






小肽:调节植物-根际微生物相互作用的新靶点
Small peptides: novel targets for modulating plant–rhizosphere microbe interactions

Review,2024-04-25,Trends in Microbiology,[IF15.9]
第一作者:WeiyiTan(谭唯一)
通讯作者:
TengxiangLian(连腾祥,liantx@scau.edu.cn)
JingJin(金晶,jingjin@scau.edu.cn)
Lam‑SonPhanTran(Son.Tran@ttu.edu)
DOI:
https://doi.org/10.1016/j.tim.2024.03.011
原文链接:
https://www.cell.com/trends/microbiology/abstract/S0966-842X(24)00085-4
- 摘要 -
根际微生物在植物生长和抵御环境压力方面发挥着至关重要的作用,这凸显了微生物与植物之间错综复杂的交流。虽然我们对这些机制的理解仍在不断发展,但植物已配备了一系列促进不同生物界之间交流的信号分子。植物(SPPs)和微生物(SPMs)产生的小肽在细胞内信号传导中发挥着关键作用,对协调植物的各个发育阶段至关重要。在本综述中,我们认为SPPs和SPMs是植物与根圈微生物之间进行双向跨界交流的重要信号媒介。我们探讨了发生这种交流的几种潜在机制途径。此外,我们还提出,受植物-根际微生物相互作用的启发,利用小肽是整体生物工程学领域的一种创新方法。
- 主要内容 -
植物与根际微生物之间的交流
与动物不同,植物是无法移动的,必须迅速适应一系列环境挑战,如极端温度(寒冷和炎热)、干旱、盐胁迫以及食草动物或病原体的攻击。为了应对这些逆境,植物与根际微生物建立了共生关系。这些微生物通过根部渗出物从寄主植物中获取有机和无机物质,包括碳和氮代谢物。作为交换,这些微生物中的许多会给宿主带来益处,包括帮助获取养分、保持水分、防御病原体和合成促进生长的激素。植物的适应性在很大程度上取决于这种共生交流,这种交流也促进了植物和微生物的互利互惠。这种相互依存的关系使我们能够将植物及其相关微生物视为一个整体,其特点是频繁而复杂的互利交流。
植物与微生物的共同进化和全基因组理论支持了植物与微生物之间跨界交流的存在。该理论认为,植物及其相关微生物的基因组是一个整体,作为整个生物体共同发挥作用。植物功能基因可通过微生物的吸收直接影响微生物基因的表达,并通过调节根系形态和渗出物的释放间接改变根圈微生物群的组成。最近,人们越来越关注植物源小肽(SPPs)和微生物源小肽(SPMs)(表1)在促进跨生物界交流方面的作用,特别是它们的传递能力。在本文中,我们提出小肽的传递在植物宿主及其相关的根际微生物之间建立了一个双向、跨界的交流通道。
