










Establishing Komagataella phaffii as a Cell Factory for Efficient Production of Cholesterol Sulfate
将 Komagataella phaffii 建立为高效生产胆固醇硫酸的细胞工厂

摘要
胆固醇硫酸(CS)是一种胆固醇的硫酸化衍生物,由于其广泛的生物学和药理学活性,在医学领域具有重要应用。目前,CS 的供应主要依赖于从羊毛脂提取胆固醇后进行化学转化,这促使人们对通过工程化微生物细胞工厂进行大规模生产 CS 产生了浓厚兴趣。本研究首次报道了在酵母 Komagataella phaffii(也称为 Pichia pastoris)中实现 CS 的微生物生产。
通过组合代谢工程策略,包括阻断竞争分支、过表达限速的甲瓦龙酸途径基因、调控脂质代谢以及增强关键辅因子 3′-磷酸腺苷-5′-磷酸硫酸(PAPS)的供应,与初始菌株相比,CS 的产量提高了 6.8 倍以上。优化后的菌株(CHS0505)在摇瓶培养中达到了 249 mg/L 的 CS 产量,而在补料分批发酵中产量达到了 545 mg/L,这突显了 K. phaffii 作为类固醇生产细胞工厂的潜力。
更重要的是,CS 生产细胞工厂的开发将通过合成生物学促进糖胺聚糖和其他硫酸化天然产物的微生物生产。
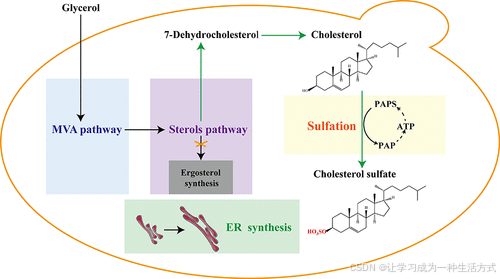
引言
我们的研究不仅确立了 K. phaffii 作为类固醇生产的有前景微生物底盘,还将推动糖胺聚糖及其他硫酸化天然产物的可持续生产。
类固醇广泛分布于自然界中,并在生长、生殖、代谢调节以及内分泌稳态中发挥重要作用。(1) 类固醇药物的工业合成发展迅速,已成为继抗生素之后的第二大药物类别,全球市场规模超过 100 亿美元。(2) 胆固醇是一种重要的类固醇,是合成大多数类固醇的重要原料和中间体。(3) 胆固醇硫酸(CS)是一种胆固醇的硫酸化衍生物,广泛分布于各种组织和体液中。(4) 它在人类身体中发挥着重要的生理作用,例如调节表皮脱落和屏障功能,调节脑代谢,发挥神经保护作用,并参与 T 细胞受体库的形成。(5−9) 目前,CS 的供应依赖于从羊毛脂中提取胆固醇的化学转化,这种方法高度依赖有限的自然资源,并引发
对有毒有机溶剂的使用和多种副产物的产生的担忧。这不仅导致大量废水的生成,还导致生产成本高昂。因此,迫切需要开发绿色细胞工厂,从简单的碳源(如葡萄糖和甘油)中从头合成CS。
酵母中存在内源性麦角固醇生物合成途径,使其成为通过失活与晚期相关的ERG 基因并引入异源途径基因实现类固醇化合物从头合成的理想底盘。(10) 例如,酿酒酵母(Saccharomyces cerevisiae)已被广泛用于从头合成类固醇,包括7-脱氢胆固醇 (7-DHC)、(11−15) 胆固醇、(16) 植物甾醇、(17−19) 薯蓣皂素、(20) 氢化可的松、(21) 和孕烯醇酮。(22) 此外,像Yarrowia lipolytica、Cyberlindnera jadinii 和 Komagataella phaffii (也称为 Pichia pastoris)等非常规酵母也被工程化以生产某些类固醇,如菜油甾醇、(23) 孕烯醇酮、(24) 和胆固醇。(25,26)
为了提高酵母中异源类固醇的生产,通常采用优化三个模块的策略,即中心碳代谢模块、甲瓦龙酸 (MVA) 模块和后甾醇模块。(27) 具体而言,通过破坏相关 ERG 基因(如 ERG4、ERG5 或 ERG6)减少竞争途径中的产品消耗,通过过表达 MVA 模块中的关键基因(如 ERG20、IDI 和 tHMG1)增加甾醇代谢流量,以及通过增强后甾醇模块(如 ERG1、ERG2 和 ERG11)加速甾醇向固醇的转化。(11,12,14,18,27) 调控转录因子(如 ROX1、MOT3 和 UPC2)和脂质代谢(如 INO2、INO4 和 PAH1)也被证明有助于提高酵母中甾醇的积累。(12,27−29) 当前,已经通过引入异源 C-7 甾醇还原酶基因(DHCR7 或 DWF5)和 C-24 甾醇还原酶基因 DHCR24,结合 ERG5 和 ERG6 基因的双敲除(Δerg5Δerg6),在酵母中实现了胆固醇的生产。(16,20,25,26) 而胆固醇可以进一步通过硫转移酶 SULT2B1b (30,31) 催化转化为 CS,目前尚未有微生物细胞工厂实现这一工程化。
Komagataella phaffii 是一种嗜甲醇酵母,被广泛用于生产各种重组蛋白。(32) 随着 CRISPR/Cas9 基因组编辑技术的快速发展,K. phaffii 已被用于生产各种增值天然产物,如长春胺、(33) 广藿香醇、(34) (+)-柠檬醛、(35) 黄红素、(36) α-烯烃、(37) α-檀香烯、(38) 龙胆二烯 A、(39) 和 α-双环烯。(40) 然而,有关工程化 K. phaffii 异源生产类固醇的报道仍然非常少。因此,值得尝试各种策略来探索 K. phaffii 的类固醇,特别是 CS 的高效过量生产潜力。
