










A near-complete genome assembly of Catharanthus roseus and insights into its vinblastine biosynthesis and high susceptibility to the Huanglongbing pathogen
长春花的近乎完整基因组组装及其长春花碱生物合成与对黄龙病病原高易感性的见解

长春花(Catharanthus roseus),一种原产于马达加斯加的药用植物,是两种重要抗癌药物——长春花碱(vinblastine)和长春新碱(vincristine)的唯一来源,这两种药物通过单萜来源的吲哚生物碱(MIA)途径合成(Das et al., 2020)。它还是研究黄龙病(HLB)的重要模式植物,黄龙病是导致全球柑橘产业巨大经济损失的毁灭性病害(Ma et al., 2022)。长春新碱是一种获得FDA批准的药物,已用于急性淋巴细胞白血病、慢性髓性白血病的淋巴母细胞危机以及霍奇金/非霍奇金淋巴瘤的治疗(Below and M Das, 2021)。在长春花中,长春花碱的生物合成受多种关键酶的复杂调控;不同的前体物质被定位于特定的组织,并可能在不同组织和细胞类型之间转运(Caputi et al., 2018; Liu et al., 2021)。然而,长春花碱生物合成途径中的几个基因仍未被鉴定(Zhang et al., 2022)。长春花还是研究多种农业重要的难培养细菌病原的优秀模式植物(Zeng et al., 2022),如黄龙病的主要致病菌——Candidatus Liberibacter asiaticus(CLas)。因此,长春花的高质量基因组组装不仅有助于我们理解MIA的生物合成和基因调控途径,还为黄龙病的研究提供了重要的基因组资源。2015年,基于短读长测序的长春花草种基因组草图已经完成(Kellner et al., 2015),但它存在碎片化和不完整的问题。因此,需要改进的基因组序列以便进行更详细的基因组分析,特别是MIA的生物合成和黄龙病防控研究。
为了实现这一目标,我们使用Illumina、PacBio、BioNano和Hi-C平台(补充表1)结合全面的组装策略(补充图1)组装了‘Pacifica XP Burgundy’品种的长春花基因组。经过contig组装、染色体锚定、空隙填充和精修后,我们获得了一个高质量的染色体级基因组组装,基因组总长度为561.7 Mb,包含89个scaffold,scaffold N50为71.1 Mb(补充表2)。其中,一条染色体从端粒到端粒完全组装,四条染色体组装为单端端粒(图1A;补充表3)。为了评估组装质量,我们将Illumina和PacBio的测序读段分别映射到已组装的基因组上,得到的总体读段映射率分别大于96%和99%(补充表4)。我们在组装基因组中识别出了98%的BUSCO基因(补充表2),并且Merqury评估的共识质量和完整性值分别为38.3和97.5。根据Hi-C数据,我们没有检测到任何大规模结构的错误定向或组装错误(补充图2)。因此,该组装在连贯性和完整性上代表了显著的改进。其质量可与最近发布的两个长春花基因组(Li et al., 2023; Sun et al., 2023)相媲美,并且具有更高的连贯性(补充表2)。对重复元素的注释显示,组装中有73.71%是重复序列:转座元素占68.59%,长末端重复逆转录转座元件占43.11%(补充表5)。Ty3/Gypsy和Ty1/Copia元素分别占组装基因组的16.17%和19%。通过同源性、转录组引导和自发基因预测的整合,我们鉴定出了30,085个编码蛋白的基因,且识别到了96.9%的完整BUSCO基因(补充表2)。超过98.57%的基因通过NR和TrEMBL数据库搜索进行了功能注释。这一高连贯性的基因组组装使我们能够识别出25个基因簇,其中七个基因簇包含了与长春花碱生物合成途径相关的基因(补充图3)。