植物源小肽和微生物源小肽触发的信号通路
环境胁迫可诱导SPPs的产生,SPPs可介导植物对胁迫的适应,并有可能促进植物与根际微生物的交流。SPPs类似于传统的植物激素,通过与特定受体结合,在极低浓度下发挥细胞间信号转导功能。这一信号传递过程包括信号感知、信号转导和后续反应。在植物中,来自前SPPs(Box1)的成熟肽到达靶细胞后,会被特定的质膜定位受体识别,这些受体通常被归类为类受体激酶(RLKs)或类受体蛋白(RLPs),包括富亮氨酸重复类受体激酶(LRR-RLKs)。配体与受体的相互作用会引发构象变化和/或(自身)磷酸化,从而引发一系列细胞内事件。这些事件会激活其他激酶,如丝裂原活化蛋白激酶(MAPKs)和钙依赖蛋白激酶(CPKs),然后使信号通路中的下游元件磷酸化。例如,植物EPIDERMAL PATTERNING FACTOR(EPF)家族通过激活拟南芥中的MAPK通路来增强信号传递,而PHYTOSULFOKINE(PSK)肽则通过启动番茄中的CPK通路来发挥作用。下游元件包括转录因子,它们调节基因表达并决定细胞对SPPs信号的反应。值得注意的是,MAPKs和CPKs有时会表现出协同作用,ROOTMERISTEM GROWTH FACTOR(RGF)肽在激活这两种途径以调节拟南芥与微生物之间的相互作用中的作用就是证明。病原体诱导的RGF7肽激活MAPK通路,并通过感应质膜上的RGF1 INSENSITIVE4/5(RGI4/5)-BRASSINOSTEROID INSENSITIVE 1-ASSOCIATEDKINASE 1(BAK1)/SOMATIC EMBRYOGENESIS RECEPTOR KINASES 4(SERK4)受体复合物,触发细胞外钙离子内流,从而激活CPK。这种对转录因子的协调作用促进了RGF7前体编码基因(preRGF7)的表达,并增强了反馈回路。SPP受体信号通路为细胞应对环境刺激提供了快速有效的途径(图1)。
SPP受体结合激活的信号通路在调节植物与微生物的相互作用中起着关键作用。这些受体将信号从细胞外环境传递到细胞内空间,是细胞通信对外部刺激做出反应的重要组成部分。例如,C-TERMINALLY ENCODED PEPTIDES(CEPs)在根系中发挥着系统性氮需求信号元件的作用。低氮水平或氮饥饿会激活C-TERMINALLY ENCODED PEPTIDE RECEPTOR1(CEPR1)和CEPR2通路。这些信号通路通过上调硝酸盐转运体编码基因,特别是NRT1.1的表达,显著调节水稻中硝酸盐相关基因的表达和代谢谱,从而增强根际氮补充微生物的招募。这些微生物有助于将有机氮转化为植物更容易获得的形式,如硝酸盐和铵,从而提高植物有效利用可用氮资源的能力(图2,关键图)。