






 本文报道了链霉菌通过产生精氨酸酮(ARGK)与真菌如曲霉和青霉进行跨界互作,特别是阿扎霉素F的发现,揭示了细菌信号如何影响真菌的天然产物生物合成。研究还探讨了精氨酸酮在塑造土壤微生物群落中的作用,以及其在不同真菌间的普遍性。
本文报道了链霉菌通过产生精氨酸酮(ARGK)与真菌如曲霉和青霉进行跨界互作,特别是阿扎霉素F的发现,揭示了细菌信号如何影响真菌的天然产物生物合成。研究还探讨了精氨酸酮在塑造土壤微生物群落中的作用,以及其在不同真菌间的普遍性。
作者:张金龙,南京农业大学硕士在读,主要研究土壤链霉菌与mVOCs。
周刊主要展示LorMe团队成员优秀周报,每周定期为您奉上学术盛宴!本期周刊为您介绍链霉菌聚酮所介导的微生物互作,原文于2023年发表在《Nature Microbiology》。

导读
虽然原核微生物与真核微生物的互作对生态系统的功能至关重要,但目前相关的研究仍然较少。作者的研究表明,链霉菌产生的一种聚酮—精氨酸酮(Arginoketides)介导了该菌和曲霉属、青霉属真菌的跨界互作,并促进了真菌产相应的天然产物。精氨酸酮有环状和线状两种,其中环状精氨酸酮—阿扎霉素F(Azalomycin F,这是一种链霉菌生产的抗生素)由伊朗链霉菌产生,它诱导了构巢曲霉(Aspergillus nidulans)中隐性BGC(生物合成基因簇)苔色酸(Orsellinic acid,苔色酸盐具有清除自由基的活性)基因的表达。合成精氨酸酮的细菌和响应该信号的真菌从同一土壤样本中分离出来,基因组分析和文献检索表明,精氨酸酮生产者在世界各地都有发现。因此,除了它们的直接影响,精氨酸酮诱导真菌天然产物的间接作用,可能会有助于改善整个土壤微生物群落的结构和功能。
结果
伊朗链霉菌bld基因突变体不会诱导真菌基因表达
为了确认真菌中苔色酸BGC是如何激活的,作者敲除了链霉菌的Bld调节因子。据报道,BldD蛋白和BldH蛋白以及由bldA基因编码的识别TTA密码子的tRNA调控着链霉菌的生长发育和天然产物合成,所以作者敲除了相关的基因,然后发现当bldD,bldA和bldH的缺失突变株与构巢曲霉共培养时,发现苔色酸并未被诱导。因此,由Bld调节因子可能控制了细菌诱导真菌BGC活化的过程。
基于蛋白质组学、转录组学数据,与野生型相比,ΔbldA缺失突变株的许多基因转录水平下调,蛋白质表达水平下降,其中就包括了阿扎霉素F。代谢组的数据也表明,ΔbldA缺失突变株的培养提取物中缺乏与阿扎霉素F对应的分子质量。
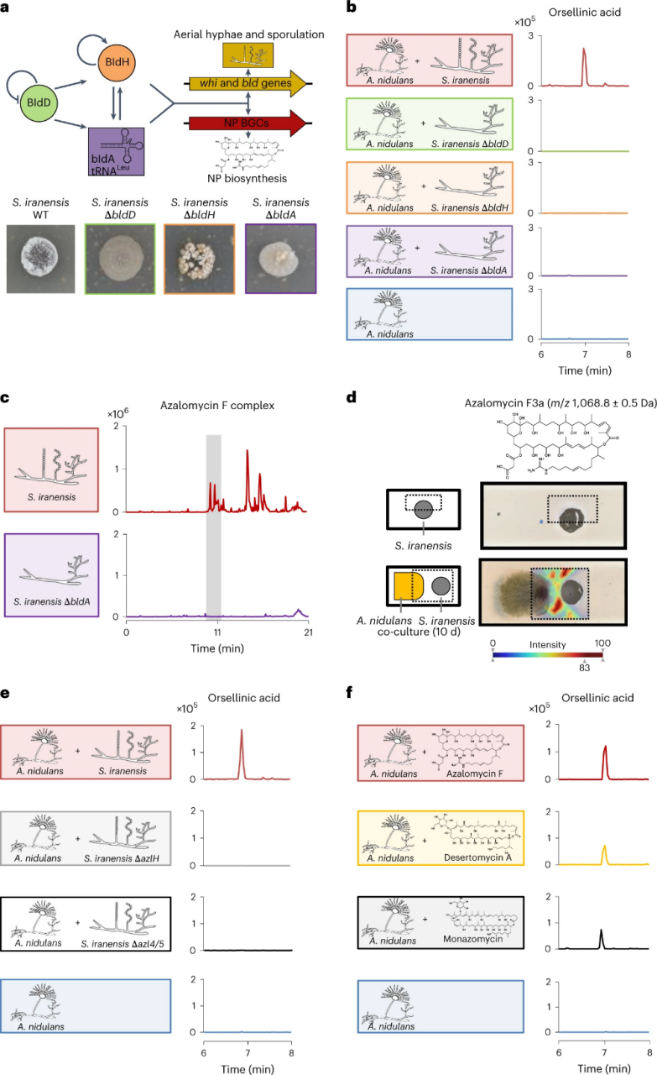
图一 链霉菌产精氨酸酮诱导构巢曲霉生产苔色酸及其衍生物的鉴定