此外,硫化反应由多种硫转移酶催化,通过将磺酸基团(−SO3H)转移至羟基上。(41,42) 这是自然界中最重要的修饰反应之一。(43,44) 除了 CS,许多化合物(如多糖、(45) 黄酮类、(46) 神经递质、(47) 肽、(48) 次生代谢产物、(49) 和脂质 (50))都依赖于硫酸化修饰以发挥生物活性。3′-磷酸腺苷-5′-磷酸硫酸(PAPS)是所有生物硫酸化反应中唯一的活性磺酸基团供体。(51,52) 充足的 PAPS 供应是高效生物合成这些硫酸化化合物的前提条件。然而,硫酸化天然产物的酶促和微生物合成仍然受到限制,(45,46,53−56) 因为 PAPS 在体外价格昂贵且不稳定,并且在像大肠杆菌和酵母等微生物细胞中具有较差的细胞内可用性,这阻碍了工业规模化的生产。PAPS 是通过硫酸盐对 ATP 的两步激活过程合成的,由 ATP 硫化酶 (ATPS) 和 APS 激酶 (APSK) 催化。(57−59) 然而,PAPS 在体内的合成调控非常复杂,包括硫酸盐的运输、ATP 的合成与转化、PAPS 的消耗以及防止 PAPS 积累的负反馈机制。(60) 目前,尚无关于通过系统代谢工程增强体内 PAPS 合成以生产硫酸化化合物的报道。因此,迫切需要探索各种代谢工程策略,以增强 PAPS 的供应,从而实现高效微生物生产硫酸化天然产物。
本研究通过组合代谢工程策略对 K. phaffii 进行了改造,使其能够高效生产 CS。所采取的策略包括阻断竞争分支、过表达限速的 MVA 途径酶、调控脂质代谢以及增强 PAPS 的供应(见图 1)。在 K. phaffii 中建立的高产 CS 平台展现了微生物生产类固醇和含硫酸盐天然产物的巨大潜力。
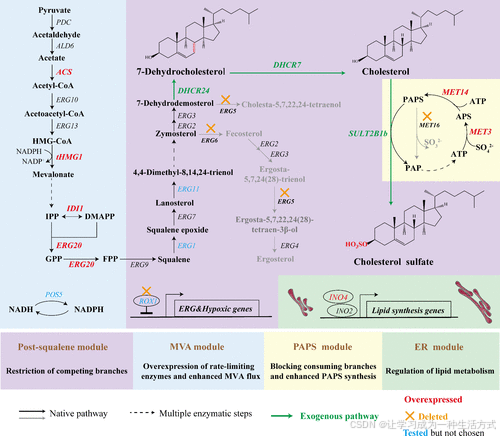
图1. 高水平CS生产中使用的模块化工程策略示意图 在K. phaffii中通过过表达DHCR24、DHCR7和SULT2B1b,以及敲除ERG5和ERG6(Δerg5Δerg6),重建了CS生物合成途径。模块化工程策略包括以下四部分:
-
过表达限速酶和增强MVA通量(MVA模块);
-
限制竞争分支(后鲨烯模块);
-
增强PAPS供应(PAPS模块);
-
调节脂质代谢(ER模块)。
黑色箭头表示天然代谢途径;虚线箭头表示多个酶促步骤;绿色箭头(或基因)表示外源途径(或基因);过表达的基因用红色高亮显示;敲除的基因用黄色高亮显示;蓝色基因表示这些基因曾被尝试过表达或敲除,但最终未被选中。
代谢物缩写:
-
HMG-CoA: (S)-3-羟基-3-甲基戊二酰辅酶A
-
IPP:异戊烯二磷酸
-
DMAPP:二甲基丙烯基二磷酸
-
GPP:香叶基二磷酸
-
FPP:法尼基二磷酸
基因缩写:
-
ACS:乙酰辅酶A合成酶
-
tHMG1:截短型HMG-CoA还原酶
-
IDI1:异戊烯基二磷酸异构酶
-
ERG20:法尼基焦磷酸合成酶
-
ERG1:鲨烯环氧化酶
-
ERG11:羊毛甾醇14-α-去甲基酶
-
ERG5:C-22甾醇脱氢酶
-
ERG6:Δ(24)-甾醇C-甲基转移酶
-
DHCR24:Δ(24)-甾醇还原酶
-
DHCR7:7-脱氢胆固醇还原酶
-
SULT2B1b:羟基甾体硫酸转移酶2
-
POS5:线粒体NADH激酶
-
ROX1:低氧基因抑制因子
-
MET3:ATP硫酸化酶
-
MET14:腺苷-5′-磷酸硫酸激酶
-
MET16:3′-磷酸腺苷硫酸还原酶
所用基因的完整列表见补充表S1。
方法与材料
菌株、培养基和试剂
-
Escherichia coli DH5α:用于基因克隆与质粒扩增,在Luria–Bertani培养基(10 g/L胰蛋白胨、5 g/L酵母提取物、10 g/L氯化钠)中培养,37 °C,补充100 μg/mL氨苄青霉素。
-
K. phaffii GS115-Cas9:用于基因组工程和生产7-DHC、胆固醇及CS,来源于先前研究。
-
S. cerevisiae Syvdb1和Syvdb2:工业二倍体菌株,用于7-DHC生产,来自浙江大学许志南教授。
-
酵母菌株培养在YPD培养基(10 g/L酵母提取物、20 g/L蛋白胨、20 g/L葡萄糖)或YPG培养基(10 g/L酵母提取物、20 g/L蛋白胨、20 g/L甘油),30 °C,220 rpm。必要时加入100 mg/L Zeocin、200 mg/L潮霉素B和/或200 mg/L G418抗性筛选剂。
所用试剂:
-
Gibson拼接试剂盒、T4 DNA连接酶及限制性内切酶来自NEB(美国Ipswich);
-
