





 本文研究了不同冥想传统对大脑动态的影响,特别是内观冥想导致的α和γ波段振荡的熵增加。通过脑电图(EEG)分析,发现内观冥想实践增加了α和γ波段的样本熵,表明大脑状态的复杂性和不确定性增加。此外,研究还观察到低/高γ波段的相干性和亚稳态的变化,这与意识丰富度和大脑功能的整合有关。机器学习分类器能够根据这些熵变化有效地区分冥想状态。这些发现扩展了对冥想神经机制的理解,为意识研究提供了新的视角。
本文研究了不同冥想传统对大脑动态的影响,特别是内观冥想导致的α和γ波段振荡的熵增加。通过脑电图(EEG)分析,发现内观冥想实践增加了α和γ波段的样本熵,表明大脑状态的复杂性和不确定性增加。此外,研究还观察到低/高γ波段的相干性和亚稳态的变化,这与意识丰富度和大脑功能的整合有关。机器学习分类器能够根据这些熵变化有效地区分冥想状态。这些发现扩展了对冥想神经机制的理解,为意识研究提供了新的视角。
本文探讨了一个假设,即神经动态的熵指数可以反映意识内容的强度和质量。以往的研究已经证实,5-羟色胺类迷幻药物可能对大脑活动产生紊乱作用,导致主观效果与某些冥想状态的现象学有很大的重叠。在这里,我们认为长期的冥想实践会导致内源性大脑振荡活动熵的增加。我们估计了被归类为“专注注意力”(喜马拉雅瑜伽),“开放性监控”(内观)和“开放性觉察”(伊莎舒尼雅瑜伽)传统冥想状态中波段特定振荡的熵。在所有传统中,内观导致了最高的熵增加,主要在α波和低/高γ波段。与之前的研究一致,所有冥想传统都增加了γ波段的全局一致性,但也通过降低超稳定性来稳定γ范围的动态。最后,机器学习分类器可以根据γ波段熵的头皮分布在某些冥想传统对之间进行有效概括。我们的研究结果扩展了之前关于冥想过程中观察到的频谱变化的发现,展示了长期练习如何实现高熵的大脑状态。这构成了一个内源性、自我诱导的高熵状态的例子。本文发表在NEUROSCIENCE杂志。
关键词:冥想,脑电图,熵,α波振荡,γ波振荡,内观。
引言
人类意识的范围丰富多样,但这种多样性的神经基础仍然难以捉摸(Koch 等,2016)。最近,Carhart-Harris 及其同事提出了一个假设,即自发脑活动的熵程度指标了意识主观体验的“丰富性”(Carhart-Harris 等,2014)。熵是一种来自统计力学理论的无量纲量,可以解释为物理系统中存在的无序程度(Ben-Naim,2012)。正如这一假设的支持者所说:“将熵度量应用于认知神经科学的极大优点在于,它能够独特地弥合物理和主观之间的鸿沟;只是同一枚硬币的两面——但仍然是不同的面。”根据这一提议,低熵脑状态表现为潜在配置的减少,因此在主观上被体验为意识觉醒度降低的状态(Schartner 等,2015;2017b;Hudetz 等,2016;Liu 等,2018)。相反,高熵脑状态被体验为内容丰富,但同时是不可预测的,因此是不确定的(Tagliazucchi 等,2014;Cavanna 等,2017;Carhart-Harris,2018),从而使主观体验超出日常意识的范围。例如,已经将5-羟色胺类致幻剂(即5-羟色胺5-HT2A受体激动剂)、谷氨酸类解离剂(即NMDA受体拮抗剂)(Schartner 等,2017a;Viol 等,2017;2019)以及非选择性腺苷拮抗剂兴奋剂(咖啡因)(Chang 等,2018)的急性效应与高熵脑振荡相关,或者等效地,与可压缩性降低相关。
从信息论的角度来看,熵表示确定系统微观状态所需的信息量,或者等同于其不确定性和不可预测性的水平。1 大脑熵的生物学意义取决于所研究的实验变量。使用脑电图(EEG)测量的头皮电位的大规模振荡经常被用作生理、病理和药理诱导全球大脑状态(如睡眠、癫痫、昏迷、麻醉)的标记。因此,这种全球大脑振荡的熵可以近似于潜在大脑配置分布的不确定性,从而产生主观感知的意识体验“丰富度”的客观标志。 我们提出,熟练冥想者达到的大脑状态与大脑振荡活动的熵增加有关。这一假设得到了观察的支持,即某些冥想传统的长期实践与普通清醒意识相比,主观体验的可选择范围更大(Brandmeyer等人,2019)。特别是,其中许多特征与5-羟色胺类迷幻药物的急性效应相同。