值得注意的是,单个SPP可与多个受体相互作用,这凸显了其多变的信号能力。例如,在拟南芥中,CLAVATA3(CLV3)肽通过与包括CLV1和CLV2在内的一系列受体结合,表现出很强的杂合性,从而通过miR169/核糖转运因子Y亚基ALPHA(NF-YA)模块在调节植物对青枯雷尔氏菌(Ralstonia solanacearum)的防御反应中发挥了不可或缺的作用。相反,单个受体往往可以被多个SPPs靶向,例如,CLAVAT3/EMBRYO SURROUNDING REGION(CLE)肽MtCLE12和MtCLE13与SUPER NUMERIC NODULES(SUNN)受体相互作用,调节Medicago truncatula结瘤。此外,不同SPP受体对之间的相互作用经常汇聚到共享的下游信号成分上,包括核心受体和MAPK信号级联。这些发现强调了SPPs调节植物与其根际微生物群之间相互作用的多方面途径。
迄今报道的SPMs的序列和功能与SPPs相似,但对其合成过程和分类仍知之甚少。SPMs作为植物信号调节因子,通过与多种受体相互作用来调节植物免疫和激素信号,从而影响植物与微生物之间的相互作用。例如,Colletotrichum tofieldiae产生了一种新型SPM类似物RAPID ALKALINIZATION FACTOR(RALF),称为CtRALF,其结构和功能与RALF SPPs相似。CtRALF直接与拟南芥RLK FERONIA(FER)结合,通过一系列反应增强拟南芥植物的微生物共生能力,包括触发钙波、抑制活性氧(ROS)爆发和增加MAPK磷酸化。由于SPPs和SPMs在与其受体结合后发挥作用,因此鉴定将受体激酶与MAPK和/或CPK信号通路连接起来的分子对于了解小肽在植物-微生物相互作用中的信号作用至关重要。
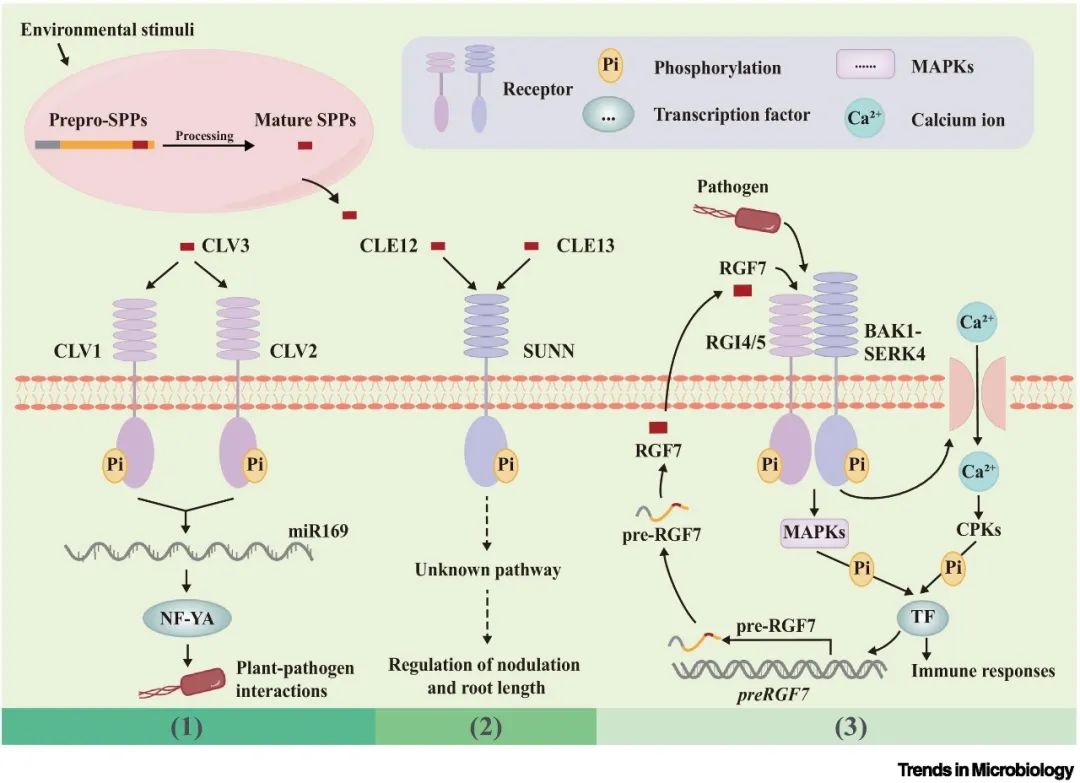
图1 小肽的合成及其传递信号的多种机制。
植物源小肽:促进植物-根际微生物跨界交流
在整个生命周期中,植物依靠一系列复杂的信号途径来协调各种生理过程。这些信号因其流动性和调节功能,在吸引微生物进入植物体内方面至关重要。信号转移的一种机制是分泌性多泡体(MVBs)将SPPs带到细胞外空间,在那里它们与膜融合,并以细胞外囊泡(EVs)的形式释放SPPs。众所周知,EVs参与了跨界的信号传递过程。例如,拟南芥寄主植物可以利用EVs直接向真菌病原体Botrytis cinerea运送信使RNA(mRNA)。进入真菌病原体后,传递的mRNA被翻译成蛋白质,有可能靶向干扰真菌线粒体的活动,从而减少病原体感染。此外,Pečenková和Potocký观察到,在计算共表达和聚类分析中,SPPs和外囊亚基EXO70蛋白共同表达,它们形成了一种新型的双膜结合细胞器,产生不同类型的EVs。