KOD Flash DNA聚合酶来自TOYOBO(日本);
-
质粒小提及DNA纯化试剂盒购自Thermo Fisher Scientific(美国Waltham);
-
标准品7-DHC与胆固醇购自阿拉丁生化科技公司(中国上海);
-
CS标准品购自熙雅化工有限公司(中国山东)。
除特别说明外,其余化学试剂均购自Sangon Biotech(中国上海)。
质粒构建
-
整合供体质粒:含有基因组整合同源臂以及异源基因表达的启动子和终止子,基于之前研究设计。
-
外源基因DHCR24(Gallus gallus来源)、DHCR7(来自G. gallus、Danio rerio、Homo sapiens或Rattus norvegicus)及SULT2B1b(来自H. sapiens)经密码子优化后由Generay Biotech合成(中国南京)。
-
内源基因(如MET3、MET14、ERG20、tHMG1等)通过PCR从酵母基因组DNA扩增。
-
突变基因ACS*(ACSL641P):编码不受反馈抑制的乙酰辅酶A合成酶突变体(来源于Salmonella enterica),由质粒ACS*Opt扩增。
-
其他候选基因(如CysC、TrxA等)通过PCR从E. coli或Pseudomonas aeruginosa基因组DNA扩增。
这些基因/片段通过酶切连接和/或Gibson拼接组装到相应的供体质粒中。基因、质粒及引物序列见补充表S1-S4,sgRNA序列和整合位点见补充表S5。
菌株构建
基于之前描述的CRISPR/Cas9系统(61),进行K. phaffii的基因敲除和定点整合。具体步骤包括:
-
途径基因表达盒与同源臂PCR扩增后,与sgRNA质粒共转化到Cas9表达菌株中;
-
通过电转化方法完成K. phaffii转化;
-
用菌落PCR验证克隆,并使用抗生素去除sgRNA质粒。
所有构建的菌株见表1。菌株构建流程示意图见补充图S1。
Table 1. Yeast Strains Used in This Study
| strain name | genotype/description | source |
|---|---|---|
| GS115-Cas9 | K. phaffii GS115-his4::HIS4-Cas9 | our lab |
| C01 | GS115-Cas9, Δerg5 | this study |
| C02 | C01, Δerg6 | this study |
| DHC01 | C02, Int12::GAPp-GgDHCR24-AOX1t | this study |
| CH01 | C02, Int12::(AOX1t-GgDHCR24-GAPp)-(TEF1p-DrDHCR7-0547t) | this study |
| CH02 | C02, Int12::(AOX1t-GgDHCR24-GAPp)-(TEF1p-GgDHCR7-0547t) | this study |
| CH03 | C02, Int12::(AOX1t-GgDHCR24-GAPp)-(TEF1p-HsDHCR7-0547t) | this study |
| CH04 | C02, Int12::(AOX1t-GgDHCR24-GAPp)-(TEF1p-RnDHCR7-0547t) | this study |
| CH05 | DHC09, Int21::TEF1p-DrDHCR7-0547t | this study |
| CH06 | CH05, Int1::ADH2p-GgDHCR24-AOX1t | this study |
| CHS01 | CH01, Int1::ADH2p-HsSULT2B1b-AOX1t | this study |
| CHS02 | CHS01, Int15::GAPp-tHMG1-AOX1t | this study |
| CHS03 | CH-S02, Int18::(AOX1t-ScERG20-GAPp)-(TEF1p-ScIDI1-PMP20t) | this study |
| CHS04 | CH-S03, Int36::TEF1p-SeACS*-AOX1t | this study |
| CHS05 | CH-S04, Δmet16 | this study |
| CHS06 | CH-S04, Int37::(AOX1t-KpSUL1-PDC1p)-(CDC19p-KpSUL2-0547t) | this study |
| CHS07 | CH-S04, Int21::(AOX1t-KpMET3-GAPp)-(TEF1p-KpMET14-0547t) | this study |
| CHS08 | CH-S04, Int21::TEF1p-KpMET3-(G4S)6-EcCysc-0547t | this study |
