







【Title】Cell fate roadmap of human primed-to-naive transition reveals preimplantation cell lineage signatures
【Publication Book】Nature communications
【Publication Time】2022 Jun
【Abstract】Human naive pluripotent stem cells offer a unique window into early embryogenesis studies. Recent studies have reported several strategies to obtain cells in the naive state. However, cell fate transitions and the underlying mechanisms remain poorly understood. Here, by a dual fluorescent reporter system, we depict the cell fate dynamics from primed state toward naive pluripotency with ALPG activation followed by the activation of OCT4-distal enhancer. Integration of transcription profiles and the chromatin accessibility landscape reveals the appearance of primitive endoderm and trophectoderm signatures in the transitioning subpopulations, with the capacities for derivation of extra-embryonic endoderm and trophoblast stem cell lines, respectively. Furthermore, despite different fluorescent dynamics, all transitioning intermediates are capable of reaching the naive state with prolonged induction, showing their developmental plasticity and potential. Overall, our study describes a global cell roadmap toward naive pluripotency and provides hints for embryo modeling-related studies.
【Background】
人类天真多能干细胞(Naive PSCs)捕获了与植入前上胚层(preimplantation epiblast)相对应的地面多能状态,与处于原始多能状态的传统多能干细胞(Primed PSCs)相比,它表现出更强的可塑性和无偏差分化潜能,从而为发育研究和治疗应用提供了一个取之不尽的模型。
由于在培养系统优化方面取得了重大突破,目前已开发出多种实现幼稚多能性的策略。天真多能性造血干细胞可通过直接从植入前胚胎中衍生、体细胞重编程或传统多能性造血干细胞在原始状态下的转换产生。
据报道,在建立幼稚多能性过程中发生了一些分子事件: 表面标志物 SSEA4 在直接从人类胚胎植入前囊胚或处于primed状态的原始多能干细胞衍生出naive多能干细胞的过程中消失;OCT4 增强子的活性在细胞从primed状态过渡到naive状态的过程中从近端增强子(PE)转向远端增强子;ALPG(又称 ALPPL2)的表达在primed状态或体细胞状态的细胞建立naive多能性的过程中获得。
在本研究中,为了精确监测细胞在唤醒状态下的幼稚多能性建立,我们构建了一个由ALPG-promoter-RFP和OCT4-ΔPE-GFP组成的双荧光报告系统,并追踪了这一过程中的荧光动态。通过大量RNA-seq(scRNA-seq)和单细胞RNA-seq(scRNA-seq)分析,转录谱分析显示了向幼稚多能性过渡的轨迹,ALPG激活后,OCT4-远端增强子也随之激活。染色质可及性动力学(CAD)的整合分析表明,在从primed诱导到naive诱导的过程中,过渡亚群中先后出现了原始内胚层(PrE)特征和TE特征,具有强PrE或TE特征的中间产物可分别生成胚外内胚层细胞系或滋养层干细胞(TSC)系。此外,尽管荧光动态不同,但所有过渡中间细胞都能在5iLAF培养系统的长期诱导下达到幼稚多能状态,这显示了它们的发育可塑性和潜力。总之,我们的研究描述了通向幼稚多能性的高分辨率细胞路线图,为人类囊胚建模和早期胚胎发生研究提供了宝贵的资源。
【Result】
1.双报告系统监控从primed-to-naive过渡过程中的荧光动态
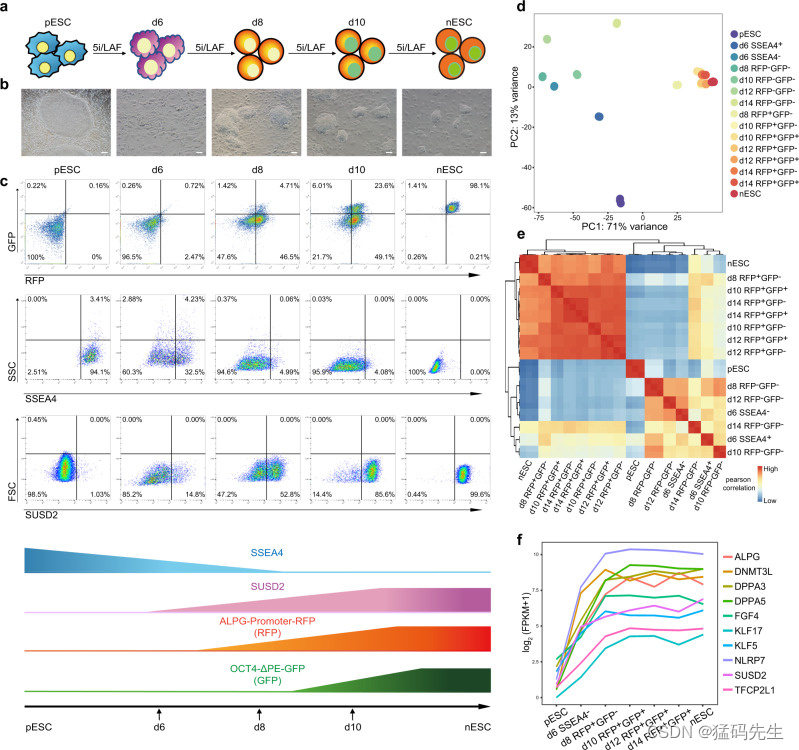
为了精确研究引物向天性转化过程中的动态变化,我们构建了一个双荧光报告系统。一个是ALPG-promoter-RFP(以下简称RFP),其中RFP与ALPG转录起始位点(TSS)上游3 kb的DNA序列融合21;另一个是OCT4-ΔPE-GFP(以下简称GFP),其中OCT4 PE元件被删除,OCT4的表达主要依赖于其远端增强子的活性,如之前报道的那样12。然后,我们用该报告系统对原始胚胎干细胞(pESCs)进行了基因工程,并在 5iLAF 培养条件下进行了primed-to-naive的重置(图 1a)。