






摘要:性和性别与人类在整个生命周期中以及健康和疾病中的行为有关,但它们是否与相似或不同的神经表型有关尚不清楚。在这里,我们证明,在儿童中,性和性别独特地反映在大脑的内在功能连接中。躯体运动、视觉、控制和边缘网络优先与性别相关,而性别的网络相关性更分布在整个皮层中。这些结果表明,性和性别不仅在社会中而且在生物学中都是不可简化的。
1. 引言
在过去的二十年里,性、神经生物学和行为之间的相互作用得到了广泛的研究。然而,这些研究经常报告相互矛盾的结果并且无法复制。关于性别差异的文献越来越多和许多报告的差异缺乏可重复性表明我们在如何研究、解释和报告与性别相关的结果方面存在潜在的偏见和/或误解。最近,研究人员开始质疑这些观察到的男性和女性之间的差异是由生物学(例如,性别)驱动的,还是它们是否是社会建构(例如,性别)的表现。现实情况更复杂,因为性和性别都受到生物和社会因素的影响。至关重要的是,生物和社会因素之间的关联在本质上是交织在一起和相互的。例如,个人一生中的经历是由个人的性别和性别以及他们所处的社会文化环境决定的;复杂的关系汇聚在一起,影响大脑的组织和功能。在这里,我们使用术语“性别”来表示个体出生时的身体解剖学、生理学、遗传学和/或激素的特征,我们使用术语“性别”来表示个体的态度、感受和行为特征。有关这些术语以及性与性别之间可能存在的复杂关系的详细讨论,我们建议读者参考我们的补充文本。迄今为止,生物医学研究主要集中在了解性对大脑和行为的影响上。因此,性别的贡献在很大程度上是未知的。
我们人类经验的一个基本方面是我们的性和性别,我们如何看待它们,以及他人如何看待它们。性和性别可以解释我们的行为,并影响我们整个生命周期的健康和疾病。女性、出生时被指定为女性的人 (AFAB) 和性别/性别少数群体历来被排除在生物医学研究之外。因此,这组人更有可能被低诊或误诊为常见的脑部疾病 [例如,注意力缺陷/多动障碍 (ADHD),并因治疗干预(例如药物)而产生不良反应。在脑科学中,精神疾病的患病率和表达以及寻求治疗的行为存在性别和性别差异。AFAB人更有可能符合情绪和焦虑障碍的标准,而出生时被指定为男性(AMAB)的人更有可能被诊断出患有物质使用和注意力缺陷障碍。AFAB 患者更有可能报告情绪问题并寻求精神疾病的治疗。近年来,研究人员试图将精神疾病表现的这些差异与功能性大脑组织模式联系起来。然而,这一领域的工作在很大程度上是假设观察到的差异是性别的产物,而不是性别的产物。此外,研究性和性别神经科学的研究历来试图确定(二元)性别之间的基本生物学差异。在生物医学研究中,性和性别经常被混为一谈,基于错误的假设,即它们是由相同的因素决定的,并且两者彼此直接相关。然而,性和性别是与许多生物、社会和环境因素相关的复杂多维结构。了解性别和性别的独特大脑功能性相关性对于研究男性和女性之间表现出差异的大脑相关疾病至关重要。