这些特征包括情绪和情感向正面转变,认知灵活性增加,情绪失调或情绪不稳定,自我意识改变,呈现“梦幻般”状态,以及主体/客体意向结构的崩溃(非二元意识)(Carhart-Harris 和 Nutt 2014;Travis,2014;Nichols,2016;Nour 和 Carhart-Harris,2017;Millie`re等人,2018)。冥想和使用5-羟色胺类迷幻药物经常与获得关于“客观现实”和自我的“洞察”意图相关(Barrett 和 Griffiths,2018)。此外,证据表明,某些冥想实践(Strauss等人,2014;Haukaas等人,2018)和5-羟色胺类迷幻药物(Griffiths等人,2016;Carhart-Harris等人,2017,2018;Dos Santos等人,2018;Garcia- Romeu和Richards,2018;Palhano-Fontes等人,2018)可以在患有抑郁和焦虑的患者中产生持久的积极效果,提高了大脑熵增加作为这些效果共同机制的可能性(Carhart-Harris,2018)。
现在人们普遍认识到,“冥想”这个词本身是非特异性的,并且经常被用来指代在文化背景和冥想指导方面都存在异质性的实践(Braboszcz等人,2017)。为了评估我们的假设,同时考虑这种异质性,我们研究了三组遵循不同传统的冥想专家的脑电图数据:喜马拉雅瑜伽(HT),内观(VIP)和伊莎瑜伽(SNY)。这些传统可以沿着一个从“专注关注”(FA)到“开放监控”(OM)冥想的连续谱排列(Lutz等人,2008;Lippelt等人,2014)。喜马拉雅瑜伽使用曼特拉来保持注意力集中,可以被归类为FA(“专注关注”)。伊莎瑜伽的冥想者练习“Shoonya”冥想,这是一个有意识的“不做”的过程,同时对内部或外部刺激保持不响应,可以归类为OM(“开放监控”)。在这两个极端之间,内观要求从业者在心理扫描身体感觉的过程中保持专注(“samatha”);然而,这种冥想的一个关键组成部分是对分心元素的洞察(或元认知,“vipasyana”),这些元素必须被识别并释放,以便继续FA(“专注关注”)过程(Lutz等人,2007)。 我们对每个头皮电极的频段特定振荡活动的局部熵进行了补充分析,并分析了两个全局指标:振荡的长程相干性以及其随时间的波动(即转换稳定性)。这两个指标受到意识理论描述的启发,并已经在实证研究中证明与意识水平有关(Carhart-Harris等人,2014;Schartner等人,2015, 2017a, b)。
实验方法
我们研究了一个公开可用的脑电图数据集。关于实验方法的更多信息可以在之前的出版物中找到(Braboszcz等人,2017)。本文所使用的所有代码可以在https://github.com/enzotagliazucchi/meditation-entropy 找到。
参与者
根据年龄、性别和练习年限,招募了30名对照组(CTR)成员、27名喜马拉雅瑜伽(HT)冥想者、20名内观(VIP)冥想者和27名伊莎瑜伽(SNY)冥想者。对照组受试者的选择依据年龄、性别和没有冥想经历。实验得到了当地伦理委员会和加州大学圣地亚哥分校伦理委员会的批准。实验在印度里士满的冥想研究所(MRI)进行。各组按年龄和性别进行匹配,结果为16名对照组(45±10岁,5名女性)、16名喜马拉雅瑜伽冥想者(43±12岁,2名女性)、16名伊莎瑜伽冥想者(40±10岁,2名女性)和16名内观冥想者(47±15岁,5名女性)。一个单独的受试者组作为所有三种冥想传统的对照组。
实验设计
所有参与者都被要求睁开眼睛,并在坐在地板上的毯子上或椅子上之间进行选择。实验分为两个各持续20分钟的阶段,一个标为“冥想”,另一个标为“指导性漫游思维”。阶段顺序在参与者之间进行了平衡处理,以防止顺序效应。在“冥想”条件下,参与者准备进行冥想练习(专注于呼吸或呼气/吸气)10分钟,然后被要求在最后10分钟内进行冥想。对照组在整个20分钟的条件下执行与冥想者相同的指示。在“指导性漫游思维”条件下,参与者被要求回忆从童年到近期的自传式事件。本次分析基于比较冥想者和对照组在“冥想”条件的后半部分。作为额外检查,我们还比较了对照组的“指导性漫游思维”条件与冥想者的“冥想”条件,以及两组之间的基线条件(准备冥想)。
