










Multilayered regulation of secondary metabolism in medicinal plants
药用植物次生代谢的多层调控

摘要
药用植物是次生代谢产物(SMs)的巨大储库,这些物质具有重要的医药和工业潜力。然而,由于药用植物中次生代谢产物的积累量较低,获取这些化合物仍然是一项挑战。此外,这些次生代谢产物的合成涉及多个空间、时间及环境调控步骤,形成高度协调的代谢途径。第一层调控涉及一个由转录因子组成的复杂网络;而近年来发现的第二层更复杂的调控机制是表观遗传修饰,包括DNA甲基化、组蛋白修饰和基于小RNA的机制,这些因素可以共同或分别通过调控基因表达来影响次生代谢产物的合成。在本文中,我们总结了关于遗传和表观遗传调控的研究成果,特别强调了药用植物中的次生代谢产物调控机制,为基因表达的多层调控提供了新的视角。
引言
植物作为活体化学工厂,能够合成多种次生代谢产物(SMs)。这些代谢物虽然不直接参与生长和繁殖等主要生命过程,但通常具有重要的生态功能。大多数植物共享基本的生物合成途径,其中大部分初级代谢产物存在于所有组织中。这种代谢核心的维持导致了有限的代谢框架。然而,频繁的修饰,如糖基化、甲基化、酰基化和磷酸化,以及由特定酶介导的少量化学变化,使得基本结构产生多种变体(Morreel et al. 2014)。
药用植物中的不同化学成分具有生物活性,不仅能通过医药和食品工业造福人类健康,还在香水、农用化学品和化妆品行业中具有重要价值(Hassan 2013)(图1)。根据其生物合成途径(图2),次生代谢产物可分为三大类:在莽草酸途径中合成的酚类化合物,在甲羟戊酸途径中合成的萜类化合物,以及在三羧酸循环途径中合成的含氮化合物(Jamwal et al. 2018; Sanchita and Sharma 2018)(图3)。
然而,这些代谢产物的合成面临诸多挑战,例如其在植物组织中的含量较低。因此,研究人员正在探索提高药用植物中这些化合物含量的方法。其中,代谢工程是一种重要策略,可用于增强目标化合物的产量或减少不需要化合物的合成。
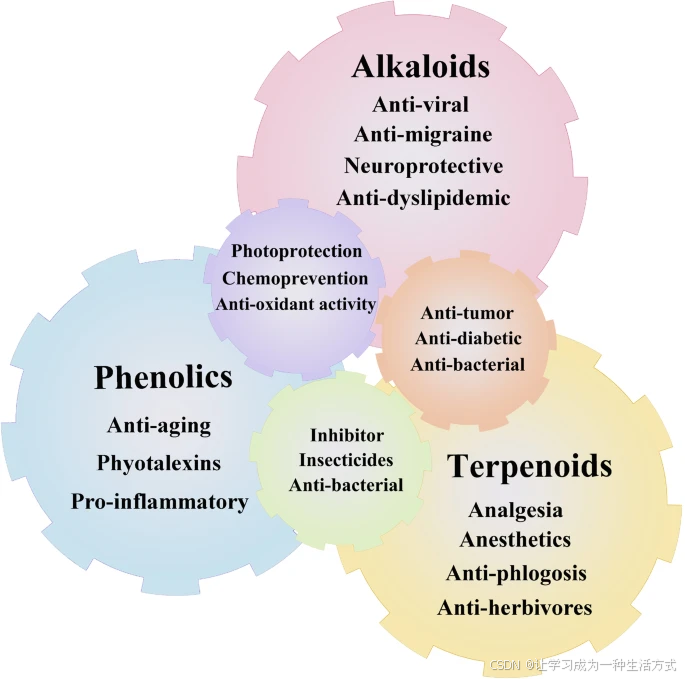
药用植物中次生代谢产物的物理化学性质。酚类、萜类和生物碱等化合物的丰富性使药用植物具有重要的营养保健和药用价值。药用植物中的生物活性化合物具有多种健康益处,包括抗糖尿病、抗癌、催乳、助消化、护肝、调节功能及抗氧化作用,并可用于治疗厌食症、抗结石及抗病原体感染等多种疾病,同时还具有其他多种药用特性。
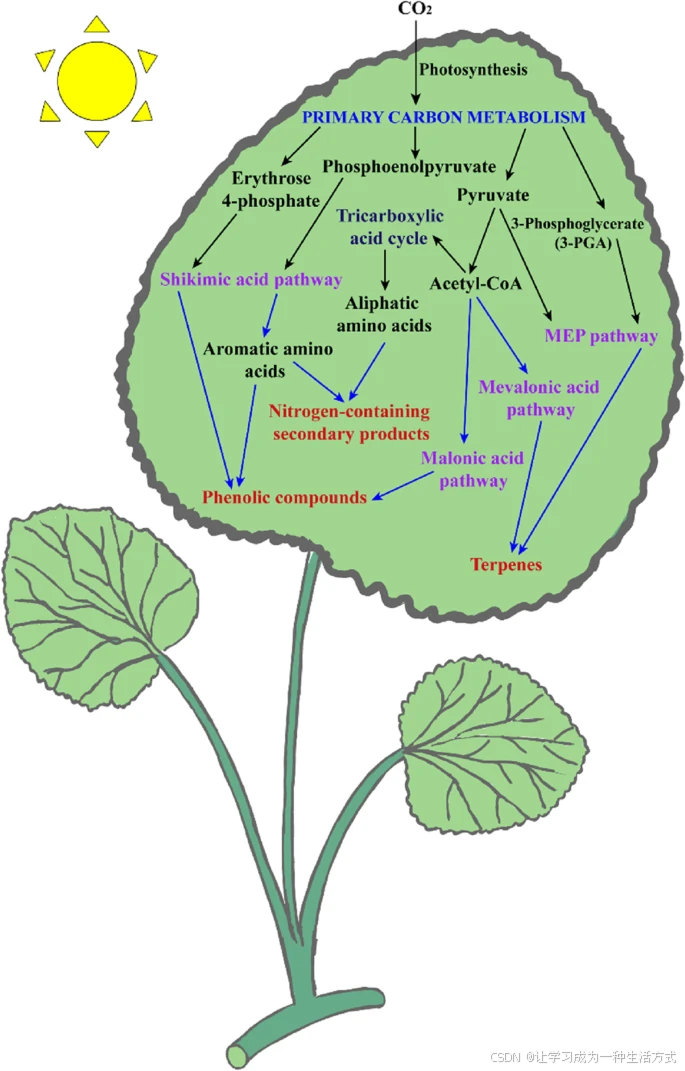
次生代谢产物生物合成主要途径及其与初级代谢相互关系的示意图。初级代谢物通过生物合成途径转化为次生代谢产物,从而与次生代谢相互关联。植物次生代谢的主要生物合成途径包括莽草酸(shikimate)途径、丙二酸(malonate)途径、甲羟戊酸(mevalonate)途径和甲基赤藓醇磷酸(methylerythritol-phosphate)途径。植物次生代谢产物(SMs)主要分为三大类:萜类、酚类和含氮化合物,它们均来源于光合作用这一主要碳代谢过程,并通过上述四种生物合成途径形成。
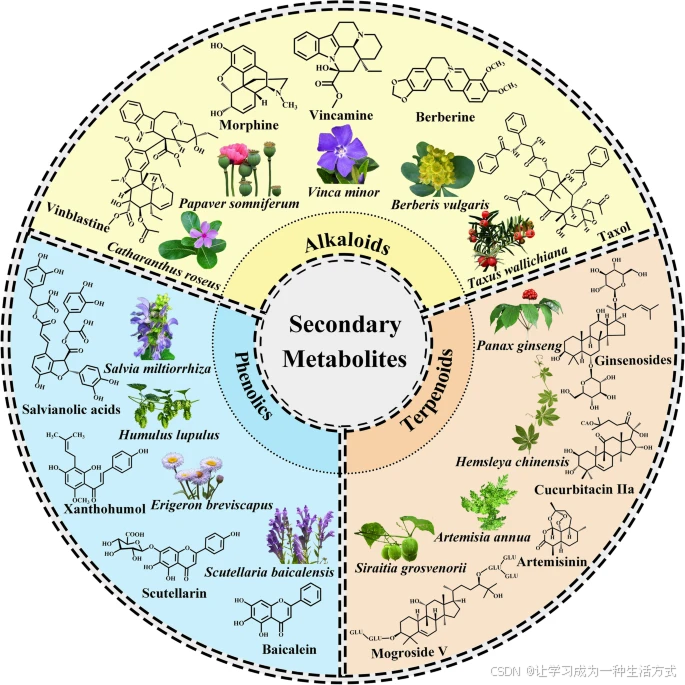
药用植物产生的次生代谢产物的化学结构示例
植物能够产生多种活性成分,这些成分经过鉴定和分离后,可根据其生物合成途径进行分类。
-
酚类化合物:
-
丹参酚酸(Salvianolic acids)——来源于丹参(Salvia miltiorrhiza)
-
黄腐酚(Xanthohumol)——来源于啤酒花(Humulus lupulus)
-
角蕨苷(Scutellarin)——来源于灯盏细辛(Erigeron breviscapus)
-
黄芩素(Baicalein)——来源于黄芩(Scutellaria baicalensis)
-
-
萜类化合物:
-
人参皂苷(Ginsenosides)——来源于人参(Panax ginseng)
-
胡芦巴碱 IIa(Cucurbitacin IIa)——来源于华南胡芦(Hemsleya chinensis)
-
青蒿素(Artemisinin)——来源于黄花蒿(Artemisia annua)
-
罗汉果苷 V(Mogroside V)——来源于罗汉果(Siraitia grosvenorii)
-
-
生物碱类化合物:
-
长春花碱(Vincristine)——来源于长春花(Catharanthus roseus)
-
吗啡(Morphine)——来源于罂粟(Papaver somniferum)
-
长春胺(Vincamine)——来源于小长春花(Vinca minor)
-
小檗碱(Berberine)——来源于小檗(Berberis vulgaris)
-
紫杉醇(Taxol)——来源于喜马拉雅红豆杉(Taxus wallichiana)
-
近年来,科学家对药用植物中次生代谢产物的调控机制(包括转录调控)的理解不断加深。转录因子(TFs)在植物防御过程中发挥作用,能够检测压力信号并调控下游防御基因的表达。然而,目前对如何在不影响植物正常生长发育的情况下调控次生代谢产物积累的分子机制仍不十分清楚(Zheng et al., 2023)。
此外,植物的生存能力、耐受性和生产能力依赖于次生代谢产物的合成增加(即诱导作用)。研究表明,各种生物诱导因子(如真菌、细菌等)和非生物诱导因子(如外源激素)可用于提高植物次生代谢产物的产量,以增强其对环境胁迫的抵御能力(Jan et al., 2021)。
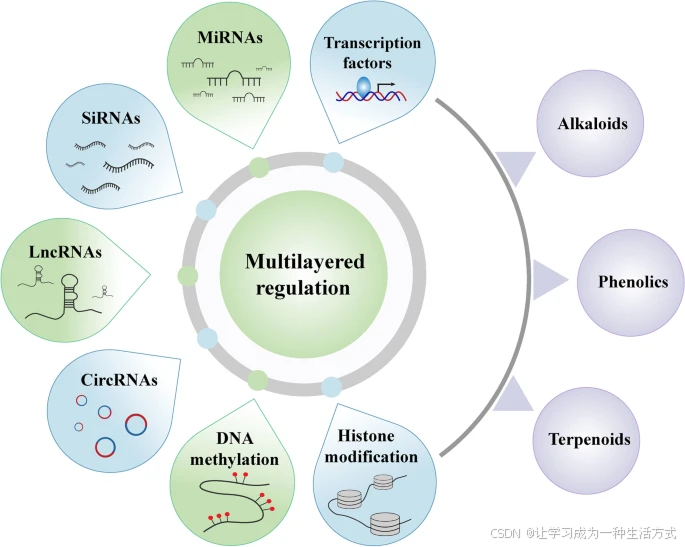
近年来,药用植物次生代谢产物的表观遗传调控引起了越来越多的关注。表观遗传学指的是基因组中非遗传性的可遗传分子修饰,这些修饰可能影响基因表达(Meyer et al., 2018)。表观遗传修饰影响并调控着植物的生长发育和生理过程。例如,DNA甲基化(发生在胞嘧啶位点)已被证实能够影响基因表达、转座子活性和染色体相互作用,从而调节植物的生长发育并响应环境信号(Zhang et al., 2018a; Vidalis et al., 2016)。
最新研究表明,DNA甲基化模式的变化可能通过小RNA影响顺式(cis)*和*反式(trans)*的基因表达,进而影响模式植物*拟南芥(Arabidopsis thaliana)*中的初级代谢和特化代谢途径(Kooke et al., 2019)。此外,**组蛋白修饰**和*DNA修饰可能会影响细胞代谢物与基因表达的关系(Leung et al., 2020)。植物的特化代谢途径受到发育和环境因素的调控,而表观遗传调控可能在其中发挥重要作用。
已有研究表明,DNA甲基化抑制剂能够显著提高丹参(Salvia miltiorrhiza)毛状根培养中的酚类产物合成(Yang et al., 2018)。
在研究次生代谢产物的遗传调控时,需要认识到基因组是它们的“硬件”,而DNA及相关蛋白的表观遗传修饰则是影响基因表达的“软件”。
