






点击蓝字 关注我们
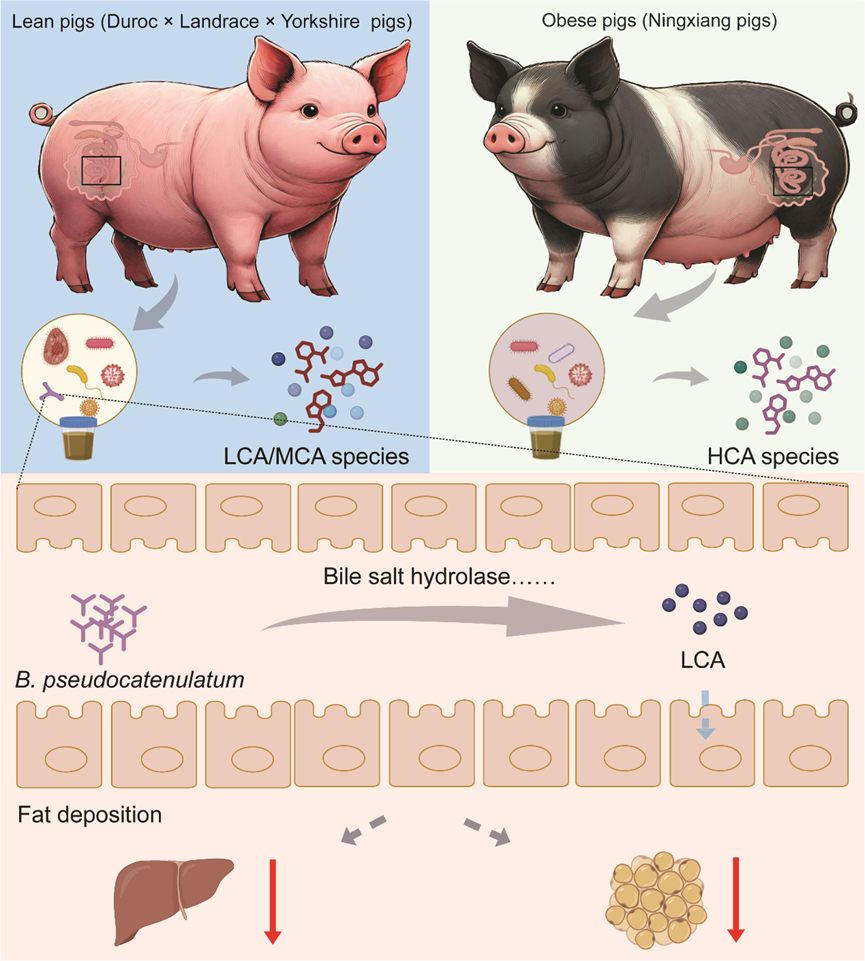
肠道假小链双歧杆菌通过增强次级胆汁酸生物合成降低脂肪沉积

iMeta主页:http://www.imeta.science
研究论文
● 原文: iMeta (IF 23.8)
● 原文链接DOI: https://doi.org/10.1002/imt2.261
● 2024年12月30日,湖南农业大学谭碧娥、王婧和印遇龙院士团队等在iMeta在线发表了题为“Gut Bifidobacterium pseudocatenulatum protects against fat deposition by enhancing secondary bile acid biosynthesis”的文章。
● 本研究采用16s rDNA测序、宏基因组和代谢组技术,系统评估了瘦肉型猪和脂肪型猪的肠道微生物组成和功能差异,并通过高脂饲喂小鼠模型和宁乡猪验证了假小链双歧杆菌通过影响次级胆汁酸生物合成影响宿主的脂肪沉积,揭示了假小链双歧杆菌降低脂肪沉积的分子机制,并提示胆盐水解酶是预防人类和动物脂肪过度沉积的潜在靶点。
● 第一作者:查安东
● 通讯作者:谭碧娥(bietan@hunau.edu.cn)、 印遇龙(yinyulong@isa.ac.cn)、王婧(jingwang023@hunau.edu.cn)
● 合作作者:齐鸣、邓远坤、李浩、王楠、王城名、廖思梦、万丹、熊霞、廖鹏
● 主要单位:湖南农业大学动物科技学院、岳麓山实验室、中南大学基础医学院、中国科学院亚热带农业生态研究所
亮 点
● 宏基因组和代谢组揭示瘦肉型猪和脂肪型猪的肠道微生物组成和功能存在显著差异,并利用结肠菌群移植试验证明不同品种的结肠菌群差异调节宿主脂肪沉积;
● 肠道中的假小链双歧杆菌通过促进次级胆汁酸的生物合成来减少机体过度脂肪沉积;
● 膳食添加石胆酸也有助于减少脂肪沉积。
摘 要
肠道微生物在调控人类和动物脂质代谢中起着关键作用,但特定微生物及其代谢产物如何影响脂肪沉积尚不清楚。本研究通过结肠菌群移植试验发现,瘦肉型猪和脂肪型猪的结肠菌群对脂肪沉积的影响显著不同。宏基因组分析显示,瘦肉型猪结肠假小链双歧杆菌丰度较高,且与低背膘厚正相关。同时,瘦肉型猪中石胆酸等微生物代谢产物显著富集,并与假小链双歧杆菌丰度呈正相关。在HFD喂养的小鼠中,补充活假小链双歧杆菌显著减少脂肪沉积,并促进结肠次级胆汁酸的生物合成。然而,抑制胆盐水解酶会削弱假小链双歧杆菌的抗脂肪沉积作用。此外,饮食添加石胆酸也显著减少了HFD饲喂大鼠和脂肪型猪的脂肪沉积。这些发现揭示了假小链双歧杆菌降低脂肪沉积的分子机制,并提示胆盐水解酶可以作为预防肥胖的重要潜在靶点,为调节人类和动物脂肪沉积提供了新思路。
视频解读
Bilibili:https://www.bilibili.com/video/BV1Jt6GYyEDE/
