结构
肌管系统是指包绕在每一条肌原纤维周围的膜性囊管状结构,他们实际是由功能不同的俩组独立的管道系统组成。包括:
1.横管系统
横管,又称T管,走向和肌原纤维相垂直。它由肌膜向细胞内凹入而成,凹入位置与各Z线水平,横穿于肌原纤维中肌节之间,呈环状环绕每条肌原纤维,同一水平的横管互相沟通,横管内腔与细胞外液相通,其
作用是
将肌细胞兴奋时出现在细胞膜上的电位变化传入细胞内。
2、纵管系统
纵管,又称L管,走向和肌原纤维平行。纵管包绕每个肌小节的中间部分,在近横管时管腔膨大成终池。每一横管和两侧的终池构成所谓
三联管结构。
纵管和终池是Ca2+的储存库,在肌肉活动时实现Ca2+的储存、释放和再积聚。三联管是把肌细胞膜的电变化和肌细胞的收缩过程耦联起来的关键部位。
二、肌肉收缩与舒张过程
在肌肉收缩和舒张过程中,与肌丝滑行有关的蛋白质,称为
肌肉收缩蛋白,
包括肌球蛋白和肌动蛋白。
(一)兴奋在神经-肌肉接点的传递(乙酰胆碱)
1、
神经-肌肉接点:
是实现兴奋由运动神经传递到肌肉的装置,肌肉的收缩是由运动神经以冲动的形式传来刺激引起的。运动神经纤维在到达所支配的骨骼肌时发出分支,形成末端膨大的神经末梢。神经末梢与肌纤维接触前失去髓鞘,再以裸露末梢嵌入肌膜上,被称为终板膜在凹陷中,形成所谓的
神经-肌肉接点。
2、
传递机制:通过化学递质乙酰胆碱(Ach)和终板膜电位变化来实现。
3、
传递过程
①当运动神经元兴奋时,神经冲动沿运动神经纤维传至轴突末梢,并刺激接点前膜。接点前膜去极化使膜上的钙通道开放,使得细胞外液中的Ca2+进入接点前膜,触发轴浆中的囊泡向接点前膜的内侧面靠近。
②囊泡与接点前膜融合,其中所含的乙酰胆碱被释放进入突触间隙,随后立即与突触后膜的乙酰胆碱受体结合,引起突触后膜的 Na+和K+的通透性改变,突触后膜除极化,形成终板电位。终板电位通过局部电流作用,使邻近肌细胞膜去极化而产生动作电位,实现了兴奋由神经传递给肌肉。
③由于突触间隙中和终板膜上有大量的胆碱酯酶,在它的作用下每次冲动从轴突末梢释放的乙酰胆碱,在短时间内(2ms)全部水解失活,从而维持神经-肌肉接头下次正常的传递功能。
4、兴奋在神经-肌肉接点传递的特点:
(1)
化学传递:
神经和肌肉之间的兴奋传递是通过化学递质乙酰胆碱进行传递的
(2)
兴奋传递节律是1对1的:
即每一次神经纤维兴奋都可引起一次肌肉细胞兴奋。因为神经末梢每次动作电位所引起的乙酰胆碱释放量相当大,足以产生较大的终板电位,从而激发肌肉细胞兴奋。
(3)
单向传递:
兴奋只能由神经末梢传递给肌肉,而不能反向传递
(4)
时间延搁:
兴奋的传递要经历递质释放、扩散和作用等多个环节,因而传递速度缓慢