EEG 数据采集
使用具有10-20 Headcap标准64通道帽子的64+8通道Biosemi Active-Two放大器系统记录EEG数据。记录右侧和左侧乳突电极以及垂直和水平电眼图(EOG)。实验室隔音,地板电气屏蔽和接地。所有电极的偏移量都保持在Biosemi系统测量阻抗的50毫伏以内。
EEG 数据处理
使用EEGLAB(版本12)数据首先参考右侧乳突并从1024Hz降采样至256Hz。在EEG时间序列中应用截止频率为1Hz(无限响应滤波器)、过渡带宽0.3Hz和六阶的高通滤波器。还应用了EEGLAB自动伪迹拒绝程序。遵循之前的出版物(Braboszcz等人,2017)详细描述的手动程序,拒绝具有非常高/低频率内容的8秒时期。通过视觉识别并从分析中移除坏电极(每个受试者0-18个坏电极,每个受试者平均5个电极)。应用Infomax独立成分分析算法来拒绝与眼动和肌肉活动相关的伪迹。
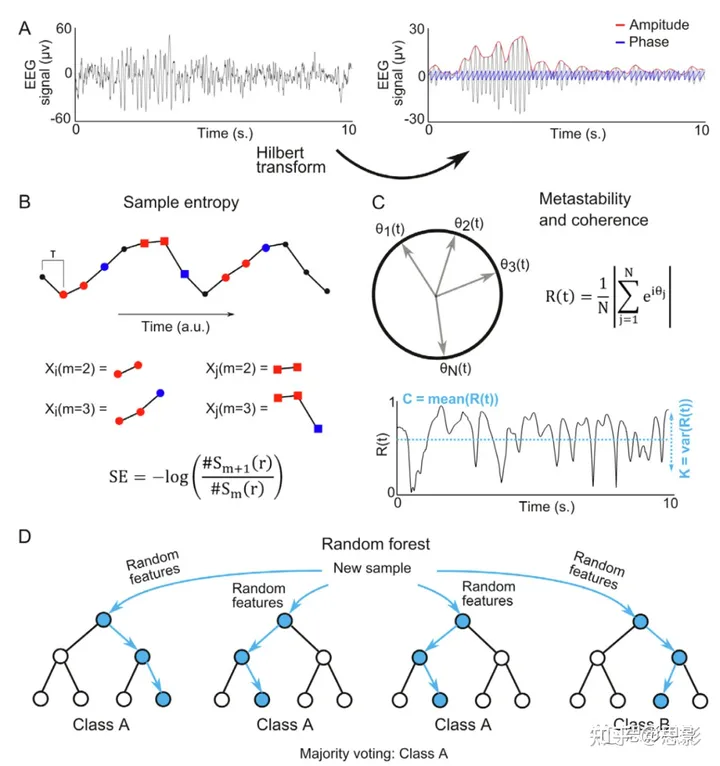
图1. 方法概述。
(A) 带通滤波和希尔伯特变换的应用,以获得EEG信号的瞬时幅度和相位。(B) 嵌入维度m = 2的SE算法示例。展示了长度为m的两个模板向量,X2(i=2)和X2(i=6), 以及相关联的嵌入维度m + 1的模板向量。通过计算模板向量对(对于m和m + 1)的距离在容差r范围内的数量,即Sm(r)和Sm+1(r), 然后应用等式(1)得到样本熵。 (C) 定义一组具有相的N振子的瞬时同步R(t)。显示了R(t)的代表性时间过程,作为相干度C和易失性K的定义的例子,分别为R(t) 的均值和方差。 (D) RF(随机森林)算法图示。对于每个训练样本,选择一个随机特征子集,然后使用这些特征训练一个决策树。通过评估使用选定特征子集训练的决策树来预测新样本的类别,然后应用投票或多数规则(在本例中,投票预测类别A)。
带通滤波和希尔伯特变换
我们将分析重点放在以下典型频段上:delta(1-4 Hz)、theta(4-8 Hz)、alpha(8-12 Hz)、beta(12-20 Hz)、低 gamma(20-60 Hz)和高 gamma(60-100 Hz)。我们选择计算特定频段振荡包络的熵是基于以下两个考虑。首先,窄频段已经与特定大脑功能、认知领域以及特别是冥想状态联系在一起(Buzsaki,2006;Braboszcz等人,2017)。其次,尚不清楚宽带信号波动的频谱内容如何偏向熵估计,或者等效地说,宽带信号的熵在多大程度上提供了与频谱分析互补的信息。
对于每个EEG通道,我们应用了四阶Butterworth带通滤波器以获得上述范围内的窄带信号。然后,我们使用希尔伯特变换得到了过滤时间序列的解析表示。如图1A所示,给定一个实值信号x(t),Hilbert变换产生一个复数表示(解析信号),表示为
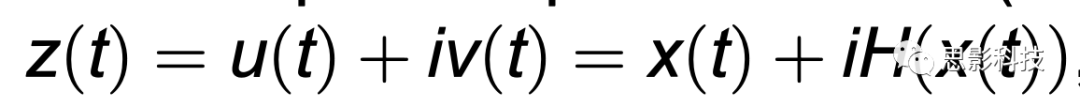
,
其中H(x(t))
表示实值信号x(t)的Hilbert变换。从这个表示中,可以计算出瞬时幅度
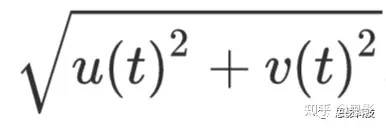
,以及瞬时相位tan(v(t)/u(t))的负一次方
。
样本熵
样本熵(SE)是基于计算数据中的模式重复次数的时间序列复杂度(或等效地,规律性或可预测性)的估计(Richman和Moorman,2000)。SE测量在时间序列中观察到相似(在一定容差范围[r]内)固定长度的模式的概率,与基于将长度增加一个样本的模式进行相同计算所得到的结果相比。模式的长度由嵌入维数(m)确定。与最近的研究相反,我们选择了SE( 样本熵)而不是Lempel-Ziv(LZ)复杂度,因为LZ估计算法意义上的信息(即可压缩性)并且需要对数据进行任意二值化(Schartner等人,2015),而SE可以应用于连续信号,并产生与统计力学(吉布斯)和信息理论(香农)熵定义更直接相关的熵估计。