本文将讨论药用植物次生代谢的多层调控机制,包括:
-
转录调控
-
表观遗传酶的作用(包括“写入者”(writers)、“擦除者”(erasers)和“读取者”(readers))
-
表观遗传修饰在次生代谢产物调控中的关键作用
最后,我们提出一个多层次的基因与表观遗传调控的理解框架,探讨它们在调节基因表达和次生代谢产物积累中的作用。
药用植物次生代谢产物的生物合成
药用植物中的次生代谢产物(SMs)由不同的代谢途径生成,其数量和质量受到不同环境和温度的影响。此外,次生代谢产物的生物合成与植物细胞内的初级代谢高度相关。萜类化合物的合成主要涉及两条途径:甲羟戊酸(MVA)途径和2-C-甲基-D-赤藓糖醇-4-磷酸(MEP)途径,其中MEP途径发生在质体中。在大肠杆菌(Escherichia coli)*中,MEP途径最早被阐明,随后通过生化和基因组学方法鉴定了植物中的同源基因(Rodriguez-Concepcion and Boronat, 2002)。糖酵解的产物,如*丙酮酸(pyruvate)*或*乙酰辅酶A(acetyl-CoA),可用于合成异戊烯基焦磷酸(IPP)*和*二甲基烯丙基焦磷酸(DMAPP),后者是所有萜类化合物的通用前体,分布于植物细胞的多个亚细胞区室(Nagegowda et al., 2010)。
在植物中,酚类化合物主要通过莽草酸(shikimic acid)*和*丙二酸(malonic acid)*途径合成(Ghasemzadeh and Jaafar, 2011)。此外,已有研究表明*丙二酸途径也存在于真菌和细菌中,参与酚类化合物的合成(Cheynier et al., 2013)。在面对多种胁迫因素时,苯丙氨酸解氨酶(PAL)和查尔酮合酶(CHS)共同调控酚类化合物的合成(Sharma et al., 2019)。
含氮次生代谢产物(如生物碱)在其分子结构中含有氮元素,赖氨酸(lysine)、酪氨酸(tyrosine)和色氨酸(tryptophan)等氨基酸是其生物合成的前体。研究发现,不同部位的药用植物中次生代谢产物的组成和复杂程度存在差异,表明这些化合物可能通过特定的调控途径和运输途径在某些器官中合成。SMs 的形成和积累可以通过不同层面的调控实现,从胞外营养物质的运输和代谢开始。因此,研究药用植物中与次生代谢产物生物合成相关的基因表达模式变化至关重要。
次生代谢产物的遗传调控涉及通过基因调控机制控制次生代谢产物的合成和产量。其调控机制可发生在不同水平,包括转录调控、转录后调控、翻译调控和翻译后调控。本文将重点总结药用植物次生代谢产物的转录调控和转录后调控。
次生代谢的转录调控机制解析
转录因子(TFs)通过调控诱导性次生代谢产物的合成以及生物合成基因的表达水平来影响SMs的积累。TFs 是能够与靶基因启动子区域结合的DNA结合蛋白,并通过调节RNA聚合酶的活性来改变转录起始速率。此外,TFs 还可整合外部和内部信号,从而调控次生代谢途径中关键酶基因的表达(Yang et al., 2012)。近年来,科学家们对参与SMs生物合成的TFs及其调控机制进行了大量研究。以下将介绍独立或协同调控SMs生物合成的主要转录因子家族。
bHLH家族
bHLH(basic helix-loop-helix,碱性螺旋-环-螺旋)蛋白是植物中最大的转录因子家族之一,其含有高度保守的bHLH结构域,由螺旋-环-螺旋(HLH)结构和碱性区域组成(Li et al., 2021)。bHLH结构包含45个氨基酸,形成二聚化基序,支持同源二聚化(homodimerization)*或*异源二聚化(heterodimerization)(Kavas et al., 2016; Mao et al., 2017; Xiang et al., 2015)。
已有研究表明,在模式植物拟南芥(Arabidopsis thaliana)*中,共鉴定出162个bHLH转录因子,分为21个亚家族(Toledo-Ortiz et al., 2003);人参(*Panax ginseng*)中含有169个bHLH TFs,分为24个亚家族(Chu et al., 2018);水稻(*Oryza sativa*)中含有167个bHLH TFs,分为22个亚家族(Li et al., 2006);葡萄(*Vitis davidii*)中含有115个bHLH TFs,分为25个亚家族(Li et al., 2021);苹果(*Malus × domestica*)中含有188个bHLH TFs,分为18个亚家族(Mao et al., 2017);白菜(*Brassica rapa ssp. pekinensis*)中含有230个bHLH TFs,分为24个亚家族(Song et al., 2014)。截至目前,在植物转录因子数据库(Plant Transcription Factor Bank, PlantTFDB - Plant Transcription Factor Database @ CBI, PKU)中,共鉴定出166种植物中的28,698个bHLH TFs,其中至少20种植物属于*药用植物。
许多bHLH TFs参与植物的生长发育、非生物胁迫应答(Zhou et al., 2020),并影响次生代谢产物的合成,如生物碱、萜类和黄酮类化合物(Heim et al., 2003; Zhou et al., 2020)。
例如:
-
CjbHLH1调控黄连(Coptis japonica)*中*喹啉类生物碱的生物合成(Yamada et al., 2011)。
-
Catharanthus roseus(长春花)中CrMYC1和CrMYC2参与甲基茉莉酸(MeJA)信号调控,并调控萜吲哚生物碱(TIA)的合成(Chatel et al., 2003; Zhang et al., 2011a, b)。
此外,bHLH TFs 也调控黄酮类化合物的合成,例如:
-
MYC-RP/GP(紫苏,Perilla frutescens)(Gong et al., 1999)
-
GtbHLH1(龙胆,Gentiana)(Nakatsuka et al., 2008)
-
ScbHLH17(千日红,Senecio cruentus)(Li et al., 2020)
近年来,越来越多的bHLH TFs 被发现调控药用植物中萜类化合物的合成。例如,PnbHLH1可与E-box核心序列结合,并调控三萜皂苷的合成(Zhang et al., 2017)。此外,TSAR1和TSAR2调控苜蓿(Medicago truncatula)*中的三萜皂苷合成,并对*茉莉酸信号产生响应(Mertens et al., 2016; Ribeiro et al., 2020)。
总之,bHLH家族在次生代谢产物的调控中起着关键作用,其具体机制值得进一步研究。
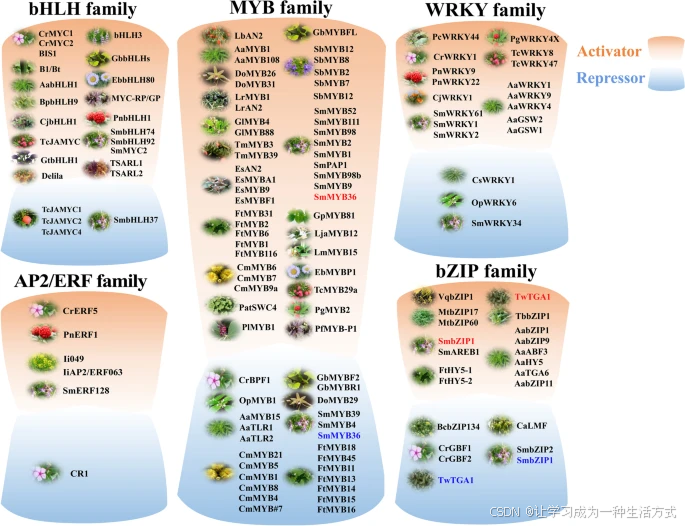