(5)
高敏感性:
易受化学和其他环境因素变化的影响,易疲劳。
(二)肌肉兴奋-收缩耦联 (Ca²⁺是媒介物)
肌细胞兴奋过程是以膜的电变化为特征的,而肌细胞的收缩过程是以肌纤维机械变化为基础,它们有着不同的生理机制,肌肉收缩时存在某种中介过程把它们联系起来,这一中介过程称为
肌肉的兴奋-收缩耦联。
包括以下三个过程/步骤:
1、电兴奋通过横管系统传向肌细胞深处
2、三联管结构处的信息传递
3、肌浆网中Ca²⁺释放入胞质以及Ca²⁺由肌质向肌浆网再聚积。
详细过程:当肌细胞兴奋时,动作电位可沿着凹入肌细胞内部的横管系统传导,深入到三联管结构和肌小节近旁。兴奋传至三联管后,引起横管膜去极化,致使终池上Ca²⁺释放通道大量开放,终池中 Ca²⁺顺浓度梯度迅速进入肌浆中,使肌浆Ca²⁺浓度比静息时提高了100倍,实现了肌细胞的兴奋-收缩耦联,Ca²⁺被认为是肌细胞兴奋-收缩耦联的媒介物。
(三)肌细胞的收缩与舒张过程
肌丝滑行理论认为
1、肌肉收缩时,虽然外观上可以看到整个肌肉缩短,但在肌细胞内并无肌丝和它们所含的分子结构的缩短,而只是在每个肌小节内发生细肌丝向粗肌丝之间的滑行
2、出现明带的长度缩短,而暗带长度不变,相应H区变窄。即由Z线发出的细肌丝在某种力量作用下主动向暗带中央移动
3、结果各相邻Z线都相互靠近,肌小节长度变短,造成整个肌原纤维、肌细胞、乃至整条肌肉长度缩短。
4、从分子水平上来看
肌肉收缩实际上是构成粗肌丝的肌球蛋白和构成细肌丝的肌动蛋白相互作用的结果,而细肌丝上的原肌球蛋白和肌钙蛋白则起着控制作用。
基本过程:
(1)当肌细胞兴奋动作电位引起肌浆Ca²⁺的浓度升高时,Ca²⁺与细肌丝上肌钙蛋白结合,引起肌钙蛋白分子构型发生变化,这种变化又传递给原肌球蛋白分子,使后者构型亦发生变化。其结果,原肌球蛋白从肌动蛋白双螺旋结构的沟沿滑向沟底,安静时抑制肌动蛋白和横桥结合的因素被接触,暴露出肌动蛋白上能与横桥结合的位点。
(2)横桥与肌动蛋白结合形成肌动球蛋白。肌动球蛋白可激活横桥上ATP 酶活性,在Mg²⁺参与下,ATP 分解释放能量,引起横桥头部向粗肌丝中心方向摆动,牵引细肌丝向粗肌丝中央滑行。当横桥角度发生变化时,横桥头部与肌动蛋白解脱,并恢复到原来垂直的位置。紧接着横桥又开始与下一个肌动蛋白的位点结合,重复上述过程,进一步牵引细肌丝向粗肌丝中央滑只要肌浆中Ca²⁺浓度不下降,横桥循环运动就不断进行下去,将细肌丝逐步拖向粗肌丝中央,于是,肌小节缩短,肌肉出现缩短。