详细计算SE的步骤在补充材料1。关键计算是
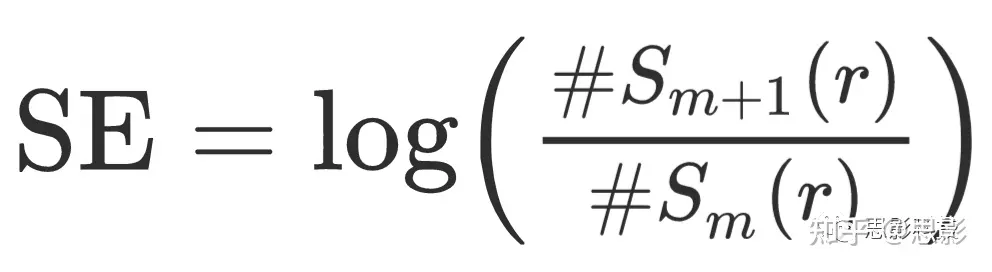
,其中#Sm和#Sm+1分别表示嵌入维数m和m+1的模板向量数量,容差为r。根据以往的出版物(如Al-Angari和Sahakian,2007,Bruce等人,2009,Song等人,2012,Zhang和Zhou,2012,Yentes等人,2013),我们将嵌入维数的值设定为m=2。通常也会将容差表示为与时间序列的标准差(SD)成正比;在这里我们设定为r=0.2xSD。最后,为了减少数据中的自相关影响,我们考虑了。
亚稳态和相干性
考虑N个耦合振荡器(在这种情况下,每个64个带通滤波的EEG通道都可以被概念化为一个振荡器),每个振荡器都有自己的特征振荡频率xi。振荡器集合之间的瞬时同步量可以用Kuramoto序参数来衡量。序参数R(t)的范围在0到1之间,分别表示最小和最大同步(Shanahan,2010)。易变性(K)计算为R(t)的时间方差,表示振荡器的动态变化范围(即随时间发生的瞬态同步和去同步)。相干性C计算为R(t)的平均值,表示所有振荡器之间的平均同步水平。计算K和C的过程在图1C中概述,并在附录中进一步详细解释。
随机森林(RF)分类器
为了评估SE中的变化是否代表每种冥想传统的特定特征,我们使用这些数据训练并评估一类称为RF(Breiman,2001)的机器学习分类器。RF建立在简单的决策树分类器基础上,对于每个特征,都会引入阈值,以便在树的每个节点处将训练集分成两部分。阈值选择以最大化划分后的样本纯度度量;当一个类的所有样本都被分配到划分的一边时,将获得最高可能的纯度值,反之亦然。如果情况并非如此,接下来将评估下一个特征,并递归地确定阈值,直到获得最高纯度值或所有特征都用尽。
对于每个频率带,我们考虑了与每个EEG电极位置的SE熵值相对应的64个特征,并训练了200个独立的RF分类器来区分CTR(对照组)、HT(喜马拉雅瑜伽)、VIP(内观)和SNY(伊莎瑜伽)(Abraham等人,2014)。作为分类器准确性的衡量指标,我们获得了ROC曲线下的面积(AUC)。对于每个分类器,我们采用分层分割方法将其分为两个相等大小的集合(训练和测试)。我们还在随机洗牌样本标签后应用相同的过程,以评估AUC的统计显著性。通过计算洗牌后数据得到的AUC与未洗牌数据得到的AUC相比,构建了经验p值。
最后,我们通过使用一个二元决策问题(例如CTR与VIP)的数据训练RF分类器,然后将其应用于另一个二元决策问题(例如CTR与HT),来研究泛化准确性所获得的AUC。
统计分析:
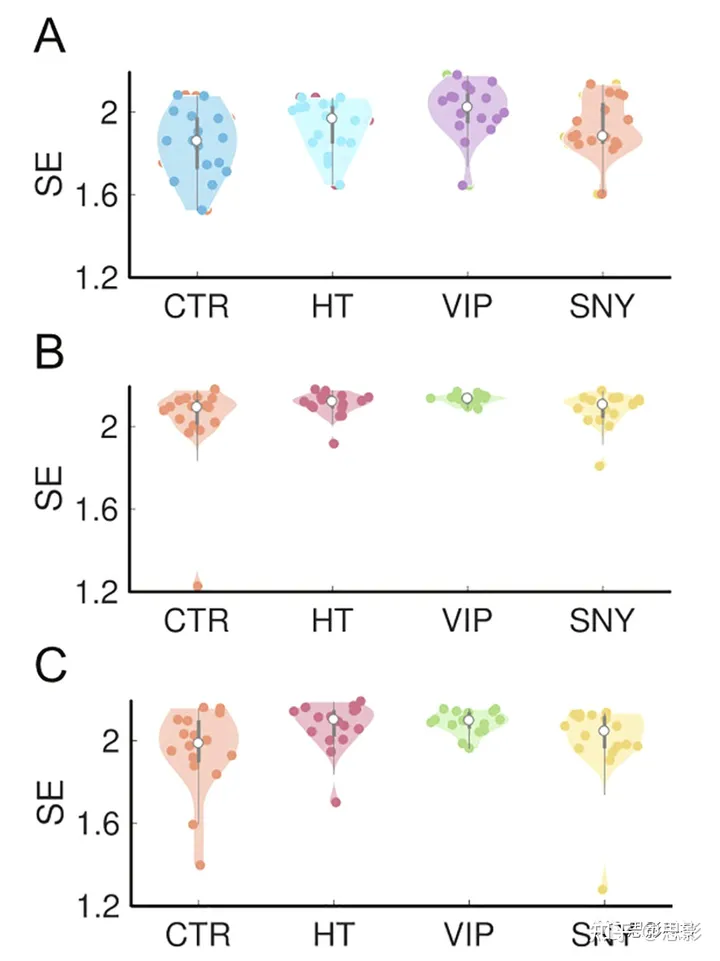
图2. 冥想对大脑振荡熵的影响
冥想状态改变了α波(面板A)、低γ波(面板B)和高γ波(面板C)频带的大脑振荡熵。小提琴图显示了个体参与者之间SE值(对所有电极进行平均)的分布。统计数据在结果部分中报告。
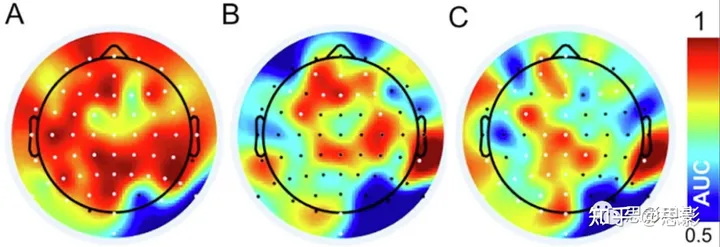
图3. 熵变化的定位
相对于CTR,在α波(面板A)、低γ波(面板B)和高γ波(面板C)频带中,VIP中较高SE值的拓扑分布。白色电极表示显著较高的SE(p ≤ 0.05,Wilcoxon检验,FDR校正)。颜色刻度表示接收操作特征曲线下面积(AUC)的效应大小。
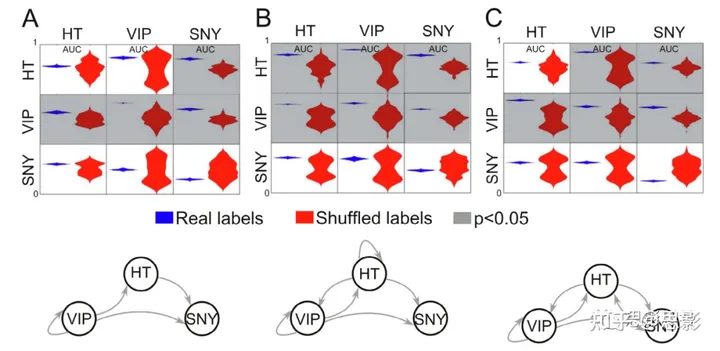
图4. 冥想传统之间熵变化的泛化
基于训练和测试RF分类器的多变量分析,使用α波(面板A)、低γ波(面板B)和高γ波(面板C)频带中64个EEG通道的SE值。
上面板:RF分类器被训练来区分冥想传统(行)与CTR组。之后,它们在区分列中的冥想传统与CTR的问题中进行评估。对角线上的条目对应于使用相同数据集的训练/测试分割来训练和评估分类器。条目显示从未混洗(蓝色)混洗(红色)数据获得的AUC值的分布(小提琴图)。不透明条目表示实证p值(从比较两个AUC分布中获得)为≤0.05(FDR校正)。