| CHS09 | CH-S04, Int21::(AOX1t-TrxA-PPK2N-GAPp)-(TEF1p-KpMET3-(G4S)6-EcCysc-0547t) | this study |
| CHS10 | CHS04, Int21::(AOX1t-ScMET3-GAPp)-(TEF1p-ScMET14-0547t) | this study |
| CHS11 | CHS04, Int21::TEF1p-ScMET3-(G4S)6-EcCysc-0547t | this study |
| CHS12 | CHS04, Int21::(AOX1t-PPK2N-GAPp)-(TEF1p-ScMET3-(G4S)6-EcCysc-0547t) | this study |
| CHS0501 | CHS05, Int21::(AOX1t-ScMET3-GAPp)-(TEF1p-ScMET14–0547t) | this study |
| CHS0502 | CHS05, Int21::(AOX1t-KpMET3-GAPp)-(TEF1p-KpMET14–0547t) | this study |
| CHS0503 | CHS05, Int37::(AOX1t-KpERG1-GAPp)-(TEF1p-KpERG11–0547t), Δrox1 | this study |
| CHS0504 | CHS0503, Int21::(AOX1t-KpMET3-GAPp)-(TEF1p-KpMET14–0547t) | this study |
| CHS0505 | CHS0502, Int32::AOX1t-ScINO4-GAPp | this study |
| CHS0506 | CHS0502, Int32::(AOX1t-ScINO4-GAPp)-(TEF1p-KpPOS5–0547t) | this study |
菌株培养
为了比较7-DHC的生产情况,将工业二倍体S. cerevisiae菌株(Syvdb1和Syvdb2)或K. phaffii菌株(DHC01)的单个菌落接种到4 mL的YPD培养基中,分别在30°C、220 rpm下培养12或24小时。随后,将约0.6 mL的预培养液接种到含30 mL YPD培养基的250 mL三角瓶中,在30°C、220 rpm下培养。培养36小时后,添加2%的额外葡萄糖。酵母细胞在发酵72小时后收获。用于生产7-DHC、胆固醇和CS的工程化K. phaffii菌株在YPG培养基中进行摇瓶发酵。具体步骤如下:将单个工程化酵母菌落接种到4 mL的YPD培养基中,在30°C、220 rpm下培养24–30小时。然后,将约0.6 mL的预培养液接种到250 mL三角瓶中的30 mL YPG培养基中,在30°C、220 rpm下培养36小时后,添加2%的额外甘油。发酵72小时后收获酵母细胞。所有发酵实验均以生物学三重复进行。
生物反应器发酵
将CHS0505的单个菌落接种到4 mL的YPD培养基中,培养24小时后,将1 mL(2%)种子液转移至三个各含50 mL新鲜YPD培养基的250 mL三角瓶中,在30°C、250 rpm下培养24小时。所得培养液(150 mL)转移至一个5 L生物反应器(T&J-A型,上海天骄生物工程有限公司)中,反应器内含2.5 L的YPG培养基(40 g/L甘油)。发酵过程在30°C和250–600 rpm下进行,气流速率为1–3 vvm,以保持溶解氧(DO)高于20%;通过自动添加8%的氨水和36%的乙酸将pH维持在5.5–5.8。当发酵罐内甘油浓度低于1 g/L时,连续补充甘油,补料液含500 g/L甘油和20 g/L酵母提取物。在特定时间间隔取样,检测OD600、甘油浓度、乙醇浓度和类固醇产量。
7-DHC和胆固醇的GC-MS定量
从酵母细胞中提取7-DHC和胆固醇,采用三种不同的提取方法:
-
方法A:取500 μL酵母培养液,用去离子水清洗两次,收集酵母细胞后,用1 mL 3 M HCl在100°C下处理5分钟,将细胞残渣用蒸馏水清洗至pH中性。随后加入500 μL 3 M KOH和500 μL甲醇,在80°C下皂化90分钟。冷却后,加入500 μL正己烷进行类固醇提取,震荡提取2小时。
-
方法B:与方法A相同的步骤处理酵母细胞,但加入500 μL甲醇和500 μL正己烷直接提取类固醇,震荡2小时。
-
方法C:取500 μL酵母培养液的细胞,悬浮于1 mL ddH₂O中,用自动样品研磨仪(TissueLyser 24,中国上海)以65 Hz破碎5分钟。离心去除上清液后,加入500 μL甲醇和500 μL正己烷震荡提取2小时。