诱导第6天观察到小菌落,此时SSEA4(引物状态特异性表面标记)的表达急剧下降,一小部分细胞开始表达SUSD2,这是最近报道的天真状态特异性表面标记28(图1b,c)。第 8 天出现了类似天真克隆的圆顶形菌落,表达 RFP 的细胞亚群大大增加,比例与 SUSD2+ 细胞相似(图 1b,c)。此时,几乎所有细胞都变成了 SSEA4 阴性。GFP 表达与 RFP 表达不同步。直到第 10 天才能观察到 GFP+ 亚群。在诱导过程中,RFP+GFP+细胞的比例不断增加,在第14天时,挑选出类似天真细胞的集落,建立天真ESC(nESC)系(图1b、c)。
2.从primed-to-naive过渡期间中间体的转录谱分析
接下来,我们收集了primed-to-naive过程中具有不同荧光动态的过渡中间体,并对它们进行了批量RNA-seq分析(补充数据1)。主成分分析(PCA)表明,从第 8 天开始收集的 RFP+ 细胞与 nESCs 紧密聚集在一起,并与 pESCs 区分开来(图 1d,e)。当与人类胚胎数据集整合时,RFP+ 细胞和 nESCs 与 ICM 细胞紧密聚类,而 pESCs 在转录组上与已报道的 P0 和 P10 期囊胚衍生的 hESCs 极为相似(补充图 1a)。

在趋向于幼稚多能性的过程中,外胚层特异性标记基因从第 6 天开始上调(图 1f;补充图 1b),而引物状态特异性基因则逐渐下调(补充图 1b,补充数据 2)。

接下来,我们根据基因表达的动态(至少一个样本的 FPKM ≥ 5)对表达集群进行了表征,并确定了六种主要的表达模式(补充图 1c-h,补充数据 1)。参与干细胞维持和胚胎形态发生的原始多能性相关基因迅速下调,包括 OTX2、ZIC2 和 ZIC3(补充图 1c),而天真多能性相关基因在三类中被激活: 其中一组富含对幼稚多能性调控和 mRNA 处理至关重要的基因,如 NANOG、TFAP2C、LIN28B 和 DPPA3,这些基因从第 6 天开始表达增加(补充图 1d);另一组富含与胚胎发育和蛋白质修饰相关的基因,如 DNMT3L 和 NODAL,这些基因在第 8 天达到表达峰值(补充图 1e);第三组富含与胚胎发育和蛋白质修饰相关的基因,如 DNMT3L 和 NODAL,这些基因在第 8 天达到表达峰值(补充图 1f)。1e);第三组富含参与氧化磷酸化代谢的基因以及核心幼稚多能性标记 ALPG 和 UTF1,其表达在第 10 天达到峰值(补充图 1f)。在第 6 天,SSEA4- 细胞表现出独特的特征:一个群集富含与 PrE 发育(GATA4、GATA6)、细胞外基质组织(KRT8、COL1A1)和胚胎形态发生(HAND1、HAND2、HOXB4)相关的基因(补图 1g),另一个群集富含与共享多能性标记相关的基因,包括 POU5F1、SOX2、SALL4 等、 补充图 1h)。因此,批量 RNA-seq 分析揭示了primed-to-naive过渡过程中不同的基因表达模式。