a,bld基因的级联调控(上)。伊朗葡萄球菌的ΔbldD、ΔbldH和ΔbldA缺失突变株缺乏气生菌丝和孢子形成(下)。b,分别对野生型、突变株(ΔbldD,ΔbldH和ΔbldA)伊朗链霉菌与构巢曲霉的共培养物的上清液进行HPLC-MS分析,提取的苔色酸(m/z 167 [M-H]−)的离子色谱图。c,野生型链霉菌和ΔbldA缺失突变株培养物的总离子色谱图。灰框表示仅在伊朗链霉菌野生型中发现的阿扎霉素 F复合物的洗脱时间。d,在琼脂培养基上对共培养10 d的伊朗链霉菌与构巢曲霉进行MALDI-IMS分析。样品制备的可视化示意图(左);方框表示测量面积。MALDI-IMS分析阿扎霉素F3a的分布(m/z 1068.8±0.5Da;右)。使用热图来描述分析质量的丰度,从低(蓝色)到高(红色)丰度。e,从高效液相色谱-质谱分析得出的苔色酸(m/z 167 [M-H]−)的提取离子色谱图,该色谱图来自于对构巢曲霉与阿扎霉素F缺陷株伊朗链霉菌的共培养提取物的分析。f,苔色酸(m/z 167 [M-H]−)的提取离子色谱图,来源于用纯化的阿扎霉素F处理的构巢曲霉培养提取物的HPLC–MS分析。
伊朗链霉菌在构巢曲霉存在下分泌阿扎霉素F
为了可视化阿扎霉素 F的空间分布,作者采用了基质辅助激光解析-电离(MALDI)成像MS (IMS)技术对伊朗链霉菌和构巢曲霉共培养。检测到两种可能的代谢物:阿扎霉素F 3a和阿扎霉素F 4a,它们与计算出的阿扎霉素 F3的质量[M + H]+ 1068.6583很好地吻合。同时它们的分布表明,阿扎霉素F的生物合成只在靠近真菌的细胞中被激活,所以阿扎霉素F的产生是受到了构巢曲霉的影响。
阿扎霉素F激发构巢曲霉中天然产物的生物合成
为了证明阿扎霉素F是触发真菌天然产物生物合成的细菌信号,作者分析了缺失阿扎霉素F生物合成基因的突变菌株。分别将伊朗链霉菌突变株ΔazlH、Δazl4、Δazl5与构巢曲霉共培养,发现并没有产生苔色酸或其衍生物。作者将10µg ml−1体外分离纯化的阿扎霉素F加入到单培养的构巢曲霉中,观察到诱导产生了苔色酸。这种处理下的阿扎霉素F浓度与共培养处理下的阿扎霉素浓度相当。
总的来说,阿扎霉素F是一种受欢迎的细菌诱导剂,可以激活构巢曲霉中天然产物的生物合成。为了探究该反应是否是一种特异性的反应,作者在另一种名为FGSC A4的野生型构巢曲霉中加入了等量的阿扎霉素F,该菌株也与阿扎霉素F发生了反应,产生了苔色酸, 茶渍酸(Lecanoric acid)以及化合物F-9775a和F-9775b 。
精氨酸酮激发真菌中天然产物的生物合成
环状精氨酸酮由宏内酯骨架和含有胍基或氨基的侧链组成,例如分别由大链霉菌(S. macronensis)和马休链霉菌(S. mashuensis)产生的沙漠霉素A(desertomycin A)和单唑霉素(monazomycin),鉴于作者发现了纯化的沙索霉素A和单唑霉素也可以诱导构巢曲霉中苔色酸及其衍生物的产生,作者又提出产生这些化合物的放线菌也能够诱导真菌苔色酸BGC表达,随后在大链霉菌、马休链霉菌与构巢曲霉的共培养物中确实发现了苔色酸及其衍生物。.