次生代谢产物的转录调控通过转录因子(TFs)改变SMs的诱导合成以及生物合成基因的转录水平。转录因子是能够结合到靶基因启动子区域的DNA结合蛋白,通过RNA聚合酶调节转录起始速率。研究表明,转录因子通过整合外部和内部信号,调控酶基因的表达,从而控制SMs的积累。本文总结了参与药用植物中次生代谢产物生物合成的转录因子网络,其中大量的正向调控转录因子被鉴定出来,但负向调控转录因子的数量较少。负向调控转录因子也在维持植物次生代谢的动态平衡中起着至关重要的作用。
MYB家族
MYB(骨髓母细胞瘤)蛋白是一类广泛参与次生代谢产物合成的转录因子,此外还参与植物的生长、繁殖和应对胁迫等多种生物过程。MYB转录因子的结构域由1至4个重复单元组成,每个单元包含50-53个氨基酸,编码三条α-螺旋(Martin and Paz-Ares, 1997)。根据MYB结构域的数量和位置,MYB转录因子可以分为四类:1R(R1/2,R3-MYB)、2R(R2R3-MYB)、3R(R1R2R3-MYB)和4R(R1R2R2R1/2MYB)(Dubos et al., 2010)。R2R3-MYB是MYB因子的主要类别,已根据其最C端MYB结构域中的基序将其分为28个亚组(Stracke et al., 2001)。自从首次在玉米(Zea mays)中发现MYB转录因子以来,约22,032个MYB转录因子和15,369个相关序列在165种植物中被鉴定出来(Plant Transcription Factor Database, PlantTFDB - Plant Transcription Factor Database @ CBI, PKU)。其中,21种为药用植物(Thakur and Vasudev, 2022)。
MYB转录因子在植物中发挥多种重要生物学功能,包括调控初级和次生代谢、控制生物钟、确定细胞的身份与命运、以及转导激素信号(Cao et al., 2020;Dubos et al., 2010;Ambawat et al., 2013)。次生代谢产物的生物合成是植物应对环境变化的一种生存策略(Jan et al., 2021)。以往对MYB调控次生代谢的研究主要集中在非药用植物中。例如,拟南芥(Arabidopsis thaliana)*中R2R3-MYB亚组4-7已被报道调控苯丙烷类化合物的合成(Liu et al., 2015;Deng and Lu, 2017):亚组4中的*AtMYB32抑制单体木质素的合成;亚组5中的AtMYB123/TT2促进原花青素的合成(Baudry et al., 2004);亚组6中的AtMYB75/PAP1、AtMYB90/PAP2、AtMYB113和AtMYB114调节花青素的合成(Gonzalez et al., 2008;Xie et al., 2006;Borevitz et al., 2000);而亚组7中的AtMYB11、AtMYB12和AtMYB111则调控黄酮醇的合成(Mehrtens et al., 2005;Stracke et al., 2007;Stracke et al., 2010)。
随着药用植物中活性次生代谢产物的开发和利用,越来越多的研究开始聚焦于药用植物中次生代谢产物的生物合成和调控。当前,MYB转录因子在药用植物中调控次生代谢产物的研究主要集中在黄酮类、酚酸类和萜类化合物的生物合成。在黄芩(Scutellaria baicalensis)*中,**SbMYB12**被发现能激活*SbCCL7-4、SbCHI-2和SbF6H-1基因的表达,积极调节黄芩苷和黄芩苷甙的生成(Wang et al., 2022a)。此外,甘草(Glycyrrhiza uralensis)*中的*GlMYB4和GlMYB88能通过茉莉酸甲酯(MeJA)诱导作用,正向调节甘草细胞中的黄酮合成(Li et al., 2020)。银杏(Ginkgo biloba)*是另一种富含黄酮类化合物的药用植物,其中*GbMYBF2和GbMYBFL分别作为抑制因子和激活因子,调控黄酮类的合成(Xu et al., 2014;Zhang et al., 2018b)。
在一系列关于淫羊藿(Epimedium sagittatum)*的研究中,**Huang et al.**识别出多个促进黄酮合成的MYB蛋白,例如**EsMYB9**、**EsMYBA1**、**EsAN2**和*EsMYBF1(Huang et al., 2013;Huang et al., 2016a;Huang et al., 2016b;Huang et al., 2017)。来自丹参(Salvia miltiorrhiza)*的两种MYB蛋白(**SmMYB1**和*SmMYB2)被证明能上调CYP98A14基因的表达,并显著促进丹参酚酸的积累(Zhou et al., 2021;Deng et al., 2020a,b)。此外,SmMYB9b和SmMYB98b在丹参毛状根中正向调节丹参酮的生物合成(Liu et al., 2020)。另一方面,SmMYB4在酚酸类和丹参酮的生物合成中起到抑制作用(Tian et al., 2022)。
此外,Lonicera macranthoides中的一个R2R3型MYB转录因子LmMYB15基因被隔离并表征,研究发现它可能通过直接转录激活4CL基因促进绿原酸的生物合成(Tang et al., 2021)。
关于萜类化合物的转录调控主要涉及单萜、倍半萜和三萜皂苷的合成。例如,作为一种具有显著抗疟作用的倍半萜内酯,青蒿素在黄花蒿(Artemisia annua)*的腺毛中合成并储存。**AaMYB1**可以正向调节腺毛的形成和青蒿素的生物合成,而*AaMYB15则导致AaADS、AaCYP、AaDBR2和AaALDH1基因的表达水平显著下降,青蒿素含量减少(Wu et al., 2021)。AaTLR1和AaTLR2通过抑制腺毛发育来降低青蒿素水平(Lv et al., 2022)。在人参(Panax ginseng)*中,**PgMYB2**被报道通过促进*PgDDS基因的表达,提高人参皂苷的产量(Liu et al., 2019a)。类似地,从三七(Panax notoginseng)*分离的*PnMYB2被认为可能调节人参皂苷的生物合成,但其具体功能尚不明确(Xia et al., 2022)。
WRKY 家族
