






今天给大家推荐一篇由清华大学自动化系戴琼海教授团队于2022年11月发表在Photonix上的一篇文章,标题为“Multi‑focus light‑field microscopy for high‑speed large‑volume imaging”。在这篇文章中,作者提出一种多焦点同步采集的球差辅助扫描光场成像方法,利用折射率不匹配引入球差相位调制,搭建无需修改硬件的球面像差辅助扫描光场显微镜(SAsLFM),通过对不同孔径分量焦点的空间位置进行再分配,实现同步多焦点体数据采集。进一步,开发相空间分块融合重建算法,可以从所搭建的SAsLFM系统采集的高维光场数据中抽离出不同深度的高分辨信息并进行匹配融合,从而还原样本的高分辨三维体信息。该方法在不需要修改硬件的情况下,可以将样本的轴向信息扩展3倍,从而为快速样本三维可视化检测提供新的方法。

文献内容
三维生物体的形态和功能动力学的快速高维可视化,如神经元钙瞬变、血管运输、胚胎发育和分子信号网络,需要高速体积成像技术来解决大时空尺度上的这些复杂性。光场显微成像技术由于其高并行性与低毒性备受研究者青睐。该技术通过将微透镜阵列(Microlens Array,MLA)插入检测路径,对快照中大体积的高维信息进行编码,可在单次拍摄中检测样本的三维空间内的高维光信息,进而通过解码算法,高分辨还原样本的三维信息。然而,LFM仍然受到时空分辨率和体积覆盖率之间的权衡,阻碍了在大规模体积中观察样本的细微细节。为突破这一限制,文章提出一种多焦点同步采集的球差辅助扫描光场成像技术,在不需要修改硬件的条件下借助折射率不匹配引入的球面像差,实现同时多焦点光场检测,并通过在分辨率板与斑马鱼模型上进行实验,验证了该方法的可行性。