3.以单细胞分辨率描述细胞群primed-to-naive转变过程中的特征

第 8 天的 RFP+ 细胞与第 10、12 和 14 天的 RFP+GFP- 或 RFP+GFP+ 细胞以及 nESCs 的转录相似(图 1d、e;补充图 1a)。多个天真多能性相关基因,如ALPG、DNMT3L、DPPA3、KLF17和SUSD2,在引物向天真过渡期间表现出强劲的上调,其表达水平从第8天开始达到与天真多能性相当的状态(图1f)。为了以单细胞分辨率进一步描述过渡时期细胞群的特征,我们对过渡过程中第 6、8、10 和 14 天收获的细胞以及人类 nESCs 和 pESCs 进行了基于液滴的 10× Genomics scRNA-seq,产生了包含 38,036 个细胞和 16,929 个常见基因的数据集。力导向布局(FDL)30 显示了中间产物的过渡轨迹以及单个细胞在有底物到无底物过渡过程中的关系(图 2a、b)。我们还利用多种降维方法,如均匀流形近似和投影(UMAP)(补图 2a-b)和 t 分布随机邻接嵌入(tSNE)(补图 2c-d),将低维空间中的细胞嵌入可视化,从而证实了上述发现。

结合共享多能性(POU5F1、PRDM14、NANOG、LEFTY1、TDGF1)、primed多能性(ZIC2、SOX11)和naive多能性(DNMT3L、DPPA3、ALPG、DPPA5、FGF4)的已知标记基因的表达模式(图 2c-e,补图 3a-b),我们通过无监督聚类分析(图 2f,补图 3c-d)确定并表征了 15 个过渡过程中的集群。通过无监督聚类分析(图 2f,补充图 3c-d),我们确定并表征了过渡过程中的 15 个集群。) 根据不同文库或聚类中的细胞比例,我们观察到具有幼稚多能性特征的细胞群从第 8 天开始大大增加(图 2g,补图 3e)。此外,第 8 天出现的 ALPG 表达亚群还显示出其他天真标志物的强健表达,如 DPPA3、DPPA5 和 FGF4(补充图 3f),这与我们的批量 RNA 序列分析结果一致。
4.primed-to-naive转化过程中染色质可及性的动态变化
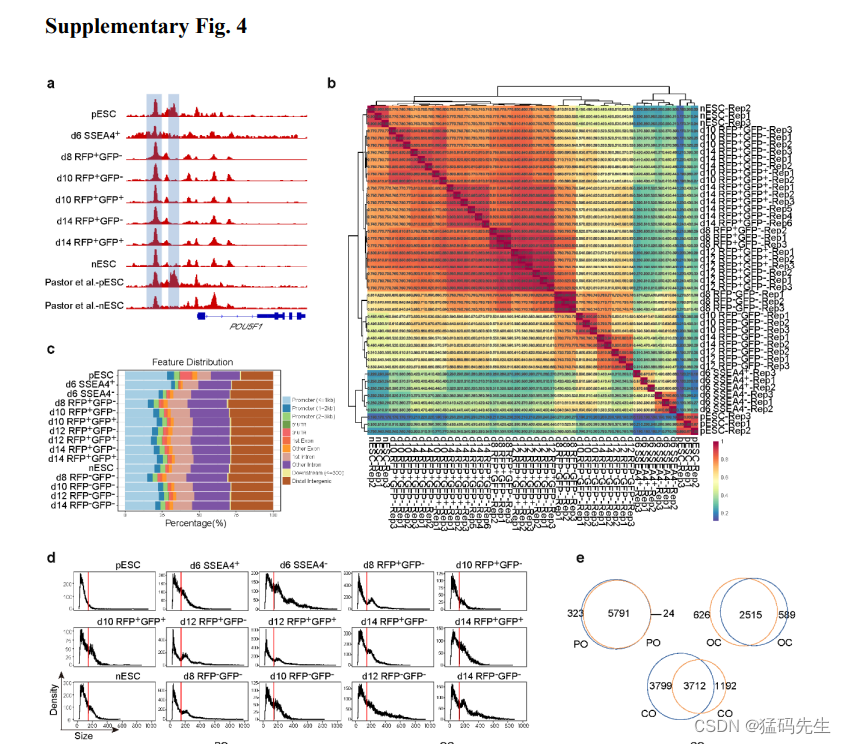
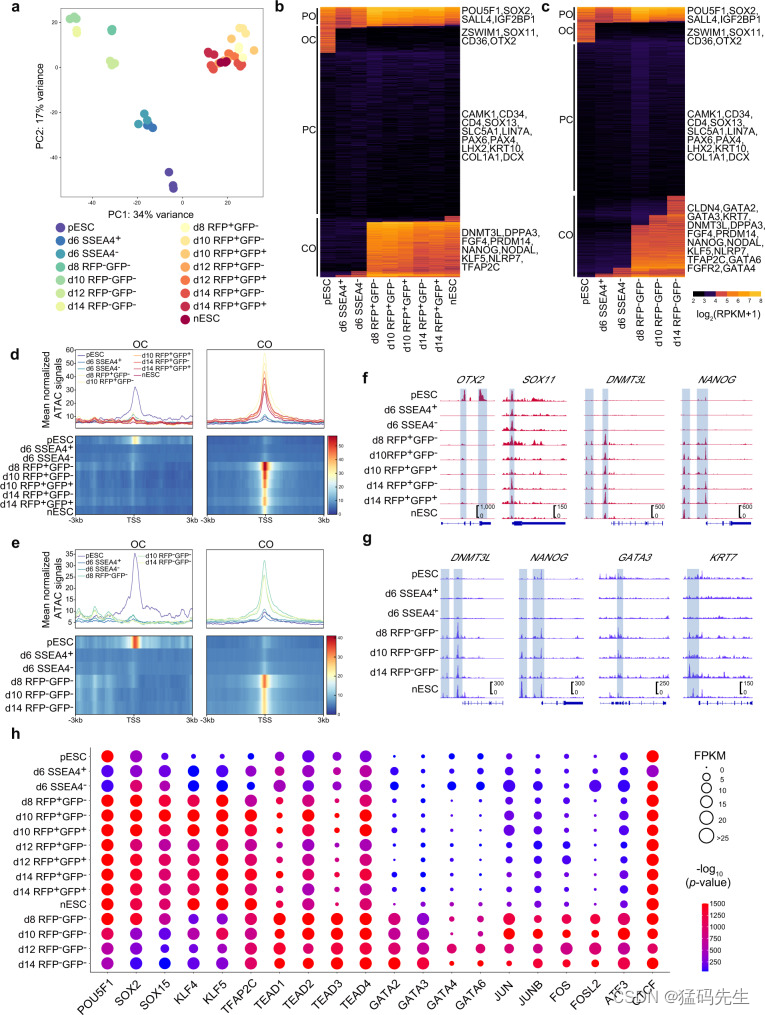
接下来,我们尝试通过转座酶可及染色质测序(ATAC-seq),根据中间体的荧光动态(补充数据 3-4)来说明primed-to-naive转变过程中染色质的可及性状况。在primed-to-naive转变过程中,观察到 OCT4 增强子活性从 PE 向远端增强子转化(补充图 4a)。对重复次数、峰值富集区和峰值大小分布的分析表明获得了高质量的 ATAC-seq 数据集(补充图 4b-d)。