每种方法所得的正己烷相通过气相色谱-质谱联用(GC-MS)分析。仪器为Agilent 8890-5977B,配备Agilent HP-5MS超惰性毛细管GC柱(30 m × 0.25 mm,膜厚0.25 μm)。离子源温度250°C,进样温度260°C,程序升温:80°C保持1分钟,然后以30°C/min升至250°C,再以10°C/min升至300°C并保持10分钟。7-DHC和胆固醇储备溶液用正己烷稀释至1、0.8、0.6、0.4、0.2和0.1 mg/L,用于生成标准曲线。
CS的LC-MS定量
从500 μL发酵液中的酵母细胞提取CS,将细胞重悬于1 mL甲醇中,并用自动样品研磨仪破碎。以12,000g离心10分钟后,将上清液转移至新管中,并用甲醇-水溶液(80/20,v/v,含0.1%甲酸)稀释。取100 μL稀释液于96孔板中,用Agilent RapidFire 4000高通量系统联用Agilent 6470三重四极杆质谱仪(RFMS/MS)分析。样品在甲醇-水溶液(10/90,v/v,含0.1%甲酸)中以1.5 mL/min流速加载至C18柱(Agilent),随后用乙腈-异丙醇溶液(80/20,v/v,含0.1%甲酸)以0.8 mL/min流速洗脱至质谱仪。通过正离子多反应监测(MRM)模式操作,CS的MRM转换设置为465.4/97和465.4/465.4。通过CS标准品的峰面积定量,并用标准曲线生成定量数据。
结果
K. phaffii中7-DHC和胆固醇生物合成途径的构建
通过敲除竞争分支基因(Δerg5Δerg6)和引入异源基因GgDHCR24,在酵母中实现了7-DHC的生物合成。二倍体工业S. cerevisiae Syvdb1和Syvdb2菌株分别携带两拷贝和四拷贝的GgDHCR24基因,生产7-DHC的浓度分别为3.3 ± 0.1 mg/L和19.8 ± 1.6 mg/L(YPD培养基,图2A)。相比之下,携带单拷贝GgDHCR24基因的K. phaffii DHC01菌株,在YPD培养基中生产7-DHC的浓度达35.6 ± 2.3 mg/L(图2A、2C),分别是Syvdb1和Syvdb2的10.6倍和1.8倍(P < 0.01)。此外,将7-DHC的产量标准化为生物量进一步比较(图S5)。
引入不同来源的DHCR7基因至DHC01,生成生产胆固醇的菌株。与前人在S. cerevisiae的研究一致(11),斑马鱼(D. rerio)的DHCR7表现出比鸡(G. gallus)、人类(H. sapiens)和大鼠(R. norvegicus)更高的催化效率,能将大部分7-DHC转化为胆固醇(图2B)。对应菌株CH01(DrDHCR7)*的胆固醇产量为108 ± 26.2 mg/L,显著高于*CH02(GgDHCR7, 6.6 ± 0.32 mg/L)、CH03(HsDHCR7, 6.8 ± 0.72 mg/L)*和*CH04(RnDHCR7, 6.2 ± 0.13 mg/L)(P < 0.01,图2B、2C)。此外,CH01中胆固醇的产量是DHC01中7-DHC产量的约三倍(108 vs 36.1 mg/L),表明高效的DrDHCR7显著提高了代谢通量向类固醇合成的转化。
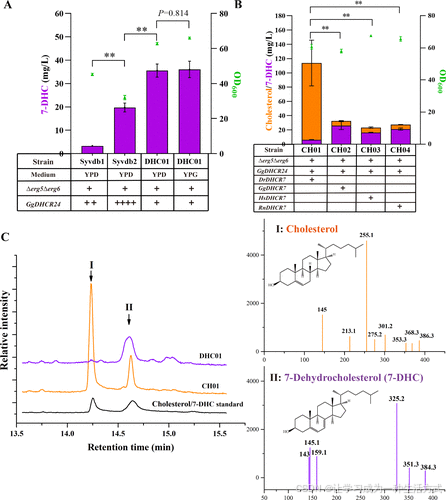
图2. 在K. phaffii中构建7-DHC和胆固醇的de novo合成途径 (A) S. cerevisiae和K. phaffii中7-DHC产量的比较。通过敲除ERG5和ERG6(Δerg5Δerg6)*并过表达不同拷贝数的*GgDHCR24,构建了重组酵母菌株。这些菌株的7-DHC产量在YPD或YPG培养基中进行评估。 (B) 在YPG培养基中筛选用于胆固醇生产的最佳DHCR7。 (C) 工程化7-DHC生产菌株和胆固醇生产菌株培养液提取物的GC-MS分析图谱。峰I和峰II分别对应胆固醇和7-DHC。标准品(7-DHC和胆固醇)和样品(DHC01和CH01)的GC-MS图谱详见补充图S4。样品提取采用方法A。未探索DHC01或CH01菌株可能生产的其他甾醇。平均值±标准差来自三次生物重复。差异显著性通过单因素方差分析(*P < 0.01,P < 0.05)评估。
三种提取方法(A、B和C)的比较 通过方法B提取DHC01样品的7-DHC产量(57.2 ± 0.1 mg/L)显著高于方法A(36.1 ± 2.9 mg/L)和方法C(44.6 ± 2.2 mg/L)(P < 0.01)。CH01样品的胆固醇产量通过方法B提取(141 ± 11.3 mg/L)略高于方法A(108 ± 26.2 mg/L)和方法C(97.9 ± 17.1 mg/L)(P分别为0.067和0.134)(图S6)。这些结果表明,方法B更适合用于提取K. phaffii中的7-DHC和胆固醇。
CS生物合成途径的构建及MVA关键基因的过表达以促进CS合成