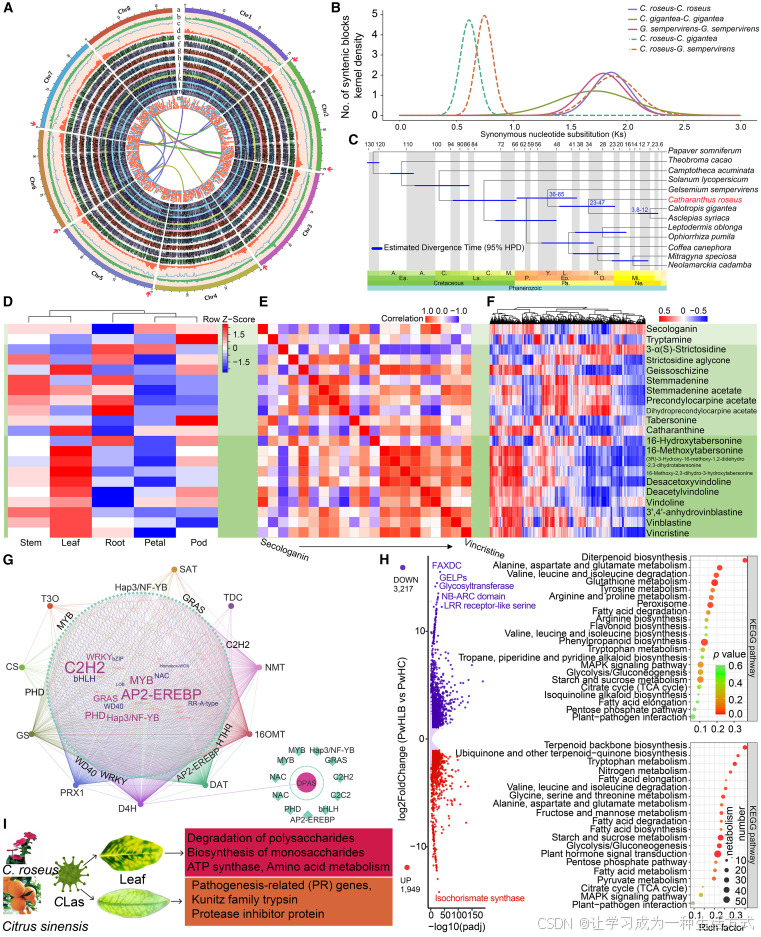
图1. 长春花基因组的组装、注释与比较分析,长春花碱生物合成途径分析及CLas感染的转录组反应
(A) 长春花基因组序列的环形图。 (a) 最外层的彩色块为8个伪染色体的圆形表示。 (b–m) GC含量、重复覆盖、基因密度以及花、柱头、茎、叶、种子、水果、幼茎、根及子叶特异性基因的表达。 (n) 长春花在CLas感染下的差异表达基因分布(调整后的p值 < 0.05 和 log折叠变化 > 2)。红色方框表示下调基因,蓝色圆点表示上调基因。内圈的线条代表共线性选择的同源基因之间的联系。染色体外层的红色箭头表示组装的端粒序列。
(B) 长春花及其他物种的同义替代率(Ks)分布,比较同源基因和旁系基因。
(C) 使用RAxML推断的最大似然系统发育树及长春花的分化时间。节点上的蓝色条表示估计日期的95%可信区间。
(D) 长春花不同组织中与长春花碱相关的代谢物概述。内标为secologanin和tabersonine。
(E) 长春花中与长春花碱生物合成相关的代谢物含量之间的相关性。
(F) 热图展示了长春花碱生物合成途径中物质含量与基因表达水平(每千碱基每百万转录本TPM)之间的相关性。
(G) 参与长春花碱生物合成的结构基因、假定转录因子及其他结构基因的调控网络。位于调控网络中心的词云代表与长春花碱生物合成相关的转录因子,字体大小表示相关转录因子的相对数量。
(H) 火山图显示了CLas感染后长春花的差异表达基因。基因的最大折叠变化和显著的p值在图中进行了注释。右侧面板显示了差异表达基因的KEGG富集分析。圆圈的颜色代表通过Benjamini和Hochberg方法校正的超几何检验的假发现率,圆圈的大小表示与给定KEGG术语相关的基因数量。
(I) 长春花和甜橙的CLas症状差异及其拟议的遗传基础。 (D)–(F)中具有相同的代谢物标签,列于(F)右侧。
古代全基因组重复是所有现存开花植物表型和进化创新的最重要推动力之一。通过同义替代值(Ks)和四重同义第三密码子反转(4DTv)频率分布的分析,清晰地识别出了长春花基因组中大约1.9的单一Ks峰(图1B;补充图4),代表了所有现存核心双子叶植物共同的古代全基因组三倍体事件(WGT-γ)(Rai et al., 2021)。长春花基因组还包含0.95%的片段重复,55.38%的分散重复,8.48%的串联重复,以及5.68%的邻近重复(补充图5;补充说明1)。系统发育分析揭示,长春花最晚在36-65百万年前与Asclepias syriaca–Calotropis gigantea的谱系发生分化,随后A. syriaca和C. gigantea在23-47百万年前发生了分化(图1C)。通过将长春花的基因家族与其他物种的基因家族进行比较,我们能够识别出与单萜类化合物生物合成功能相关的长春花特有基因(补充说明1)。
MIA(吲哚生物碱)途径可以大致分为三个部分(上游、中游和下游),每个部分都有特定的酶反应和中间体在不同组织和细胞类型间的转运(Zhang et al., 2022)。在长春花基因组中,注释出78个基因参与从secologanin和色氨酸开始的长春花碱生物合成途径(补充图6;补充表6),这些基因按簇排列在染色体上(补充图7)。高度连贯的组装和3D基因组分析表明,染色体构象在调控长春花碱生物合成相关基因的表达方面可能发挥重要作用(补充说明2)。为了进一步研究长春花碱的生物合成,我们通过基于液相色谱-质谱(LC-MS)的代谢物谱分析了长春花不同组织中的相关代谢物(图1D)。我们鉴定了21个与长春花碱相关的代谢物,其中大多数在叶片中积累较高水平,尽管色胺和tabersonine主要存在于荚果中。