









De novo biosynthesis of plant lignans by synthetic yeast consortia
植物木脂素的 De novo 生物合成:合成酵母共生体系

酵母生产二氢槲皮素-文献精读117-CSDN博客
香叶醇酵母生产+机器学习优化酵母-文献精读66_ml-enhanced peroxisome capacity enables compartmen-CSDN博客 黄酮类化合物及其衍生物生物合成的进展:构建酵母细胞工厂的系统策略-文献精读50_dahp-CSDN博客
建立高效生产胆固醇硫酸的酵母细胞工厂-文献精读91_komagataella phaffii-CSDN博客
摘要
在酵母中重构木脂素等复杂天然产物的生物合成具有挑战性,可能导致代谢多效性(metabolic promiscuity),从而影响生物合成效率。在本研究中,我们通过合成酵母共生体系(synthetic yeast consortium)对木脂素的生物合成途径进行功能分工,并利用阿魏酸(ferulic acid)作为代谢桥接物。该协作体系成功克服了代谢多效性,并合成了木脂素的常见前体松柏醇(coniferyl alcohol)。
此外,通过系统工程策略的结合,我们实现了关键木脂素骨架紫杉松素(pinoresinol)和落叶松树脂醇(lariciresinol)的De novo 生物合成,并通过合成包括抗病毒木脂素二葡萄糖苷(lariciresinol diglucoside)在内的复杂木脂素,验证了该共生体系的可扩展性。
本研究成果为异源合成木脂素提供了一个工程化起点,并特别展示了强制互利共生的酵母共生体系(yeast consortium with obligate mutualism)作为一种有效策略,可以模拟植物细胞间的代谢分工,从而提升长途径和复杂天然产物的生物合成效率。
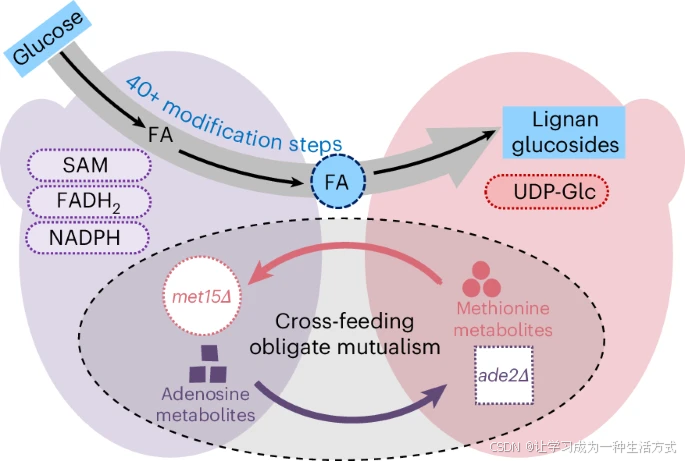
正文
由合成生物学驱动的微生物代谢工程是一种可行的方法,能够实现植物天然产物的可持续供应。然而,在微生物细胞工厂中异源重建功能性代谢途径仍然面临挑战。将生物合成途径过度表达到异源微生物中会引发代谢多效性、副反应和代谢负担等问题。这些不利因素会导致产量非常低,尤其是在合成长代谢途径的复杂分子时,如长春碱(vinblastine)、莨菪烷生物碱(tropane alkaloids)和阿片类药物(opioids)等。
在植物中,复杂的天然产物是通过不同细胞、组织、器官和细胞器之间的分工合作和代谢物运输来合成的。因此,构建类似植物细胞协作模式的微生物共生体系(microbial consortia)可能有助于合理扩展代谢能力并实现更复杂的合成。微生物共生体系是由两个或多个微生物种群(种间或种内)组成的系统,这种模式在自然界中普遍存在。
近期研究发现,特定的酵母营养缺陷型(如 ade8Δ、his3Δ 和 lys2Δ)可以形成人工共生体系,并可用于提高微生物产物的性能。例如,在提高白藜芦醇(resveratrol)合成方面已取得成功。此外,这种共生体系的“即插即用”(plug-and-play)模式使不同微生物表达长代谢途径的各个片段成为可能,从而显著减少遗传操作的时间成本,因为可以同时对所有微生物进行平行操作。 因此,构建微生物共生体系的工程技术进步对提高微生物细胞工厂的性能具有重要意义。
木脂素(lignans)是一类具有复杂结构和多样生物活性的天然产物,例如存在于板蓝根(Isatis indigotica)根部的抗病毒落叶松树脂醇葡萄糖苷(lariciresinol glucosides)。在植物中,所有木脂素(如鬼臼毒素(podophyllotoxin)、芝麻素(sesamin)、斯蒂加辛(steganacin)和落叶松树脂醇二葡萄糖苷(LDG))的合成均起始于两分子松柏醇(coniferyl alcohol)的电子偶联反应。松柏醇来源于芳香酸,与莽草酸途径(shikimate pathway)和苯丙氨酸途径(phenylalanine pathway)密切相关。
随后,不同的下游特异性酶(如漆酶(Lac)、紫杉松素-落叶松树脂醇还原酶(Plr)、细胞色素P450单加氧酶(Cyp)和尿苷二磷酸糖基转移酶(Ugt))进一步合成多样化的木脂素结构。因此,在微生物中合成木脂素的常见前体——松柏醇,是生产复杂木脂素前的优先任务。