PCA 显示,所有 RFP+ 细胞都与不同时间点的 nESC 有很好的相关性(图 3a),这表明从第 8 天开始 ALPG 激活的细胞具有相似的染色质可及性状态。有趣的是,除RFP+细胞外,仍处于RFP-状态的细胞紧密聚集在一起(图3a;补充图4b),这表明有ALPG激活和没有ALPG激活的细胞可能有不同的染色质景观和细胞命运。然后,我们重点研究了两种情况下染色质图谱的差异:在从原始细胞向幼稚细胞过渡的过程中,细胞转为 RFP 阳性和细胞保持 RFP 阴性。然而,CAD 制图显示了两种条件下的模式相似性。(图 3b-e;补充图 4e): 参与 DNA 修复、蛋白质稳定性和子宫内胚胎发育的基因位点在两种条件下都是永久开放的(PO),包括共享的多能因子 POU5F1 和 SOX2 的基因位点(图 3b,c;补充图 4e-f);分化相关基因的基因位点是永久封闭的(PC),包括 SOX13 和 PAX6 的基因位点(图 3b,c)。3b,c);参与 mRNA 代谢过程和生长因子反应的基因位点在两种条件下都经历了从开放到封闭的转变(OC),包括引物状态特异因子 SOX11 和 OTX2(图 3b-f;补充图 4e,f)。
与 PO 或 OC 基因位点在两种条件下都表现出极高的重合度不同,经历了从封闭到开放(CO)转变的基因位点在一定程度上显示出两种条件下的差异(补充图 4e)。天真多能性相关基因(如 DNMT3L 和 NANOG)的基因座不仅在向天真多能性转变的细胞中被打开,而且在 RFP 阴性的中间体中也被打开(图 3b-g),这表明这些 RFP 阴性的细胞可能仍然具有达到天真多能性状态的潜力。此外,TE 标记(GATA3、KRT7 等)和 PrE 标记(GATA4、GATA6、FGFR2 等)的位点都特别位于 RFP 阴性的中间产物的 CAD 的 CO 类别中(图 3c、e、g;补充图 4g)。对CO类别内的基因进行的GO分析表明,这些基因分别参与了胚胎上皮管形成和由向幼稚多能性发展的细胞和保持RFP阴性的细胞组成的CAD中干细胞群的维持(补充图4f)。
我们还对ATAC-seq数据集进行了基序富集分析。在primed-to-naive转化过程中,naive TFs(POU5F1-SOX2-TCF-NANOG、SOX2/15和KLF4/5)的基序在RFP+细胞中显著富集(图3h)。TE特异性TF(TEAD家族和GATA2/3)和在TSCs中高表达的TF(JUN、JUNB、ATF3、FOS和FOSL231)的基序在RFP-细胞中富集,这与其相应的TF表达相吻合(图3h)。从第8天开始,TFAP2C基序在RFP+和RFP-细胞中都被富集(图3h),这与TFAP2C基因在幼稚造血干细胞和造血干细胞中的激活一致。因此,ATAC-seq数据集的分析显示,RFP-细胞在primed向naive过渡过程中具有明显的TE特征。
5.primed-to-naive过渡期间的 TE 信号