底部面板:每个节点代表一个冥想传统,箭头表示训练用于区分源节点传统与CTR的RF分类器成功地将目标节点传统与CTR区分开来。例如,从VIP到其他节点的箭头表示训练用于区分VIP和CTR的RF分类器成功地泛化到另外两个冥想传统。
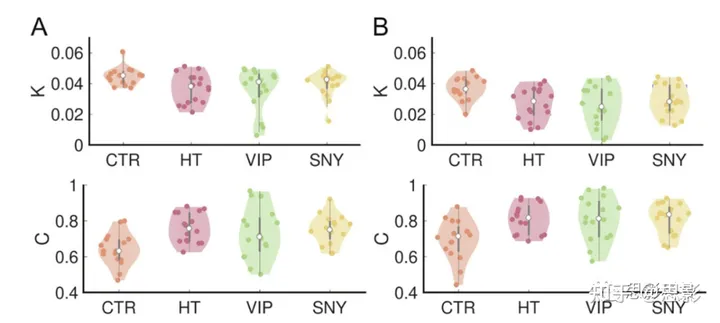
图5. 冥想对Kuramoto序参数动态的影响
冥想状态改变低γ波(面板A)和高γ波(面板B)频带振荡的相干性(C)和非稳定性(K)。小提琴图显示了不同参与者之间K和C分布。统计数据在结果部分报告。
我们采用了Kruskal-Wallis(KW)检验,这是方差分析(ANOVA)的非参数版本,用于评估实验组(CTR、HT、VIP和SNY)对SE(跨所有通道平均)、C和K的影响。每当检测到组效应显著时,我们在CTR数据和每个冥想传统数据之间进行非参数两样本Wilcoxon符号秩检验作为事后分析。Wilcoxon检验的效应量计算为

, 其中n1 = n2 = 16(正在比较的样本的大小)(Rosenthal等人,1994)。Kruskal-Wallis检验的效应量用η2表示(Lakens,2013)。多重比较(例如,图3中的头皮分布或图4中的RF分类器普遍性)使用Benjamini-Hochberg方法控制假发现率(FDR),允许假阳性率为0.05。仅在图2和图5中显示通过多重比较校正(FDR校正的Kruskal-Wallis检验)的频带,低γ波段的亚稳定性(K)除外,其呈现出显著性趋势。
结果
窄带信号包络的SE
首先,我们计算了所有受试者、电极和频率带的窄带EEG信号包络的SE(样本熵)。对于第一个分析,我们使用了“冥想”条件的第二个10分钟,这对应于对照组的关注呼气/吸气的指示,以及对冥想者的实践。我们在α(8-12Hz)、低γ(20-60Hz)(H[3]=9.56,p≤0.05,η2=0.44,FDR校正)和高γ(60-100Hz)(H[3]=8.5,p≤0.05,η2=0.41,FDR校正)频带的平均SE上发现了组效应(H[3]=8.86,p≤0.05,η2=0.42,FDR校正)。事后非参数Wilcoxon检验确定,p≤0.05,FDR校正,只有Vipassana(内观) > 控制组(VIP > CTR)的比较是显著的(Z = 2.65,r = 0.46,p≤0.005),除了高γ波段,我们观察到喜马拉雅瑜伽 > 控制组(HT > CTR)(Z = 2.12,r = 0.38,p≤0.05)和VIP > CTR(Z = 2.28,r = 0.40,p≤0.05)是显著的。平均SE值显示在图2中。
接下来,我们比较了对照组的“指导性心灵漫游”条件与冥想者的“冥想”条件。我们观察到α(8-12Hz)、低γ(20-60Hz)(H[3]=10.5,p≤0.05,η2=0.47,FDR校正)和高γ(60-100Hz)(H[3]=9.94,p≤0.05,η2=0.45,FDR校正)频带的平均SE上有显著的组效应(H[3]=11.71,p≤0.05,η2=0.49,FDR校正)。事后非参数Wilcoxon检验确定,p≤0.05 FDR校正,VIP > CTR的比较在α(Z = 2.92,r = 0.51,p≤0.005)、低γ(Z = 2.77,r = 0.48,p≤0.01)和高γ(Z = 2.65,r = 0.46,p≤0.01)频带上是显著的。
在比较基线条件(即冥想准备)获得的SE值时,我们没有观察到任何频率带上组效应对SE值的显著影响。
VIP(内观)与CTR之间SE增加的电极的拓扑分布显示在图3中。显著的电极(Wilcoxon检验,FDR校正,p≤0.05)用白色表示。颜色刻度表示接收操作特性曲线(AUC)下面积的效应大小。我们发现α波段的SE普遍增加,低γ波段前额SE增加,以及高γ波段中央和前额SE增加。
应用于SE拓扑的多变量分类器 接下来,我们研究了基于RF分类器的多变量分析是否能够区分所有冥想者群体的冥想状态和对照条件。RF分类器基于我们观察到组效应显著的频率带(α,低和高γ)的所有64个EEG通道的组合SE值。我们还评估了一个训练用于区分某个冥想状态(例如HT)与CTR的分类器是否能成功推广到区分另一个组(例如VIP)的冥想状态与CTR的问题。这种分析对于识别不同冥想传统共有的潜在“核心”特征是相关的。