植物中的木脂素合成涉及细胞间和细胞器间的协作合成网络,从而减少副反应的发生。在异源微生物(如酿酒酵母(Saccharomyces cerevisiae))中,防止代谢多效性以实现木脂素的De novo合成非常具有挑战性。尤其是对香豆酸(p-coumaric acid)、咖啡酸(caffeic acid)和阿魏酸(ferulic acid)等连续中间体具有相似结构,易通过非特异性还原途径转化为相应副产物,从而削弱指向松柏醇的关键代谢通量。
受益于不断积累的证据表明,在特定条件下酵母细胞群体可以形成种内共生体系,我们假设酵母共生体系能够模仿植物细胞的分工模式,以防止副反应并在酿酒酵母中实现木脂素的De novo合成(图1)。
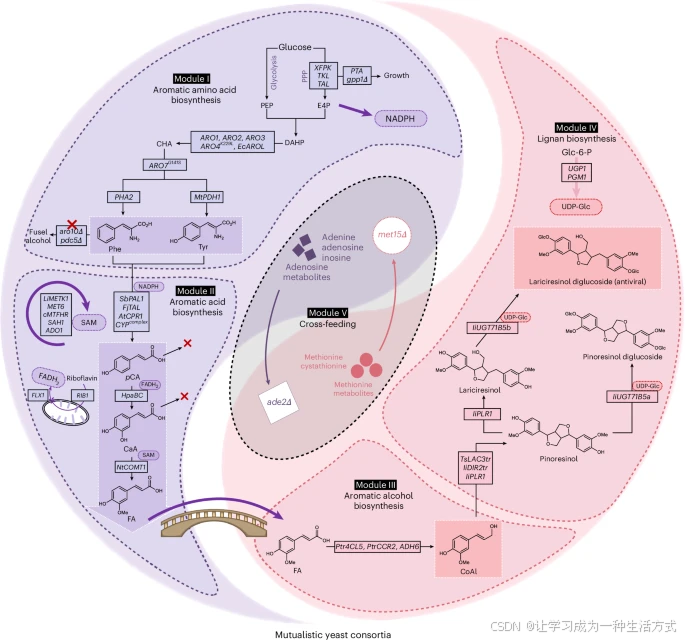
超过40个反应在met15Δ–ade2Δ依赖的酵母共生群体中发生了变化,用于木脂素葡萄糖苷的从头合成。这两个突变株,met15Δ和ade2Δ的生长依赖于甲硫氨酸相关和腺苷相关代谢物(模块V)的交叉喂养。中间体阿魏酸(FA)作为桥梁,帮助菌株避免代谢混乱,并促进木脂素葡萄糖苷的从头合成。所有工程化的途径基因都列在黑色框的矩形框中。操作的辅因子,包括NADPH、S-腺苷-L-甲硫氨酸、FADH2和UDP-葡萄糖,在彩色椭圆中表示。PEP,磷酸烯醇丙酮酸;E4P,赤藓糖-4-磷酸;DAHP,3-脱氧-D-阿拉伯糖七磷酸;CHA,香叶酸;pCA,p-香豆酸;CaA,咖啡酸;CoAl,辛弗酰醇;Glu-6-P,葡萄糖-6-磷酸;UDP-Glc,尿苷二磷酸葡萄糖。使用的基因缩写列在补充数据3中。
在这项研究中,选择了一个外源性酵母配对,met15Δ和ade2Δ,这是最常用的突变体之一,成功地形成了具有严格互惠关系的高度协同的酵母共生群体。进一步利用该共生群体将辛弗酰醇合成的工作分配给两个相互依赖的菌株,使用阿魏酸作为桥梁中间体,因为4-香豆酸辅酶A连接酶(4Cl)是一个关键的多重反应酶,它会优先与前体(例如p-香豆酸、咖啡酸和肉桂酸)反应,并不能将足够的代谢流引入阿魏酸相关的途径,从而阻止下游木脂素的合成。在这个系统中,RB57ade2Δ是阿魏酸的接收菌株,它表达来自不同物种的酶,成功地从阿魏酸合成辛弗酰醇,而阿魏酸是由另一种共培养的阿魏酸产生菌株RB218met15Δ提供的。此外,为了验证酵母共生群体的可扩展性,研究还合成了几种复杂的木脂素,包括松脂醇、紫杉醇和LDG(也称为克莱马斯坦B,是Indigofera indigotica根的抗病毒成分)。这些结果为合成更复杂的木脂素铺平了道路,因为这些分子都是必要的前体。特别是,我们证明了由两个外源性菌株met15Δ和ade2Δ组成的酵母共生体可以像多细胞生物一样分担代谢物合成的工作,有效缓解了木脂素从头生物合成中的代谢混乱,并有望扩展到其他复杂天然产物的合成。
结果
酵母中辛弗酰醇的生物合成
辛弗酰醇是所有类型木脂素的直接前体,但由于其复杂的代谢网络,无法被选择性合成。为了探索在酿酒酵母(S. cerevisiae)中合成辛弗酰醇的最佳途径,我们使用三种芳香酸(*对香豆酸、咖啡酸和阿魏酸*)作为底物,测试由4-香豆酸辅酶A连接酶(4Cl)、肉桂酰辅酶A还原酶(Ccr)和醇脱氢酶(Adh6)组成的芳香醇模块的三步酶组合,以合成相应的芳香醇(图2a)。
此前,大肠杆菌(Escherichia coli)的实验表明,从对香豆酸生成的对香豆醇可以作为可行的底物,通过FADH₂-依赖的4-羟基苯乙酸3-单加氧酶(EcHpaBC)*和*NADH-依赖的黄素氧化还原酶催化转化为辛弗酰醇,并且该转化在拟南芥(Arabidopsis thaliana)*中依赖*咖啡酸O-甲基转移酶(AtComt)。
然而,尽管对香豆酸和咖啡酸的消耗量约为50%(补充图1a),但并未检测到相应的芳香醛或芳香醇(补充图1b),表明酵母与大肠杆菌在芳香醇合成上的机制完全不同,产物可能是芳香酰辅酶A或其衍生物。
此外,我们尝试在酵母中表达PtrCCoAOMT2(黑杨(Populus trichocarpa)*来源的*咖啡酰辅酶A O-甲基转移酶2)和PtrAldOMT2(黑杨来源的咖啡酸3-O-甲基转移酶2),以通过咖啡酸甲基化来驱动辛弗酰醇合成(扩展数据图1a)。然而,这一策略仍未能实现辛弗酰醇的合成(扩展数据图1b)。
相比之下,喂养阿魏酸的酵母工程菌株能够高效转化阿魏酸为辛弗酰醇,并且不同酶组合的芳香醇模块均实现了转化(图2b)。