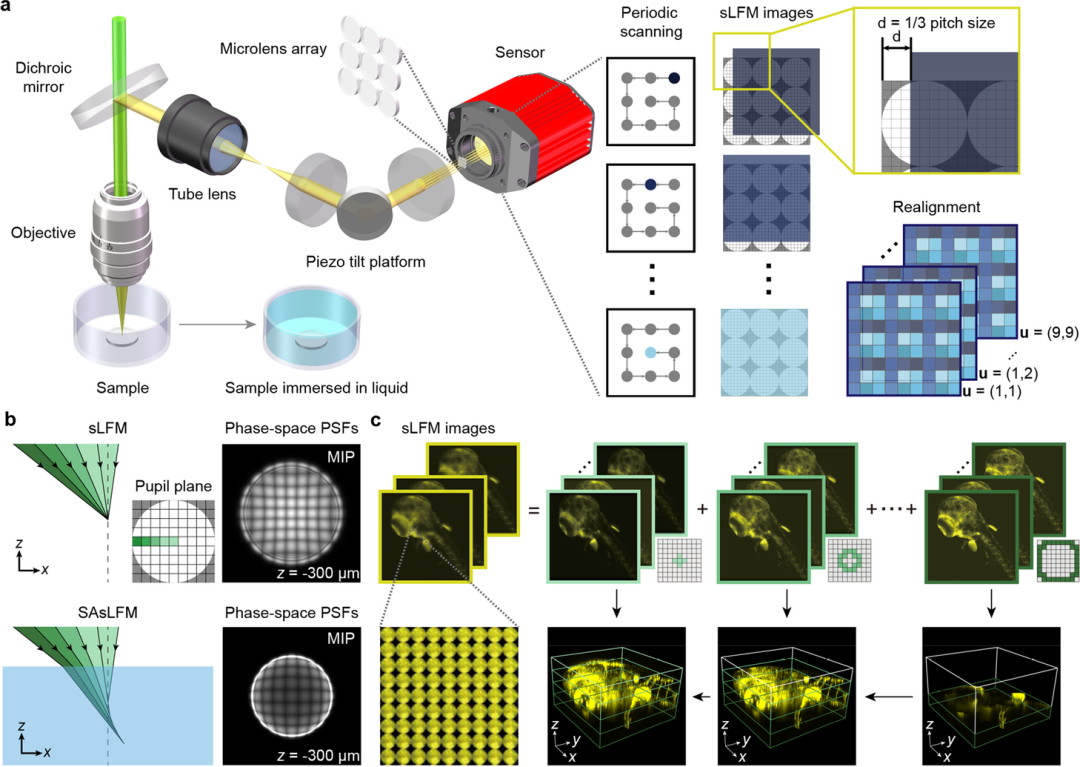
图1:SAsLFM原理;(a) SAsLFM示意图;(b) 子孔径光路编码示意图;
(c) SAsLFM数据的重新排列与重建
其中图1为本文所提方法的原理图,在这里主要选择了使用在原生图像平面插入MLA的未聚焦扫描光场显微镜,相比传统光场显微镜,主要同通过将振镜放置在共轭光瞳孔平面,能够使图像平面以小间隔移动,以增加空间采样率。基于周期性扫描路径,相对于每个微透镜的中心具有相同位置的像素可以重新排列在一起,以形成强度投影I(x,u),其中x和u分别是2D空间坐标和角度坐标。相空间的子孔径分量可以通过
![]()
来计算,其中z是物体空间中的轴向坐标,W(z,x,u)为沿相同方向的子孔径分量的点扩散函数,X(x,z)为三维样本。这些公式表明,子孔径分量之间是相互独立的,所以在理论上可以通过在光瞳孔平面引入相位调制,从而将子孔径分量的焦平面转移到不同的深度。同时考虑到相邻子孔径分量之间的串扰和由光瞳平面处的波前不连续引起的伪影,在本文中选择了引入球面像差平滑相位。其中球面像差可以通过增加样品缓冲液和物镜的浸没介质之间的折射率失配来引入,而引入的其它像差,可以经重建算法中数字自适应方法进行校正,对最终重建性能的影响可以忽略不计。然后将子孔径分量根据其相对于光瞳孔平面的中心位置分为不同的组,同一组中的子孔径分量包含来自相同深度范围内的高分辨信息,通过在相空间重建的迭代过程中合并所有信息可以在扩展的轴向范围内产生具有高分辨率的体积图像。