作者还测试了奥索霉素B(Oasomycin B),这是一种侧链中缺乏氨基的沙漠霉素家族的化合物,诱导真菌天然产物合成。氨基部分的重要性体现在抗菌活性方面,与沙漠霉素A相比,奥索霉素B失去了抗菌活性,而重建奥索霉素B的带正电片段后,即可恢复抗菌活性。实验结果表明,等浓度的奥索霉素B仅最小程度地触发真菌产生苔色酸及其衍生物。因此,作者得出结论,沙漠霉素的带正电荷部分可能是诱导真菌天然产物生物合成所必需的,这可能适用于所有环状精氨酸酮。
为了确定精氨酸酮诱导活性的特异性,作者测试了已知的抗真菌化合物,这些物质要么作用于细胞壁生物合成,如卡泊芬净(Caspofungin),要么作用于真菌细胞膜,如两性霉素B(Amphotericin B)或伏立康唑(Voriconazole)。与精氨酸酮类抗生素相反,这些结构上不同的抗真菌剂,至少在测试的浓度下,没有诱导出相应化合物的产生。
生产精氨酸酮细菌的全球分布
为了加速对激活ors BGC的化合物的测试,作者构造了GFP-orsA报告菌株,该菌株在orsA活化后发出绿色荧光。为了确定精氨酸酮生产者在自然界中出现的频率,作者随后从德国Eutingen Gäu( 48.4661535°N, 8.7291989°E)附近的土壤中分离放线菌。在600毫克土壤中,作者能够分离培养出305种代表潜在放线菌的丝状细菌。 当这些分离的菌株与报告菌株共培养时,其中 8株触发了绿色荧光,随后对这些细菌进行测序,然后使用德国微生物和细胞培养物数据库(Deutsche Sammlung von Mikroorganismen und Zellkulturen,DSMZ)进行分析,确定了不同分离株的系统发育距离。分离株48、102、124、176、219和280要么彼此亲缘关系较近,要么是同一种。根据计算出的平均核苷酸同一性值(ANI),它们很可能代表了利巴尼链霉菌(S. libani)的分离株。作者发现分离株45与其他分离株明显不同,ANI值证实了这一点,因此,作者继续研究分离株45和219,后者作为菌株7、48、102、124、176、219和280的代表。