其中,表现最佳的菌株是RB57,该菌株表达了Ptr4CL5(编码黑杨4-香豆酸辅酶A连接酶5)、PtrCCR2(编码黑杨肉桂酰辅酶A还原酶2)和ADH6(编码酿酒酵母醇脱氢酶6)。该菌株在100.0 mg/L 阿魏酸的培养条件下,成功合成了29.4 ± 1.0 mg/L 辛弗酰醇(图2b,扩展数据图2a)。
综上,我们的研究结果表明,阿魏酸是酿酒酵母中合成辛弗酰醇的最佳底物。
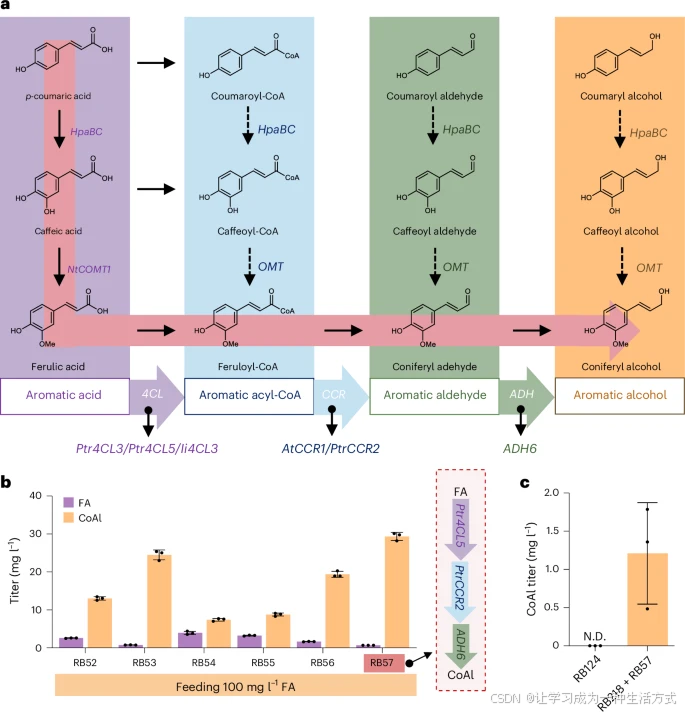
a. 简单苯丙素类化合物的复杂代谢网络,包括芳香酸(紫色)、芳香酰辅酶A(蓝色)、芳香醛(绿色)和芳香醇(黄色)。红色高亮的路径是本研究中生产辛弗酰醇(CoAl)的最佳生物合成途径。
b. 通过不同的酶组合从阿魏酸(FA)生物合成CoAl。红色高亮的RB57菌株具有最高的CoAl产量。在以100 mg/L阿魏酸喂养RB57后,经过96小时发酵测定CoAl产量和剩余FA的含量。
c. 通过RB218和RB57的混合培养(1:1接种)进行CoAl的从头合成。N.D.表示未检测到。研究中使用了三种4Cl基因(黑杨(P. trichocarpa)Ptr4CL3、Ptr4CL5和北刘寄奴(I. indigotica)Ii4CL3)、两种Ccr基因(拟南芥(A. thaliana)AtCCR1和黑杨PtrCCR2)以及一种内源性Adh基因(酿酒酵母(S. cerevisiae)ADH6**)作为候选基因。
除非另有说明,所有后续的摇瓶发酵均在Delft-D培养基中进行,并补充40 mg/L组氨酸和60 mg/L尿嘧啶。发酵液在96小时后进行提取,用于代谢物定量。所有数据均为三个独立生物重复的平均值,误差棒表示标准差(s.d.)。
为此,我们在之前构建的阿魏酸高产菌株RB79中引入了芳香醇模块基因(Ptr4CL5、PtrCCR2和ADH6),试图进行辛弗酰醇的从头合成。然而,工程菌株RB124未能合成辛弗酰醇(扩展数据图2b),表明未知的副反应竞争性地消耗了前体,阻碍了向阿魏酸和辛弗酰醇的代谢通量。
鉴于植物4Cl酶具有广泛的底物特异性,我们推测Ptr4Cl5可能在对香豆酸和咖啡酸转化为阿魏酸之前,就已将它们转化为相应的羟基肉桂酰辅酶A衍生物(图2a和补充图2a)。研究中的三种4Cl候选基因均能催化对香豆酸、咖啡酸和阿魏酸(补充图2b)。
利用双酵母体系优化CoAl合成
为了减少4Cl对对香豆酸和咖啡酸的作用,我们设计了一种双酵母共培养体系,在阿魏酸合成步骤处拆分生物合成途径(图1)。
首先,我们验证了阿魏酸在两种酵母菌株之间可以被有效地分泌和摄取(扩展数据图3a)。实验表明,阿魏酸能有效分泌至阿魏酸高产菌株RB218的培养基中(扩展数据图3b),而且喂养阿魏酸能够被高效利用以合成辛弗酰醇(图2b),这表明阿魏酸可以自由跨越酵母细胞。
一致地,我们观察到共培养RB218和RB57菌株后,能够实现1.2 ± 0.7 mg/L的辛弗酰醇从头合成(图2c),证明了微生物共培养体系是一种可行策略,能够减少副反应。然而,产量波动较大(偏差达52.4%),表明需要构建更稳定的共培养体系以提高合成效率和稳定性。
为了优化代谢分工,我们尝试构建营养缺陷型酵母菌株的共培养体系。我们从CEN.PK113-11C母株中构建了11种常见的敲除突变体,并在摇瓶发酵条件下评估其生长情况。
通过600 nm光密度(OD600)*测定最终生物量(图3a,扩展数据图4a),筛选出*三组最强的互补菌株组合。然而,这些菌株在发酵过程中表现出较长的生长滞后期(图3b),表明发酵时间需要延长。
虽然ade2Δ–ser2Δ和ilv1Δ–arg4Δ组合在144小时发酵后的OD600值略高,但met15Δ–ade2Δ组合具有最高的比生长速率(μmax)。
值得注意的是,ser2Δ突变株在固体培养基上不能生长,但在液体培养基中生长良好(扩展数据图4b),这不符合严格共生的原则。因此,我们最终选择met15Δ–ade2Δ组合进行进一步研究。
随后,我们验证了met15Δ–ade2Δ组合的共培养依赖交叉营养供给,而不是由接种发酵液残留代谢物提供养分(扩展数据图4c)。
在继续研究met15Δ–ade2Δ组合应用于辛弗酰醇生物合成之前,我们希望进一步明确在交叉供养中起关键作用的代谢物。这项研究对理解酵母共培养体系的交叉供养机制具有重要意义。
理论上,不仅甲硫氨酸和腺苷,其相关生物合成代谢物也可能是潜在的交叉供养物质(扩展数据图4d)。