为了构建CS生产菌株,将来自H. sapiens的羟基甾体硫酸转移酶基因SULT2B1b引入胆固醇生产菌株CH01中,所得菌株CHS01生产36.6 ± 4.5 mg/L CS和77.7 ± 6.5 mg/L胆固醇(图3A)。通过LC-MS分析确认该新化合物(峰III)为CS(图3B和补充图S7)。
为提高CS产量,需要增强CS合成途径的代谢通量。在后鲨烯模块中,该通量主要依赖于MVA途径,ACS1、tHMG1(截短型HMGR)、ERG20和IDI1是主要的限速酶。(38) 在MVA途径中过表达tHMG1使得CHS02生产的CS浓度达到87.0 ± 10.5 mg/L,比母菌株CHS01(36.6 ± 4.5 mg/L)高出2.4倍(P < 0.01)(图3A)。
接下来,将ScERG20、ScIDI1和SeACS*(反馈不敏感的ACS1突变体)的表达盒依次整合到K. phaffii基因组中。所得菌株CHS03和CHS04的CS产量分别为81.9 ± 5.2 mg/L和84.8 ± 2.5 mg/L,与CHS02相似(图3A)。在MVA模块中,过表达tHMG1显著提高了CS水平,但进一步过表达ScERG20、ScIDI1和SeACS*未能进一步提高CS产量,表明其他因素(如PAPS供应)可能限制了CS的生物合成。
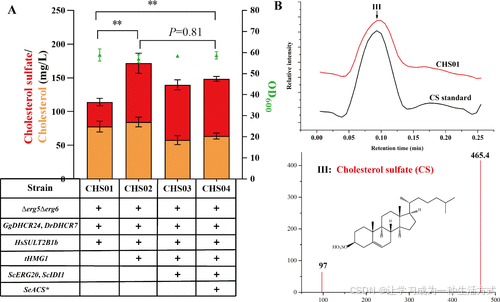
图3. 在K. phaffii中构建CS生物合成途径并通过过表达MVA途径关键基因提高CS水平 (A) 通过敲除ERG5和ERG6,引入GgDHCR24、DrDHCR7和HsSULT2B1b,构建了CS生产起始菌株。随后在MVA途径中过表达限速基因tHMG1、ScERG20、ScIDI1和SeACS*以提高CS合成。 (B) 工程化CS生产菌株培养液提取物的LC-MS分析图谱,峰III对应CS。CS标准品和样品CHS01的LC-MS图谱详见补充图S7。所有数据为三次生物重复的均值±标准差,显著性差异通过单因素方差分析评估。*P < 0.05,**P < 0.01。
增强PAPS供应以促进CS合成
胆固醇向CS的硫酸化需要PAPS作为辅因子(硫酸化供体),其通过ATPS和APSK催化的两步反应由无机硫酸与ATP合成(图1)。然而,内源途径中还包括PAPS还原酶,其与硫酸转移酶竞争并将PAPS还原为无机亚硫酸盐。(54,60) 为提高PAPS供应,探索了多种代谢工程策略,包括:
-
敲除内源PAPS还原酶基因(Δmet16);
-
过表达KpSULT1和KpSULT2以增强无机硫酸的摄取;
-
过表达来自S. cerevisiae(ScMET3和ScMET14)或K. phaffii(KpMET3和KpMET14)的ATPS和APSK;
-
过表达融合了ATPS(ScMET3或KpMET3)和来自E. coli(EcCysC)的APSK的双功能ASAKS5;
-
过表达ATP再生酶PPK2N(PPK2CpaN)。
通过策略1(Δmet16,见补充图S2和S3)敲除内源PAPS还原酶基因可轻微提高CS水平,所得菌株CHS05生产100 ± 13.2 mg/L CS,相比母株CHS04(84.8 ± 2.5 mg/L CS)提高了1.18倍,但差异不显著(P = 0.254)。
相较之下,单独过表达来自S. cerevisiae或K. phaffii的ATPS和APSK(策略3)未能提高CS产量,菌株CHS07和CHS10分别生产83 ± 1.2和87.6 ± 2.4 mg/L CS,与母株CHS04(84.8 ± 2.5 mg/L)相似(P = 0.901和P = 0.850)。
然而,将策略1(Δmet16)与策略3(过表达ATPS和APSK)结合显著提高了CS水平。相应菌株CHS0501和CHS0502分别生产148 ± 11.3和157 ± 15.7 mg/L CS,较CHS04(84.8 ± 2.5 mg/L)分别提高了1.74倍和1.85倍(P < 0.01)。此外,CHS0501和CHS0502的硫酸化程度分别达到了71.2%和68.1%,显著高于CHS04(52.4%)(P < 0.01)(图4)。
遗憾的是,策略2、4和5均未能提高CS产量。相应菌株CHS06(策略2)、CHS08和CHS11(策略4)*以及*CHS09和CHS12(策略4结合策略5)*分别生产51.5 ± 6.5、23.4 ± 11.7、39.3 ± 11.5、67.6 ± 3.7和47.1 ± 17.1 mg/L CS,均低于母株*CHS04(84.8 ± 2.5 mg/L)(图4)。值得注意的是,单独在菌株CHS11中过表达ScMET3-(G4S)6-EcCysC(策略4)使CS产量下降至39.3 ± 11.5 mg/L,但胆固醇水平达到峰值95.1 ± 3.8 mg/L(图4)。
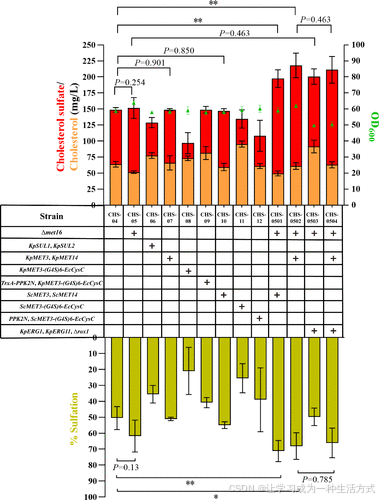