虽然这种共表达在统计学上并不显著,但作者将其归因于CLE肽表达的延迟和分析算法的偏差。值得注意的是,已知EVs可促进信号分子在不同植物物种间的高效远距离运输,并在植物与微生物的跨界交流中充当小RNA(sRNA)的新型载体。此外,一些细菌和真菌,如放线菌属(Aggregatibacter actinomycetemcomitans)、日本农杆菌(Bradyrhizobium japonicum)、霍尔德布氏菌(Blumeria hordei)和灰霉病菌(Botrytis cinerea),可以通过膜融合使EVs内化。基于这些发现,我们认为SPPs可通过EVs促进植物与根际微生物之间的交流(图2)。
在广义根际微生物的定义下,有大量文献记载,根瘤菌和枝状菌根真菌(AMFs)依靠SPPs与宿主植物建立共生关系。值得注意的是,尽管调控方式多种多样,但现有的关键调控因子和共享的下游途径在增强各种SPPs的协同效应方面发挥着关键作用。例如,在紫花苜蓿(M. truncatula)中,MtSUNN的参与使MtCLE35能够通过下调miR2111的表达来调节结瘤的积极影响,而MtCLE53则通过降低绞股蓝内酯生物合成相关基因的表达和绞股蓝内酯的水平来调节AMF的定植。此外,SPPs还能决定特定根际微生物的命运,通过影响根系结构与宿主建立共生关系。例如,在番茄中,CEP2-CEPR1模块通过减少吲哚乙酸(IAA)的合成和侧根(LR)的萌发来抑制AMF共生。此外,根系结构的改变会产生不同的微生物附着点,并调节土壤有机物和养分的利用效率,从而重塑根际微生物群落的资源可用性。这些发现共同表明,SPPs通过调节微生物(如根瘤菌和AMF)与其寄主植物之间的共生关系以及影响根系形态,在跨界交流中发挥作用。
最近的研究表明,植物免疫系统与根际微生物的招募有关。优化免疫反应对植物抑制有益微生物的过度繁殖至关重要。然而,过于强大的植物防御能力会阻碍生长并降低自身免疫力,这就凸显了在防御机制和促进生长之间保持微妙平衡的必要性。最近的一项调查发现,PSK RECEPTOR 1(PSKR1)通过减轻水杨酸(SA)介导的免疫反应,帮助拟南芥根际假单胞菌的定殖,同时促进拟南芥中属于生长相关基因本体(GO)术语群的基因的表达,尤其是与光合作用和糖运输相关的基因。此外,在磷酸盐(Pi)缺乏的条件下,磷酸盐饥饿反应1(PHR1)介导的RALF23-FER通路会调节FLAGELLIN-SENSING2(FLS2)和BAK1受体复合物的形成以及植物根部次生代谢物的生物合成。这种调节机制降低了植物的免疫力,从而将黄杆菌、假单胞菌和芽孢杆菌等有益微生物招募到根际。这一过程不仅能促进韧皮部的生长,还能扩大钙吸收相关基因的表达,从而缓解钙饥饿。然而,通过产生ROS,RALF23-FER复合物也被确定能协调调节根际中的有益假单胞菌,这与它在标准条件下组装模式识别受体(PRR)复合物的预期免疫作用不同。这些发现凸显了SPPs通过调节植物免疫在招募根际微生物方面的关键作用。