HPLC-MS/MS代谢分析表明,甲硫氨酸、胱硫醚、S-腺苷-L-高半胱氨酸、腺嘌呤、腺苷、肌苷和鸟苷5′-单磷酸可能是交叉供养物质(扩展数据图5a,b)。
代谢通路分析显示,甲硫氨酸、胱硫醚、腺嘌呤、腺苷和肌苷是支持严格共生的关键代谢物,它们通过代谢通量补偿相关代谢缺陷,从而拯救met15Δ和ade2Δ突变株(图3d)。
此外,胞外代谢物浓度(扩展数据图5c,d)与细胞生长行为(图3b)之间的强相关性,再次表明代谢物交叉供养在维持共培养体系稳定性方面至关重要。
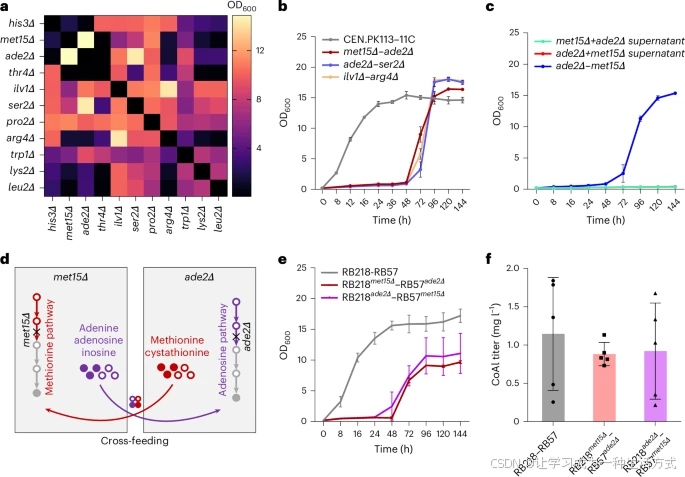
a. 11 种营养缺陷型菌株配对共培养 144 小时后最终 OD600 值的热图分析。两种营养缺陷型菌株的初始接种 OD600 值均为 0.05,总初始 OD600 值为 0.1。在共接种之前,营养缺陷型菌株的种子细胞用等体积超纯水清洗三次。营养缺陷型酵母细胞在不含组氨酸、尿嘧啶和其他额外成分的 Delft-D 培养基中生长。颜色条表示不同营养缺陷型菌株配对(初始接种比 = 1:1)在无外源氨基酸添加的情况下的最终 OD600 值。
b. 三种生长表现较优的酵母共生体的生长曲线,其最终 OD600 值均超过 14。
c. 细胞裂解上清液在与接种体积等效的浓度下,无法支持酵母共生体的建立。
d. 提出的模型描述了 met15Δ–ade2Δ 酵母共生体中潜在的交叉喂养机制。
e,f. 简单混合培养(RB218–RB57)及酵母共生体(RB218met15Δ–RB57ade2Δ 和 RB218ade2Δ–RB57met15Δ)的生长曲线(e)和 CoAl 产量(f)。所有数据均表示三个(a–d)或五个(f)生物学独立样本的均值,误差线表示标准差(s.d.)。
考虑到 RB218 和 RB57 表现出不同的表达模式和生长状态,我们构建了两个方向相反的潜在共生体系(即 RB218met15Δ–RB57ade2Δ 和 RB218ade2Δ–RB57met15Δ 组合;补充图 3)。研究发现,交叉喂养的方向对生长和产物产量具有显著影响(图 3e,f)。RB218met15Δ–RB57ade2Δ 组合的生长比 RB218ade2Δ–RB57met15Δ 组合更稳定,并能够较稳定地产生 0.9 ± 0.2 mg l⁻¹ 的芥子醇。这些结果表明,代谢途径表达差异导致生长变化和共生体系性能的变化。met15Δ 营养缺陷型菌株的致死表型可能更容易通过交叉喂养得到缓解,因此 met15Δ 营养缺陷型菌株应引入生长较慢的菌株(即本研究中的 RB218)。
酵母中紫杉醇和落叶松酚的生物合成
由于紫杉醇和落叶松酚是多种具有活性的木脂素的关键骨架,我们尝试基于酵母共生体系中从头合成的芥子醇,进一步探索其下游合成途径(图 4a)。为了简化实验,采用阿魏酸补料的方法模拟酵母共生体系中的阿魏酸交叉喂养,从而加速下游合成途径的探索与构建。
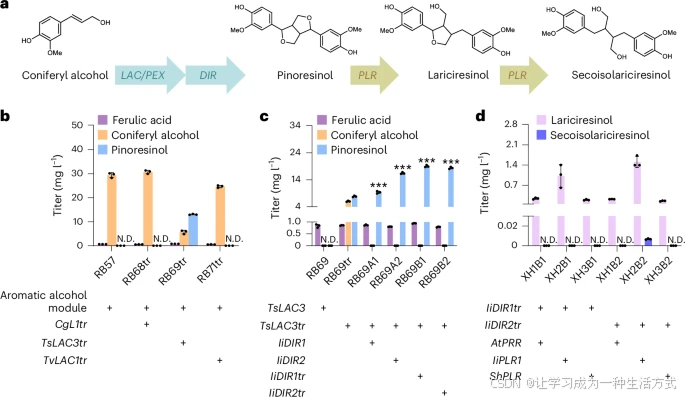
a. 由芥子醇(coniferyl alcohol)生成木脂素(紫杉醇、落叶松酚和松脂醇)的生物合成途径涉及 Lac/Pex、Dir 和 Plr 三种酶催化的三个酶促反应。
b. 经过工程改造的菌株携带芳香醇模块和不同信号肽截短的 Lac,用于将芥子醇(黄色)转化为紫杉醇(蓝色)。
c. Dir 对紫杉醇(蓝色)合成的影响。
d. 针对落叶松酚(粉色)和松脂醇(浅紫色)合成筛选最高效的 Plr 酶。所有菌株在 30°C 下生长于 20 mL Delft-D 培养基中,并在发酵 24 小时后加入 100 mg/L 阿魏酸(FA)。统计分析采用双因素方差分析(***P < 0.001),以 RB69tr 为对照。所有数据均表示三个生物学独立样本的均值,误差线表示标准差(s.d.)。
植物木脂素合成途径的第一步是酚氧自由基偶联反应,以芥子醇为底物合成紫杉醇。该过程可由过氧化物酶(Pex)和漆酶(Lac)两大家族催化。此前研究表明,从 Corynebacterium glutamicum 克隆的 Lac 基因 CgL1 和来自 大肠杆菌(E. coli) 的信号肽截短 Pex 基因 YfeX(本研究中 PEX1tr) 可通过丁香酚供料在 E. coli 中催化合成紫杉醇。然而,这两种酶在 S. cerevisiae(酿酒酵母)(菌株 RB68 和 RB94)中未能合成紫杉醇(扩展数据图 6a,b),可能是由于不同物种中酶糖基化水平的差异。