Youtube:https://youtu.be/pZQN2mIADtU
中文翻译、PPT、中/英文视频解读等扩展资料下载
请访问期刊官网:http://www.imeta.science/
全文解读
引 言
过多的脂肪沉积容易导致代谢综合征的发生,包括肥胖、2型糖尿病、非酒精性脂肪肝和心血管疾病等。在畜牧业中,脂肪沉积过多还会对饲料转化率和生产效率产生负面影响。近年来的研究强调了肠道微生物群在能量稳态和脂肪沉积中的关键作用。肠道微生物及其相关代谢产物干预可能成为减少脂肪沉积的一种有前景的策略。
肠道微生物的相对丰度会因饮食、地理位置和生理状态等因素而显著变化。作为能量稳态重要调节因子的双歧杆菌在肥胖个体中的丰度明显低于健康个体。已有研究表明,假小链双歧杆菌C95株定殖可显著降低体重、脂肪重量、空腹血糖和胰岛素抵抗,并改善餐后血糖反应。此外,假小链双歧杆菌C95株接种还通过调节包括胆汁酸(BAs)在内的代谢产物,降低了用来自2型糖尿病患者的粪便微生物群定植的无菌小鼠的空腹血糖。但是,关于双歧杆菌如何影响宿主脂肪沉积的机制还不清楚。已有研究表明,由共生菌产生的胆汁酸在调节宿主能量稳态中起着关键作用。例如,猪胆酸(HCA)已被鉴定为代谢紊乱的生物标志物,而微生物衍生的去氧胆酸(HDCA)通过法尼酰X受体(FXR)和过氧化物酶体增殖物激活受体α(PPARα)缓解非酒精性脂肪肝。此外,研究发现双歧杆菌与次级胆汁酸代谢密切相关,这表明次级胆汁酸可能在双歧杆菌介导的效应中发挥关键作用。
猪的消化生理和杂食性进食行为与人类相似。因此,以宁乡猪(宁乡猪)为代表的地方猪种(以背膘厚著称)是研究脂肪沉积和肥胖的理想模型。在本研究中,我们通过整合宏基因组测序和结肠代谢组学,全面分析了瘦肉型猪和脂肪型猪在微生物组成和代谢功能的差异,以期筛选影响脂肪沉积的关键菌株或关键代谢途径,为减少人类和动物脂肪沉积提供潜在的治疗策略。
结 果
瘦肉型猪和脂肪型猪的肠道微生物与背膘厚度显著相关
本研究发现与瘦肉型猪(杜长大猪)相比,脂肪型猪(宁乡猪)的背膘厚度和皮下脂肪组织中的脂肪细胞大小明显增大(图 1A、B)。猪的后肠微生物群对能量稳态和脂肪沉积至关重要,且主要富集在结肠中。我们收集了结肠食糜进行16S rDNA 测序,结果显示宁乡猪的结肠微生物群多样性和丰度在统计学上显著高于杜长大猪(图 1C)。主坐标分析显示,杜长大猪和宁乡猪的结肠微生物群结构存在显著差异(图 1D)。为了探究结肠微生物群在脂肪沉积中的作用,我们使用Mantel检验将微生物群落组成的差异与背膘厚度进行了相关性分析,结果显示二者之间存在显著相关(r = 0.1492,p < 0.001,图 1E)。通过PICRUST2分析发现,脂质代谢途径在宁乡猪的结肠微生物群中显著富集(图 1F)。此外,与次级胆汁酸合成、初级胆汁酸合成、甘油三脂代谢、不饱和脂肪酸合成、酮体合成与降解、甘油磷脂代谢和亚油酸代谢相关的途径也在宁乡猪中富集(图 S1)。这些结果表明,结肠微生物群的组成可能在调节猪的脂肪沉积中发挥重要作用。