作者使用antiSMASH和BlastN,分析了分离株45和219基因组数据,以寻找精氨酸酮生物合成所需的聚酮合酶中的精氨酸负载结构域。分析表明细菌分离株219的基因组具有潜在的利地霉素(lydicamycin)BGC,而分离株45携带了(线性霉素)linearmycins的推定BGC。这一发现很有趣,因为它将诱导化合物合成的线索扩展到线性精氨酸酮上。随后对细菌分离株219和45的培养提取物进行的高性能LC-MS(HPLC-MS)、高分辨率MS(HR-MS)和串联MS (MS/MS)分析,证实了30-去甲基裂地霉素和linearmycin家族化合物的产生。同时将市售的linearmycin A和30-去甲基裂地霉素在添加到构巢曲霉培养物中,发现也诱导了苔色酸及其衍生物的形成。作者假设产生该类物质的菌株也应该能够诱导真菌产化合物。情况确实如此,因为在之前分离株的共培养实验中,链霉菌分离株2和219就是分别通过产生利地霉素和linearmycin触发苔色酸及其衍生物的形成的。因此,不仅环状精氨酸而且线性精氨酸能够诱导真菌天然产物的生物合成。
使用分析技术鉴定土壤中的天然产物具有挑战性,因为它们倾向于吸附到土壤颗粒上。因此,作者评估了报告菌株对阿扎霉素F的敏感性,发现它对低至10ng.ml-1的浓度有反应。将土壤上清液添加到构巢曲霉的培养物中,与没有土壤上清液的培养物相比,nLuc(荧光素酶)的活性明显增加。由于之前的研究表明ors BGC是由精氨酸酮特异性诱导的,所以精氨酸确实存在于土壤中。
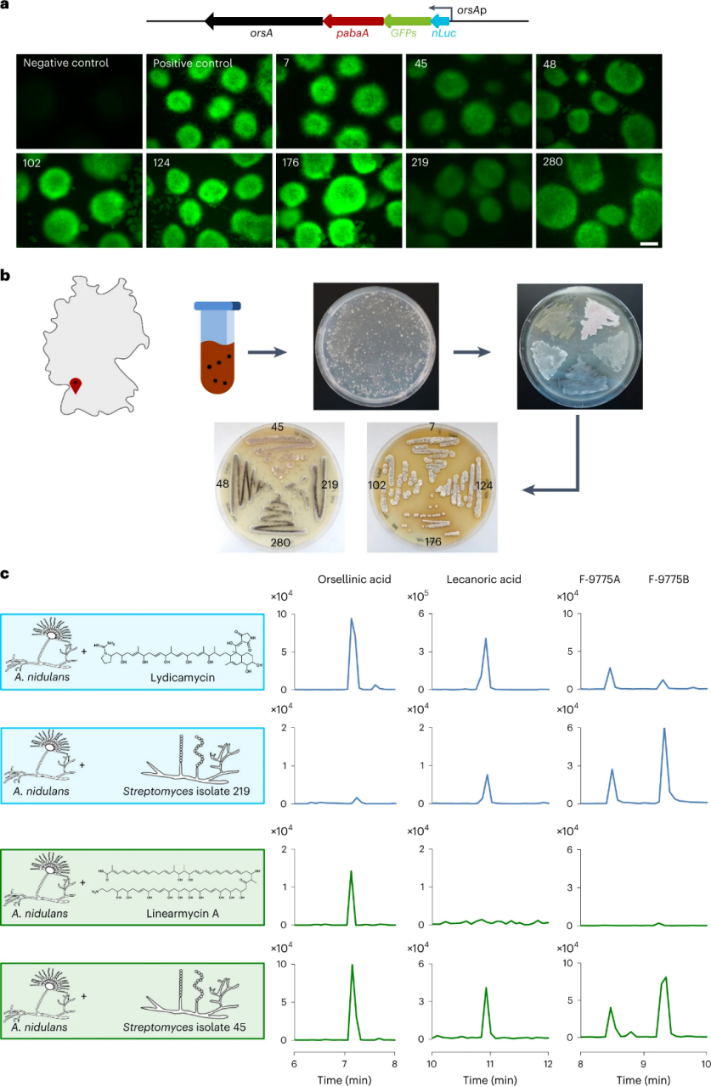
图二 产生精氨酸酮的土壤细菌的分离及激活构巢曲霉的BGC
a,orsAp-nluc-GFPs报告基因融合示意图(上),链霉菌土壤分离株7、45、48、102、124、176、219、280与含报告基因的菌株共培养6小时的图像(下)。阴性对照为单培养的含报告基因的构巢曲霉,阳性对照为与已知的伊朗链霉菌菌共培养的含报告基因的构巢曲霉。b,德国地图,注明了土壤样品的产地(左上)和细菌分离的工作流程(右)。c、用利地霉素、linearmycin和土壤分离株链霉菌219和45处理构巢曲霉,从共培养提取物的HPLC-MS分析中提取的奥塞林酸及其衍生物的离子色谱图(右)。从培养提取物的HPLC-MS分析中得到奥塞利酸(m/z 167 [M - H]-)、茶渍酸(m/z 317 [M - H]-)以及F-9775A和F-9775B (m/z 395 [M - H]-)的离子色谱图。