Lac 属于多铜氧化酶超家族,能够将电子从底物(本研究中为芥子醇)转移至 O₂,生成 H₂O 并产生相应的自由基产物。因此,我们分别表达了 木层孔菌(Trametes sp. C30) 的 TsLAC3 和 多孔菌(T. versicolor) 的 TvLAC1,它们在真菌中表现出较高的氧化能力(扩展数据图 6c)。此外,考虑到 DyP 型 Pex B 家族参与芥子醇的偶联,我们从 PeroxiBase 数据库筛选了四个 PEX 基因,并在细胞质中表达了其信号肽截短版本(PEX2-5tr),用于紫杉醇合成(http://peroxibase.toulouse.inra.fr/)。然而,这些尝试均未成功合成紫杉醇(扩展数据图 6b)。
值得注意的是,TsLAC3 表达菌株 RB69 消耗了所有阿魏酸(扩展数据图 6a),表明 TsLAC3 能催化阿魏酸的转化。已有研究表明,Lac 通常集中于细胞壁,以高效催化木质素合成的自由基偶联反应。因此,我们截短了 Lac 的信号肽以便其在细胞质中功能性表达。最终,在 RB69tr 菌株中表达信号肽截短的 TsLAC3tr 后,从 100 mg/L 阿魏酸成功合成 13.0 ± 0.19 mg/L 的紫杉醇(图 4b)。同时,Lac 与 ABTS 结合的显色反应也验证了 TsLac3 在 S. cerevisiae 中的活性,并表明去除信号肽有助于提高活性(扩展数据图 6d)。
当以两轮供料方式加入 100 mg/L 阿魏酸(扩展数据图 7a)时,第二轮供料导致紫杉醇合成不稳定,产量下降,表明阿魏酸过量时更容易发生未知的副反应(扩展数据图 7b)。此外,TsLac3tr 可催化 对香豆酸、绿原酸、阿魏酸和芥子醇 向多种产物的转化(扩展数据图 7c)。即使涉及芥子醇的双分子自由基反应,也可能随机生成多个二聚体产物,甚至进一步形成高分子聚合物,而阿魏酸的降解可能是主要副产物之一。
为了减少随机自由基偶联导致的副反应,我们引入了 指向蛋白(Dir),其能够控制 β–β′ 键的区域选择性和对映选择性。我们从 I. indigotica(青蓝草) 中筛选了两个 DIR 基因(IiDIR1 和 IiDIR2),并分别表达其完整形式和截短版本(IiDIR1tr 和 IiDIR2tr),以抑制 TsLac3tr 的非特异催化反应。结果表明,所有 Dir 变体均可促进芥子醇消耗,并通过 β–β′ 键区域选择性提高紫杉醇产量(图 4c),且倾向于合成 (−)-紫杉醇(补充图 4b)。特别是,当 IiDir1tr 与 TsLac3tr 共定位于细胞质 时,紫杉醇产量比对照菌株 RB69tr(7.9 ± 0.2 mg/L)提高 143%,达到 19.1 ± 0.4 mg/L(100 mg/L 阿魏酸为底物)。
Plr 是一种 NADH 磷酸(NADPH)依赖性酶,通过两步反应将紫杉醇转化为 落叶松酚 和 松脂醇(图 4a)。拟南芥 AtPrr1 可催化紫杉醇的第一步转化,但对落叶松酚无亲和力,而 I. indigotica 的 IiPLR1 可催化该两步反应,但其对落叶松酚的催化活性极低。因此,我们分别表达了 AtPRR1(拟南芥 Plr1)、IiPLR1(青蓝草)和 ShPLR1(六瓣黄精) 进行筛选。结果表明,这些酶均可在 S. cerevisiae 中合成落叶松酚(图 4d)。
在所有表达菌株中,XH2B1 和 XH2B2(表达 IiPLR1) 的落叶松酚产量最高,分别达到 1.0 ± 0.3 mg/L 和 1.2 ± 0.5 mg/L,并含有近 等量的(−)-落叶松酚和(+)-落叶松酚(补充图 4b)。此外,在 XH2B2 菌株中检测到 微量的松脂醇(6.75 ± 0.48 μg/L),表明 IiPlr1 具有催化落叶松酚向松脂醇转化的能力。
综上所述,我们解析并构建了一个高效的紫杉醇和落叶松酚生物合成途径,这为未来拓展不同结构木脂素的异源合成奠定了基础。
在酵母中生物合成复杂的木脂素葡萄糖苷
接下来,我们尝试利用先前鉴定的催化元件,在酵母中合成LDG(一种来源于I. indigotica 根部的抗病毒成分)。为了实现木脂素葡萄糖苷在酵母中的生物合成,我们将I. indigotica 中先前鉴定出的三种 UGT 基因(IiUGT71B2、IiUGT71B5a 和 IiUGT71B5b)整合到产紫杉醇醇(lariciresinol)的菌株 XH2B2 中。