图4. 探索PAPS供应和后鲨烯途径对提高K. phaffii中CS合成的影响 通过不同的代谢工程策略提高PAPS积累,包括:(1) 消除PAPS消耗途径(Δmet16);(2) 增强无机硫酸的摄取(过表达KpSULT1和KpSULT2);(3) 过表达来自S. cerevisiae(ScMET3和ScMET14)和K. phaffii(KpMET3和KpMET14)的ATPS和APSK;(4) 过表达融合了ATPS(ScMET3或KpMET3)和来自E. coli的APSK(EcCysC)的双功能ASAKS5;(5) 过表达ATP再生酶PPK2N(PPK2CpaN)。此外,还过表达了后鲨烯途径关键基因ERG1和ERG11,并敲除了低氧基因转录抑制因子ROX1,以研究它们对CS水平的影响。硫酸化程度根据CS/(CS+胆固醇)的摩尔比计算。所有数据为三次生物重复的均值±标准差,显著性差异通过单因素方差分析评估。*P < 0.05,**P < 0.01。
重调节羊毛甾醇合成途径以提高CS合成
如前人在S. cerevisiae中的研究所示,过表达后鲨烯模块中的ERG1和ERG11可提高7-DHC产量。(12,28,63) 此外,敲除ROX1(血红素依赖的低氧基因抑制因子)可增加多个ERG基因的表达,从而提高甾醇和倍半萜的水平。(12,63,64) 基于此,在菌株CHS05中过表达内源基因ERG1和ERG11,并敲除ROX1,所得菌株CHS0503生产109 ± 9.9 mg/L CS,略高于母株CHS05(100 ± 13.2 mg/L CS)。此外,CHS0503的胆固醇产量比母株CHS05高出78%(91.4 ± 8.1 vs 51.4 ± 1.6 mg/L)。
随后,这些策略进一步应用于性能最佳的菌株CHS0502(Δmet16,过表达KpMET3和KpMET14)。然而,所得菌株CHS0504生产148 ± 17.0 mg/L CS,与母株CHS0502(157 ± 15.7 mg/L)相似(图4)。
鉴于多种羊毛甾醇合成酶定位于内质网(ER),(65) 通过扩展ER可提高萜类化合物、甾体化合物(12,28) 和分泌蛋白的产量。(66,67) 酵母中的INO2和INO4是磷脂生物合成的转录因子编码基因,对ER扩展起关键作用。在K. phaffii中,与INO2相比,过表达INO4能更高效地扩展ER并提高人乳白蛋白的产量。(67) 因此,在菌株CHS0502中过表达INO4构建了菌株CHS0505,其CS产量达到249 ± 8.4 mg/L,是菌株CHS0502(157 ± 15.7 mg/L)的约1.6倍(图5)。
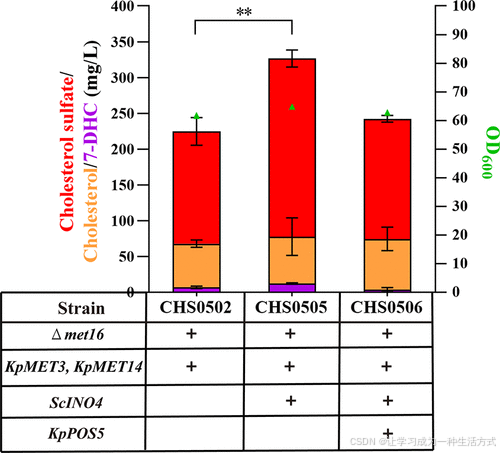
图5. 脂质调控和辅因子NADPH供应对K. phaffii中CS合成的影响 INO4是酵母磷脂生物合成中的转录因子,在扩展内质网(ER)的空间方面发挥关键作用。在羊毛甾醇合成途径中,许多酶(如tHMG1和细胞色素P450酶(CYPs))依赖于NADPH。所有数据均为三次生物重复的均值±标准差,显著性差异通过单因素方差分析评估。*P < 0.05,**P < 0.01。
此外,NADPH(烟酰胺腺嘌呤二核苷酸磷酸)是氧化还原酶的重要辅因子。在羊毛甾醇合成途径中,许多酶(如tHMG1和细胞色素P450酶(CYPs))都依赖NADPH,每合成一个羊毛甾醇分子需要消耗16个NADPH分子。(68) 此前的研究表明,过表达POS5在S. cerevisiae中显著提高了鲨烯的产量。(69) 因此,在CS产量最高的菌株CHS0505中整合了内源性POS5表达盒。然而,结果显示,所得菌株CHS0506的CS产量为168 ± 3.8 mg/L,低于菌株CHS0505(249 ± 8.4 mg/L)(图5)。
CS高产菌株的补料分批发酵
在对CS生物合成的四个模块进行优化后,最终的工程化菌株CHS0505在摇瓶中实现了249 ± 8.4 mg/L的CS产量,相比起始菌株CHS01提高了6.8倍(图6A)。
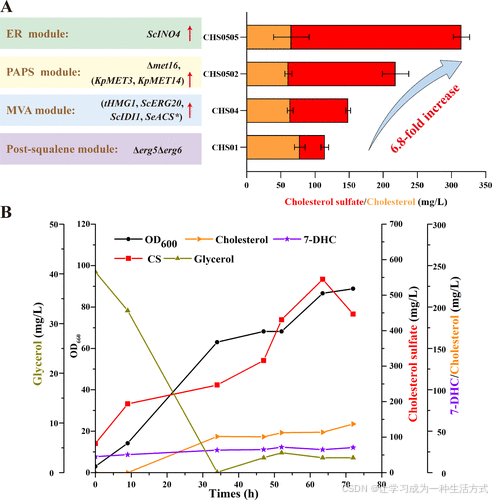
图6. 通过模块化路径优化和补料分批发酵提高CS产量 (A) 使用四步模块化工程策略构建的工程酵母菌株中的CS产量。 (B) 在CHS0505菌株中通过补料分批发酵实现CS的生产。