一些SPPs还能以根系分泌物的形式释放到根际,从而可能影响根际微生物的数量和群落结构。对苜蓿根培养物渗出物进行的肽组分析发现了多种SPPs,包括CEPs、CLEs和XYLEM SAP ASSOCIATED PEPTIDEs(XAPs)。其中,MtCEP肽能调节LR数量和结核共生,CLE家族的MtCLE5和MtCLE17肽对主根生长有反作用。已知MtXAP1和MtXAP5肽可抑制酪氨酸硫酸化后的LR发育。与传统植物激素的作用模式类似,SPPs根据其功能被概念化为类似植物激素的信号分子。植物以根渗出物形式释放的植物激素在根际微生物招募中的作用已得到广泛认可,这表明SPPs通过根渗出物在植物-根际微生物相互作用中发挥着关键作用。然而,要阐明根渗出物中的SPPs是否能特异性地招募根际中的有益微生物或病原微生物,还需要对相关分子机制有更深入的了解(图2)。
被称为pri-miRNA的microRNA初级转录本含有能够编码肽的短开放阅读框(ORF)序列。以这种方式获得的肽被称为microRNA编码肽(miPEPs),标志着SPP研究领域的重大进展。其应用的一个例子是使用合成的miPEP172c来促进大豆(Glycine max)结节的生长,说明了SPPs可以调节植物与微生物相互作用的新机制。这种方法对改变作物根际微生物的结构和组成具有潜在的意义。
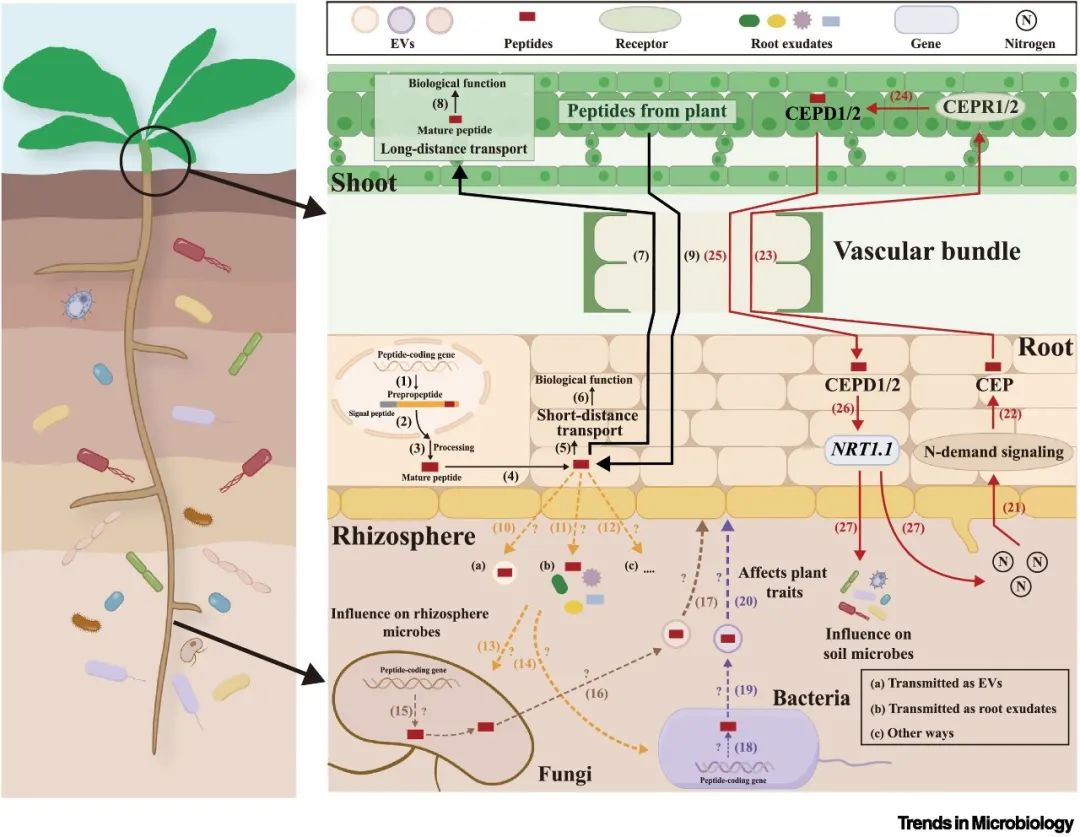
图2 SPPs在植物体内的信号传递过程并影响根际微生物的群落结构。该图还包括小肽(SPPS和SPMs)参与植物-微生物跨界信号交流的可能途径。
微生物源小肽:促进根际微生物与植物的跨界交流
SPM通过与植物受体结合并诱导防御相关基因的表达来激活植物免疫。例如,生物营养型病原体黄单胞菌(Xanthomonas oryzae pv. oryzae,Xoo)会产生一种硫酸化的SPM,称为RaxX,其中含有硫酸化的酪氨酸,并以含硫酸化酪氨酸的植物肽(PSYs)(SPPs的硫酸化酪氨酸形式)命名为MSY。RaxX的硫酸盐衍生物(RaxX13sY)MSY在拟南芥和水稻中诱导根系生长的方式类似于PSYs。此外,RaxX还是PSY的分子模拟物,可促进Xoo感染。在真菌中也观察到类似的现象,真菌Fusarium oxysporum产生一种类似植物RALF的肽,命名为F-RALF,这种肽有助于病原体在宿主体内定殖并增强其在番茄植株中的毒力。与植物RALF肽一样,F-RALF肽也能显著抑制番茄和拟南芥幼苗根部和根毛的生长。禾谷镰刀菌等其他镰刀菌也具有编码RALF的基因,可能会促进病原体在小麦(Triticum aestivum)上的定殖。这些研究结果表明,植物的健康也与其根瘤微生物群中高水平的SPMs编码基因有关。