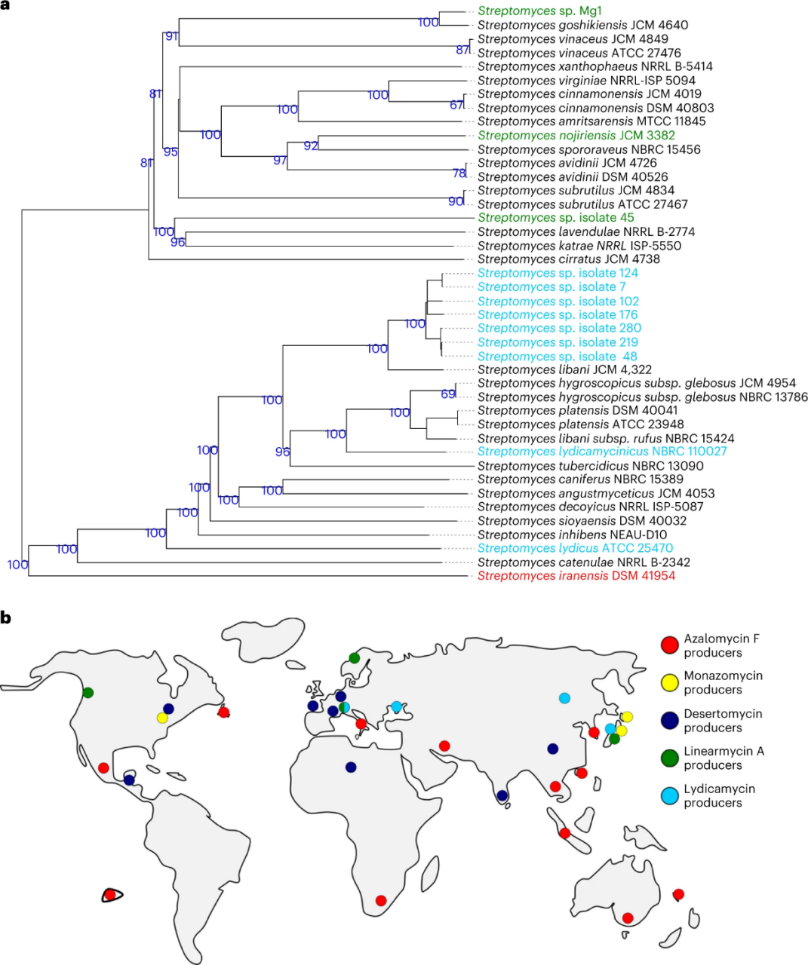
图三 精氨酸酮生产者的系统发育树及地理分布
真菌对精氨酸酮反应的频率
为了深入了解精氨酸酮是否会被构巢曲霉以外的真菌感知,作者又分析了烟曲霉,这是一种感染后可以引起生命危险的人类病原体。尽管在系统发育上与构巢曲霉关系遥远,但烟曲霉也会在雷帕霉素链霉菌(S. rapamycinicus)和伊朗链霉菌的特异性诱导下激活烟环素和烟熏素的BGCs。同时,作者将纯化的阿扎霉素F添加到烟曲霉ATCC 46645的单一培养物中时,它产生了烟环素和烟熏素。类似地,用阿扎霉素F处理烟曲霉CEA10分离株时也产生了烟曲霉素,表明阿扎霉素F介导了链霉菌与这些真菌的互作。
在这一基础下,作者试图确定整个土壤环境中潜在的响应该信号的真菌。为此,作者从之前分离链霉菌的土壤样品中分离出真菌菌株,共获得了 106 株真菌,并测试了它们对伊朗链霉菌 WT 和伊朗链霉菌 ΔazlH 缺失突变株的反应。结果表明,在106株真菌中,有31株在与伊朗链球菌WT共培养时表现出培养颜色的变化,与ΔazlH缺失突变体共培养时并未发现该变化。此外,作者还对这些共培养物进行了HPLC-MS分析。发现只有当分离的真菌与伊朗链霉菌野生型共培养时,四种代表性共培养物的全离子色谱图才显示出新的质量峰,即有新的天然产物被诱导产生。作者在鉴定出了其中一株菌株的诱导产生的物质,基于HPLC-MS,HR-MS和MS / MS分析,这种被诱导产生的物质是卡维林素(carviolin),卡维林素是青霉属真菌产生的红色色素,具有潜在的吸蚕特性(即吸引蚕的特性)。与这一发现一致,真菌分离株27与产linearmycin的细菌分离株45、产30-去甲基裂解二霉素的细菌分离株219的共培养也过表达了卡维林素。
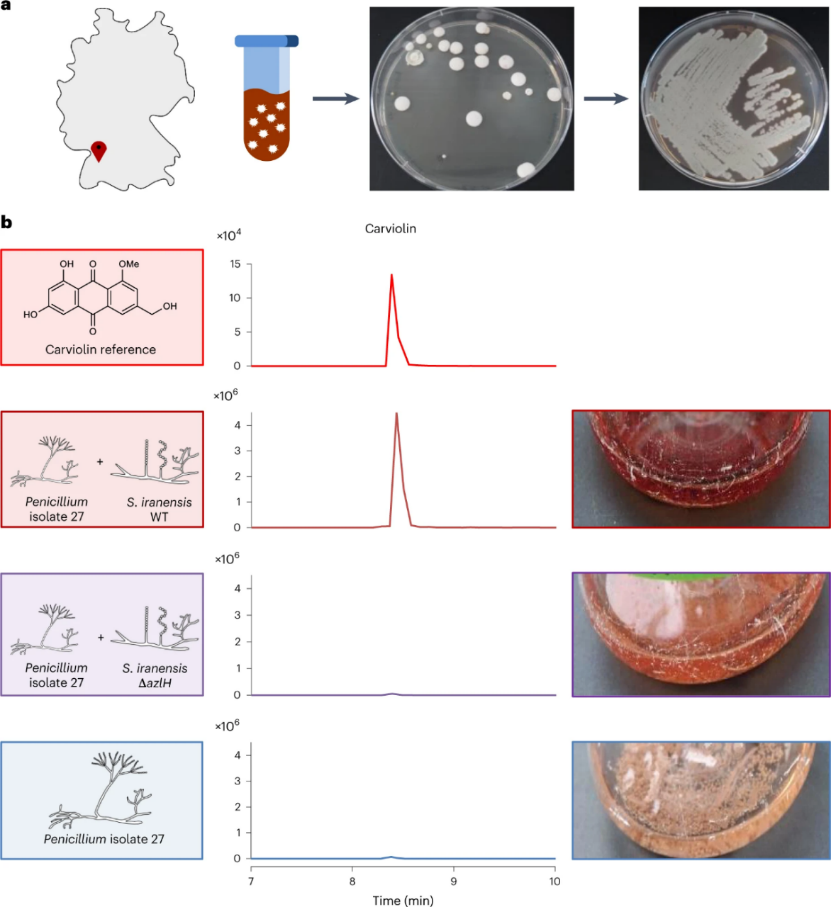
图四 青霉菌27号分离株的分离及产红色色素-卡维林素对阿扎霉素F的反应鉴定
a,德国地图,注明土壤样本的产地(左)和真菌分离的工作流程(右)。b,通过HPLC-MS分析青霉分离株27与伊朗链霉菌野生型、阿扎霉素F缺陷突变株伊朗链霉菌ΔazlH共培养后的提取物以及单独培养的提取物,获得carviolin (m/z 299 [M - H]-)的提取离子色谱图。