WRKY 转录因子在植物的种子萌发、种子休眠和应对逆境中的作用非常重要(Rushton 等,2010)。第一个 WRKY 基因是从甘薯中克隆出来的(Ishiguro 和 Nakamura,1994),大量的 WRKY 基因已从多种药用植物中分离并鉴定出来,如青蒿 (A. annua,Ma 等,2009)、人参 (P. ginseng,Di 等,2021)、三七 (P. notoginseng,Zheng 等,2022)、细叶蛇床子 (Ophiorrhiza pumila,Wang 等,2022)、艾草 (Artemisia argyi,Zhang 等,2022) 和大麻 (Cannabis sativa,Liu 等,2021)。WRKY 转录因子具有显著的结构特征;它们的结构包含1到2个 WRKY 域,这是由大约60个高度保守的氨基酸残基组成的DNA结合域。WRKYGQK 是位于 N 端的核心七肽序列,C 端的序列由 C2H2(C-X4-5-C-X22–23-H-X-H)或 C2HC(C-X7-C-X23-H-X-C)锌指结构组成。根据它们的 DNA 结合域(DBD)和锌指结构,WRKY 转录因子分为 I、II 和 III 三个组,II 组进一步分为 IIa、IIb、IIc、IId 和 IIe,分别对应于初级氨基酸序列(Eulgem,2000;Jiang 等,2017)。C2H2 和 C2HC 基序对于 WRKY 蛋白与 W-box(TTGACT/C)顺式元件的相互作用至关重要,因为它们通过识别并结合 W-box(TTGACT/C)启动基因的转录,进而激活或抑制转录,并识别自身的启动子或其他靶基因以实现调控作用(Bakshi 和 Oelmüller,2014)。WRKY 基因的多个 W-box 显示出自我调节和交叉调节是 WRKY 转录因子调控网络的特点。
WRKY 转录因子响应病原体和与防御相关的植物激素,如水杨酸(SA)或茉莉酸(JA),这意味着 WRKY 基因家族在植物免疫中发挥着重要作用(Wani 等,2021)。WRKY 蛋白通过结合病原感染过程中的启动子区域的 W-box 元件,在调节许多基因的表达方面发挥着至关重要的作用,这与 JA 和 SA 信号通路的调节密切相关。甲基茉莉酸(MeJA)增强了三七对 F. solani 的抗性,这表明 JA 信号通路在三七应对 F. solani 感染中起着至关重要的作用(Liu 等,2019b)。WRKY 基因的表达水平在 MeJA 处理后和随后的 F. solani 感染中增加。观察到 PnWRKY9 重组蛋白特异性地结合到 JA 响应和 F. solani 抵抗相关防御素基因(PnDEFL1)启动子中的 W-box 序列。在烟草中过表达 PnWRKY9 显著提高了对 F. solani 的抗性,而 RNAi 介导的 PnWRKY9 表达水平降低使得三七叶片对 F. solani 更易感(Zheng 等,2022)。PnWRKY22 在抗根腐的防御反应中起着枢纽基因的作用。瞬时过表达 PnWRKY22 提高了三七叶片中的水杨酸水平(Ning 等,2021)。
类似于许多其他转录因子,WRKY 转录因子调节次生代谢,包括酚丙烷类、萜类和生物碱代谢(Schluttenhofer 和 Yuan,2015)。关于其他药用植物中参与次生代谢合成的 WRKY 转录因子的相关信息见图 4 和补充表 3。此外,WRKY 转录因子调节多种具有多种生物学功能的植物特有代谢产物。青蒿素是一种广泛用于抗疟疾药物的倍半萜内酯,最初在 Artemisia annua 中发现。之前的研究表明,AaWRKY1 在调节青蒿素生物合成中起着重要作用,并且 amorpha-4,11-diene synthase(ADS)是 AaWRKY1 的靶基因(Ma 等,2009)。此外,腺毛特异性 WRKY 1 AaGSW1 直接结合到 AaCYP71AV1 和 AaORA 启动子中的 W-box,积极促进 Artemisia annua 中青蒿素的生物合成(Chen 等,2017)。进一步研究表明,AaWRKY9 通过直接结合 AaDBR2 和 AaGSW1 启动子的方式积极调节青蒿素的生物合成(Fu 等,2021)。人参属植物,常见的名字是人参,属于五加科,包含传统的药用植物,广泛用于生产人参皂苷。从人参基因组中识别出了 137 个 PgWRKY 基因。共表达分析识别出了 11 个 PgWRKY 基因,可能在皂苷生物合成过程中具有潜在的调控作用(Di 等,2021)。PgWRKY4X 结合到角鲨烯环氧化酶(PgSE)启动子中的 W-box。PgWRKY4X 的过表达显著增强了 P. ginseng 转基因细胞中的人参皂苷积累。此外,分析了 PgWRKY 基因的转录水平,相关性分析表明 GPS、SS、CYP716A47、CYP716A53v2、UGT74AE2、UGT94Q2、PgWRKY1、PgWRKY3 和 PgWRKY8 与总人参皂苷含量显著相关(Yao 等,2020)。蛇床子(Ophiorrhiza pumila)是一种药用植物模型,用于研究抗癌药物广泛使用的五环喹啉生物碱——喜树碱(CPT)的生物合成。在蛇床子基因组中鉴定了 46 个 OpWRKY 基因。OpWRKY6 的过表达显著减少了喜树碱的积累,而在 OpWRKY6 基因敲除株系中,喜树碱的积累反而增加(Wang 等,2022)。大麻中的大麻素是重要的次生代谢物。CsWRKY1 是 delta-9-四氢大麻酚酸合成酶表达的对立调节因子(THCAS)(Liu 等,2021)。
AP2/ERF 家族
AP2/ERF(APETALA2/乙烯响应因子)转录因子家族主要负责调节植物的逆境反应(Shukla 等,2006;Navarro 等,2009;Tang 等,2007;Liu 等,2011)、调节植物的生长发育(Feng 等,2020)以及参与药用植物一些次生代谢途径的调节(Xiao 等,2023;Menke 等,1999;Yu 等,2012)。AP2/ERF 序列包含至少一个由大约 60 个氨基酸组成的 AP2 域。根据其顺式元件,AP2/ERF 具有五个亚家族:ERF(乙烯响应因子)、AP2(APETALA2)、DREB/CRT(脱水响应元件结合因子)、RAV(与 ABI3/VP1 相关)和 Soloist。ERF 亚家族包含一个 AP2/ERF 域(识别基序:AGCCGCC),其主要功能是调节植物对部分逆境的响应和次生代谢途径。AP2 亚家族具有两个高度相似的 AP2/ERF 域(识别基序:GCAC(A/G)N(A/T)TCCC(A/G)ANG(C/T)),其主要功能是调节植物的生长发育。DREB/CRT 亚家族仅包含一个 AP2/ERF 域(识别基序:GCAC(A/G)N(A/T)TCCC(A/G)ANG(C/T)),其主要功能是调节植物对逆境的响应。DREB/CRT 亚家族和 ERF 亚家族的区别在于,两个亚家族的第14和第19个氨基酸不同。RAV 亚家族包含一个 AP2+B3 域(识别基序:CAACA)。其主要功能是调节植物生长发育并响应部分逆境。Soloist 亚家族具有一个与其他亚家族同源性较低的 AP2 域,主要功能是调节植物对部分逆境的响应。
ERF 转录因子在调节次生代谢途径中起着重要作用,尤其是在生产具有高药用价值化合物的植物中。例如,一些人参中的 ERF 基因在冷应激下的表达水平会受到影响,而 PgERF 基因家族对 MeJA 有响应(Chen 等,2020)。在 C. roseus 中,CrERF5 对乙烯和 JA 信号做出反应。关于参与次生代谢合成的 AP2/ERF 转录因子的更多信息见图 4 和补充表 4。CrERF5 在 C. roseus 花瓣中的过表达会导致 MIAs(单萜类生物碱)的生物合成途径上游和下游关键基因的表达水平显著增加,而沉默 CrERF5 则会导致这些基因表达水平下降,表明 CrERF5 通过调节 MIA 生物合成途径中的基因来影响 MIAs 的积累水平(Pan 等,2019)。