SPMs参与微调宿主植物体内生长与防御机制之间的平衡,例如PSKR1通过SPMs调节生长与防御之间的权衡。具体来说,假单胞菌被推测具有结构特异性SPM同源物,能增强植物根部PSKR1基因的表达,这是维持植物生长与防御之间复杂平衡的关键机制。在拟南芥中,共生细菌具有编码大量鞭毛蛋白(flg22)型多肽的能力。通过与植物细胞表面的FLS2受体及其共受体BAK1相互作用,植物可以识别这些flg22肽并做出反应。虽然大多数flg22肽都能逃过植物免疫系统的检测,但也有一部分flg22肽能够通过拮抗作用和信号调节改变免疫反应,增强微生物群落的多样性并维持微生物的平衡。
微生物中也存在EVs,植物和细菌的EVs具有相似性或同源性。细菌的EV是由外膜的外部出芽形成的,而真核生物的EV则是由MVB介导的外泌体形成的。微生物EV很可能在将SPM运送到植物体内、影响宿主免疫基因方面发挥作用。这表明,在EVs的介导下,通过SPMs进行跨界信号传递有了新的途径(图2)。
小肽的工程与应用
植物保持抗逆性和耐压性的能力非常复杂,不断变化的外部环境仍然是当前研究的一个重要课题。在微生物工程领域,由于对植物-根际微生物相互作用的深入研究,创新农业应用的新机会正在出现。然而,在实际田间应用中,仅仅在农业中应用有益微生物,而忽视有益微生物与其寄主植物之间的联系,往往无法长期持续下去。大量研究发现了多种信号分子参与植物-根际微生物相互作用的信号转导过程。生物化学、蛋白质组学、遗传学和生物信息学方面的最新进展与质谱分析相结合,极大地促进了肽组学的快速发展。这一进展阐明了SPPs和SPMs在调节植物-根际微生物相互作用中的作用,为小肽工程在全生物体中的应用提供了新的视角。
合成小肽的应用可以直接调节植物的生长过程,如免疫反应和养分调节,也可以通过塑造根际微生物间接改变植物的性状,从而直接或间接地改变植物的性状。关于小肽工程及其在植物与微生物相互作用中的应用,已有多篇报道。例如,将小肽与表面活性剂结合对茶叶(Camellia sinensis(L.)Kuntze)叶层微生物群的群落结构有显著影响。此外,还有报道称发现miPEPs可抵抗真菌病原体,从而提高植物的抗病性。最近的研究结果表明,参与植物与微生物相互作用的SPPs可以螯合血液中的游离血红蛋白,从而有助于治疗由需要血红蛋白才能存活的细菌或寄生虫引起的疾病。然而,这些发现只是冰山一角,还有更多令人兴奋的小肽应用和相关机制有待发现。
值得注意的是,微生物天然产生水解小肽所需的酶。因此,微生物基因编辑可能是合成小肽的一种潜在方法。此外,与其他方法相比,微生物培养需要的营养更少,生长期更短,因此是一种在短时间内生产大量SPPs的经济有效的方法。例如,一种经过基因工程改造的枯草杆菌菌株被用于体外分泌RALF23。这些都表明,微生物可以成为前景广阔的小肽体外合成工厂,包括SPPs和SPMs(图3)。