在这里,我们试图描述和解开儿童性和性别的神经生物学基础。为此,我们使用基于大脑的预测量化了来自青少年脑认知发展 (ABCD) 研究的大量儿童样本中与指定性别和性别相关的功能网络。值得注意的是,我们在机器学习上下文中使用术语“预测”来指代估计个人性别或性别的算法的输出。首先,使用基于大脑的预测建模方法,我们证明了性别和性别都与功能连接的个体变异性有关。接下来,评估共享或不同的功能连接是否与性和性别相关,我们确定尽管关联存在一些重叠,但性和性别在大脑中是唯一代表的。最后,表征性和性别的功能网络相关性,我们揭示了性别优先与躯体运动、视觉、控制和边缘网络相关,而性别的网络相关性在整个大脑中分布更多。总的来说,这些结果表明,性和性别都与个体功能连接有关,这些关联可能是大脑相关疾病中存在的性别和性别差异的基础。
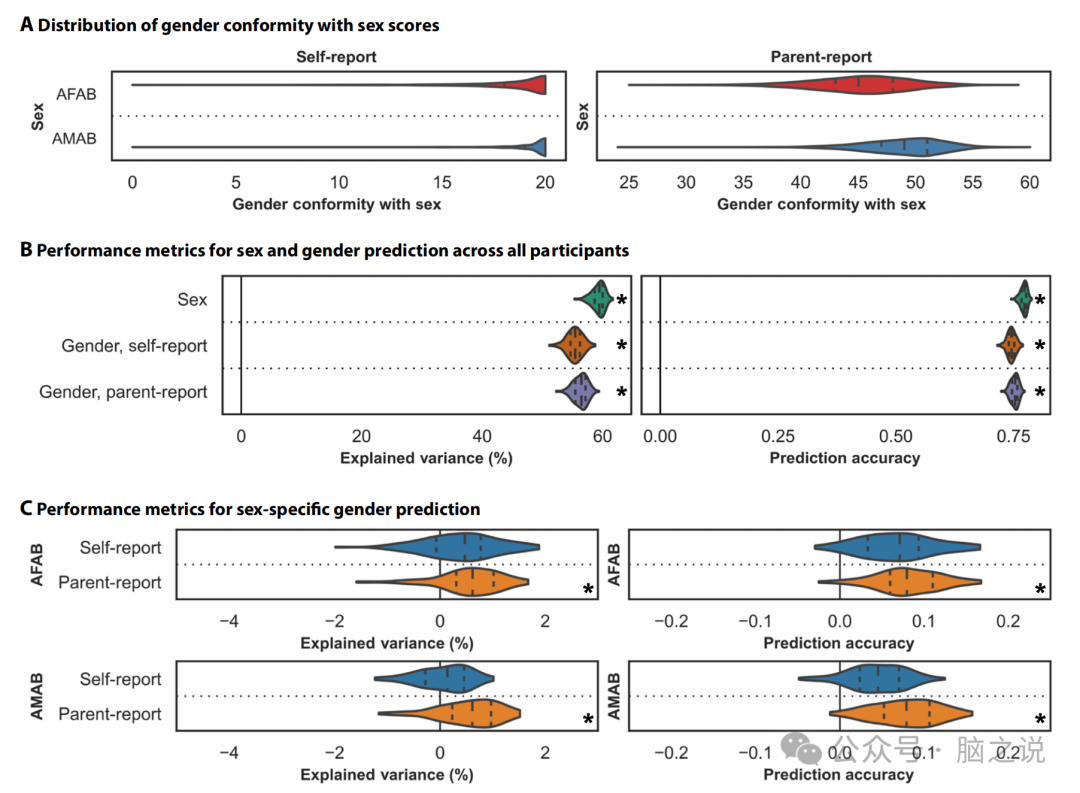
图1 功能连接与指定的性和性别有关
2. 结果
2.1 儿童 AMAB 的性别一致性高于儿童 AFAB
利用基线时 ABCD 研究的神经影像学数据,以及 1 年随访时间点自我和父母报告的性别数据,我们首先评估了性别评分的性别差异(所有参与者的图 1A)。自我报告的性别分数衡量感受性别、性别表达和性别满足度,而父母报告的性别分数衡量游戏期间的性别类型行为和性别焦虑。在自我报告和父母报告的性别测量中,分数越高表示性别一致性越高,性别一致性是指个人的性别与其指定性别的一致程度。AFAB 儿童的自我报告和父母报告的性别评分更相似,并且 AMAB 儿童在父母报告中表现出比 AFAB 儿童更高的性别一致性 和自我报告措施。这些趋势与之前在整个 ABCD 样本中报告的趋势一致,表明这些分析中使用的具有神经影像学数据的子样本代表了整个队列。现存文献表明,AMAB 儿童比 AFAB 儿童感受到更大的符合性别规范的压力。这可能在一定程度上解释了我们的结果,其中AMAB儿童报告的性别一致性比AFAB儿童强。
2.2 性和性别与功能连接的个体差异有关
使用交叉验证的线性脊回归模型,我们量化了功能连接与性别以及性别之间的关联。在所有个体中,59.27% 的功能连接方差与性别相关,55.37%的方差与自我报告的性别相关,56.30%的方差与父母报告的性别相关(图 1B)。这些对所有个体的性别预测可能会因性别而混淆(反之亦然),因为性和性别无疑是相互关联的。
为了进一步区分性别与性别的功能相关性,我们量化了功能连接与性别之间的性别特异性关联。这些模型在 AFAB 或 AMAB 儿童中分别训练以预测性别。我们的模型没有成功预测任何性别的自我报告的性别评分。另一方面,0.56%和 0.55%功能连接方差分别与 AFAB 和 AMAB 个体的父母报告的性别评分相关(图 1C)。作为对照,我们还评估了大约一半的参与者的功能连接与性别相关的程度(大致对应于用于性别特异性性别分析的样本量)。在这里,54.14%的功能连接方差与性别相关。这表明性别和性别预测之间报告结果的差异不仅仅是由样本量驱动的。此外,多项研究表明,功能连接受性别影响。在这里,我们在儿童中复制了这些发现,并进一步证明功能连接也与父母对孩子性别的报告有关。