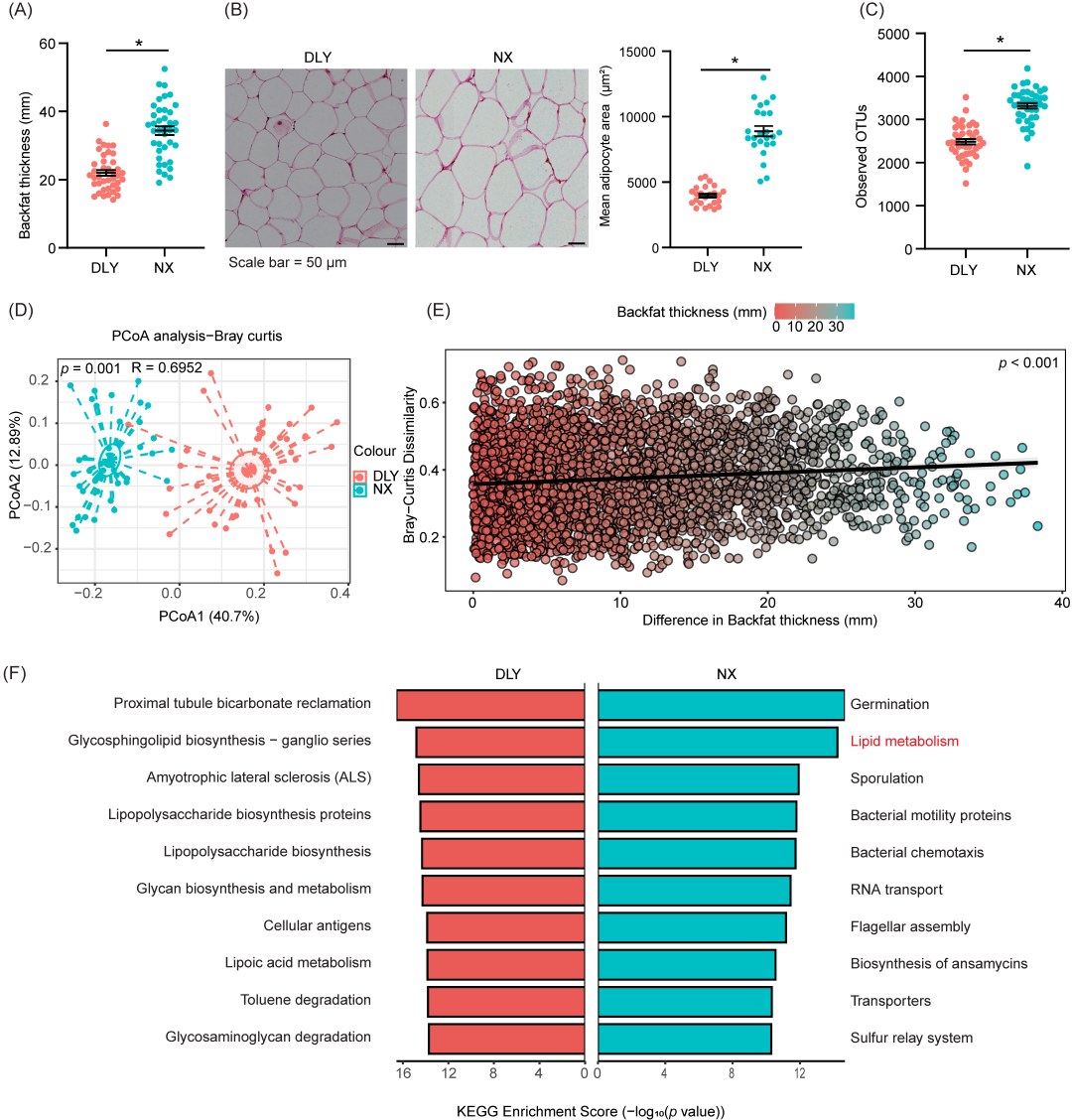
图1. 瘦肉型猪和脂肪型猪的肠道微生物与背膘厚度显著相关
(A)背膘厚度;(B)皮下脂肪组织的苏木精-伊红(H&E)染色图像;(C)结肠微生物α多样性;(D)基于Bray-Curtis距离的结肠微生物主坐标分析;(E)肠道微生物组与背膘厚度的相关性;(F)PICRUSt 2.0分析。
瘦肉型猪和脂肪型猪的结肠微生物群影响脂肪沉积
为了探讨猪结肠微生物群对脂肪沉积的影响,我们将杜长大猪和宁乡猪的结肠微生物群分别移植到7-8周龄的雄性C57BL/6J小鼠体内(图2A)。移植品种特异性结肠微生物群未对饲喂常规饮食的小鼠的体重变化或累计食物摄入量产生影响(图2B、C)。然而,移植宁乡猪的结肠微生物群显著增加了饲喂常规饮食小鼠的肾周白色脂肪组织(pWAT)的重量(图2D),而对皮下白色脂肪组织(sWAT)和附睾白色脂肪组织(eWAT)的重量和平均脂肪细胞面积没有显著影响(图2E、F)。此外,移植不同品种猪结肠菌群对饲喂常规饮食的小鼠在血清脂质谱,包括总胆固醇、葡萄糖、甘油三酯、高密度脂蛋白胆固醇(HDLC)、低密度脂蛋白胆固醇(LDLC)、肝脏重量和肝脏脂肪变性等方面没有显著影响(图S2A-C)。移植宁乡猪结肠微生物群的小鼠肝脏中Fxr和Takeda G蛋白偶联受体5(Tgr5)的mRNA表达显著低于移植杜长大猪结肠微生物群的小鼠(图S2D)。
在高脂饮食(HFD)饲喂的小鼠中,移植宁乡猪的结肠微生物群显著增加了体重和累计食物摄入量(图2G、H)。这些小鼠的sWAT、eWAT、pWAT、和WAT重量均显著增加,同时eWAT和sWAT的脂肪细胞平均面积也显著增大(图2I-K)。宁乡猪结肠微生物群移植的小鼠血清中葡萄糖、甘油三酯、总胆固醇、HDLC、LDLC和非酯化脂肪酸(NEFA)水平均显著增加(图S2E)。而在接受宁乡猪菌群小鼠的血清中,参与脂肪吸收的重要代谢产物——胆汁酸(BAs)的水平显著降低(图S2E)。HFD已知会诱导肝脏损伤,移植自宁乡猪的结肠微生物群显著增加了HFD喂养小鼠的肝脏重量和肝脏脂肪变性评分(图S2F、G)。在转录水平上,宁乡猪结肠微生物移植小鼠的肝脏中,参与脂肪生成的基因(如乙酰辅酶A羧化酶(Acc)、脂肪酸合酶(Fasn)、Pparγ和类固醇调节元件结合蛋白1(Srebp1))的mRNA表达显著上调(图S2H)。
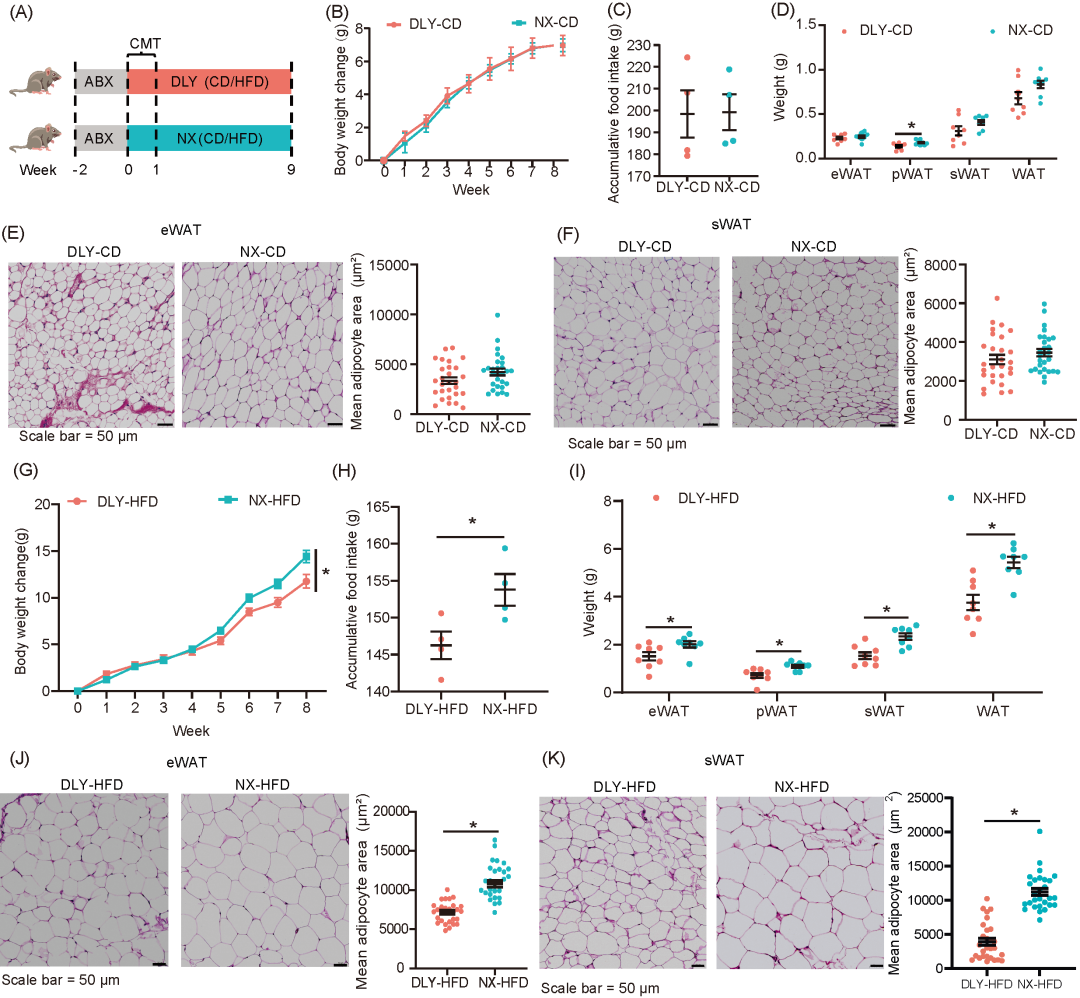
图2. 瘦肉型猪和脂肪型猪的肠道微生物群对脂质沉积变化的影响