qPCR 确认了菌株CHS0505中关键基因(包括GgDHCR24、DrDHCR7、HsSULT2B1b、tHMGR、KpMET3、KpMET14 和 ScINO4)的表达水平(补充图S8)。此外,与野生型K. phaffii菌株相比,工程化CHS0505菌株表现出显著更长的迟滞期和较低的细胞密度(补充图S9)。由于PAPS生物合成的第一步涉及无机硫酸与ATP的活化,因此研究了硫酸补充对CS合成的影响。将两种硫酸形式(硫酸钠Na₂SO₄和硫酸铵(NH₄)₂SO₄)分别补充到在YPG培养基中培养的最佳菌株CHS0505中。结果表明,补充2 mM硫酸钠对CS产量仅有轻微促进作用(P = 0.764),但在高浓度(10 mM Na₂SO₄)下,额外的硫酸并未进一步提高CS产量(补充图S10)。这一结果与之前的研究一致,表明添加硫酸镁对E. coli中苄基葡糖苷酸盐水平没有显著影响。(70) 然而,补充硫酸铵(NH₄)₂SO₄对CS合成产生了负面影响(补充图S10和S11)。
因此,在2.5 L YPG培养基中,不补充额外硫酸的情况下,最佳菌株CHS0505被用于5 L发酵罐中的补料分批发酵。发酵过程中,在初始甘油(40 g/L)消耗完后,采用碳源限制策略,通过调节供料速率将甘油浓度维持在0.1–3 g/L的低水平(图6B)。经过72小时发酵,酵母细胞密度(OD600)达到88.8。在总计450 g甘油的连续供给下,乙醇未积累(<1 g/L),发酵罐中CS和总类固醇(7-DHC、胆固醇和CS)的产量分别在63.5小时后达到最大值545 mg/L和621 mg/L(图6B)。
讨论
类固醇在制药领域具有高度价值。然而,传统的类固醇制备方法(如植物或动物提取及化学合成)存在产量低、成本高的问题。近年来,利用微生物细胞工厂进行类固醇化合物的大规模生产受到越来越多的关注。
目前,Y. lipolytica和K. phaffii等非传统酵母已被工程化用于探索类固醇生产的潜力。在7-DHC的生产中,工程化K. phaffii菌株(Δerg5Δerg6,携带GgDHCR24)在摇瓶中达到35.6 mg/L的产量,优于工业二倍体S. cerevisiae菌株(Δerg5Δerg6,携带四拷贝GgDHCR24),其产量为19.8 mg/L。在胆固醇的生产中,K. phaffii菌株(Δerg5Δerg6,携带GgDHCR24和DrDHCR7)达到108 mg/L,高于S. cerevisiae(Δerg6Δatf2,携带DrDHCR24、DrDHCR7和三个ERG基因)生产的16 mg/L,以及C. jadinii(Δerg5Δerg6,携带GgDHCR24、StDWF5(DrDHCR7的同源基因,具有更高催化活性)和HMG1)生产的81.8 mg/L。(25,71) 本研究首次在K. phaffii中实现了高效CS的合成。通过组合代谢工程策略,CS产量在补料分批发酵中达到545 mg/L,凸显了K. phaffii作为类固醇生产细胞工厂的潜力。
尽管取得了这些进展,但CS产量仍低于g/L级,尚无法替代传统工业生产工艺。因此,需要进一步进行代谢和途径工程以优化K. phaffii中的CS合成。删除ERG6已被证明是提高酵母中胆固醇衍生类固醇化合物生产的成功策略。(11) 然而,这一改造导致了较长的迟滞期(补充图S9),这是由于ERG6的缺失导致膜中羊毛甾醇耗竭,从而损害了膜流动性,并增加了渗透和氧化应激的敏感性。(72) 因此,可以考虑通过CRISPRi系统维持基本的羊毛甾醇含量(12) 或通过启动子替换下调ERG6的表达(20),而不是完全敲除,以进一步促进菌株生长并提高CS产量。
此外,研究表明,过量胆固醇的合成会显著改变K. phaffii中膜双层的组成和结构,从而对细胞生理产生严重影响,包括异常生长、膜蛋白定位不当以及对多种试剂的敏感性增加。(73) 因此,在本研究中观察到的延长的迟滞期和降低的细胞密度可能是由脂毒性引起的(补充图S9)。为此,可以通过促进甾醇酯化途径、增强三酰基甘油(TAG)生物合成来扩大脂滴的大小,从而提高其储存甾醇的能力(13,17),以减少脂毒性并提高CS产量。
此外,导入甾醇细胞外运输基因以实现CS的分泌生产,可能是提高产量并简化下游纯化的有前景的策略,这在7-DHC(14) 和薯蓣皂素的生产中已有成功的应用。(20)
鉴于甾醇生物合成消耗大量能量并需要大量NADH和/或NADPH,通过辅因子工程重新平衡细胞内氧化还原状态对提高CS产量至关重要。(13,15) 进一步优化策略还可包括整合关键基因的多拷贝表达,以及在内质网、脂滴、过氧化物酶体和质膜等不同亚细胞区室之间重新分配酶的分布,以提高CS水平。(27)
本研究通过对PAPS(3'-磷酸腺苷-5'-磷酸硫酸盐)生物合成的探索和代谢工程策略的实施,显著提高了CS的产量。PAPS作为一种重要的中间代谢物,已被证明对其他硫酸化化合物(如软骨素硫酸、肝素硫酸、苄基葡糖苷酸盐和硫酸化酪氨酸)(46–80) 的微生物合成具有广泛的应用潜力。本研究中积累的见解不仅为生产CS提供了基础,也为其他硫酸化天然产物的微生物合成提供了指导。


















 861
861

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