有趣的是,与体外实验不同,只有IiUGT71B5b(XH12) 在酵母中能够催化 LDG 的合成(当补充100 mg L⁻¹ 阿魏酸时,图 5a)。最终,在 XH18 菌株中过表达三拷贝 IiUGT71B5b,并通过过表达磷酸葡糖变位酶(PGM1)和 UDP-葡萄糖焦磷酸化酶(UGP1) 增强UDP-葡萄糖前体的合成,使得 LDG 产量提高了 217%,达到 133.8 ± 38.6 μg L⁻¹。
我们还测试了这些 UGTs 催化合成 紫杉醇醇葡萄糖苷 的能力。所有测试的 UGTs 均能够在100 mg L⁻¹ 阿魏酸 供给条件下,同时合成 紫杉醇醇单葡萄糖苷 和 双葡萄糖苷(图 5b)。结果表明,IiUGT71B2(XH13) 和 IiUGT71B5b(XH9B1) 更倾向于催化紫杉醇醇单葡萄糖苷的合成,而连续催化生成紫杉醇醇双葡萄糖苷的活性较低。相反,IiUGT71B5a(XH8B1) 更适合催化紫杉醇醇双葡萄糖苷的合成。
本研究通过对高效代谢途径的优化、功能酶的挖掘、亚细胞定位修饰以及自由基偶联反应的区域选择性调控,最终实现了通过向酵母中补充阿魏酸来生物合成 LDG,为其从头合成提供了新的可能性。
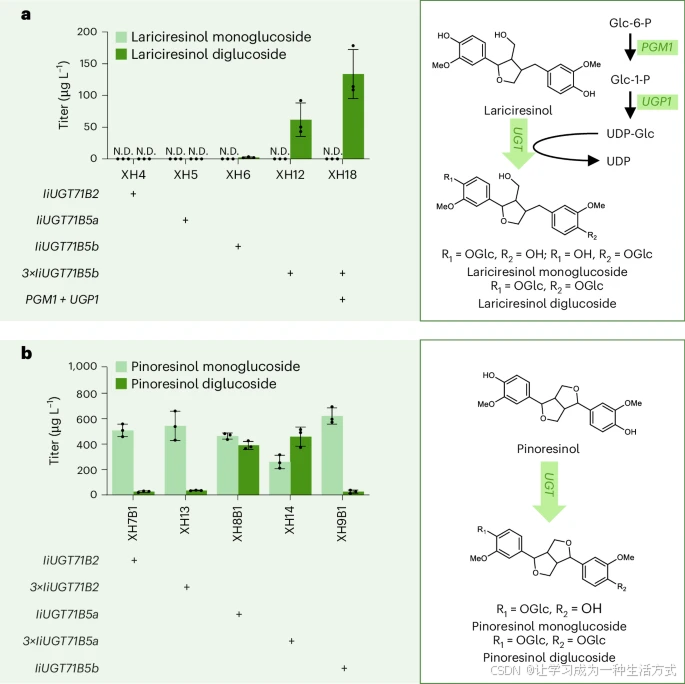
a,b,工程化酵母菌株中表达不同的 UGT 基因以生产紫杉醇醇葡萄糖苷(a)和松脂醇葡萄糖苷(b)。
所有菌株均在 30°C 条件下培养于 20 mL Delft-D 培养基中,并在发酵 24 小时后补充 100 mg L⁻¹ 阿魏酸(FA)。Glu-1-P 代表 葡萄糖-1-磷酸,其中 3× 表示基因的三拷贝表达。所有数据均为三个生物学重复的平均值,误差棒表示标准差(s.d.)。
酵母共培养体系中的木脂素葡萄糖苷生物合成
最后,我们构建了一个用于从头合成抗病毒 LDG 的酵母共培养体系,这一体系提供了一种有利的共生系统,能够缓解酚酸下游代谢途径中的代谢混杂性(图 6a)。
LDG 合成的完整途径 被分解并分别引入以下两类突变菌株中:
-
RB218 菌株:已经构建了阿魏酸生物合成途径。
-
XH12 菌株:表达了所有将阿魏酸转化为 LDG 的关键基因。
在该系统中,阿魏酸 作为桥梁连接两个代谢模块,实现从头合成(图 1),同时,蛋氨酸(methionine)和腺苷(adenosine)途径及相关代谢物充当共生互惠的“交换货币”,促进强制共生关系的建立。
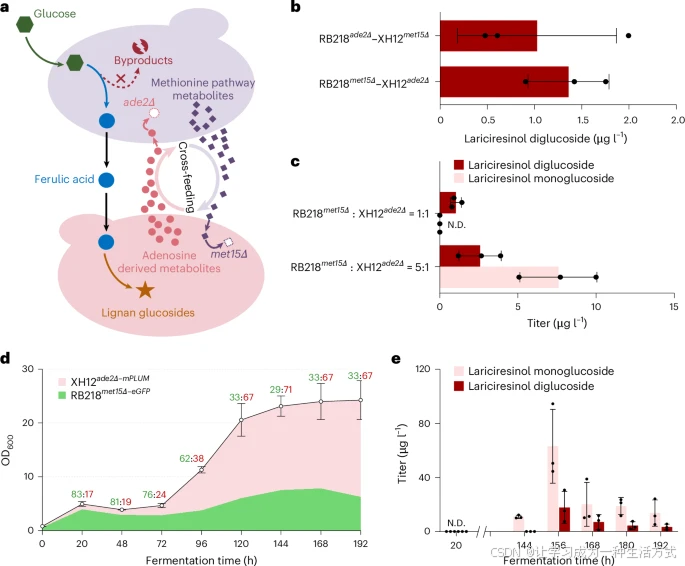
a. 基于 met15Δ–ade2Δ 突变株的酵母共培养体系的工作模型
为了实现木脂素葡萄糖苷的从头生物合成,本研究构建了一个酵母共培养体系,该体系由两个营养缺陷型突变酵母株(met15Δ 和 ade2Δ)*组成,利用*蛋氨酸和腺苷相关代谢物的互补共给,并协同利用阿魏酸(FA) 作为代谢桥梁,从葡萄糖合成木脂素葡萄糖苷。
b. 酵母共培养体系从头生物合成 LDG,即I. indigotica(北板蓝根) 的主要抗病毒活性成分。
c. 在 met15Δ–ade2Δ 共培养体系中,不同接种比下 LDG 和紫杉醇醇单葡萄糖苷的产量。
d,e. 采用RB218met15Δ–eGFP 和 XH12ade2Δ–mPLUM 组成的酵母共培养体系,在补料分批发酵过程中,监测细胞生长及比例变化(d) 和木脂素葡萄糖苷累积情况(e)。在发酵 166 小时前,LDG 的产量可能因积累水平较低而未被检测到。酵母共培养体系的补料分批发酵初始接种比 为 5:1,并于 20 小时后开始补加葡萄糖。所有数据均为三个独立生物学重复的均值,误差棒表示标准差(s.d.)。
本研究利用阿魏酸高产菌株 RB218 和木脂素生物合成菌株 XH12 构建了两种不同的 met15Δ–ade2Δ 共培养体系(图 6b),即:
-
RB218met15Δ–XH12ade2Δ 组合
-
RB218ade2Δ–XH12met15Δ 组合
其中,RB218met15Δ–XH12ade2Δ 组合在合成效率和稳定性 方面表现最佳,能够从 2% 葡萄糖中合成 1.4 ± 0.4 μg L⁻¹ LDG。考虑到阿魏酸的消耗,进一步提高 RB218met15Δ 菌株的阿魏酸供给能力 可能会提高产量。因此,采用 RB218met15Δ:XH12ade2Δ = 5:1 的初始接种比,使 LDG 的从头合成 产量提高至 2.6 ± 1.4 μg L⁻¹,同时合成了前体化合物紫杉醇醇单葡萄糖苷(7.6 ± 2.5 μg L⁻¹)(图 6c)。
此外,通过设计:
-