为定量评估球面像差对成像性能的影响,该研究团队采用数值模拟方法,对20 X/0.5 NA扫描光场显微镜系统(sLFM)的光束传播特性进行了系统分析。通过在光瞳平面上引入不同强度的球面像差,深入研究了像差对成像质量的影响规律。如图2所示,在无球面像差的理想情况下,所有子孔径的点扩散函数均聚焦于同一深度,系统景深仅为50 μm。然而,随着球面像差泽尼克系数的增加,各子孔径分量的聚焦深度呈现显著差异,如当主球面像差的泽尼克系数分别增至5和10时,系统景深可分别扩展至120 μm和185 μm,充分证明了适度球面像差在扩展成像景深方面的积极作用。
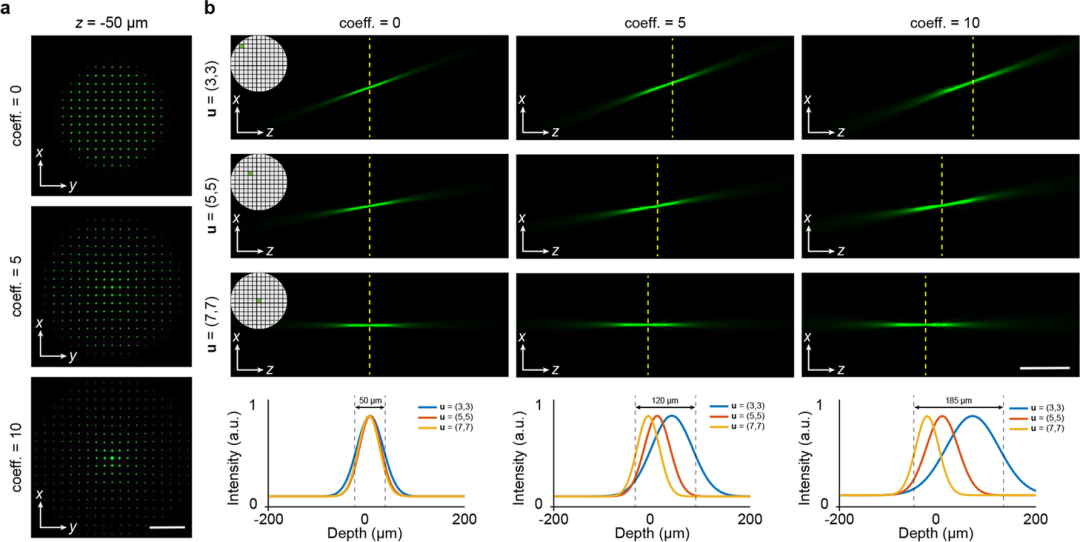
图2:模拟的不同水平球面像差调制的点扩散函数示意图;(a)基于波动光学理论,在光瞳平面附加不同水平的初级球面像差,生成离散的空间PSF;(b) 基于波动光学理论,模拟了具有20 X /0.5 NA物镜的13×13 SAsLFM系统的相空间PSF
为验证该方法在大景深范围内的高分辨率体积成像能力,该文章采用10 X/0.28 NA长工作距离物镜,在不同扫描周期下对轴向不同深度的分辨率板进行了系统性成像实验。如图3所示,虽然SAsLFM由于点扩散函数的横向扩展导致原生焦平面的分辨率出现轻微下降,但其仍能保持显著的横向分辨率。更重要的是,与传统的扫描光场显微镜相比,SAsLFM在较大轴向深度范围内展现出更优异的分辨能力,能够清晰分辨间距更小的条纹结构,充分证明了该方法在三维成像中的优越性能。