该分析的结果显示在图4中。在上面板中,行对应于用于训练RF分类器的组,列对应于用于评估其性能的组。对角线条目对应于使用相同组训练和评估RF分类器(即一半数据用于训练,另一半用于评估)。在每个条目内,蓝色/红色小提琴图表示使用未洗牌/洗牌类标签获得的结果。阴影条目表示相应的AUC大于偶然水平(p < 0.05,FDR校正)。图4的下面板显示了所有产生显著大于偶然水平的AUC的训练/评估对。
在所有情况下,都可以高度准确地区分VIP(内观)和CTR(对照)组。其他冥想传统则不然,除了低γ波段的HT;然而,我们注意到,训练用于区分VIP和CTR的RF分类器成功地推广到了其他两种冥想传统的所有三个频率带。这些结果与图2、图3中呈现的结果一致,并暗示,尽管SE的增加是VIP的特征,但SE的拓扑分布可以提供关于其他冥想传统共有的特征的信息。
相干性和亚稳态
最后,我们研究了所有EEG电极间的相干性C,计算为同步性的时间平均值。我们还从方差中获得了亚稳态K。图5的上/下面板分别显示了所有实验组的K/C。我们评估了低(H[3] = 5.97,p≤0.1,η2 = 0.33)和高γ波段(H[3] = 9.95,p≤0.05,η2 = 0.45)的组效应对K的影响,以及低(H[3] = 12.03,p≤0.01,η2 = 0.50)和高γ波段(H[3] = 11.2,p≤0.01,η2 = 0.48)的组效应对C的影响。
事后非参数Wilcoxon检验发现,在低γ波段,冥想期间K值减少,用于比较HT < CTR(Z = 2.24,p≤0.01,r = 0.39)和Isha瑜伽 > 控制(SNY < CTR)(Z = 1.16,p≤0.05,r = 0.29);在高γ波段,比较HT < CTR(Z = 2.27,p≤0.005,r = 0.39)、VIP < CTR(Z = 2.35,p≤0.01,r = 0.41)和SNY < CTR(Z = 2.31,p≤0.01,r = 0.40)。相反,在低γ波段,C值增加了HT > CTR(Z = 3.10,p≤0.001,r = 0.54)、VIP > CTR(Z = 1.90,p≤0.05,r = 0.33)、SNT > CTR(Z = 3.03,p≤0.001,r = 0.53);在高γ波段,C值增加了HT > CTR(Z = 2.80,p≤0.005,r = 0.49)、VIP > CTR(Z = 2.35,p≤0.01,r = 0.41)、SNY > CTR(Z = 2.88,p≤0.005,r = 0.51)。
讨论
以前的研究报告了不同冥想实践中局部振荡功率和远距离相干性的变化,这些变化在神经解剖学上与报告的主观体验一致,例如,枕叶γ波的增加已经与视觉图像的变化紧密联系在一起(Lutz等人,2004;Lutz等人,2007;Braboszcz等人,2017;van Lutterveld等人,2017;Kakumanu等人,2018;Luft等人,2019)。这些报告中发现的异质性突显了“冥想”一词的模糊性质(Brandmeyer等人,2019;Schoenberg和Vago,2019)。我们的分析通过显示冥想如何增加α和γ波段的信号复杂性来扩展了以前的发现,以样本熵(SE)为指标。特别是,VIP(内观)中的SE变化表现为典型的,意味着它们允许机器学习分类器正确地将EEG信号熵中的其他冥想状态推广为检测问题。与对照组相比,观察到了在毗婆舍那禅修中α和γ波段功率包络熵的增加,无论是比较禅修与呼吸集中还是在匹配对照组中指导心灵漫游。然而,我们在比较冥想者和对照组之间的“专注呼吸”条件时没有观察到差异,这表明我们的结果是冥想参与的积极体现,而不是反映特质效应。