(3)当刺激终止后,终池膜对Ca²⁺通透性降低,Ca²⁺释放也停止。肌浆膜上的钙泵迅速回收Ca²⁺,使肌浆 Ca²⁺浓度下降,钙与肌钙蛋白结合解离,肌钙蛋白恢复到原来构型,继而原肌球蛋白也恢复到原来构型,肌动蛋白上与横桥结合的位点被重新掩盖起来,横桥与肌动蛋白分离,粗、细肌丝退回到原来位置,肌小节变长,肌肉产生舒张。
三、兴奋在神经纤维与在神经肌肉点头处传播的区别(补充)
(1)两者兴奋传播的机制不同。兴奋在神经纤维传播是以电信号的方式进行的,是建立在局部电流学说机制上的,兴奋在神经—肌肉接点处传播是以化学信号进行的
(机制)
(2)兴奋在神经纤维上传导是双向性的,而在神经--肌肉接点传播是单向性的
(方向)
(3)在神经纤维传播速度快,而在神经肌肉接点传播存在较长时间延搁
(速度)
(4)兴奋在神经纤维传导具有相对不疲劳性,而在神经--肌肉接点传播者易疲劳,并受药物和环境因素的影响
(疲劳)
第三节 肌肉的收缩形式与力学特征
不依赖于前、后负荷,可影响肌肉收缩效果的肌肉内在功能状态称为
肌肉收缩能力。
一、肌肉的收缩形式
依肌肉收缩的张力和长度变化,将肌肉收缩的形式分以下三类:
(一)缩短收缩
①
缩短收缩:
指肌肉收缩所产生的张力大于外加的阻力时,肌肉缩短,并牵引骨杠杆做相向运动的一种收缩形式。缩短收缩时,肌肉起止点互相靠近,又称
向心收缩。
②力学特征:
A、缩短收缩时,因负荷移动方向和肌肉用力方向相一致,肌肉做正功;
B、缩短收缩时肌肉起止点相互靠近,又称向心收缩。
③应用:如运动练习中的前臂弯举、高抬腿跑、挥臂扣球等练习,参与工作的主动肌就是做缩短收缩。
依据整个关节运动范围肌肉张力与负荷的关系,缩短收缩又有非等动收缩和等动收缩。
①非等动收缩(等张收缩)。是肌肉
克服恒定负荷的一种收缩形式。由于不同关节角度杠杆得益不同和受肌肉收缩长度变化的影响,在整个关节范围内,肌肉收缩产生的张力和所遇负荷阻力是不同的,收缩速度也不相同。等张收缩发展力量只有关节力量最弱点得到最大锻炼。
应用:例如屈肘举起恒定负荷时,肱二头肌的张力在关节角度115度-120度时最大,关节角度30度时最小。
②等动收缩是通过专门的等动练习器来实现的。该器械使负荷随关节运动进程得到精确调整,即在关节角度的张力最弱点负荷最小,在关节角度张力的最强点负荷最大,因此,在整个关节范围内肌肉产生的张力能始终与负荷等同,肌肉能
以恒定速度或等同的强度收缩。是使肌肉在整个关节运动范围,都能得到最大锻炼。
应用:自由泳的手臂划水。
(二)拉长收缩
①
拉长收缩:
当肌肉收缩所产生的张力小于外力时,肌肉积极收缩但被拉长,这种收缩形式为拉长收缩。
②
力学特征:
A、肌肉收缩产生的张力方向与负荷移动方向相反,肌肉做负功;
B、拉长收缩时肌肉起止点相离,又称离心收缩;