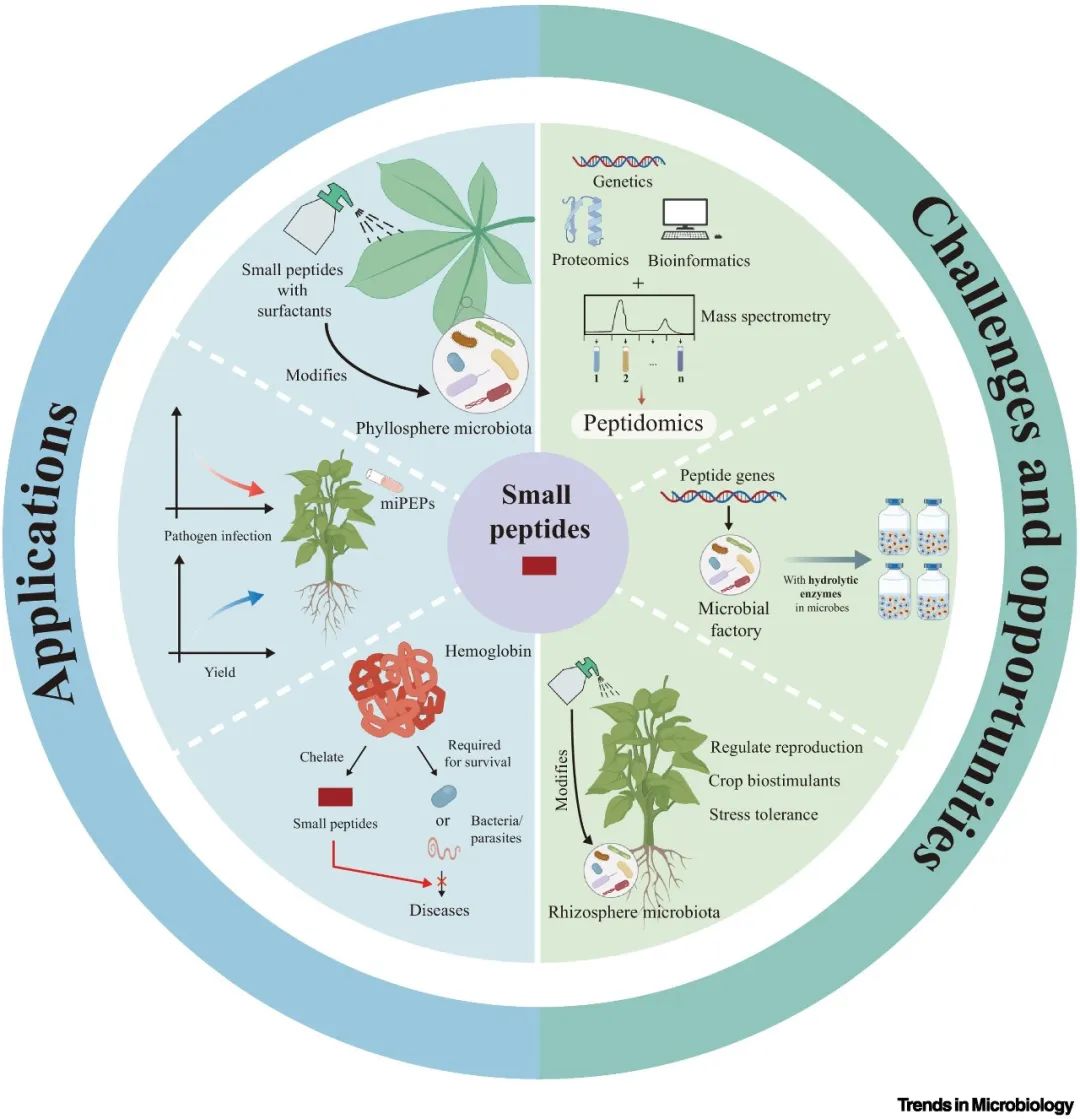
图3 以小肽为基础的肽工程的实际应用及肽工程面临的机遇和挑战。
- 总结及展望 -
鉴于SPPs和SPMs可以进入EV,然后与微生物相互作用的发现,并结合目前对SPPs在植物组织中的流动性及其通过配体-受体相互作用的作用机制的了解,我们认为根际微生物群落的形成部分是由这些小肽介导的。这一观点意味着,在全生物体内参与植物-根瘤微生物相互作用调控的SPPs和SPMs在农业和林业领域具有相当大的应用潜力。虽然研究SPPs和SPMs的各种工具和方法(包括高通量筛选和生物信息学)已经开发出来,但我们对其功能的了解仍不全面。如果我们关于植物与根际微生物之间由SPPs和SPMs介导的双向信号交流的假设被证明是正确的,那么这将为植物与根际微生物之间的相互作用与全生物工程学之间的联系提供一种新的途径。这将产生一系列新的问题有待探索,以开发和描述涉及SPPs和SPMs的各种调控过程。
- 作者简介 -
华南农业大学农学院硕士研究生谭唯一为论文第一作者,国家大豆改良中心广东分中心连腾祥副教授、华南农业大学农学院/岭南现代农业科学与技术广东省实验室金晶副教授和德克萨斯理工大学Lam‑Son Phan Tran教授为论文共同通讯作者,本研究得到国家自然科学基金(批准号:32170115)、国家重点研发(批准号:2022YFF1001800),双一流学科推进项目(批准号:2021B10564001),广州市科技创新发展基金(批准号:2024A04J5487)和广东省自然科学基金(批准号:2023A1515030275)的资助。
通讯作者

华南农业大学
连腾祥
博士、副教授、硕士生导师
目前主要以大豆为研究对象,整合微生物组学、代谢组学、分子生物学和遗传学等手段和技术体系,揭示大豆-微生物-环境互作分子机制,研发大豆抗逆高产相关技术。主持国家自然科学基金、国家重点研发计划及广东省广州市等省部级项目10余项。在Trends in Microbiology, Plant Biotechnology Journal,ISME Communications,Soil Biology & Biochemistry, mSystems, Applied and Environmental Microbiology, Biology Fertil Soils, Microbiology Spectrum, Journal of Hazardous Materials, Plant Physiology and Biochemistry, Journal of Integrative Agriculture, Biochar, Applied Soil Ecology, 作物学报、华南农业大学学报等国内外专业期刊上发表论文30余篇,授权发明专利2项。担任European Journal of Soil Biology,Frontiers in Microbiology,土壤与作物编委;iMeta和广东农业科学青年编委。