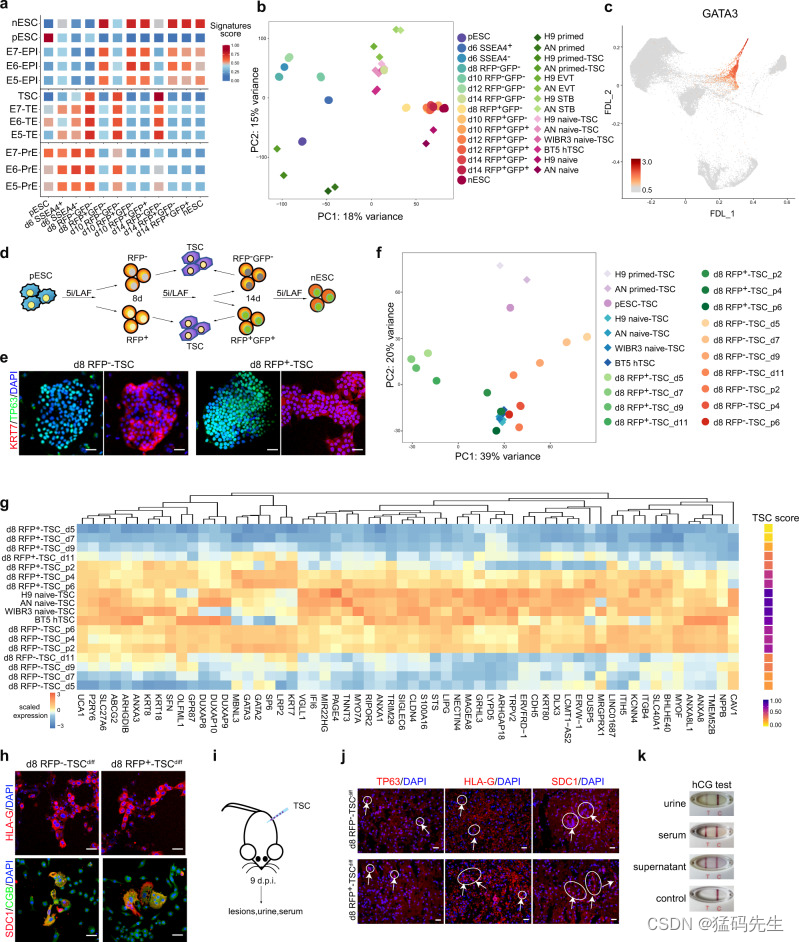
我们通过 RNA-seq 分析进一步确定了 RFP- 中间体的 TE 特征。我们进行了评分分析,以研究中间细胞在过渡过程中的不同胚胎特征(图 4a;补充数据 2)。不出所料,在原始细胞向幼稚细胞过渡期间,幼稚状态和 EPI 的特征在 RFP+ 细胞中上调并保持不变,而原始状态的特征仅在 pESCs 中富集(图 4a)。我们还观察到,在 RFP- 中间体中,TE/TSC 的特征非常丰富(图 4a)。此外,当我们的批量 RNA-seq 数据集与已发表的 TSC 数据集32 整合时,第 14 天的 RFP-GFP- 细胞与来自 nESCs 或囊胚的 TSCs 紧密聚类(图 4b;补充图 5a)。具有代表性的 TE 标记,如 GATA2/3、KRT7 和 TP63,在 RFP- 中间体中特异表达(补充图 5b),证实了它们与 TE/TSC 在转录程序上的相似性。scRNA-seq 分析还显示,RFP- 细胞的一个亚群特异表达 TE 相关基因(图 4c;补充图 5c)。因此,结合 ATAC-seq 分析的结果,我们证实在 RFP- 中间体中出现了具有 TE 特征的 TE 样亚群。
6.从过渡中间细胞中衍生出 TSCs
接下来,我们推测这种类似 TE 的亚群可能会在体外产生 TSCs。我们通过流式细胞术收集了第 8 天和第 14 天的 RFP- 中间体,并按照之前的报道33 将其培养在 TSC 培养基中(图 4d)。由于 TSCs 可从幼稚 PSCs 中生成7,32,我们也对中间 RFP+ 细胞进行了 TSC 诱导作为对照。不出所料,第 8 天或第 14 天,RFP- 和 RFP+ 细胞都成功生成了具有代表性菌落形态和高表达 TP63 和 KRT7 等 TSC 标记的 TSCs(图 4e;补充图 5d-e),而 pESCs 则未能建立稳定的 TSC 株系,这与之前的研究结果一致32(补充图 5f)。转录谱分析显示,在primed-to-naive重置的第8天,从RFP-和RFP+中间产物衍生的TSCs与已发表报告中描述的TSCs具有高度相似性32(图4f;补充数据5),这进一步表明这些过渡中间产物具有生成TSCs的能力。有趣的是,在TSC诱导过程中,RFP-细胞和RFP+细胞可能采用不同的途径建立TSCs,最终都会激活TSC相关基因表达程序(图4f,g)。此外,我们还观察到,与 RFP+ 细胞相比,RFP- 中间体在早期时间点显示出滋养细胞诱导加速的现象(补图 5g)。