③应用:在人体运动中拉长收缩起着制动、减速、克服重力等作用,跑步时支撑腿后蹬前的屈髋、屈膝等,使臀大肌、股四头肌等被预先拉长,为后蹬时的伸、伸膝发挥更大的肌肉力量创造了条件。
(三)等长收缩
①等长收缩:
肌肉收缩产生的张力等于外力,不做功,肌肉积极收缩,但长度不变,这种收缩形式被称之为等长收缩。
②力学特征:等长收缩时负荷未发生位移,从物理学角度认识,肌肉没有做功,但仍消耗很多能量
③应用:等长收缩是静力性工作的基础,在人体运动中对运动环节固定、支持、保持身体某种姿势起重要作用。
二、肌肉收缩的力学特征
肌肉收缩的力学特征是指肌肉收缩时的张力与速度长度与张力的关系,此外,还有肌肉做功和功率问题等。
(一)肌肉收缩的张力与速度的关系/肌肉收缩的张力-速度曲线图
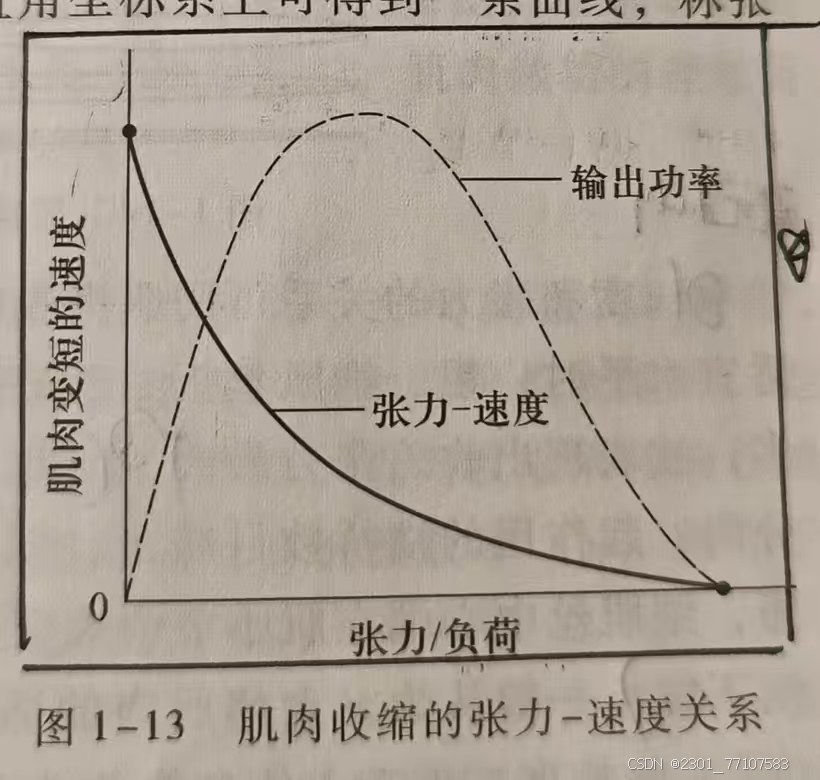
肌肉在后负荷(肌肉开始收缩时才遇到的负荷或阻力)作用下表现的张力与速度的关系描绘在直角坐标系上可得到一条曲线,称
张力-速度曲线。
1、表现(该曲线说明)
(1)在一定的范围内,肌肉收缩产生的张力和速度大致
呈反比;
(2)当后负荷增加到某一数值时,张力可达到最大,但收缩速度为 0,肌肉只能做
等长收缩;
(3)当后负荷为
零时,张力理论上为0,肌肉收缩速度达到最大;
(4)提示:要获得
收缩的较大速度,负荷必须相应减少;要克服较大阻力,即产生较大张力,收缩速度必须慢。
2、机制:
(1)张力和速度关系由肌肉性质决定,
(2)肌肉张力和收缩速度可能分别由两种独立的机制所控制。张力的大小取决于活化的横桥数目,收缩速度取决于横桥上能量释放的速率。
(3)当后负荷增大时,使更多的横桥处于活化状态,从而增大肌肉收缩的张力,同时抑制ATP 水解,降低能量释放率,使收缩速度变慢。
(二)肌肉收缩的长度与张力关系
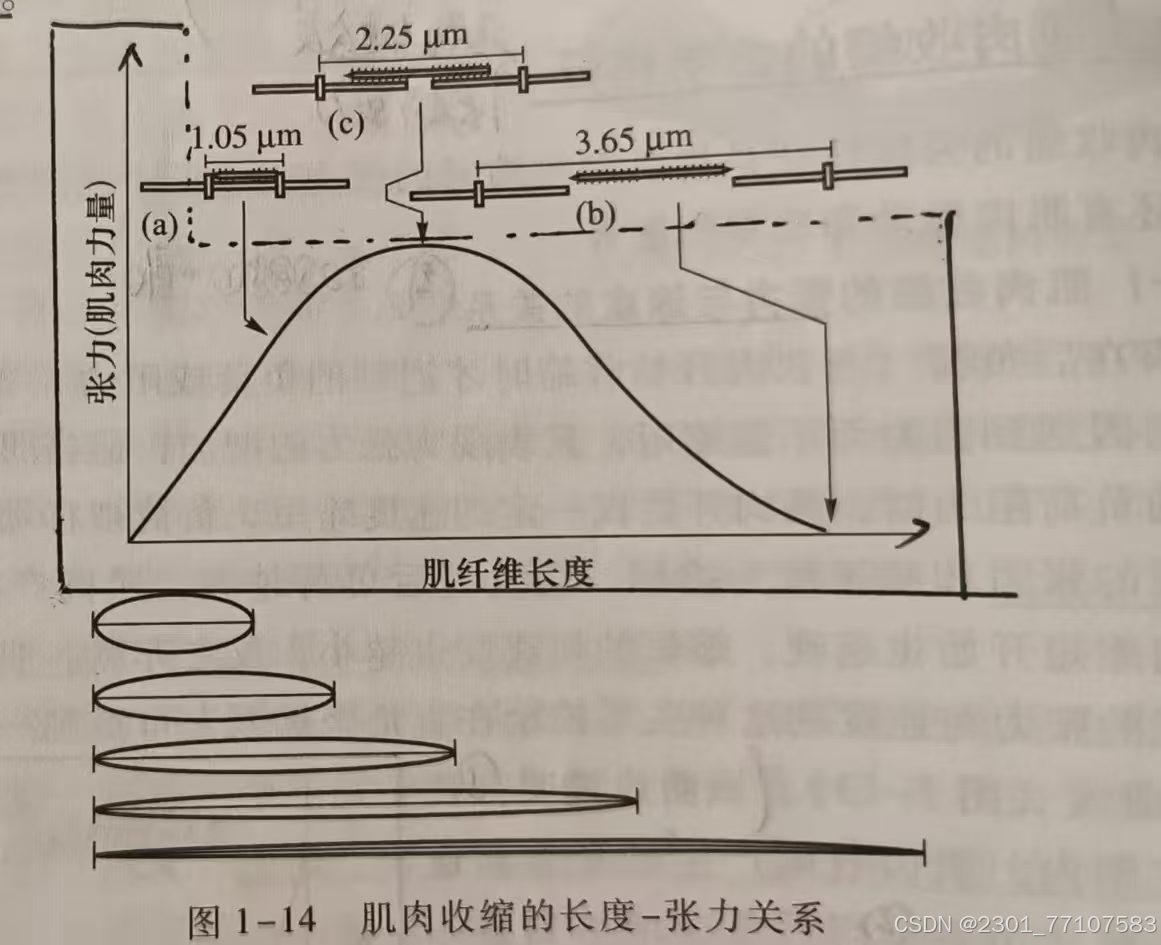
在坐标系上将肌肉在不同前负荷(肌肉收缩前施加于肌肉一定负荷)作用下张力与长度的变化绘制下来,可以得到一条曲线,称为
肌肉收缩的长度-张力曲线。
1、特点:
(1)在肌肉收缩前,加于肌肉一定负荷(称前负荷),使肌肉收缩前就处于被拉长的状态,即改变肌肉初长度。
(2)最初增大肌肉收缩初长度,肌肉收缩时产生的张力也增加。
(3)初长度增大超过某一长度时,张力反而减少(可联想到蹲踞式起跑)。适宜初长度时,肌肉收缩产生的张力最大。
2、机制:可用肌肉收缩的肌丝滑型理论加以解释
(1)肌肉长度处于适宜水平时,粗、细肌丝处于最理想的重叠状态,因而起作用的横桥数目最多,表现收缩张力最大。
(2)肌肉拉得太长,粗、细肌丝趋向分离,起作用的横桥数目减少,肌肉张力下降。