bZIP 家族
bZIP(basic leucine zipper motif,碱性亮氨酸拉链结构)转录因子(TFs)是真核生物中最丰富且保守的基因家族之一(Nijhawan et al. 2008)。bZIP TFs 以其共同的 bZIP 结构域命名(Dröge-Laser et al. 2018)。bZIP 结构包含 60 至 80 个氨基酸,由两个部分组成:一个由 20 个氨基酸组成的高度保守的 DNA 结合碱性区域,以及一个相对多样化的亮氨酸拉链区域(Talanian et al. 1990)。碱性氨基酸区域位于 C 端区域,通过固定的 N-x7-R/K 结构进行序列特异性 DNA 结合。亮氨酸拉链区域位于 N 端区域,由几个七肽重复序列或疏水性氨基酸残基(如甲硫氨酸、异亮氨酸、缬氨酸等)组成。
根据 A. thaliana(拟南芥)中 78 个 bZIP TFs 的碱性区域及其他保守区域的相似性,AtbZIPs 被分为 13 个亚家族,包括 A~K、M 和 S(Dröge-Laser 和 Weiste 2018)。不同的亚家族因其功能而命名。例如,A 亚家族在 ABA(脱落酸)信号转导中起核心作用,并能通过直接结合 ABA 反应元件(ABRE;ACGTGG/TC)响应顺式元件,调控靶基因的表达(Ali et al. 2016)。
自 bZIP TFs 在拟南芥和水稻等模式植物中被发现以来,植物转录因子数据库(Plant Transcription Factor Database)已鉴定出 165 种植物中约 15,498 个 bZIP 序列 (PlantTFDB - Plant Transcription Factor Database @ CBI, PKU)。bZIP TFs 在植物信号转导(Hossain et al. 2010)、生物和非生物胁迫响应(Ying et al. 2012)、生长发育调控(Alonso et al. 2009)以及次生代谢物生物合成(Zhang et al. 2015a, b)方面起着重要作用。例如,拟南芥中的 bZIP TFs HY5 和 HYH 可调控花青素合成(Zhang et al. 2011a, b)。柿子(Diospyros kaki)中的 DkbZIP5 参与 ABA 信号响应,其过表达可上调 DkMYB4 的表达,从而促进原花青素的积累(Akagi et al. 2012)。此外,番茄(Solanum lycopersicum)中的 SiHY5 作为 CRY1a 的下游基因,可直接识别并结合花青素生物合成基因 CHS1、CHS2 和 DFR 启动子中的 G-box 和 ACE 元件,进而激活其转录,从而促进花青素生物合成(Liu et al. 2018)。
在 T. brevicorniculatum(短角大戟)中,TbbZIP1 通过 ABA 信号通路调控 TbSRPP1 基因的转录,进而影响天然橡胶的合成(Fricke et al. 2013)。在石榴(Punica granatum)中鉴定出的 PgbZIP16 和 PgbZIP34 可促进烟草叶片中的花青素积累(Wang et al. 2022)。利用 CRISPR/Cas9 技术敲除 VvbZIP36 的等位基因后,VvbZIP36 敲除植株中的一系列花青素生物合成基因被激活,导致相关代谢物的积累(Tu et al. 2022)。
关于 bZIP TFs 在药用植物次生代谢调控方面的研究主要集中在萜类、黄酮类和生物碱的生物合成(详见图 4 和补充表 5)。例如,ABA 诱导可促进青蒿素的积累,并且在 ABA 诱导下,黄花蒿(Artemisia annua)中的 AabZIP1 表达显著上调。研究表明,AabZIP1 可结合青蒿素合成关键酶基因 ADS 和 CYP71AV1 启动子序列中的 ABA 反应元件(ABRE),并激活其表达,从而正向调控青蒿素合成(Shu et al. 2022)。AabZIP9 也可结合 ADS 启动子的顺式元件并激活其表达,进而正向调控青蒿素生物合成(Shen et al. 2019)。
此外,ABA 诱导的 AaABF3(bZIP TFs A 亚家族成员)可直接结合并激活青蒿素生物合成关键基因 ALDH1 的表达,从而调控青蒿素合成(Zhong et al. 2018)。AaHY5(bZIP TFs H 亚家族成员)是光依赖青蒿素合成的核心调控因子,可直接结合泛素 E3 连接酶 AaCOP1,激活与青蒿素合成相关的 AaGSW1 基因的表达,从而调控青蒿素合成(Hao et al. 2019)。
研究表明,柴胡(Bupleurum chinense)中的 BcbZIP134 可能在柴胡皂苷的生物合成中起负调控作用(Xu et al. 2019)。在丹参(Salvia miltiorrhiza)中,bZIP TFs 可通过调控丹参酮的生物合成影响根部形态。SmbZIP7 和 SmbZIP20 与丹参酮生物合成途径关键基因 SmKSL1 和 SmCYP76AH1 共同表达,可能通过影响后两者的表达调控丹参酮合成(Zhang et al. 2018a, b, c)。
SmAREB1 通过正向调控 SmPAL、SmTAT、SmRAS 和 SmHPPD 的表达促进丹酚酸生物合成(Jia et al. 2017)。SmbZIP1 负调控丹参酮生物合成,SmbZIP1 过表达的丹参毛状根中丹参酮含量低于对照组,而 SmbZIP1 敲除突变株中的丹参酮含量显著增加(Deng et al. 2020a, b)。此外,SmbZIP1 可直接结合 C4H1 基因的启动子并激活其表达,从而促进丹酚酸生物合成。
在喜树(Camptotheca acuminata)中,CaLMF 负调控喜树碱合成途径基因 CaTDC1、CaG8O、CaCYC1 和 Ca7DLS 的表达,从而抑制喜树碱的生物合成(Chang et al. 2019)。在长春花(Catharanthus roseus)中,CrGBF1 和 CrGBF2 可特异性结合萜类吲哚生物碱合成关键基因 Str 的 G-box,并通过下调 Str 的表达抑制萜类吲哚生物碱的合成(Sibéril et al. 2001)。因此,bZIP TFs 在调控次生代谢物合成中的生物学功能可以有效提高药用植物的产量和质量。
NAC 家族
NAC 转录因子(TFs)在植物的生长、发育和抗胁迫能力方面发挥重要作用。NAC 转录因子因其包含相似 DNA 结合域的三种蛋白 NAM(no apical meristem,无顶端分生组织)、ATAF1-2 和 CUC2(cup-shaped cotyledon,杯状子叶)而得名。研究表明,NAC 蛋白在植物中广泛存在(Ernst et al. 2004)。在黄花蒿(Artemisia annua)中,已鉴定出 AaNAC1,其可被脱水、低温、水杨酸(SA)和茉莉酸甲酯(MJ)诱导表达。研究表明,AaNAC1 可能有助于提高黄花蒿的青蒿素含量和抗旱能力(Lv et al. 2016)。此外,在喜树碱(CPT)生物合成和调控的共表达网络研究中,OpNAC1 通过抑制脱氧野靛碱酸 O-甲基转移酶(OpLAMT)的表达,参与调控喜树碱的生物合成(Hao et al. 2023)。