我们进一步评估了从第8天的中间产物中提取的TSCs的分化潜力。在特定条件下培养时,这些细胞可分化为具有特异性 HLA-G 表达的绒毛外滋养层细胞(EVT)和具有 SDC1 和 CGB 表达的合体滋养层细胞(ST)33(图 4h)。当皮下注射到 NOD-SCID 小鼠体内时,这些 TSCs 会产生病变,如免疫染色法观察到的 TP63+细胞、SDC1+ ST 样细胞和 HLA-G+ EVT 样细胞所示(图 4i,j)。此外,通过妊娠试验棒测定,在注射了 TSCs 的宿主小鼠的 ST 细胞培养物上清液、尿液和血清中检测到了人绒毛膜促性腺激素(hCG)(图 4k;补充图 5h)。综上所述,这些研究结果表明,具有功能分化能力的TSCs可以从RFP-细胞中建立,而RFP-细胞在原始细胞向无细胞过渡期间表现出TE特征。
7.primed-to-naive过渡期间的 PrE 信号
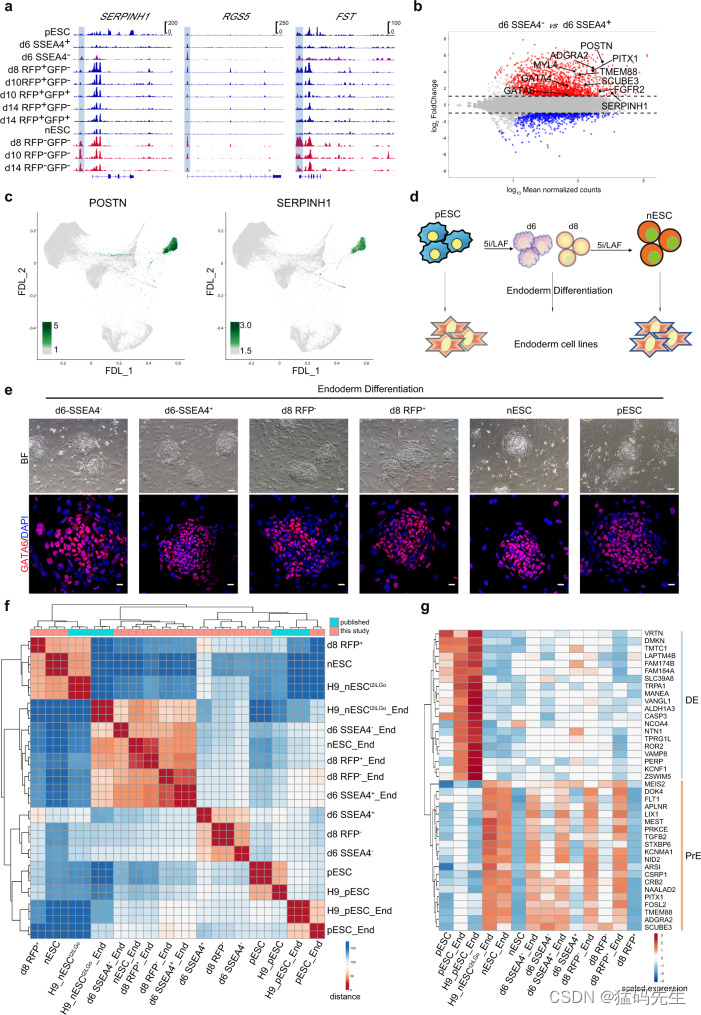
我们还观察到在原始细胞向幼稚细胞过渡的过程中出现了 PrE 标志(图 3h;图 4a;补充图 1g)。ATAC-seq 数据集的基序富集分析表明,RFP-细胞中的内胚层标志物 GATA4 和 GATA6 的基序富集(图 3h)。与 RFP+ 中间体相比,PrE 相关基因位点的 ATAC-seq 信号在 RFP- 和第 6 天-SEA4-中间体细胞中更为富集(图 5a)。通过评分分析,我们还观察到第 6 天-SSEA4-中间细胞和第 8 天-RFP-细胞中的 PrE 特征都非常丰富(图 4a)。总之,这些数据进一步表明了 PrE 标志的出现。
接下来,我们试图鉴定过渡过程中的类PrE亚群。如上文所述,RNA-seq分析显示,在第6天的过渡中间体中,PrE相关标记短暂激活,核心多能性标记失活(补充图1g-h,补充数据1)。差异表达分析证实了 SSEA4 表达缺失后 PrE 相关基因的快速上调(图 5b;补充图 6a)。第 6 天 SSEA4- 细胞亚群的特征是 30 个差异表达(DE)最多的基因,如 POSTN 和 PITX1,也可以通过一系列 PrE 标记的表达来确定,包括 SERPINH1、FGFR2、MYL4 和 TMEM88(图 5b, c; 补充图 6b)。
然后,我们将过渡过程第 6 天和第 8 天的中间产物以及 nESCs 和 pESCs 分别进行内胚层分化34(图 5d)。所有衍生的内胚层细胞都能作为稳定的细胞系维持和扩增,具有胚胎内胚层细胞的典型形态,内胚层标记基因 GATA6 的表达也很强(图 5e)。由 pESC 衍生的内胚层细胞(命名为 pESC_End 细胞)显示出最终内胚层(DE)特征,一系列 DE 基因高表达(图 5g,补充图)。图 5g,补图 6c),而在原始细胞向幼稚细胞过渡的过程中,由第 6 天的 SSEA4+ 和 SSEA4- 细胞、第 8 天的 RFP- 细胞以及 nESC 衍生的内胚层细胞系表现出强烈的 PrE 特征,富集表达 PrE 相关基因(图 5g,补图 6c)。因此,从有丝分裂到无丝分裂的过渡中间体以及 nESCs 具有分化成具有 PrE 特征的内胚层细胞的潜力。
7.长期诱导向幼稚多能性过渡的 RFP- 中间体
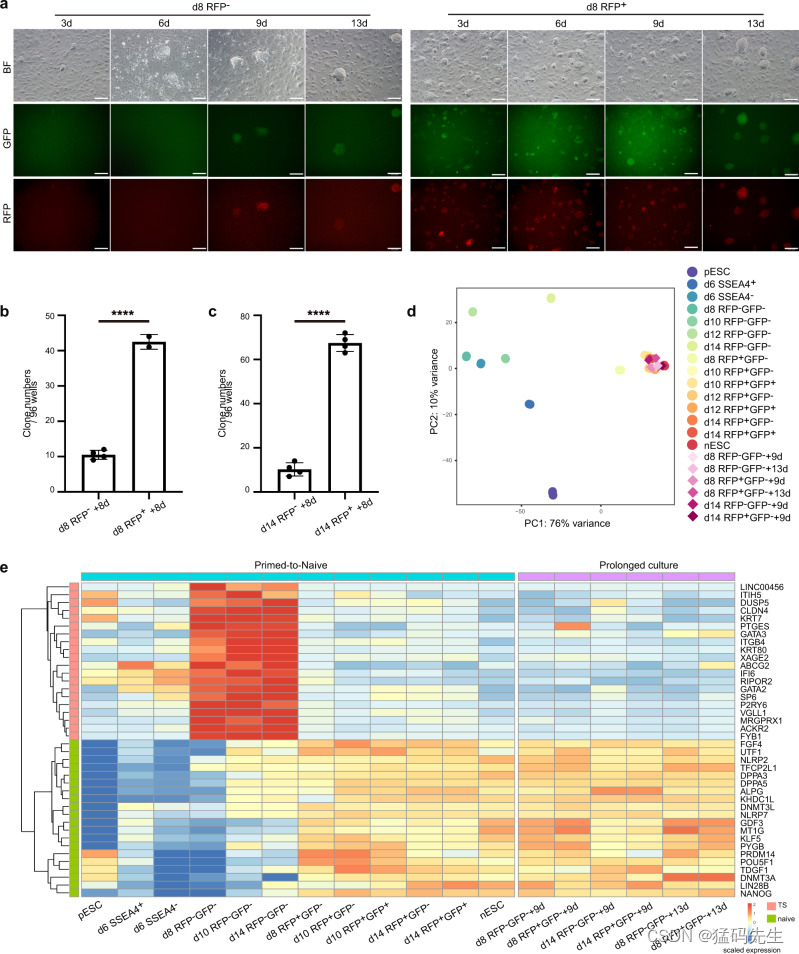