(A)结肠微生物群移植实验示意图;(B)体重变化;(C)累计食物摄入量;(D)器官重量;(E)附睾白色脂肪的H&E染色代表性图像,以及平均脂肪细胞面积的定量分析;(F)皮下脂肪的H&E染色代表性图像,以及平均脂肪细胞面积的定量分析;(G) 体重变化;(H) 累计食物摄入量;(I)脂肪组织重量;(J)附睾白色脂肪的H&E染色代表性图像,以及平均脂肪细胞面积的定量分析;(K)皮下脂肪的H&E染色代表性图像,以及平均脂肪细胞面积的定量分析。DLY-CD:接受来自杜长大猪结肠微生物群且用普通饲料喂养的小鼠;NX-CD:接受来自 宁乡猪结肠微生物群且用普通饲料喂养的小鼠;DLY-HFD:接受来自杜长大猪结肠微生物群且用高脂饲料喂养的小鼠;NX-HFD:接受来自宁乡猪结肠微生物群且用高脂饲料喂养的小鼠。
结肠特定微生物调控瘦肉型猪和脂肪型猪间次级胆汁酸代谢的差异
为了进一步确定调控背膘厚度的特定肠道微生物,我们对50个结肠食糜样本进行了高通量宏基因组测序。结果鉴定出559种细菌和12种古菌,其中宁乡猪的结肠微生物群中细菌种类的数量显著较多。9种细菌在杜长大猪中显著富集,而10种细菌则在宁乡猪中较为常见(图3A)。使用randomForest包,我们构建了一个稀疏随机森林模型,突出了12种细菌物种,例如假小链双歧杆菌,它们作为背膘厚度的重要预测因子,能够解释59.71%的方差(图3B)。接下来,Spearman相关分析确定了与背膘厚度相关的前20种细菌,包括假小链双歧杆菌(图3C)。通过整合差异分析、随机森林和Spearman分析,假小链双歧杆菌和 Akkermansia glycaniphila 等12种菌被识别为背膘厚度的统计学显著相关因子和关键生物标志物(图3D)。此外,线性判别分析效应量分析(LEfSe)也揭示了假小链双歧杆菌在杜长大猪中的富集(图S3)。综合来看,这些发现表明假小链双歧杆菌 和 A. glycaniphila 是调控背膘厚度变化的关键微生物。
为了阐明杜长大猪和宁乡猪结肠微生物群的功能差异,我们采用HUMAnN2进行功能预测。分析结果显示,宁乡猪结肠微生物群中与脂肪酸生物合成、次级胆汁酸生物合成及其他多个代谢功能相关的途径显著富集。相反,与错配修复、同源重组和核苷酸切除修复相关的途径则在杜长大猪中显著富集(图S4A)。随后进行的代谢组学分析鉴定出147种结肠代谢物,显示宁乡猪和杜长大猪之间的代谢物谱存在明显差异(图3E)。其中,16种代谢物在杜长大猪和宁乡猪之间差异显著(FDR<0.05,Log2 FC > 1.32 或 < -1.32,图3F)。猪脱氧胆酸和石胆酸(LCA)是两种主要的胆汁酸,在两种猪的结肠内容物中均有分布,但其分布模式不同。LCA类胆酸和鼠胆酸类胆酸在杜长大猪的结肠中显著富集,而HCA类胆酸在宁乡猪中显著富集(图S4B)。在杜长大猪中,LCA类胆酸,包括LCA、iso-LCA和GLCA均显著富集,而HCA类胆酸在宁乡猪中更为丰富(图S4C)。进一步分析发现LCA和GLCA的水平与背膘厚度呈负相关,提示这些胆汁酸浓度较高可能有助于减少脂肪沉积(图S4D)。为了识别特定的微生物对LCA生成的影响,我们进行了胆汁酸与细菌物种的Spearman相关性分析。结果显示,LCA及其衍生物与结肠中假小链双歧杆菌的相对丰度显著正相关(图S4E)。综合以上结果,我们提出假小链双歧杆菌通过调节次级胆汁酸代谢,特别是LCA的生成,发挥预防脂肪沉积的作用。