2.3 共享和独特的功能网络与儿童的性别和性别有关
将 Haufe 变换应用于从模型中提取的特征权重,以提高其可解释性和可靠性,并将绝对 Haufe 变换的权重平均以计算平均绝对特征重要性分数。我们评估了从不同预测模型中提取的特征之间的相关性(图 2A)。对于与性别无关的性别预测模型,与性别相关的功能连接在很大程度上与与性别相关的功能连接重叠,这表明经过训练以捕获性别变异性的模型正在捕获与性别相关的变异,反之亦然。对于性别特异性模型,与性别相关的功能连接与 AFAB 中与性别相关的功能连接不同 和 AMAB子项。对于自我报告测量,与性别相关的功能连接在性别之间呈弱相关性,但对于父母报告测量则不同。最后,与 AFAB 儿童的自我报告和父母报告测量相关的功能联系呈中等相关性,但在 AMAB 儿童中不相关。这些结果表明,性和性别虽然密切相关,但在功能网络中具有唯一代表性。
最后,我们通过将区域特征权重映射到 17 个大规模皮层网络和一个非皮层网络(图 2B)上,建立了性别和性别的网络级功能相关性。在这里,我们将分析的重点放在性别的功能相关性和性别特异性父母报告的性别测量之间的重叠上,因为自我和父母报告的性别测量的性别无关相关性与性别的相关性几乎相同,而性别特异性自我报告测量的功能相关性(图 S3)对应于表现不佳的模型。我们评估了图 2B 所示的网络级特征权重之间的相关性。与性别相关的网络水平连接在性别之间呈中等相关性。这种关系比在 AFAB或 AMAB儿童中观察到的性别和性别网络相关性之间的关系更强。这进一步证明了性别的功能网络相关性与性别的相关性不同。这些结果还表明,两性之间存在共享和独特的性别相关性。
性别的功能网络相关性主要存在于躯体运动、视觉、控制和边缘网络中,而性别的网络相关性在整个皮层网络中更加分散。在 AFAB 儿童中,性别的网络相关性主要涉及颞顶叶、边缘、背侧/腹侧注意力和躯体运动网络内部和之间的联系。在 AMAB 儿童中,性别的网络相关性包括颞顶叶内部和之间的连接、默认模式、边缘、背侧/腹侧注意力、躯体运动和视觉网络。基于这些发现,我们可以推测单峰和异峰网络内部/之间的不同功能联系与性和性别有关。
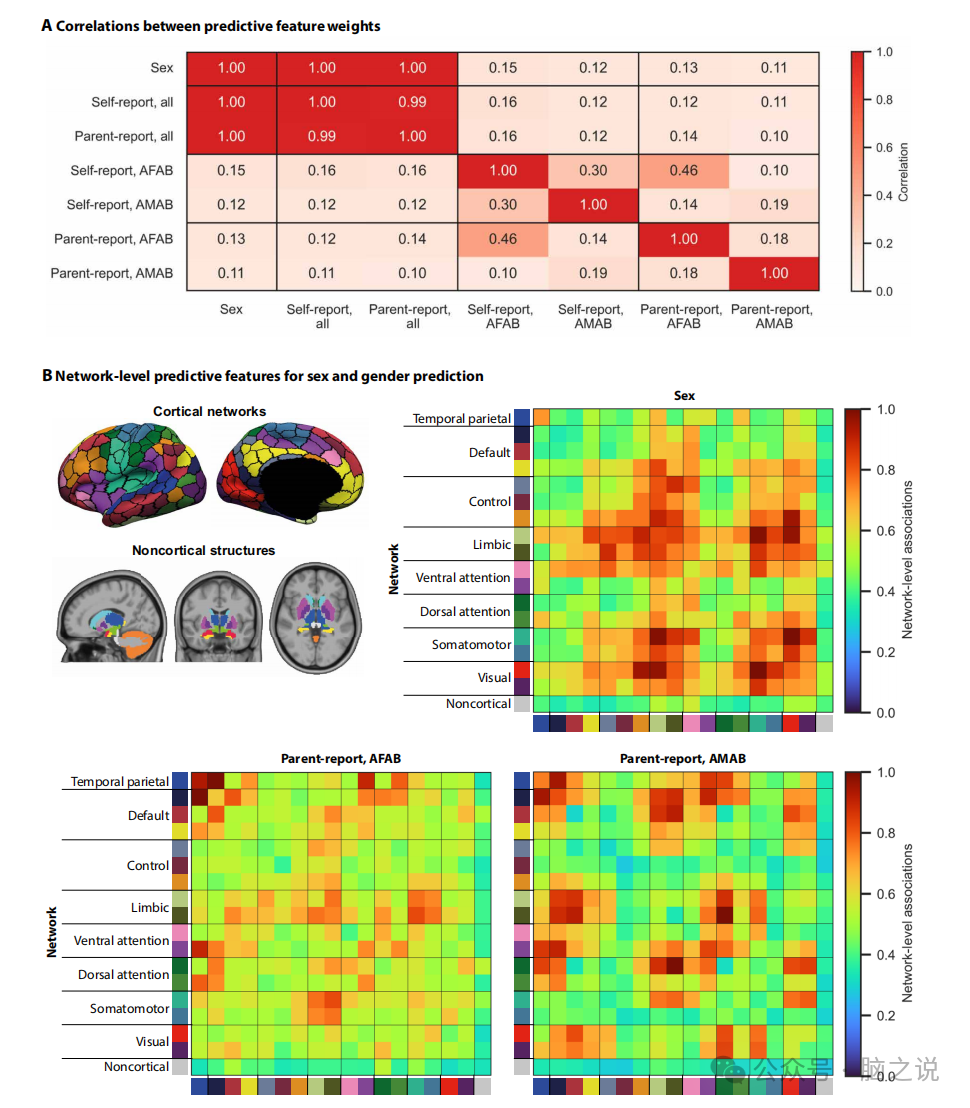
图2 不同的功能网络与指定的性和性别有关
3. 讨论
性别和生物学和行为的性别差异与整个生命周期的健康结果有关。了解性和性别的神经生物学基础对于随后确定性和性别如何影响健康和疾病以及开发性别特异性和以性别为导向的诊断和预后工具至关重要。在这里,我们证明了功能连接与性别和父母报告的性别有关。