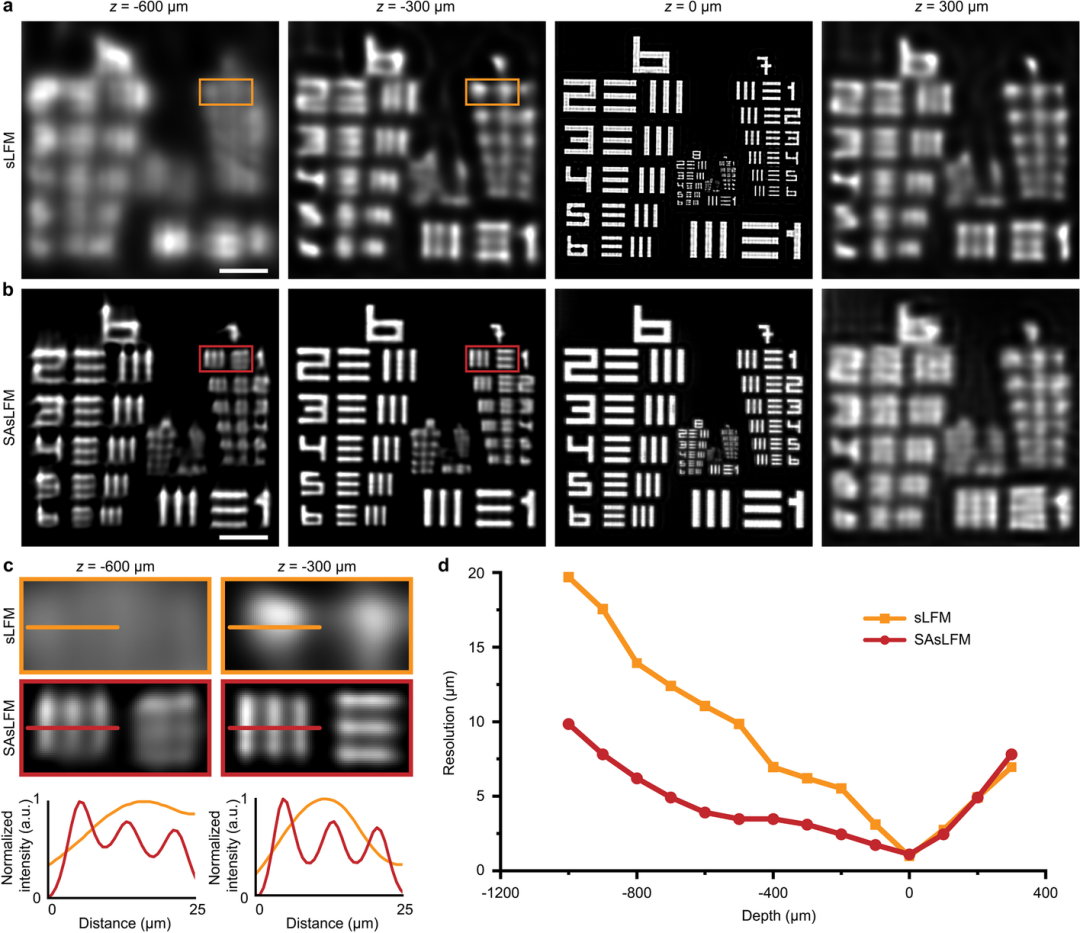
图3:不同深度的分辨率板的重建结果;(a)(b) sLFM和SAsLFM的分辨率板重建结果的比较;(c)图a与图b相同区域的放大细节以及在相同位置上归一化强度线的比较;(d) sLFM和SAsLFM在不同轴向位置的分辨率
为验证SAsLFM在体积成像中的性能优势,该研究团队在相同实验条件下,分别采用sLFM和SAsLFM对斑马鱼全身血管进行了对比成像实验。如图4所示,SAsLFM的子孔径分量能够清晰分辨出原始sLFM图像中模糊的细微血管结构,展现出更优异的细节分辨能力。重建结果的对比分析进一步表明,随着轴向深度范围的增加,sLFM成像的血管结构逐渐模糊,而SAsLFM在整个大轴向深度范围内均能保持高保真的血管结构分辨能力,充分证实了其在三维体积成像中的显著优势。
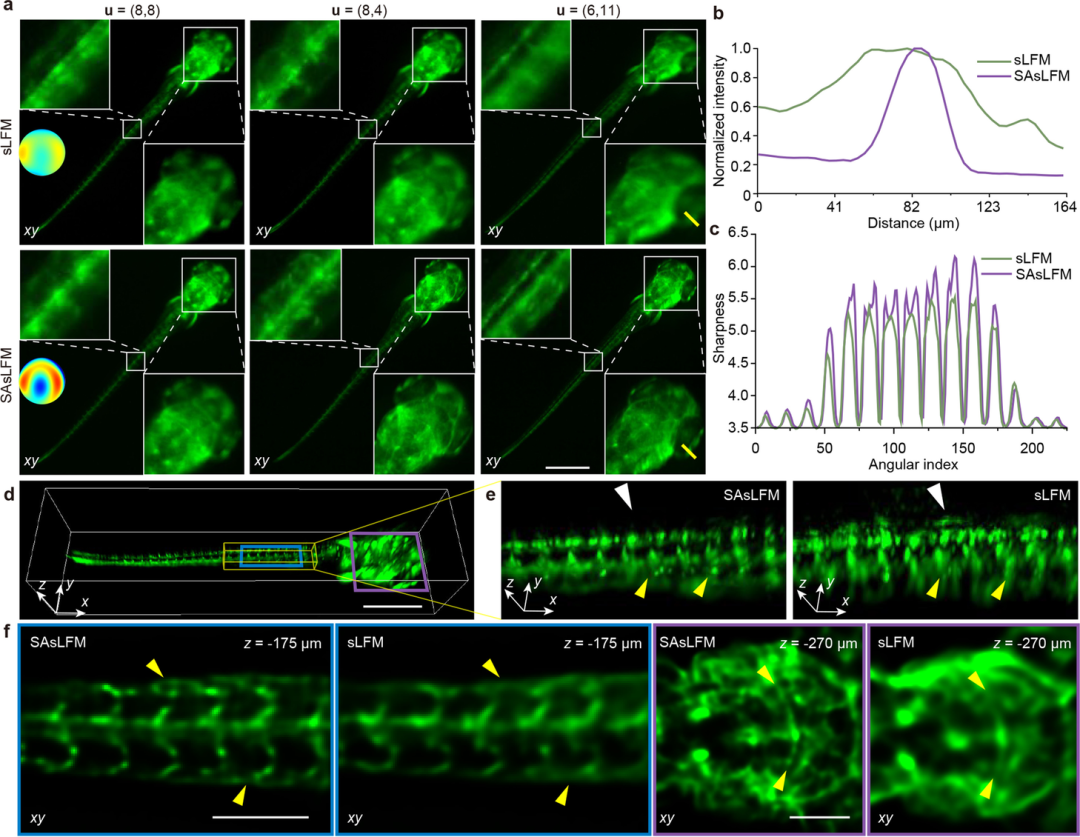
图4:斑马鱼幼虫血管结构的体积成像;(a) sLFM和SAsLFM图像重新排列的子孔径分量比较;(b)图a中黄色虚线的归一化强度比较;(c)清晰度与子孔径指数的强度分布;(d) SAsLFM获得的3D渲染体积;(e)d中黄色区域的放大图;(f) SAsLFM的不同深度的重建切片
为全面验证SAsLFM的多功能成像能力,该研究团队开展了一系列活体生物样本的三维成像实验。首先,通过对清醒小鼠大脑的钙活动进行大规模三维成像,其中主要在光路中引入重量小于2克的水容器以产生球面像差,成功实现了约2000×2000×330 μm³大成像体积内的单神经元分辨率,且无需轴向扫描,结果如图5(a)所示。