由于我们通过FDL观察到中间细胞有向幼稚多能性过渡的趋势(图2a),并且通过CAD图观察到RFP-中间细胞中与幼稚状态相关的基因位点逐渐开放(图3c),我们想知道RFP-中间细胞是否也能通过在5iLAF培养基中长期诱导达到与RFP+细胞类似的幼稚状态。虽然第 8 天-RFP-细胞在 5iLAF 长时间培养的前 6 天仍为荧光阴性,但在 5iLAF 培养基中长期诱导 9 天后,大多数细胞都变成了 RFP+GFP+ 细胞(图 6a)。同时,第 8 天 RFP+ 细胞比第 8 天 RFP- 细胞更快转化为 RFP+GFP+ 细胞(图 6a)。亚克隆实验也验证了这些结果,实验结果表明,在长时间的 5iLAF 培养 8 天后,第 8 天-RFP+细胞比第 8 天-RFP-细胞形成幼稚集落的效率更高(图 6b)。在延长 5iLAF 培养 RFP+ 和 RFP- 细胞的第 14 天,也能观察到类似的结果(图 6c)。在 5iLAF 培养基中长时间诱导后,这些 RFP- 中间体最终达到了类似于 nESCs 的状态(图 6d、e;补充数据 5)。总之,这些结果证明了 RFP- 中间体在从原始向幼稚过渡过程中的幼稚状态诱导潜能和发育可塑性。
8.细胞从 primed-to-naive过程的命运路线图
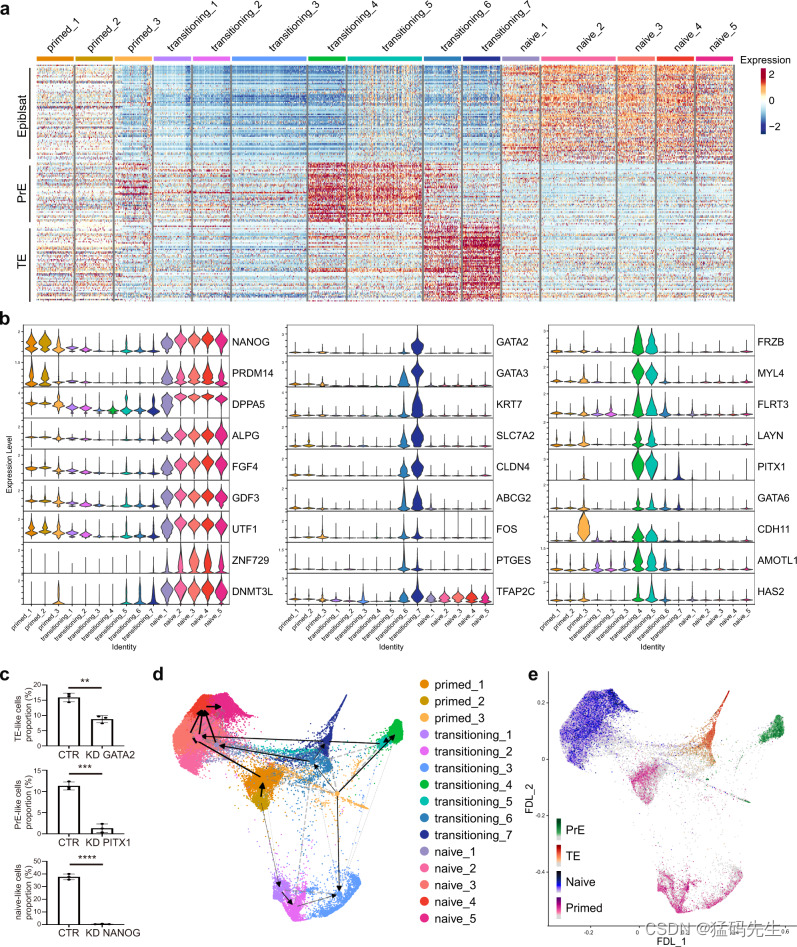
最后,我们试图描述从primed到naive过渡过程中的全局细胞命运轨迹。结合上述结果,我们根据相应的基因特征和标记基因的表达(图 7a;补充图 7a),确定了具有 PrE 标志的过渡亚群(过渡亚群 4 和 5)、具有 TE 标志的过渡亚群(过渡亚群 6 和 7)和具有 naive 标志的过渡亚群(图 7b),如 PITX1 和 GATA6 代表 PrE 标志,GATA2 和 GATA3 代表 TE 标志,NANOG 和 ALPG 代表 naive 标志。此外,与对照组相比,敲除其中一些分支依赖性转录因子可显著降低相应亚群的比例(图 7c)。
我们还进行了基于分区的图抽象(PAGA)35 轨迹推断(图 7d,补图 7b-c)和 RNA 速度36,37 计算(图 7d,补图 7d),以预测单个细胞的未来状态。这些结果与可重建转录组事件时序的潜伏时间推断分析(补图 7e-f)一起表明,虽然一小部分处于原始状态的细胞可直接达到幼稚状态,但大多数细胞仍经历了更复杂的过渡:首先失去原始多能性,然后在亚群中相继出现 PrE 和 TE 标志,最终获得幼稚多能性(图 7e)。
总之,这些结果显示了细胞从原始状态走向幼稚状态的命运路线图,其中具有 PrE 和 TE 标志的亚群相继出现,大多数细胞最终达到具有幼稚标志的状态。















 9952
9952











 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