我们对性别的预测(超越性别)远不如对性别或单独的性别预测准确,这表明性别可能是一个更复杂的结构,在功能连接模式中没有得到明确表示。性别是一个多维结构,包括个人的内在身份及其外部互动和行为,这两者都极难量化。至关重要的是,这里使用的自我报告测量评估了性别一致和性别不一致的感觉性别,以及性别表达和性别满足度,而父母报告测量则评估了玩耍过程中的性别一致和性别不一致的行为以及青少年的性别焦虑。因此,他们试图捕捉个人性别的不同方面。在这里,我们观察到相对于父母报告的分数,自我报告的分数存在有限的总体水平变异性。然而,这两个指标的相关性显著,尽管相关性很弱。我们无法捕捉自我报告的性别和功能连接之间有意义的关联可能是由于这些测量的有限可变性和/或由于自我和父母报告的测量中捕捉的性别维度的固有差异。但是,其他几个因素也可能促成这些结果。性别和性别在此数据集中高度相关;因此,反映性别效应(与性别无关)的功能连接性差异比例相当小也就不足为奇了。另一种解释是,性别对大脑的影响在个体之间更稳定,而性别的影响则更可变。性别认同和表达是流动的,这种流动性在非二元和跨性别个体中可能尤其明显。性别的这种动态性质可能反映在网络相关性中,因此很难使用横断面预测建模方法进行捕获。尽管如此,我们检测到功能连接与父母报告的性别之间存在显着关联,这表明性别确实会影响儿童大脑网络的组织。
在青春期,功能性脑网络在皮层网络和性别中不均匀地成熟。负责对一种感觉模态中的刺激做出反应的单峰感觉网络首先成熟,其次是参与高阶认知和社会过程的异峰关联网络。在这里,我们发现单峰和异峰网络都参与了性别和性别预测,这表明性和性别对大脑的影响是广泛的。此外,一组独特的功能连接与性别相关(在考虑性别之后)。性别与躯体运动、视觉、控制和边缘网络内部/之间的连接性更密切相关,而性别的网络相关性在整个皮层中更加分散。功能连接和性别之间的关联在性别之间有些重叠,但也表现出关键差异。在 AFAB 儿童中,在颞顶叶和注意力网络中观察到最强的关联,而在 AMAB 儿童中,我们看到涉及多个异峰网络以及视觉和躯体运动网络的更分散的关联模式。总之,这些发现表明性别的功能相关性与性别的功能相关性不同,并且构成性别的独特多维结构与 AFAB 和 AMAB 儿童的功能连接模式存在差异相关性。因此,性和性别必须同时研究,以充分捕捉男性和女性之间、男孩和女孩之间以及其他性别之间存在的差异和相似之处。
这些发现受到几个限制。首先,性不是二元的。然而,在此处分析的 ABCD 样本中,所有参与者都报告了他们的性别为女性或男性。因此,我们只考虑了二元性别的神经相关性以及 AFAB 和 AMAB 儿童的性别相关性。在性别和性别更加多样化的样本中进行的额外分析可能会揭示进一步的见解。其次,我们将性别视为一个连续体,而不是一个二元变量,以代表其真实性质并防止丢失与性别个体差异有关的宝贵信息。然而,自我报告的性别数据的变异性有限。这可能在一定程度上限制了我们捕捉功能连接和性别之间准确关系的能力,以及解开性别和性别的网络相关性的能力。在表现出更大性别不一致的个体中富集的数据集中的后续分析可能具有更高的检测这些关系的能力。与此相关,对具有不同性别一致性水平的个体亚群的分析可以揭示性别的功能网络相关性在人群中保持一致的程度。