为进一步评估SAsLFM在活体动物动态追踪中的性能,研究团队在22 Hz的成像频率下对自由移动的水母蝶状幼体进行了实时成像。实验结果表明,SAsLFM不仅能够清晰解析大深度范围内的精细三维结构,还可在约2000×2000×500 μm³的体积内实现对折叠触手的精确追踪,结果如图5(d)所示。此外,为验证SAsLFM的高光子效率和低光毒性特性,研究团队对表达His2Av-mRFP1的果蝇胚胎发育过程进行了长时间观测,结果如图5(g)所示。即使在胚胎肌肉高度活跃的发育后期,SAsLFM仍能实现单细胞分辨率的全胚胎无伪影成像,充分证明了其在大视场长期三维观测中的独特优势,为发育生物学研究提供了强有力的技术支持。
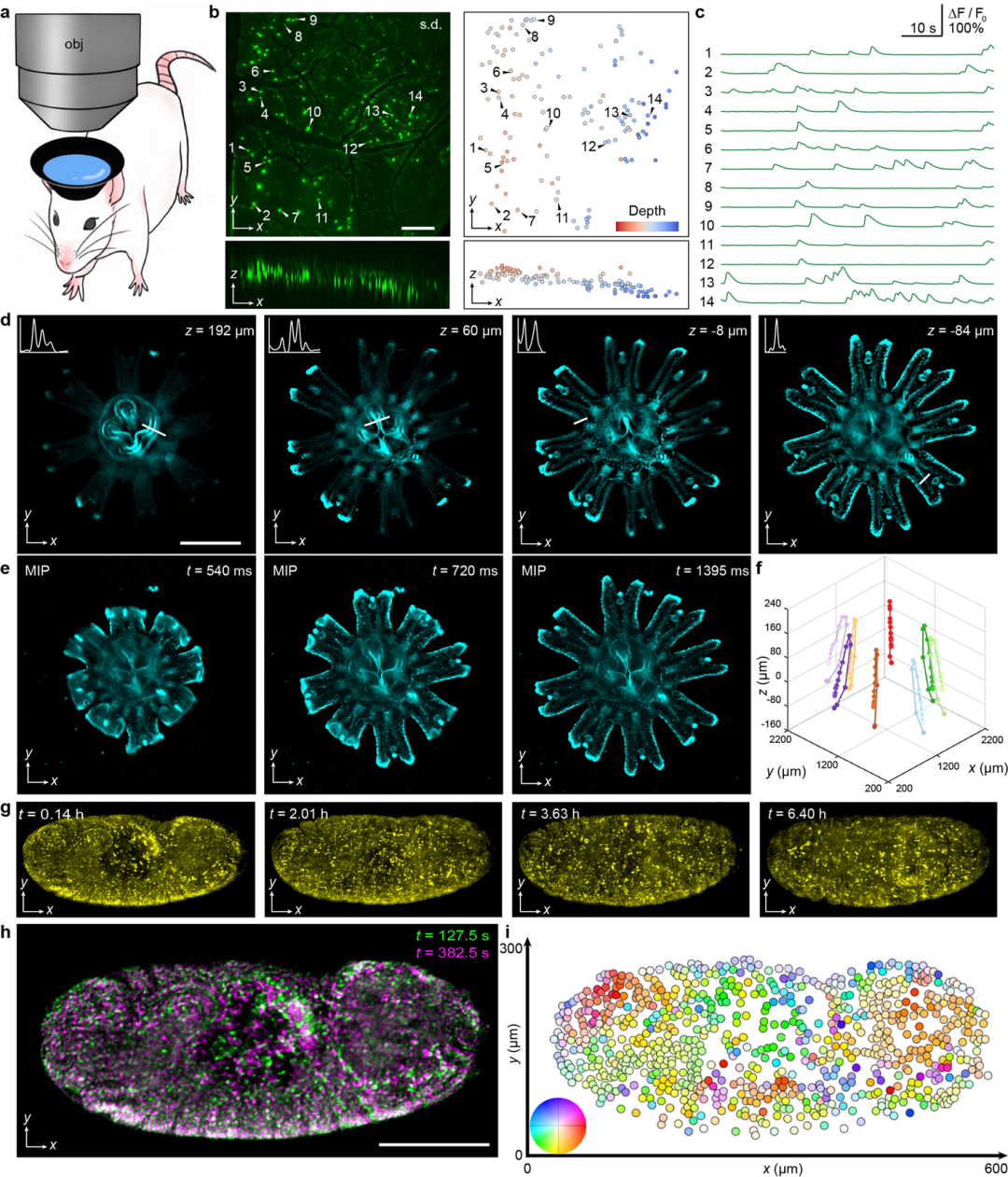
图5:SAsLFM的不同种类生物样本的成像;(a)小鼠实验模型;(b)清醒小鼠大脑的神经元在1009个体积上标准偏差的正交MIP;(c) 在b中标记的14个选定神经元的时间轨迹;(d)在不同深度重建的水母蝶状幼体的切片显示;(e)(f)在不同时间段的水母蝶状幼体重建结果的最大强度投影以及3D跟踪;(g)果蝇胚胎发育后期细胞动力学的时间序列;(h)(i)以255秒的时间间隔分离的两个体积的MIPs以及整个果蝇胚胎的半自动跟踪
为实现SAsLFM的实时三维重建,该研究团队创新性地提出了一种基于U-net架构的相空间反卷积算法,该算法可实现每秒167个体积的超快重建速度。如图6所示,该方法首先采集高分辨率三维体积数据,并通过正向投影获取相空间SAsLFM图像;随后,采用数据增强和噪声添加等技术对数据进行预处理,并与高分辨率体积数据配对,用于网络训练。