非编码 RNA 对次生代谢的调控
虽然转录因子是次生代谢产物的关键转录调控因子,但植物非编码 RNA(ncRNAs)同样参与生物活性化合物的合成。ncRNA 可分为四类,包括微小 RNA(miRNAs)、小干扰 RNA(siRNAs)、长非编码 RNA(lncRNAs)和环状 RNA(circRNAs),它们在调控负责次生代谢合成途径的植物基因方面发挥重要作用。近年来,调控性 ncRNAs 在药用植物中的作用,尤其是 miRNAs,受到了广泛关注。本文总结了当前关于药用植物非编码 RNA 的研究进展,并探讨其在生物活性化合物合成中的作用。
miRNAs
miRNAs 是一类小型非编码 RNA 分子,通过与靶 mRNA 3' 非翻译区(3' UTR)的互补结合,在转录后水平调控基因表达(Bartel 2009;Agarwal et al. 2015)。当 miRNAs 结合其靶 mRNA 时,可导致 mRNA 降解或抑制其翻译,从而影响基因表达。在药用植物中,miRNA 介导的调控有助于控制次生代谢产物的合成,涉及生物合成途径和转录调控因子的调节。此外,miRNAs 还可靶向参与生物碱、黄酮类和萜类等重要次生代谢产物合成的基因(图 5)。例如,在罂粟(Papaver somniferum)中,miR13、miR408 和 miR2161 被认为是调控生物碱生物合成的关键 miRNAs(Boke et al. 2015)。其他研究也报道了多种 miRNAs 参与植物生物碱合成途径,包括小长春花(Vinca minor)(Verma et al. 2020)、印度蛇根草(Rauvolfia serpentina)(Prakash et al. 2016)、长春花(Catharanthus roseus)(Pani 和 Mahapatra 2013;Shen et al. 2017)以及六雄蕊八角莲(Podophyllum hexandrum)(Biswas et al. 2016;Kumar et al. 2018)。
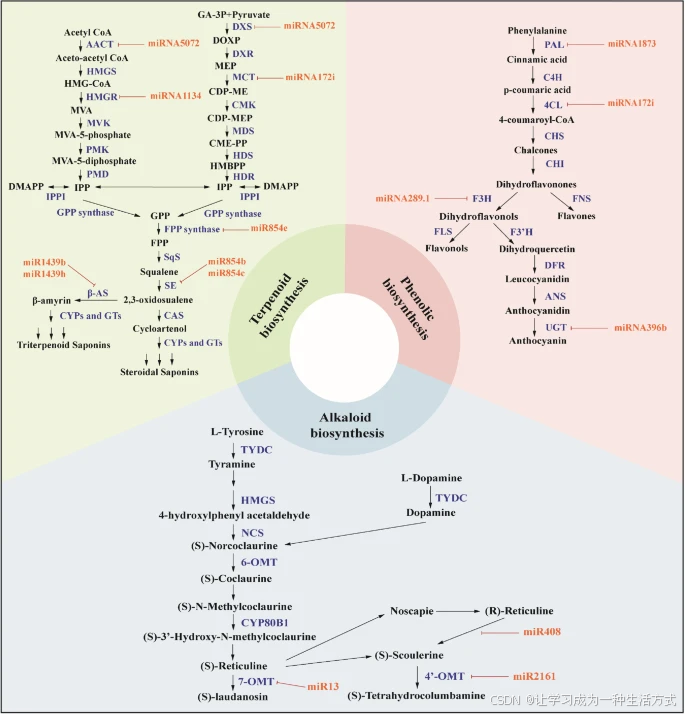
miRNA在萜类、酚类和生物碱生物合成中的调控机制 大量miRNA通过裂解或抑制靶mRNA参与次生代谢物的合成。miRNA的调控作用使植物细胞内的次生代谢物重新定向到特定的生物合成路径,从而提高了治疗性植物代谢产物的产量。该图总结了当前关于miRNA及其在植物次生代谢物(如生物碱、萜类化合物和类黄酮)的调控、生物合成及积累中的作用的研究认知。
关键酶和缩写
-
AACT:乙酰乙酰辅酶A硫解酶
-
HMGS:HMG-CoA合酶
-
HMGR:HMG-CoA还原酶
-
MVK:甲羟戊酸激酶
-
PMK:磷酸甲羟戊酸激酶
-
PMD:甲羟戊酸二磷酸脱羧酶
-
IPPI:异戊烯基二磷酸异构酶
-
DXS:1-脱氧-D-木酮糖-5-磷酸合酶
-
DXR:1-脱氧-D-木酮糖-5-磷酸还原异构酶
-
MCT:2-C-甲基-D-赤藓糖-4-磷酸胞苷转移酶
-
CMK:CDP-ME激酶
-
MDS:2-C-甲基-D-赤藓糖-2,4-环二磷酸合酶
-
HDS:4-羟基-3-甲基丁烯基二磷酸合酶
-
HDR:4-羟基-3-甲基丁烯基二磷酸还原酶
-
FPPS:法尼基二磷酸合酶
-
SQS:鲨烯合酶
-
SE:鲨烯环氧化酶
-
β-AS:β-香树脂醇合酶
-
CAS:环麦角甾醇合酶
-
CYPs:细胞色素P450酶
-
GTs:腺毛
-
PAL:苯丙氨酸解氨酶
-
C4H:肉桂酸-4-羟化酶
-
4CL:4-香豆酰辅酶A连接酶
-
CHS:查尔酮合酶
-
CHI:查尔酮异构酶
-
F3H:黄烷酮-3-羟化酶
-
FNS:黄酮合酶
-
FLS:黄酮醇合酶
-
F3'H:黄酮类3'-羟化酶
-
DFR:二氢黄酮醇-4-还原酶
-
ANS:花色素合酶
-
UGT:UDP-糖基转移酶
-
TYDC:酪氨酸/多巴脱羧酶
-
NCS:去甲可卡因合酶
-
6-OMT:6-O-甲基转移酶
-
CYP80B1:N-甲基可卡因-3'-羟化酶
-
7-OMT:7-O-甲基转移酶
-
4'-OMT:4′-O-甲基转移酶
在萜类生物合成路径中,已识别出其他miRNA并分析了它们的靶标。萜类或异戊二烯类化合物是一个庞大且多样的挥发性有机化合物家族。这些化学物质对于植物在多种生态区域中的生存和进化至关重要,能够对抗生物胁迫、传播种子、增强耐热性并吸引传粉者(Dudareva等,2006;Tetali,2019)。此外,萜类化合物在香料、香精、制药和生物燃料等领域也具有巨大的潜力(Tetali,2019)。
鉴于这些化合物有多种应用,了解它们的多重功能有助于通过基因工程调控和操纵它们的生物合成(Abbas等,2017)。例如,药用植物P. kurroa含有一种名为3-脱氧-7-磷酸庚糖合酶的酶,其mRNA受miR-4995调控,从而调节picroside的合成(Vashisht等,2015)。Saifi等(2015)进一步鉴定了在甜叶菊(Stevia rebaudiana)的甜菊糖苷生物合成路径中起作用的11种miRNA。
类似地,miR7539、miR5021和miR1134可能通过靶向萜类路径上游基因来调节萜类生物合成(Fan等,2015)。在茶树(Camellia sinensis)*中,研究也表明miRNA影响萜类合成(Zhao等,2018)。对*Ferula gummosa的五种miRNA(miR2919、miR5251、miR838、miR5021和miR5658)的分析表明,它们与萜类合成路径相关(Najafabadi和Naghavi,2018)。此外,高通量测序和降解组分析还确定了银杏(Ginkgo biloba)中与萜类三内酯合成调控相关的miRNA(Ye等,2020)。
在濒危药用植物Podophyllum hexandrum Royle中,研究表明miR1438和miR1873通过调节咖啡酰辅酶A O-甲基转移酶和二氢黄酮醇4-还原酶C基因来调控木质素和类黄酮的合成(Biswas等,2016)。 植物miRNA的表达受到环境和遗传因素的调节,如胁迫、病原体感染和激素信号。miRNA在药用植物次生代谢物合成的调控中起重要作用,理解这些过程的调节机制可以为提高有价值药用化合物的产量提供新的策略。
小干扰RNA(siRNA)
小干扰RNA(siRNA)的长度约为21–24个核苷酸,它们参与植物的非生物胁迫响应和次生代谢调控。植物内源性siRNA根据其来源和生物发生途径可分为以下几类:转作siRNA(ta-siRNA)、天然反义链衍生的siRNA(nat-siRNA)和异染色质siRNA(hc-siRNA)(Vazquez等,2010)。在拟南芥(Arabidopsis)中,一个名为OnSEN的类copia逆转座子在热胁迫下通过靶向HSFA1和HSFA2激活siRNA生物合成突变体(Ito等,2011)。