第三,这些分析是在相对年轻的队列中进行的。随着这些孩子进入青春期,他们对性别的理解和表达可能会发生明显变化。这种变化将与大脑结构和功能的成熟并行。因此,考虑大脑成熟和青春期之间的相互作用如何影响性别的个体差异将至关重要。未来的分析应寻求评估这些功能性网络表示的性别和性别在青春期和从童年到青春期的整个过渡期以及整个成年期如何变化。第四,使用一组假设静态形式的性别认同和表达的问题在单个时间点评估性别。未来跨多个时间点的工作可以改用允许在一系列时间尺度上评估性别流动性的问题。第五,性别受当地文化规范和共同社会经验的影响。ABCD 数据集完全在美国收集,不能代表全球人口。随后的分析应调查其他国家是否存在类似的关系。最后,我们使用了与之前工作一致的全脑方法。然而,表现出性别差异并与复杂行为有关的边缘网络容易出现信号丢失,并且比其他网络具有较低的重测可靠性。尽管我们实施了严格的质量控制措施,但我们不能排除整个大脑信噪比的固有差异不会影响我们的预测和/或解释,这应该在未来的分析中进行评估。
我们的分析揭示了儿童性别和性别的潜在神经生物学相关性,表明复杂而微妙的生物和环境因素共同影响大脑组织。然而,这些结果并不能为性别本质主义提供证据,我们强烈警告不要对这项工作进行任何简化或(错误)解释以提出相反的建议。我们建议读者查阅我们的补充文本以获取更多信息。
如果我们要了解性别和性别多样化样本中的健康和疾病,就必须全面了解性和性别的神经生物学相关性。在这里,我们确定了发育中人脑中指定性别和性别表达的不同功能网络相关性。这些相关性在发育和成年过程中如何维持或改变,以及它们如何与任何年龄的性别流动经历相关联,仍有待确定。
4. 材料和方法
4.1 数据
ABCD 数据集是一个基于社区的儿童和青少年的大型样本,他们接受了一套全面的神经影像学、行为学、发育学和精神病学组合的评估。在这里,我们使用了在基线时间点获取的经过最低限度预处理的神经影像学数据,以及来自美国国家心理健康研究所(NIMH) ABCD 版本 2.0.1 数据档案的1年随访时间点的自我和父母报告的性别数据。使用 GE 和 Siemens 扫描仪的协调协议在美国的 21 个地点采集了磁共振 (MR) 图像。与我们之前的工作一致,我们使用排除标准来确保质量控制。对于 T1 数据,我们删除了未通过侦察所有质量控制的个体。对于功能连接数据,我们排除了基于边界的注册成本大于 0.6 的功能运行。此外,我们用帧位移毫米或体素差分信号方差体积,以及之前的一个体积和之后的两个体积。我们还删除了包含少于 5 个连续卷的未删数据片段。我们删除了超过一半体积被删失和/或最大的功能性运行。我们还排除了没有至少 4 分钟数据的人。根据 ABCD 联盟的建议,我们排除了由于预处理不正确而使用 Philips 扫描仪扫描的个体。我们还排除了没有所有行为(例如性别)数据或彼此相关的个体,以防止由于神经生物学和/或行为测量的固有遗传性而导致的意外偏倚。最后,我们从少于 50 人的网站中删除了数据。我们的最终样本包括来自 ABCD 2.0.1 版本的 4757 名儿童。数据集的研究方案由加州大学圣地亚哥分校的中央机构审查委员会审查和批准,在某些情况下,由各个站点 IRB 审查和批准。父母或监护人提供了书面知情同意书,儿童在参与前表示同意。