实验结果表明,尽管该网络的重建质量与基于ADMM的相空间去卷积算法相当,但其计算效率提升了三个数量级,为活体样本的实时三维动态观测提供了强有力的技术支持。
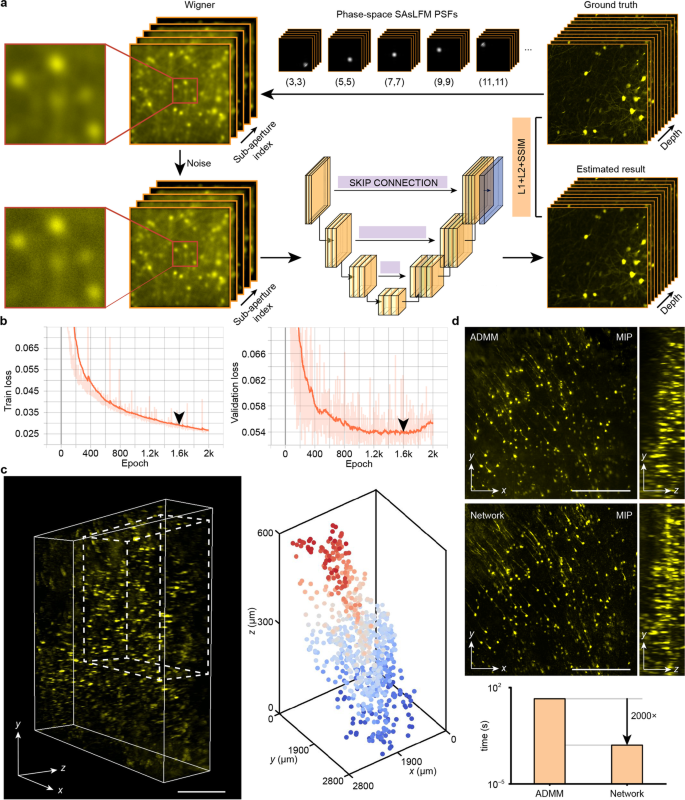
图6:深度学习重建方法示意图;(a)网络数据生成与训练流程;(b)网络训练损失与验证集损失;(c)网络训练后的输出数据的合成体积,由25个重叠50%的重建图像堆栈组成;(d)基于ADMM的相空间反卷积方法与网络重建方法的比较
本研究创新性地提出了一种基于子孔径点扩散函数调控的多焦点成像技术。该技术通过调节物镜与样品间浸没介质的折射率,在光路中引入可控的大球面像差,从而实现对子孔径点扩散函数沿z轴能量分布的精确调制,显著扩展了系统的轴向高分辨率成像范围。实验结果表明,该方法不仅有效缓解了传统成像中远离焦平面区域分辨率显著下降的问题,同时显著减少了焦平面附近的成像伪影,为厚样品中精细三维结构的快速测量提供了强有力的技术支持。展望未来,该研究团队将进一步探索更复杂的相位调制模式以优化系统性能,并致力于拓展SAsLFM在生物医学等领域的应用范围。
文章转自:生物医学光子学与分子影像实验室
声明:转载此文目的在于传递更多信息,仅供读者学习、交流之目的。文章版权归原作者所有,如有侵权,请联系删除。

















 546
546

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