ORCID主页:https://orcid.org/0000-0003-1131-2491

华南农业大学农学院
金晶
博士、副教授、硕士生导师
博士毕业于中国农业大学,现就职于华南农业大学农学院,主要从事野生稻优异基因的挖掘与利用及植物-微生物互作等方面的研究。主持多项国家自然科学基金、广东省自然科学基金项目。以第一作者或通讯作者在The Plant Cell、Molecular Plant、Plant Physiology、mSystems等国际权威期刊发表SCI论文。

德克萨斯理工大学
农业科学与自然资源戴维斯学院
Lam‑Son Phan Tran
博士、副教授、硕士生导师
生物科学和生物技术领域的专家,于1994年在匈牙利圣伊斯特万大学获得生物技术硕士学位,并于1997年获得生物科学博士学位。从2020年9月开始,加入德克萨斯理工大学农业科学与自然资源戴维斯学院最新的研究机构-作物非生物抗逆性基因组学研究所,担任德克萨斯理工大学植物与土壤科学系教授。
宏基因组推荐
本公众号现全面开放投稿,希望文章作者讲出自己的科研故事,分享论文的精华与亮点。投稿请联系小编(微信号:yongxinliu 或 meta-genomics)
猜你喜欢
iMeta高引文章 fastp 复杂热图 ggtree 绘图imageGP 网络iNAP
iMeta网页工具 代谢组MetOrigin 美吉云乳酸化预测DeepKla
iMeta综述 肠菌菌群 植物菌群 口腔菌群 蛋白质结构预测
10000+:菌群分析 宝宝与猫狗 梅毒狂想曲 提DNA发Nature
一文读懂:宏基因组 寄生虫益处 进化树 必备技能:提问 搜索 Endnote
16S功能预测 PICRUSt FAPROTAX Bugbase Tax4Fun
生物科普: 肠道细菌 人体上的生命 生命大跃进 细胞暗战 人体奥秘
写在后面
为鼓励读者交流快速解决科研困难,我们建立了“宏基因组”讨论群,己有国内外6000+ 科研人员加入。请添加主编微信meta-genomics带你入群,务必备注“姓名-单位-研究方向-职称/年级”。高级职称请注明身份,另有海内外微生物PI群供大佬合作交流。技术问题寻求帮助,首先阅读《如何优雅的提问》学习解决问题思路,仍未解决群内讨论,问题不私聊,帮助同行。
点击阅读原文,跳转最新文章目录阅读


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