此外,在大豆(Glycine max)*中,siRNA通过靶向CHS基因(如CHS7和CHS8)生成初级siRNA,从而抑制所有CHS基因的表达并抑制类黄酮的生物合成。有趣的是,这种沉默机制仅存在于大豆的种皮中,而不影响其他器官或组织(Tuteja等,2009)。在其他植物中,如*矮牵牛杂交种(Petunia hybrids),类黄酮和花青素的合成也通过siRNA进行调控(Morita等,2012)。
长链非编码RNA(LncRNA)和环状RNA(circRNA)
长链非编码RNA(LncRNA)在植物应对非生物胁迫中发挥着重要作用(Wang等,2017)。此外,有证据表明,药用植物中的LncRNA参与了次生代谢调控。研究已在以下物种中鉴定了LncRNA:丹参(S. miltiorrhiza)(Li等,2015a, b)、人参(P. ginseng)(Wang等,2015)、毛地黄(D. purpurea)(Wu等,2012)和胭脂树(G. sylvestre)(Ayachit等,2019)。
环状RNA(circRNA)在植物发育及应对非生物和生物因素中也发挥着重要作用。例如,葡萄(Vitis vinifera)中的一个源于甘油-3-P酰基转移酶的环状RNA(Vv-circATS1)过表达后,显著提高了拟南芥(A. thaliana)的耐寒性(Gao等,2019)。
此外,环状RNA可能在次生代谢物(SM)生物合成中发挥作用。研究人员探索了环状RNA在丹参(S. miltiorrhiza)中对SM生物合成的调控作用,这种植物在东亚国家被用作药材(Jiang等,2021)。在丹参的三种植物组织中,研究共鉴定和分析了2,476个环状RNA。
其中,SMil_00026090基因编码22α-羟化酶,参与了油菜素内酯的生物合成;SMIL_00014508基因编码ent-考烯酸羟化酶,参与赤霉素的生物合成。此外,SmDXS2与其环状RNA SMscf2473-46693-46978呈显著相关性。研究发现,丹参根部的SmDXS2基因及其环状RNA的表达量显著高于叶片和茎部,这与丹参酮在根部的积累相一致,表明环状RNA可能参与了次生代谢物的合成。
由于目前对药用植物环状RNA的研究较少,上述功能可能无法全面反映它们在SM生物合成调控中的普遍作用。未来的功能性研究将有助于发现其更多的调控功能。
药用植物次生代谢的表观遗传调控
表观遗传学通过影响基因转录及多种细胞过程(如次生代谢物的生产)对植物产生作用。近年来,表观遗传学的研究增强了人们对其调控过程的理解。由于表观遗传机制在基因表达调控中扮演着复杂角色,而这部分机制尚未被完全解析,仅依靠基因操作来增加次生代谢物的产量是不足的(Sanchez-Muñoz等,2019)。
表观遗传学可以用来优化基因工程策略,通过控制关键基因的表达潜在地提高产物产量。广义的表观遗传学涵盖了多种被视为表观遗传的机制。其中,DNA甲基化等表观遗传修饰得到了广泛关注,而组蛋白修饰对于基因表达调控也至关重要(Kumar等,2017)。
这些表观遗传修饰,包括DNA甲基化和组蛋白修饰,影响了包括植物在内的多种真核生物的基因表达(Bird,2007)。本研究将总结表观遗传修饰的潜在机制,重点探讨它们对次生代谢物生产的影响。
DNA甲基化
DNA甲基化是指甲基基团附着在核苷酸的C5位置上(Kumar等,2017)。这种表观遗传修饰发生在原核生物和真核生物中(He等,2011)。许多生物(如植物)仅能对胞嘧啶残基进行甲基化,但在某些条件下,其他核苷酸也可能发生甲基化(Heithoff等,1999)。基因启动子中的甲基化通过两种机制降低转录速率。首先,附加的甲基基团阻止某些转录因子(TF)识别并结合DNA(Cedar和Bergman,2009)。此外,DNA甲基化会吸引特定结合甲基化DNA的因子,从而阻碍转录因子结合(Vanyushin和Ashapkin,2011)。
在植物中,甲基化可进一步分为新甲基化(de novo methylation)*和*维持甲基化(maintenance methylation)(Sanchez-Muñoz等,2019)。植物依赖DRM甲基转移酶通过新甲基化途径来沉默转录活性(Pribylova等,2019)。由于维持甲基化途径的稳定性,DNA甲基化可在多代生物体中遗传(Martienssen和Colot,2001)。
表观遗传修饰调节植物发育和生理过程,包括基因表达、转座子活性以及染色体相互作用,而胞嘧啶位置的DNA甲基化在这些过程中起着重要作用(Zhang等,2018a)。在植物中,DNA甲基化还影响植物对环境信号的响应和发育过程(Zhang等,2018a, b, c;Vidalis等,2016)。在拟南芥(A. thaliana)中,研究发现DNA甲基化模式可能通过小RNA在顺式和反式调控中与基因表达相关联(Kooke,2019)。
DNA修饰可能进一步影响基因表达及代谢产物水平(Leung和Gaudin,2020)。植物的代谢途径受到表观遗传因子的发育和环境调控。例如,苹果果皮的颜色来自花青素,其合成由差异甲基化的基因调控(Li等,2019b)。非模式植物中,DNA胞嘧啶甲基化可在体外调控特化代谢产物的生成。例如,研究发现野生与栽培人参的甲基化谱差异与特化代谢产物的积累相关联(Hao和Xiao,2018)。
此外,在丹参(S. miltiorrhiza)*毛状根培养中,DNA甲基化抑制剂可以显著提高*酚类产物的合成(Yang等,2018)。在鸦片罂粟(Papaver somniferum)的不同器官和栽培品种中,DNA甲基化状态与苄基异喹啉生物碱的水平密切相关(Bulut等,2020)。为探明DNA甲基化在药用植物中特化代谢途径中的作用,仍需进行多组学数据的整合分析。
DNA甲基化是植物和动物中普遍存在的生物现象,不同植物种类间的DNA甲基化水平存在显著差异(Vidalis等,2016)。例如,CG甲基化在三萜皂苷生物合成中起着表观遗传调控作用,表明这种基因家族的表观遗传变化会影响蜕皮甾类化合物(platycoside)的合成(Kim等,2020)。对长春花(C. roseus)的综合分析也揭示了DNA甲基化在调控特化代谢中的作用(Dugé de Bernonville,2020)。
表观遗传学与激素调控的潜在协同作用可能为提高植物次生代谢物的药用生产提供新的方法。此外,在自然恢复和品种依赖恢复过程中,参与次生代谢及光合作用基因的启动子区域也存在不同的DNA甲基化标记(Pagliarani等,2020)。
研究发现,长期在悬浮培养中维持的植物细胞常在次生代谢途径中表现出更高的基因甲基化水平,这已成为限制次生代谢产物大规模生产的重要障碍(Sanchez-Muñoz等,2019)。