RB218met15Δ–XH7B1ade2Δ 组合,可从头生物合成松脂醇单葡萄糖苷(1.2 ± 0.5 μg L⁻¹)。
-
RB218met15Δ–XH8B1ade2Δ 组合,可从头生物合成松脂醇双葡萄糖苷(0.5 ± 0.1 μg L⁻¹)(补充图 5)。
考虑到酵母共培养体系在摇瓶水平 下成功合成 LDG,我们进一步探索其在生物反应器 规模上的可行性。为了监测补料分批发酵过程,我们在两种营养缺陷型突变株 中分别表达荧光蛋白:
-
RB218met15Δ–eGFP(绿色荧光)
-
XH12ade2Δ–mPLUM(红色荧光)
使用荧光显微镜 计数细胞(补充图 6),结果显示,随着发酵的进行,菌株比例从接种时的 5:1 稳定调整为 3:7(图 6d)。156 小时时,LDG 和紫杉醇醇单葡萄糖苷的最高产量分别达到了 17.8 ± 11.8 和 63.1 ± 21.3 μg L⁻¹(图 6e)。
本研究在生物反应器的补料分批发酵体系 下,评估了酵母共培养体系的稳定性,并为未来优化提供了参考。
讨论
植物提取仍然是获取有价值木脂素的主要方法,但由于其浓度较低,提取效率受限。植物天然具备木脂素合成的前体分子,但由于代谢通量较弱,需要进行大量的代谢重编程以及异源途径的重构,以提高木脂素的产量。例如,在烟草叶片中瞬时表达十个参与中心代谢和木脂素合成途径的异源基因,可使(−)-去氧鬼臼毒素的含量达到0.43%【49】。然而,在植物基因组中稳定表达大量基因仍然面临巨大挑战【50】。此外,关于转基因植物的安全性问题以及大多数国家的严格法律政策,在当前背景下限制了其进一步发展和应用【51】。因此,近年来,利用工程化微生物进行复杂天然产物的异源生物合成引起了越来越多的关注。
在单一微生物细胞中重建长途径以合成复杂天然产物面临诸多挑战,例如代谢混杂、支路反应以及代谢负担等。而原始植物宿主似乎能够通过多细胞分工来缓解这些问题【4,5,6,7】。因此,人们开始探索利用微生物共培养体系来提高合成效率【3,11,12,14,52,53,54,55,56】。本研究通过酵母的互利共生体系,成功缓解了抗病毒木脂素生物合成过程中的代谢混杂问题。这一成功得益于类似植物的多细胞分工。最终构建的系统涉及超过40处基因修饰(基因表达调控及敲除),并结合了多种综合代谢工程策略,例如辅因子工程、亚细胞分区、代谢重编程和代谢物互馈等(图1)。
为了合成木脂素及其他复杂分子,多个催化反应必须有序协调进行。如果将整个生物合成途径无差别地整合到单一微生物细胞中,势必会引发一系列阻碍目标产物合成的问题,例如代谢混杂、支路反应、代谢负担以及非线性合成失衡等【52】。在木脂素合成过程中,限速酶4Cl(4-香豆酸辅酶A连接酶)具有较宽的底物谱,可将多种苯丙烷酸(例如肉桂酸、对香豆酸、咖啡酸和阿魏酸)转化为CoA硫酯中间体,这可能会导致意外的副反应,从而阻碍木脂素的合成【25】。因此,我们利用4Cl作为关键节点,将木脂素葡萄糖苷合成途径划分至两种具有互利共生关系的酵母菌株中,并以阿魏酸作为代谢传递的桥梁,实现了复杂木脂素葡萄糖苷的从头异源生物合成。该酵母共培养体系促进了生物合成途径的模块化重建,使其具备“即插即用”的特性,从而大幅降低了遗传操作的时间成本,因为可以并行操作多个微生物。此外,由于超过40种催化反应被分配至两株酵母菌中,该体系有效缓解了单细胞中表达大量酶的代谢负担和代谢混杂问题。
在稳定生物合成过程中,构建多个能协调运作的微生物群落至关重要,因为生长占优的菌株往往会在群体中竞争资源【11】。形成一个合成水平相对稳定的共培养体系十分困难(图3f),且在大规模发酵中,这种不稳定性可能会被进一步放大【1】。然而,得益于对胞外代谢组的系统研究以及全基因组挖掘,越来越多的证据表明,共培养酵母细胞之间的代谢物互馈现象十分普遍,自然的交换能力足以支持微生物群落的建立【9,13,31】。在本研究中,我们测试了11种氨基酸和核苷酸代谢相关的酵母营养缺陷型菌株组合,发现 met15Δ 和 ade2Δ 具有较强的协同作用,可形成稳定的酵母共培养体系。此外,近期的一项研究也发现 ade8Δ 能够与 met14Δ、trp1Δ、lys2Δ 和 his3Δ 等多个营养缺陷型菌株形成共培养体系,这进一步验证了我们的研究结果【13】。此外,我们在本研究中进一步验证了基于 met15Δ–ade2Δ 体系在缓解复杂生物合成代谢混杂方面的应用价值。例如,在本研究中,该体系成功应用于木脂素的合成。