本文进行的分析在先前研究中相对较少。一项先前的研究报告了在实践内观禅修(毗婆舍那)期间排列熵的减少(一种与样本熵密切相关的方法)(Venugopala等人,2017),这也与EEG电极记录到的信号之间的同步水平增加有关。冥想状态下γ波同步增加与我们的发现一致(Lutz等人,2004)。然而,与Venugopala及其同事报告的结果(即减少)相比,我们发现熵(即增加)的结果相反。我们注意到,这种差异可能归因于我们没有研究宽带EEG信号的熵,而是研究了窄带振荡包络的熵。如前所述,与先前的研究相反,我们的方法允许将包络熵的变化(可能与在载波频率振荡的细胞集合的大小有关)与宽带信号的谱内容区分开来,宽带信号包含了在非常不同频率上振荡的细胞集合的非特异性贡献(即1/f谱)。
在冥想期间,我们发现熵最显著增加的频段与Braboszcz等人之前的报告一致,该报告使用我们分析的相同数据集比较了冥想者和对照组之间的EEG谱功率(Braboszcz等人,2017)。他们报告了所有冥想传统与对照组相比,α(7-11 Hz)和γ(60-110 Hz)范围的枕部(Oz)EEG功率增加;参见Braboszcz等人(2017)的图3。γ波带增加位于枕顶头皮部位,而α波带增加则出现在所有电极上;分别参见Braboszcz等人(2017)的图4和图6。这些结果与图3中显示的α波和γ波振荡包络熵增加的拓扑定位一致。
我们的结果还与在宁玛派和噶举派佛教修行者的冥想状态下观察到的γ(25-56 Hz)波段长程相干性增加的现象一致。已报道毗婆舍那禅修可增加枕部-顶部头皮区域的局部γ(35-45 Hz)功率(Cahn等人,2010),与图3中报告的结果重叠。γ波振荡与诸如注意力、工作记忆、学习和意识等多种高阶认知过程有关(Fries等人,2007;Fries,2009)。γ波振荡还被认为是解决“束缚问题”的合理神经生物学机制(即在空间分布的皮层区域之间整合信息)(Nikolić等人,2013)。与我们的结果相反,Lutz等人讨论了一些与冥想状态有关的慢振荡报告(Lutz等人,2007)。然而,正如作者所指出的,这些结果可能受到两个基本因素的偏见:(1)没有研究快速振荡;(2)研究是基于专注禅修,这可能涉及与较慢脑振荡发作相关的自上而下的控制机制。
由于历史上冥想的神经生理学相关因素受到了多种因素的混淆,因此我们的结果与之前的研究相比较具有一定难度,特别是在几乎没有研究带特定振荡熵的文章的情况下。一个主要的混淆来源是所研究的冥想传统的性质。我们对冥想传统从专注禅修到开放性禅修的一维分类只是其他诸多表征之一。正如Lutz等人(2004)所言,在大多数冥想状态的电生理研究中没有测量EEG伽马波段。Thomas和Cohen(2014)的一篇最近的综述和荟萃分析文章也支持了这一观点(分别参见专注禅修、开放性禅修和超觉禅修的表2、表3和表4)。显然,专注禅修研究通常揭示了θ波和α波的增加,而开放性禅修的结果则较为不均匀。
尽管现象学上存在相似性,使用fMRI研究迷幻药物和冥想状态都发现了后扣带皮质(PCC)的活动减少,PCC是默认模式网络(DMN)的一个后部“中心”(Carhart-Harris等人,2012年;Berkovich-Ohana等人,2016年;Smigielski等人,2019年)。这种失活已被比作“减压阀”失效,导致对大脑活动配置的潜在剧目受到较少约束(Carhart-Harris等人,2012年;Carhart-Harris等人,2014年)。我们测量了窄带EEG功率波动的熵作为这种失调的定量指标。然而,我们注意到,熵不应该作为意识水平的绝对代理,因为非常高的熵意味着系统的组成部分(例如神经元)在统计意义上独立行为,阻止了信息整合。Giulio Tononi假设,测量状态增强剧目与信息整合水平之间的平衡可能比单独的熵更适合作为意识的度量(Oizumi等人,2014年)。所提出的度量(□)难以计算,但可以通过在经颅磁刺激(TMS)脉冲刺激后测量EEG进行经验近似,这种技术可以在未来的研究中用于研究冥想状态(Casali等人,2013年)。