4.2 性和性别数据
我们纳入了出生时分配的性别和来自青少年自我报告和父母报告性别问卷的性别数据 。这些分析中包含的所有参与者都完成了青年性别调查,其中包括四个问题,用于衡量感受性别、性别表达和性别满意度。此外,他们的父母/看护人完成了一份改编的性别认同问卷 ,其中包括 12 个问题,用于测量游戏期间的性别类型行为和性别焦虑。我们通过分别计算每个问卷中所有问题的总和来计算所有参与者的自我报告和父母报告的性别分数,并将这些总结分数用于我们的分析。我们使用非参数 Mann-Whitney U 秩检验来评估性别评分的性别差异。我们使用 Benjamini-Hochberg 错误发现率程序校正了多重比较的所有 P 值。我们还计算了每个指定性别的性别分数之间的非参数相关性,以评估可能存在的任何潜在关系。
4.3 图像采集和处理
我们如前所述处理了最低限度预处理的 MRI 数据。简而言之,我们使用 FreeSurfer 5.3.0进一步处理了经过最低限度预处理的 T1 数据,为每个个体生成皮质表面网格,然后将其注册到一个通用球坐标系。我们还通过以下步骤处理了最低限度预处理的功能性磁共振成像 (fMRI) 数据:(i) 删除初始帧,删除的帧数取决于扫描仪的类型,以及 (ii) 使用 BBR与 FsFast 与 T1 图像对齐。在计算 FD之前,我们使用带阻滤波器过滤掉呼吸假性运动。我们还从 fMRI 时间序列中回归了总共 18 个干扰协变量:全局信号、6 个运动校正参数、平均心室信号、平均白质信号及其时间导数。我们估计了未删失卷的回归系数。由于我们对行为预测感兴趣,因此我们进行了全局信号回归,并且全局信号回归已被证明可以提高行为预测性能。最后,我们使用最小二乘频谱估计跨删失帧插值脑部扫描,应用带通滤波,将它们投影到 FreeSurfer fsaverage6 表面空间,并使用 6 毫米全宽半高内核对其进行平滑。处理后,我们提取了 400皮层和19个非皮层包裹的区域时间序列。我们计算了这些时间序列之间的完全相关性,产生了 419 × 419 成对区域功能连接矩阵。如前所述,所有处理均在本地服务器上完成。
4.4 预测建模
线性岭回归模型避免了过度拟合,易于解释,并且与基于大脑的行为预测的深度学习算法相比,计算成本相对低廉。在这里,使用与我们研究团队以前使用的框架类似的框架,我们进行分析以确定指定性别和性别的大脑功能性相关性。我们使用交叉验证的脊回归模型来根据功能连接预测性别和性别。为了便于比较性别和性别预测之间的模型性能和特征贡献,我们实施了线性岭回归框架进行性别预测,而不是分类模型。在这里,我们在机器学习上下文中使用术语 “prediction” 来指代估计个人性别或性别的算法的输出。