结论
总的来说,在产生这些化合物的放线菌的广泛分布以及土壤中真菌响应该化学信号的简易程度表明,精氨酸是塑造微生物群落的微生物通讯网络的通用组成部分。
论文信息
原名:链霉菌聚酮介导土壤中细菌-真菌的跨界互作
译名:Streptomyces polyketides mediate bacteria–fungi interactions across soil environments
期刊:Nature Microbiology
DOI:10.1038/s41564-023-01382-2
发表时间:2023.6.15
通讯作者:Christian Hertweck
通讯作者单位:德国耶拿大学
猜你喜欢
【LorMe周刊】天蓝色链霉菌菌群中潜在利他性成员的突变消融
【LorMe周刊】比较代谢基因组学揭示链霉菌抗生素生产神秘机制
【LorMe周刊】土壤细菌和原生动物互作促进丛枝菌根真菌对有机氮的利用
【LorMe周刊】神秘的专业代谢物推动链霉菌探索并在与其他微生物共生时提供竞争优势
猜你喜欢
iMeta简介 高引文章 高颜值绘图imageGP 网络分析iNAP
iMeta网页工具 代谢组MetOrigin 美吉云乳酸化预测DeepKla
iMeta综述 肠菌菌群 植物菌群 口腔菌群 蛋白质结构预测
10000+:菌群分析 宝宝与猫狗 梅毒狂想曲 提DNA发Nature
一文读懂:宏基因组 寄生虫益处 进化树 必备技能:提问 搜索 Endnote
16S功能预测 PICRUSt FAPROTAX Bugbase Tax4Fun
生物科普: 肠道细菌 人体上的生命 生命大跃进 细胞暗战 人体奥秘
写在后面
为鼓励读者交流快速解决科研困难,我们建立了“宏基因组”讨论群,己有国内外6000+ 科研人员加入。请添加主编微信meta-genomics带你入群,务必备注“姓名-单位-研究方向-职称/年级”。高级职称请注明身份,另有海内外微生物PI群供大佬合作交流。技术问题寻求帮助,首先阅读《如何优雅的提问》学习解决问题思路,仍未解决群内讨论,问题不私聊,帮助同行。
点击阅读原文,跳转最新文章目录阅读


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