长期冥想(Sahdra等人,2011年;Strauss等人,2014年;Haukaas等人,2018年;Pokorski和Suchorzynska,2018年)和血清素类迷幻药物的急性效应(Griffiths等人,2016年;Carhart-Harris等人,2017年;Carhart-Harris等人,2018年;Dos Santos等人,2018年;Garcia-Romeu和Richards,2018年;Palhano-Fontes等人,2018年)均已报道可提高情绪和幸福感。在迷幻药物的急性效应期间报告了大脑熵的增加,并且已经证明可以预测人格特征的变化(Lebedev等人,2016年)。最近,Carhart-Harris和Nutt建议5-HT2A信号介导了“积极应对”,这是一个以增强可塑性为特征的过程(Carhart-Harris和Nutt,2017年)。我们研究中观察到的大部分EEG熵增加可以归因于内观禅修,这个过程可以与Carhart-Harris和Nutt提出的过程相比较。单独的专注禅修可能导致一种“迟钝”的状态;因此,需要同等程度地注意由分心带来的兴奋以及这种分心的元认知。换句话说,兴奋和专注禅修之间需要平衡。内观禅修可以被概念化为一个持续增加大脑兴奋性的过程,然后在冥想者检查并从意识中排除干扰物后减少兴奋性。未来应该对内观禅修专家进行基因表达(表观遗传学)和正电子发射断层显像(PET)研究,以确定这种“退火”过程是否仅作为类比有用,或者确实是由编码5-HT2A受体的基因表达增加以及5-HT2A受体密度水平升高所介导。5-HT2A激动剂PET放射性配体(如Cimbi-36)的研发进展(Ettrup等人,2016年),为评估这一假设提供了一个充满希望的研究途径。
我们注意到,尽管已经报道了不同化合物(例如5-HT2A激动剂和NMDA拮抗剂)的急性效应导致熵增加(Schartner等人,2017a),但我们的研究结果提供了“内源性”增加的证据,即不是由药理剂介导而是由长期冥想者执行的心理过程引起的EEG熵增加。此外,我们的分析确定了哪些频谱带变得更加混乱,这些频谱带与5-HT2A激动剂主要影响的频谱带(α节律)(Muthukumaraswamy等人,2013年;Carhart-Harris等人,2016年)以及专业冥想者冥想过程中的频谱带(γ节律)(Lutz等人,2004年)相匹配。尽管迄今为止的大多数研究都从宽带振荡的角度研究熵脑假说,但我们的工作提出了新颖的结果,这些结果应促使人们从受影响频率带的角度重新审视高熵的大脑状态。
我们观察到的一致性和超稳定性的变化表明,在冥想过程中,用于特定频带振荡的细胞群的招募在局部范围内增加,而在更全局的范围内,γ带绑定的增加减少了这种库。局部和全局脑状态潜在库的变化之间的这种冲突与LSD急性影响下获得的发现一致,这些发现表明在局部隔离活动损失的代价下,全球功能整合增加(Tagliazucchi等人,2016年)。可以进行进一步的使用5-HT2A激动剂和NMDA拮抗剂的电生理研究,以允许直接比较局部熵和全局超稳定性/一致性与在冥想状态下测量的熵和一致性。
我们的研究代表了建立现象空间增强与大脑活动熵增加之间直观关系的一步,正如Carhart-Harris等人(2014)所提出的。然而,我们的研究还存在一些局限性,这些局限性源于使用EEG推断熵变化的位置,因为由于体积传导效应,来自不同皮质来源的EEG电极信号到达头皮时会“混合”。此外,尽管参与者的心理测量评估是可行的,但它不适合与致幻状态的现象特征进行比较。为此,应使用其他问卷,如意识改变状态(ASC)问卷(Dittrich,1998)。由于这个局限性,我们只能提供间接证据,即增加的熵是冥想和致幻状态之间现象相似性的基础机制。
总之,净观禅修,一种介于开放注意和专注注意之间的实践,导致了α波和低/高γ波段脑振荡包络的熵增加。这表明,局部神经配置增加的多样性是由更高水平的局部隔离以及低/高γ波段的全局一致性增加所介导的。我们的研究提供了一个非药物诱导的高熵脑状态的例子。未来的研究应该探讨这些“内源性”的EEG熵增加是否与资深禅修者5-HT2A受体表达的变化有关,并研究熵、灵活性以及某些禅修传统对身心健康的长期积极影响之间的关系。

















 2万+
2万+

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