有关我们使用此术语的进一步讨论,我们建议读者参考我们的补充文本。对于预测性别的模型,我们包括所有个体(AMAB 和 AFAB),而对于预测性别的模型,我们要么包括所有个体,要么使用性别特异性方法(即,针对每种性别分别训练和测试)。对于包括所有个体的性别预测模型,我们颠倒了 AFAB 儿童的性别分数,使得它们的自我报告分数从 0 到 20(0 表示更女性化的性别认同和表达,20 表示更男性化的性别认同和表达)和 0 到 60 的父母报告分数(0 表示更女性化的性别认同和表达,60 表示更男性化的性别认同和表达)。对于每个模型,我们将数据分成 100 个不同的训练集和测试集(比例约为 4:1),无需替换。在拆分数据时,我们考虑了对站点进行成像,以便我们将来自给定站点的所有参与者放在训练集或测试集中,而不是在两者之间进行拆分。在每个训练集中,我们使用三重交叉验证优化了正则化参数,同时与初始训练测试拆分类似地考虑了成像位点。优化后,我们在相应的测试集上评估了模型。我们对 100个不同的训练测试拆分中的每一个重复此过程,以获得预测准确性和解释方差的分布。为了评估模型的重要性,对于每组预测模型,我们生成了一组相应的空模型,如下所示:输出变量被随机排列 1000 次,每次排列都用于使用从原始模型中的选定参数集中随机选择的正则化参数来训练和测试空模型。然后,我们将每个 null 模型的预测准确性与原始 (真实) 模型的相应模型准确性分布的平均准确性进行了比较。每个模型显著性的 P 值定义为预测精度或解释方差大于或等于对应于原始分布的空模型的比例。
4.5 特征权重
我们使用 Haufe 变换来转换从线性岭回归模型获得的特征权重,以提高其可解释性和可靠性。对于每个训练拆分,我们使用了从模型获得的特征权重 W、输入数据的协方差(函数连接性)Σx和输出数据的协方差(行为得分)Σy来计算 Haufe 变换的特征权重 A。然后,我们对 100 次拆分的绝对 Haufe 转换特征权重进行平均,以获得平均特征重要性值。我们计算了从不同模型获得的平均特征重要性之间的完全相关性,以评估它们是否依赖共享或独特特征来预测性别/性别。对于所有模型,我们还总结了网络级别的成对区域特征重要性,以支持前面描述的可解释性。为方便起见,将皮层下、脑干和小脑包裹分配给单个非皮层网络。然后,我们对区域成对绝对特征权重进行平均,以产生功能连接与性别/性别之间关联的网络级估计。最后,我们计算了这些网络级估计之间的完全相关性,以评估性别和性别的网络相关性共享的程度。
参考文献:Functional brain networks are associated with both sex and gender in children.

















 1180
1180

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