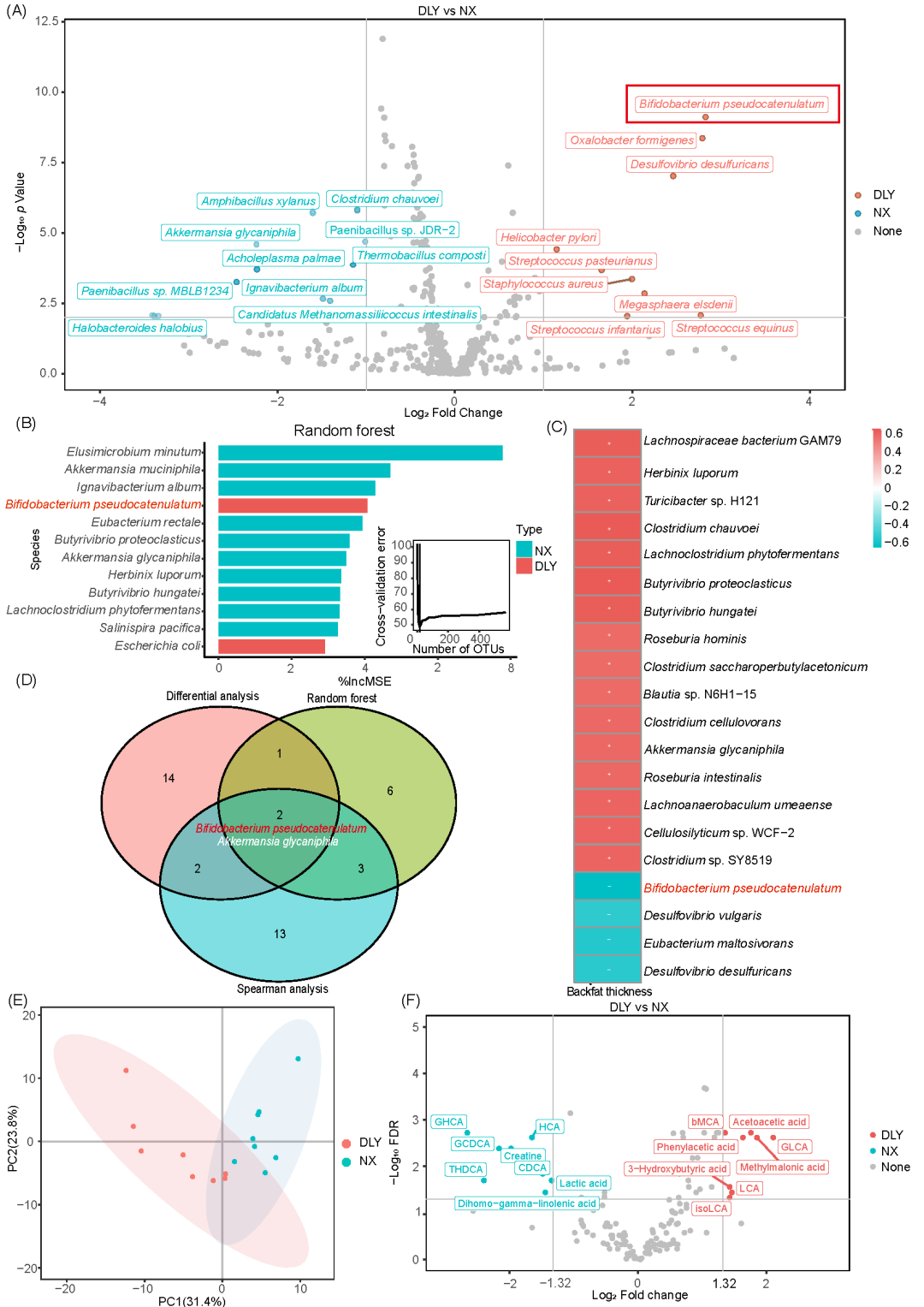
图3. 结肠特定微生物调控瘦肉型猪和脂肪型猪间次级胆汁酸代谢的差异
(A)菌种差异分析;(B)随机森林分析;(C)细菌种类与背膘厚度的Spearman相关分析;(D)差异分析、随机森林分析和斯皮尔曼分析的维恩图;(E)结肠代谢组学的主成分分析;(F)代谢物差异分析。
假小链双歧杆菌减少高脂饲喂小鼠过度脂肪沉积并增强次级胆汁酸生物合成
为了阐明假小链双歧杆菌在脂肪沉积中的作用,我们将活假小链双歧杆菌、热灭活假小链双歧杆菌、假小链双歧杆菌培养上清液和基础培养基(BBL培养基)分别给HFD小鼠灌胃9周(图4A)。通过16S rDNA定量检测确认了假小链双歧杆菌的定植(图4B)。只有活假小链双歧杆菌显著降低了HFD小鼠的体重变化(图4C、D,S5A)。活假小链双歧杆菌灌胃还显著减少了WAT总量,同时减小了sWAT和eWAT的脂肪细胞平均大小(图4E、F)。与HFD饲喂小鼠相比,活假小链双歧杆菌治疗对Fxr和Tgr5的表达没有影响(图4G)。此外,活假小链双歧杆菌还降低了肝脏重量,缓解了肝脏脂肪变性,血清丙氨酸氨基转移酶(ALT)水平下降,表明其对HFD诱导的肝脏损伤具有保护作用(图S5B-E)。与HFD小鼠相比,活假小链双歧杆菌治疗和正常饮食处理增加了肝脏中Fxr mRNA的表达(图S5F)。
为了评估对次级胆汁酸代谢的影响,我们对结肠内容物进行了超高效液相色谱-质谱分析。结果表明,与正常饮食组相比,HFD显著增加了结肠中的CA和CDCA浓度,但减少了LCA、epi LCA、HDCA和3-keto-5β-胆酸(3-k-5β-CA)的浓度。活假小链双歧杆菌灌胃显著提高了HFD小鼠结肠中LCA、3-k-5β-CA、3β-熊脱氧胆酸(3-β-UDCA)和HDCA的浓度(图4H)。
进一步的实验在抗生素处理的HFD小鼠中进行,以确定假小链双歧杆菌的抗脂肪沉积作用是否与肠道微生物群无关。假小链双歧杆菌持续减少了伪无菌HFD小鼠的体重增加、脂肪质量、肝脏重量和肝脏甘油三酯浓度(图S6)。这些结果表明,假小链双歧杆菌介导的降脂肪沉积作用可能与肠道微生物群无关。
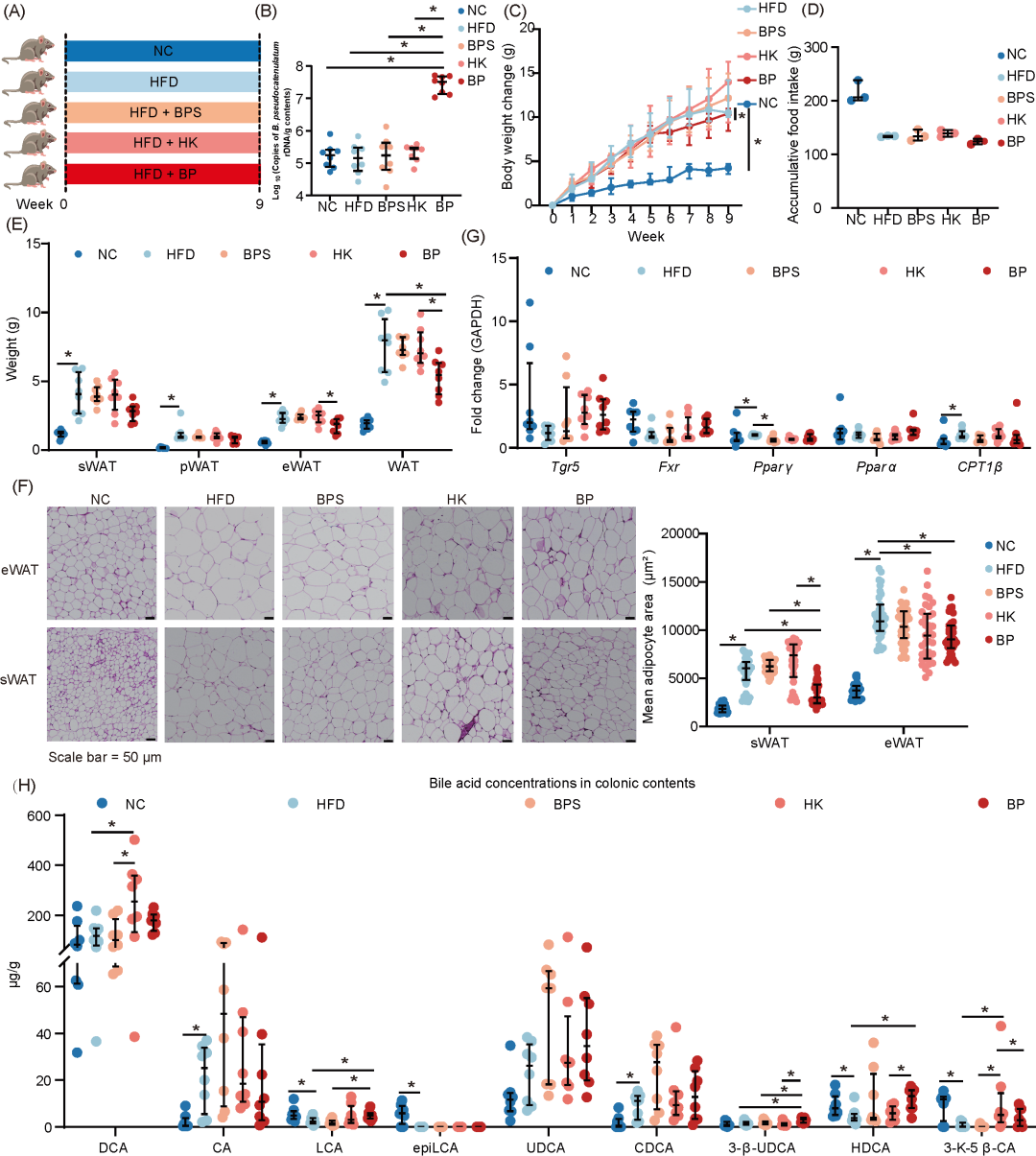
图4. 假小链双歧杆菌可降低高脂饮食喂养小鼠的过度脂肪沉积并增强次级胆汁酸的生物合成
(A)试验示意图;(B)结肠内容物中假链状双歧杆菌 16S rDNA 的拷贝数;(C)体重变化;(D)累计食物摄入量;(E)脂肪组织重量;(F)附睾白色脂肪和皮下脂肪的H&E染色代表性图像,以及平均脂肪细胞面积的定量分析;(G)附睾白色脂肪中脂质代谢相关 mRNA 的表达;(H)结肠内容物中胆汁酸浓度。NC 组:普通饲料 + BBL培养基;HFD 组:HFD + BBL培养基;BPS 组:HFD + 假链状双歧杆菌上清液;HK 组:HFD + 热灭活的假链状双歧杆菌;BP 组:HFD + 活假链状双歧杆菌。