尽管共培养体系的最终 OD600 值略高于单一野生型酵母(图3b),但其显著的生长滞后现象是一种不良表型,这可能是由于培养前48小时内代谢物交换效率较低所致(扩展数据图5和图3b)。此外,我们发现,RB218met15Δ–eGFP 与 XH12ade2Δ–mPLUM 的比例在接种时为5:1,但随着发酵的进行,逐渐稳定为3:7(图6d)。我们推测,这种变化可能源于两种营养缺陷型菌株(ade2Δ 和 met15Δ)在共培养体系中的相互依赖程度不同,这种现象可定义为“不均衡的互利共生”。具体而言,RB218met15Δ–eGFP 可能对 XH12ade2Δ–mPLUM 依赖性更高,导致 XH12ade2Δ–mPLUM 在群落中占据优势,这可能反映了其对互馈代谢物的更高需求,以及向另一方分泌代谢物的能力更强。因此,通过定制工程策略,优化蛋氨酸相关和腺苷相关代谢物的进出口效率,或许可以进一步提高酵母共培养体系的性能【13】。
综上所述,本研究以木脂素的从头合成为例,展示了酵母共培养体系可以模拟植物的多细胞分工模式,以执行代谢分工并缓解代谢混杂问题。这一体系不仅可用于改善复杂天然产物的生物合成,还可能在其他需要酵母协同作用的复杂生物功能中发挥潜在应用价值。
方法
酵母菌株与试剂
本研究使用酿酒酵母(Saccharomyces cerevisiae)CEN.PK113-11C(MATa,SUC2,MAL2-8c,his3Δ1,ura3-52)衍生的CEN.PK113-11C*(MATa,SUC2,MAL2-8c,his3Δ,ura3Δ,gal80Δ,XI5::(PTEF-CAS9-TCYC1))作为所有基因操作和构建的起始菌株29^。构建的菌株(扩展数据图8)列于补充表1。PrimeStar DNA聚合酶购自Takara BioTek。DNA凝胶纯化和质粒提取试剂盒购自Omega Biotek。除非另有说明,所有化学标准品均购自Sigma-Aldrich。松脂素单葡萄糖苷、松脂素二葡萄糖苷、落叶松树脂醇单葡萄糖苷和LDG购自ChemFaces。
基因工程
所有基因的过表达盒(补充数据2)均通过重叠延伸PCR构建,其中包括启动子、目标基因、终止子以及用于同源重组的下一个基因盒片段。所有引物(补充数据1)及所有密码子优化的酶表达载体(补充数据3)均由Sangon Biotech合成。其他内源启动子、基因和终止子均从大肠杆菌(E. coli)DH5α和酿酒酵母 CEN.PK113-11C的基因组DNA中克隆获得。
所有过表达基因盒均被整合至指定的染色体位点,以确保外源基因能够稳定且高效表达。通过CRISPR-Cas9方法29^,将凝胶纯化后的过表达基因盒与gRNA质粒共转化至酵母细胞中,以实现基因盒的整合。具体而言,利用CRISPR-Cas9网络工具CHOPCHOP选择每个目标基因及基因组位点的两个特定gRNA(补充数据4)。随后,等摩尔量(50–100 ng kb⁻¹)的纯化片段I和II与gRNA质粒共转化入酿酒酵母,采用乙酸锂介导的化学转化法,并在含0.02 g l⁻¹组氨酸的固体合成培养基(SD)平板上筛选转化子。克隆菌株通过菌落PCR和测序进行验证。随后,将经验证的克隆菌株在SD+组氨酸液体培养基中过夜培养,并接种于含SD+组氨酸+尿嘧啶+5-氟乳清酸(1 g l⁻¹)的固体培养基上,以循环去除gRNA质粒并回收URA3标记。对于基因敲除,利用500 ng的片段(由大约500 bp的同源于染色体靶位点上下游区域的序列组成),通过Cas9核酸酶切割引入的基因组断裂点进行同源修复,以实现基因敲除。
单菌株培养
酵母细胞通常培养于YPD培养基(含20 g l⁻¹葡萄糖、20 g l⁻¹蛋白胨、10 g l⁻¹酵母提取物)。含URA3或HIS3标记质粒和基因盒的菌株在SD培养基(含6.7 g l⁻¹无氨基酸酵母氮源和20 g l⁻¹葡萄糖)上筛选,相应地去除尿嘧啶或组氨酸。
在20 ml最小培养基(Delft-D)中进行摇瓶发酵,培养基成分包括2.5 g l⁻¹ (NH₄)₂SO₄、14.4 g l⁻¹ KH₂PO₄、0.5 g l⁻¹ MgSO₄·7H₂O、20 g l⁻¹葡萄糖、2 ml微量金属溶液和1 ml维生素溶液,并根据需要补充40 mg l⁻¹组氨酸和/或60 mg l⁻¹尿嘧啶。培养物以初始OD₆₀₀=0.1接种至20 ml培养基中,置于100 ml锥形瓶中,在220 rpm、30℃条件下培养96小时。


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