(3)肌肉过于缩短,细肌丝中心端在肌小节中央交错,起作用的横桥数目减少,
肌张力将急剧下降。
第四节 肌纤维类型与运动能力
一、肌纤维类型
依据骨骼肌的形态、结构和功能特征对其类别进行判别的结果,称为肌纤维类型。依据收缩速度的差异可将骨骼肌纤维划分为“慢肌”和“快肌”。
二、不同类型肌纤维的形态、代谢、生理特征
(一)快肌纤维
1、
形态学特征:快肌纤维直径粗、肌纤维数量多;肌浆少,肌红蛋白含量少,呈苍白色,其肌浆中线粒体数量少、体积小;但肌浆网发达,是慢肌纤维肌浆网的两倍左右;对钙离子的摄取速度快,因而反应速度快。接受胞体大的α-运动神经元支配
一个大α运动神经元连同它支配的快肌纤维或Ⅱ型肌纤维,称
快运动单位;
一个小α运动神经元连同它支配的慢肌纤维或Ⅰ型肌纤维,称
慢运动单位
2、
代谢特征:快肌纤维肌球蛋白ATP 酶、磷酸果糖激酶、乳酸脱氢酶等
无氧代谢酶活性高,糖原储备多,
供能方式以无氧代谢为主,因此无氧代谢能力较高。快肌纤维与无氧有关的酶的活性明显高于慢肌纤维。
3、
生理特征:快肌纤维
收缩的潜伏期短,收缩
速度快,收缩时产生的
张力大,与慢肌纤维相比,肌肉收缩的张力-速度曲线位于其上方,但收缩不能持久,易疲劳
(二)慢肌纤维
1、
形态学特征:慢肌纤维直径较细,肌纤维数量少;肌浆丰富,肌红蛋白含量高,呈红色;其肌浆中线粒体直径大、数量多、容积大;周围毛细血管网发达,受胞体小的α-运动神经元支配
2、
代谢特征:慢肌纤维氧化磷酸化酶、琥珀酸脱氢酶等活性高,氧化脂肪能力强,
慢肌纤维氧化脂肪的能力是快肌纤维的四倍。供能方式以有氧代谢为主,因而有氧氧化能力较高。
3、
生理特征:慢肌纤维收缩的潜伏期长,肌肉收缩速度慢,肌肉收缩产生的张力小,但持久、抗疲劳能力强。
三、肌纤维类型与运动的关系
(一)运动单位募集
运动单位募集指的是
运动过程中不同类型运动单位参与活动的次序和程度。
(二)肌纤维百分比与运动能力的关系,在运动实践中的意义(补充)
1、肌肉的
最大收缩速度、爆发力、纵跳高度与快肌纤维的百分组成成正相关,静力性力量和耐力则与慢肌纤维百分组成成正相关
2、
依据肌纤维百分组成与运动能力的关系,可为运动员选取更为适合他们的运动项目。
(1)腿部肌肉中快肌纤维百分比高的人适于从事短跑、举重、跳跃等短时间剧烈运动,力量、速度为主的项目;
(2)腿肌中慢肌纤维百分比高的人适于从事长距离跑、马拉松的以耐力为主的运动项目;
(3) 腿部快、慢肌纤维的百分组成接近的人,则适合进行中长距离跑的运动项目。
3、两类肌纤维百分组成与运动能力的及运动专项的对应关系并非是绝对的。在运动实践中,即使运动员不具备其所从事专项的相应肌纤维类型优势,由于充分发挥其他因素的作用,也可能取得好的成绩。所以