胆汁盐水解酶的药理学抑制削弱了假小链双歧杆菌的抗脂肪沉积作用
通过使用Proksee预测基因组功能,发现假小链双歧杆菌基因组中含有一个胆氨酸水解酶(cgh)基因,该基因与胆汁酸代谢相关(图S7A)。胆氨酸水解酶即胆盐水解酶(BSH),催化结合胆汁酸转化为游离胆汁酸。为了确定假小链双歧杆菌对脂肪沉积的影响是否通过BSH活性介导,我们在HFD小鼠模型中使用了肠道限制性BSH抑制剂GR7(图5A)。假小链双歧杆菌和GR7联合给药的结果表明,GR7显著抑制了次级胆汁酸的生物合成(图5B)。GR7治疗显著减少了假小链双歧杆菌的定植并降低了其对体重的影响(图5C、D)。尽管GR7治疗没有显著影响脂肪组织的重量,但它增加了sWAT和eWAT的脂肪细胞平均大小(图5E、F)。此外,GR7处理还导致血清葡萄糖和甘油三酯浓度的升高(图S7B)。与此同时,GR7也加剧了肝脏损伤,表现在血清AST和ALT水平升高,以及肝脏甘油三酯含量增加(图S7C、D)。
为了进一步探讨可能驱动脂质代谢变化的因素,我们测量了Fxr、Pparγ和Tgr5的转录水平。GR7的使用显著下调了肝脏Fxr的表达,并上调了eWAT中Fxr和Pparγ的表达(图5H,S7E)。这些结果共同表明,BSH活性在脂肪沉积中的重要作用。
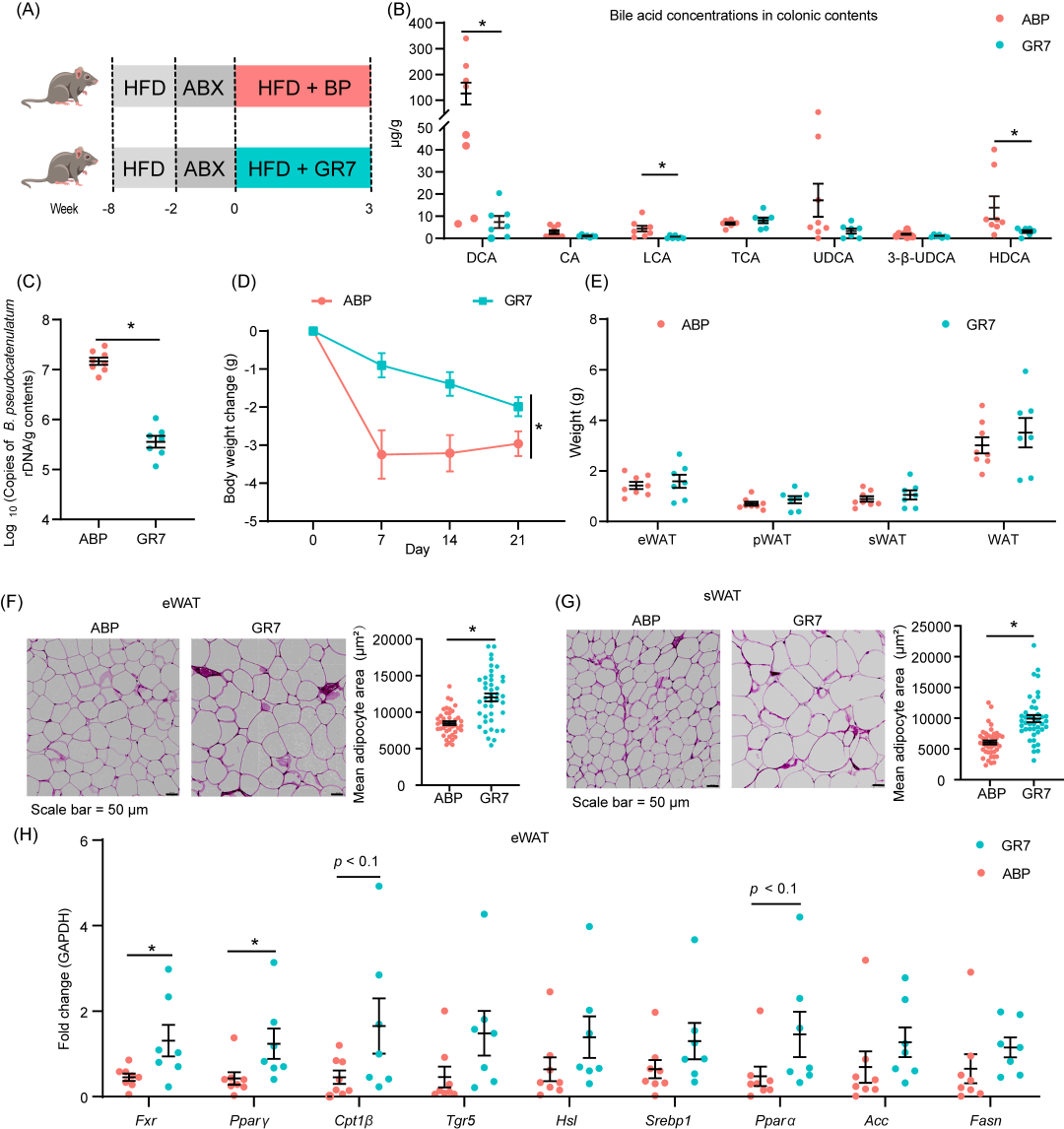
图5. 药理抑制胆盐水解酶会削弱假小链双歧杆菌对抗生素预处理的高脂饮食饲喂小鼠的降脂作用
(A)胆盐水解酶药理抑制实验示意图;(B)结肠胆汁酸浓度;(C)结肠内容物中假小链双歧杆菌 16S rDNA 的拷贝数;(D)体重变化;(E)器官重量;(F)附睾白色脂肪组织的H&E染色代表性图像,以及平均脂肪细胞面积的定量分析;(G) 皮下白色脂肪组织的H&E染色代表性图像,以及平均脂肪细胞面积的定量分析;(H)附睾白色脂肪组织中脂质代谢相关基因的 mRNA 表达。数据以平均值 ± 标准误表示, * 表示 p < 0.05(t 检验), ABP 组(n = 8),GR7 组(n = 7)。
石胆酸降低高脂饮食饲喂大鼠和宁乡猪脂肪沉积
通过对假小链双歧杆菌的单菌定植实验以及BSH抑制的药理学实验,发现这些干预持续影响了HFD小鼠结肠中的LCA浓度。为了进一步探讨LCA对脂肪沉积的影响,研究中在HFD中添加了0.3%的LCA。与HFD组相比,LCA组显著减少了体重增加和累积的食物摄入量(图6A、B)。LCA补充还显著降低了血清甘油三酯浓度,增加了LDLC浓度,并与血清脂蛋白脂肪酶(LPL)活性的增强相关,LPL是一种参与甘油三酯水解的酶(图S8A、B)。在组织水平上,LCA显著减少了eWAT、sWAT和WAT的重量,并减少了eWAT的脂肪细胞平均面积(图6C、D)。转录分析显示,LCA补充下调了eWAT中二酰甘油O-酰基转移酶1(Dgat1)、Fxr、激素敏感性脂肪酶(Hsl)和Pparγ的mRNA表达(图6E)。此外,LCA显著降低了肝脏重量和肝脏脂肪变性评分(图S8C、D)。转录分析表明,LCA上调了肝脏Fxr的表达,同时下调了脂肪酸合成酶(Fasn)和Acc的表达(图S8E)。