在人参(P. ginseng)中,DNA甲基化不仅与其驯化过程相关,还对质量控制起作用。功能分析表明,DNA甲基化与多个基因相关联,说明甲基化在驯化过程中发挥了重要作用(Li等,2017)。五加科人参(P. quinquefolium)的质量由人参皂苷的积累决定,而冷条件在这一过程中至关重要。研究表明,冬季充足的冷暴露会在早春诱发嫩叶的DNA去甲基化,促进根部人参皂苷的积累(Hao等,2020)。
在植物中,胞嘧啶-5 DNA甲基转移酶(C5-MTase)*负责维持DNA胞嘧啶的表观遗传修饰。全基因组分析在丹参中鉴定出了8种候选SmC5-MTase基因,并发现其转录丰度在*胁迫响应和次生代谢中具有重要功能(Li等,2018)。
半夏(Pinellia ternata)(又称半夏块茎)是一种典型的中药材。研究发现,半夏在遮荫条件下种植可以显著提高产量。在比较自然光和遮荫条件下生长的半夏时,研究表明,遮荫处理诱导了32.51%的基因DNA甲基化和16.25%的去甲基化,这表明甲基化变异可能与遮荫条件下半夏产量的增加有关(Shi等,2020)。
此外,研究发现,用5-氮杂胞苷(5-azacytidine)*处理*V. amurensis细胞培养物可以通过抑制DNA甲基化,将白藜芦醇产量提高两倍(Kiselev等,2011)。
组蛋白修饰(Histone Modification)
组蛋白的氨基酸残基可以发生共价修饰,包括乙酰化、甲基化、磷酸化和泛素化。当组蛋白发生乙酰化时,DNA变得更易被转录因子识别并激活基因表达;相反,组蛋白去乙酰化以及部分甲基化的组蛋白位点(如K9和K27)会紧密结合染色质,抑制基因表达(Liu等,2014;Strahl等,2000)。
组蛋白乙酰化在调节植物基因表达、种子萌发、形态建成以及胁迫响应中起关键作用(Liu等,2016)。研究表明,表观遗传变化能够影响植物的多种生长和发育过程,例如开花、种子萌发以及对生物和非生物胁迫的响应(Ahmad等,2010;Zhao等,2019)。
由于非模式植物通常具有较长的生长周期和庞大的种群规模,因此其表观遗传学研究的进展相对缓慢。然而,随着表观遗传研究方法的发展,在过去二十年中,次生代谢的表观遗传调控研究取得了显著进展。这些研究提升了我们对非模式植物次生代谢表观遗传调控机制的理解。
近年来,表观遗传修饰在许多植物次生代谢调控中被广泛研究并证实。例如,磷(Pi)缺乏条件下,苹果(Malus)叶片中的花青素积累受到miR399d与表观遗传修饰的协同调控(Peng等,2020)。
尽管我们在揭示花青素生物合成及其调控机制方面已取得重大进展,但关于此途径中表观遗传调控的基本机制仍不清楚。研究发现,在杨树(Populus)*中,**JMJ25基因**与花青素生物合成相关,并通过遗传和生化研究方法对其功能进行了表征。JMJ25通过调节染色质和DNA的甲基化状态*负调控MYB182的表达,从而抑制花青素的合成(Fan等,2018)。
结论
药用植物次生代谢产物(Secondary Metabolites, SMs)在各行业中的重要性引发了对其合成途径调控的广泛兴趣。通过操控药用植物基因组中的沉默和隐性合成途径,可以生产具有高治疗价值的天然化合物。这些调控途径的操控可能会提高次生代谢产物的产量。此外,合成生物学方法(如利用微生物或酵母的异源宿主)为提高这些化合物的生物技术生产提供了一个有前景的平台。目前,大肠杆菌(E. coli)、酿酒酵母(Saccharomyces cerevisiae)和谷氨酸棒杆菌(Corynebacterium glutamicum)等微生物,作为微生物生物转化的底盘细胞,能够生产自然界或化学合成难以获得的高丰度生物活性化合物(Ajikumar等,2010;Paddon等,2013;Galanie等,2015;Luo等,2019)。然而,对于那些含有多个P450反应或路径复杂不明的合成途径,异源重建几乎难以实现(McElroy和Jennewein,2018)。因此,由于这些在微生物中重建复杂途径的障碍,目前植物体系仍然是合成次生代谢产物的更优选择(Zhu等,2021)。但无论是微生物还是植物,鉴定生物合成途径仍是次生代谢产物合成的最大挑战。
过去几十年间,不同植物物种中已经鉴定出许多重要的次生代谢产物,并阐明了其生物合成途径。此外,鉴定参与次生代谢产物合成及修饰的基因,尤其是那些负责化合物修饰的基因,至关重要。近年来,得益于高通量技术“组学”的发展,药用植物研究取得了显著进展。在基因组学领域,人们预测了多个次生代谢产物生物合成基因簇,提升了发现新型生物活性化合物的潜力。其他组学技术(如转录组学、转译组学、互作组学、蛋白质组学及代谢组学等)正在被用来构建药用植物中生物过程的系统层面理解,而其中的表观基因组层面尤为关键。表观基因组学的目标在于探索环境如何影响基因表达,并将其他“组学层面”置于有意义且相关的生物学背景中。
目前,表观遗传学已成为增强药用植物中生物活性化合物浓度的重要工具。随着对新型药物需求的不断增加,表观遗传修饰成为识别高通量天然产物的有效方法之一。表观遗传修饰的关键功能之一在于激活沉默的次生代谢基因簇,从而提高生物活性化合物的产量。在植物细胞培养中,次生代谢产物的生产受到多种表观遗传机制(如DNA甲基化和组蛋白修饰)的调控。然而,在长期培养的植物细胞中,这些机制可能导致次生代谢产物产量的下降。尽管代谢工程方法(如诱导、合成途径基因的过表达、竞争性途径基因敲除以及转录因子工程)是提高次生代谢产物产量的有效工具,但随着培养时间延长,其效果往往会减弱。这是实现植物细胞培养商业化生产的重要障碍。
相比之下,表观遗传工程结合诱导或转录因子工程可以显著提高生物合成效率,同时减少如生长减缓等不利效应。传统代谢工程技术在短期内可提高次生代谢产物产量,而表观遗传工程技术则确保这些改变在时间上更稳定,不易被代偿性调控系统抑制。此外,鉴于某些药用植物基因组复杂、转化效率低等问题,建立强大的毛状根或悬浮细胞代谢工程平台显得尤为重要。通过使用更高效的植物转化技术(如多靶点CRISPR/Cas9技术),创建具有复杂遗传和表观遗传变化的细胞系已逐渐成为可能。这一新方法不仅能为植物细胞培养产品提供更加经济的解决方案,还能将当前尚未投放市场的新产品推向市场。通过利用表观遗传工程的潜力,植物细胞培养所生产的药物及其他天然产物将变得更加经济和易于获取。
在次生代谢产物合成后,这些化合物可能在不同的组织或细胞器内积累,并执行相应的生物学功能。这些特化代谢产物通过跨膜转运蛋白(如ATP结合盒转运蛋白(ABC)、多药和毒性化合物外排蛋白(MATE)、嘌呤渗透酶(PUP)家族)进行远距离运输(Shitan等,2014)。随着基因组学及多组学分析的进展,植物基因组中已有一些转运蛋白基因被注释。例如,三种MATE蛋白(CmMATE1、ClMATE1、CsMATE1)被鉴定为通过分泌CuB、CuE和CuC来增强植物适应性的关键蛋白(Zhong等,2022;Ma等,2023)。
未来,药用植物中代谢产物转运蛋白的鉴定将降低代谢工程的成本和时间成本,简化化合物的纯化流程,并使这些化合物能够通过细胞外泵直接从细胞中分离,从而提高整体生产效率。


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