除16-羟基tabersonine外,所有下游步骤的代谢物主要存在于叶片中。路径中相邻代谢物之间存在强相关性(图1E),它们展示了相似的基因调控特征。具体来说,上游代谢物受到多种基因的广泛调控,而下游代谢物则受限于更窄范围的基因调控(图1F)。为了进一步了解涉及长春花碱生物合成的基因调控,我们进行了加权基因共表达网络分析,识别出大量假定的调控基因和转录因子(TFs)(补充图8)。这些转录因子包括bHLH、AP2/ERF、WRKY和bZIP家族,已被报道在响应茉莉酸时调控MIA途径(Liu et al., 2021)。一些以前未报告的转录因子(如C2H2、植物同源域、GRAS、MYB和核因子Y蛋白)也被推测在长春花碱生物合成调控中发挥重要作用(图1G)。我们根据催化反应的底物和产物的化学结构特征,鉴定了几个候选基因,包括P450、氧化酶和酰基转移酶基因,这些基因可能控制从3′,4′-anhydrovinblastine合成长春新碱。这些候选基因是从基因共表达模块中识别的,这些模块与3′,4′-anhydrovinblastine、长春花碱和长春新碱的含量存在显著相关性(补充图9)。
接下来,我们想探究长春花比柑橘更快速表现出CLas感染明显症状的遗传基础(补充图10;补充说明3.1),尽管这两种物种在碳代谢和其他基本代谢过程中高度相似(补充说明3.2)。在长春花感染CLas后,共有5166个基因(占17.17%)在反应CLas感染时发生差异表达,这些基因显著富集于碳水化合物代谢、次生代谢、氨基酸代谢和脂肪酸代谢途径中(图1H;补充表7和8;补充说明3.3)。我们还比较了涉及三羧酸(TCA)循环的基因,发现柑橘中存在大量的IDH基因,而长春花则有更多的PDHB亚家族基因,这些基因在CLas感染后也表现出差异表达。CLas的基因组注释显示,这种独特的细菌需要与宿主共享基于氧化磷酸化的有氧呼吸过程、TCA循环和磷酸戊糖途径。它的厌氧呼吸依赖于与宿主的糖异生过程,并依赖宿主的氨基酸作为氮源,来获取所需的能量进行代谢活动(补充说明3.4.1)(Duan et al., 2009)。对与氧化磷酸化相关基因的检查显示,长春花中与细胞色素c氧化酶相关的基因比柑橘中多,细胞色素c氧化酶作为氧化磷酸化中的末端氧化酶,在CLas中缺失。此外,转录组分析显示,长春花中有20个(11.6%)与氧化磷酸化相关的基因在CLas感染后发生了差异表达(补充图11B)。我们还分析了CLas感染后长春花和柑橘中与氨基酸代谢相关的基因表达情况。与色氨酸和缬氨酸等必需氨基酸及非必需氨基酸脯氨酸的代谢相关基因在长春花中CLas感染后发生了差异表达,这表明CLas可能直接从长春花中获取这些氨基酸,随后这些氨基酸可进入TCA循环转化为能量或用于蛋白质合成。我们接下来检查了与病原抗性相关的长春花和柑橘基因,包括编码核苷酸结合富含亮氨酸重复蛋白、蛋白激酶、WRKY转录因子和GDSL酯酶/脂肪酶的基因。系统发育推断、基因位点和转录组分析共同表明,这些基因的数量减少且缺乏物种特异性聚集。值得注意的是,在长春花中之前报道的76.3%的抗性基因在CLas感染后没有表现出显著的差异表达(调整后的p值 < 0.05 和 |log2 fold change| ≥1),这可能解释了为什么CLas在长春花上迅速表现出明显的症状(补充图11;补充说明3.4)。综上所述,对基因组和转录组数据的综合分析提供了初步证据,表明长春花比柑橘更具作为黄龙病(HLB)研究遗传模型的潜力(图1I)。
总之,长春花的高质量基因组组装提供了有关长春新碱生物合成和CLas易感性的遗传基础的见解。它为进一步的功能基因组学研究奠定了基础,在这些研究中,长春花可以作为一个优秀的系统,用于研究CLas与宿主植物相互作用的机制,并鉴定参与长春花碱生物合成的调控基因。


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