进一步的研究中,我们对宁乡猪进行了膳食干预实验。与正常对照组(NC)相比,LCA对体重变化没有显著影响,但显著降低了背膘厚度以及血清甘油三酯和NEFA浓度(图6F、G,S8F)。LCA补充还显著减少了肝脏的NEFA浓度,并与肝脏中Pparγ和Srebp1的下调以及Cpt1b的上调相一致(图S8G、H)。此外,LCA补充显著减少了皮下脂肪组织脂肪细胞的平均面积(图6H)。
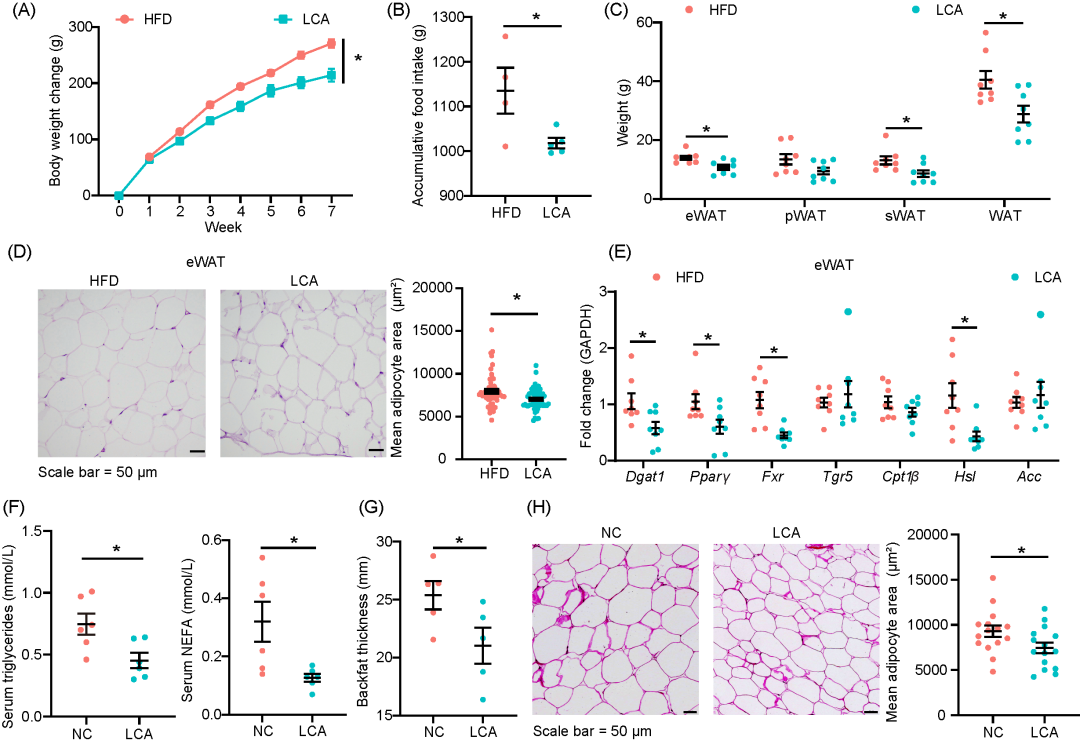
图6. 膳食补充石胆酸降低高脂饮食饲喂大鼠和宁乡猪脂肪沉积
(A)宁乡猪体重变化;(B)大鼠累计食物摄入量;(C)大鼠脂肪组织重量;(D)大鼠附睾白色脂肪组织的H&E染色代表性图像,以及平均脂肪细胞面积的定量分析;(E)大鼠附睾白色脂肪组织中脂质代谢相关基因的 mRNA 表达情况;(F)大鼠血清甘油三酯和非酯化脂肪酸(NEFA)浓度;(G)宁乡猪背膘厚度;(H)宁乡猪皮下脂肪组织的H&E染色代表性图像,以及平均脂肪细胞面积的定量分析。数据以平均值 ± 标准误表示,大鼠每组 n = 8, * 表示 p < 0.05(采用t 检验)。
讨 论
本研究通过杜长大和宁乡猪模型探讨了肠道微生物群对脂肪沉积的影响。研究表明,结肠微生物群对脂肪沉积的作用在脂肪型猪和瘦肉型猪之间存在差异,宁乡猪的微生物群特别促进脂肪积累。此外,宏基因组学和代谢组学分析显示,假小链双歧杆菌可能通过调节次级胆汁酸代谢改善脂肪沉积。研究发现,假小链双歧杆菌能有效降低HFD饲喂小鼠的脂肪沉积和肝脏脂肪变性。其机制表明,假小链双歧杆菌通过增强次级胆汁酸合成来调控脂肪沉积,而通过药理学抑制BSH则会削弱假小链双歧杆菌的抗脂肪沉积效应。此外,LCA在HFD喂养的大鼠和宁乡猪中也表现出类似的抗脂肪沉积作用。
肠道微生物群与脂肪沉积的关系已通过无菌动物和粪便微生物群移植得到了验证。以往研究表明,人类和小鼠的肥胖与健康个体之间存在显著的肠道微生物群组成差异,且肥胖个体的肠道微生物群能够增强从饮食中获取的能量。本研究还发现,杜长大猪和宁乡猪的肠道微生物群组成存在显著差异,其中宁乡猪的结肠微生物群富集了与脂质代谢相关的通路。通过将肥胖双胞胎的粪便微生物群移植到小鼠中,研究表明,接受肥胖双胞胎粪便的鼠类体内脂肪沉积有所增加。进一步地,将金华猪和长白猪的粪便微生物群分别移植到广谱抗生素处理的小鼠体内,结果表明接受金华猪粪便微生物群的小鼠肝脏和腹部脂肪沉积增加。本研究的实验中,将杜长大猪和宁乡猪的结肠微生物群分别移植到不同饮食的小鼠体内,结果表明,接受宁乡猪结肠微生物群的小鼠在HFD下肝脏和WAT中的脂肪沉积增加。此外,与接受杜长大猪结肠微生物群的小鼠相比,接受宁乡猪结肠微生物群的小鼠血清总胆汁酸浓度较低。这些结果表明,脂肪型猪源的结肠微生物群在诱导宿主脂肪沉积方面比瘦肉型猪源的微生物群具有更强的能力。