肌纤维类型的构成只是影响运动成绩的因素之一,而不是唯一因素,优异的运动成绩最终是由运动员的体能、技能和心理因素来决定的。
在实践中既要关注科学选材,又要高度重视科学训练。
(三)运动训练对骨骼肌纤维的影响
1、运动训练对骨骼肌纤维类型百分构成的影响
训练可导致
肌纤维类型改变:近年来研究认为,长期大强度耐力训练,可使快肌纤维变成慢肌纤维,而速度和力量训练只能引起肌纤维某些细微结构和代谢功能的改变;运动神经支配是决定肌纤维类型和控制其转变的可能原因。
2、运动训练对肌纤维面积的影响
训练能使
肌纤维出现选择性肥大:不同类型骨骼肌纤维的肥大主要与运动训练的形式有关,即不同形式的运动训练可优先造成主要运动肌内部某类型肌纤维的肥大,如速度和力量训练使快肌纤维增粗,而耐力训练对慢肌纤维面积无明显影响或有减小趋向。
3、运动训练对肌纤维代谢特征的影响
(1)运动训练对
肌纤维有氧能力的影响:耐力训练不仅可以明显使慢肌纤维有氧能力获得提高,而且也能使快肌纤维有氧能力获得改善,而力量训练则使肌纤维有氧氧化能力下降;
(2)运动训练对肌纤维
无氧能力的影响:田径运动中不同项目的优秀运动员的乳酸脱氢酶活性不尽相同,短跑运动员最高,长跑运动员最低,其他项目介于两者之间。人体的无氧能力可随运动专项或所经受的训练形式而改变
4、运动训练对肌纤维影响的专一性:
如以上肢活动为主的划船运动员,臀部慢肌纤维相对面积大于腿部。
第五节 肌电图
一、
肌电图:
采用引导电极将肌肉兴奋时的电变化经过引导、放大和记录所得到的电压变化图形称为肌电图。
二、肌电图的应用
1.肌肉活动的
协调性评价
采用肌电图记录运动时多块肌肉的 SEMG 信号,通过肌电信号分析和判别主要动力肌群、协同肌群和拮抗肌群的动员顺序和活动强度等,了解肌肉活动的协调性,从而为体育教学和运动训练提供科学依据。
2.局部肌肉疲劳度评价
随着肌肉疲劳的发生和发展,表面肌电信号的某些指标会出现相应的特征性变化,在一定程度上可反映出局部肌肉的疲劳程度。例如通过计算MPF、MF的下降率,可以量化评价局部肌肉对抗疲劳的能力。
3.预测肌纤维类型
SEMG信号是一定范围内许多肌纤维活动的总和,利用SEMG 预测骨骼肌纤维类型的基本理论依据是抗阻负荷过程中某些表面肌电信号特征与Ⅰ型纤维比例呈线性负相关也或与Ⅱ型纤维比例呈正相关,大多数学者的研究认为快肌纤维成分高者MPF 较高,疲劳时下降明显,而慢肌纤维成分高者 MPF 下降不明显。


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