某些肠道微生物,如Akkermansia muciniphila和双歧杆菌已被证明能够调节宿主的脂肪储存。Zhao等人通过粪便宏基因组学研究表明,假小链双歧杆菌改善了葡萄糖稳态。此外,本研究的宏基因组学分析也揭示了结肠中假小链双歧杆菌的丰度与背脂厚度呈负相关。更有趣的是,已有研究报告指出,假小链双歧杆菌的相对丰度较高与肥胖呈负相关。在本研究中,给HFD喂养的小鼠灌胃活的假小链双歧杆菌能减少脂肪沉积和肝脏脂肪变性。然而,给小鼠灌胃假小链双歧杆菌的上清液和热灭活菌未能产生类似效果,这表明假小链双歧杆菌对脂肪沉积的调节作用可能与其代谢过程中与机体的相互作用有关。上清液和热灭活菌未能产生类似效果的原因可能在于关键代谢物的不稳定或浓度不足,或缺乏与宿主之间的直接相互作用,而这些对调控脂质代谢是至关重要的。
次级胆汁酸代谢在体内和体外差异显著,胆汁酸在脂质代谢中发挥着至关重要的作用。研究还发现,LCA类胆酸在杜长大猪中显著富集,并与假小链双歧杆菌正相关。在一种胶原诱导的关节炎小鼠模型中,给小鼠灌胃假小链双歧杆菌增加了粪便中的LCA浓度。与此一致,本研究表明给HFD喂养的小鼠灌胃活假小链双歧杆菌也能增加结肠中的LCA浓度。这些结果表明,假小链双歧杆菌可能通过增强次级胆汁酸合成来缓解脂肪沉积。BSH作为次级胆汁酸合成的关键酶,其作用机制尚不明确。Targoński等人从假小链双歧杆菌中纯化了BSH。为了探讨BSH在脂质代谢中的作用,研究使用了GR7作为药理学抑制剂。结果表明,GR7抑制BSH活性,减少了假小链双歧杆菌在HFD喂养小鼠结肠中的定植,同时增加了血清甘油三酯、脂肪细胞平均面积和肝脂肪变性评分。这些结果与已有研究表明其他细菌(如Turicibacter菌株)表达胆汁盐修饰基因能降低血清胆固醇、甘油三酯和脂肪组织质量的发现一致。另一方面,研究发现,某些研究表明抑制BSH可能有助于缓解非酒精性脂肪肝病或肥胖等疾病。因此,当前研究中的差异值得进一步探讨。
LCA作为一种非12-OH型胆汁酸,在体重指数与HFD喂养小鼠中的比例关系中表现出负相关。LCA水平在袖状胃切除术后升高,有助于改善糖尿病,并通过产生胆酸-7-硫酸盐起作用。本研究中,LCA补充降低了大鼠和宁乡猪的脂肪沉积和肝脏脂肪变性。然而,有研究指出,LCA可能会加剧葡萄糖耐受性,损害正常代谢,尤其是在常规饮食的小鼠中。我们推测这可能与小鼠的生理状态和LCA浓度有关。以往研究表明,LCA既是Fxr和Tgr5的激动剂,而这两种受体的上调已被证明能降低脂肪沉积。我们的研究验证了这一发现,表明LCA通过上调肝脏的Fxr和Tgr5表达,缓解肝脂肪变性,可能通过这些途径降低脂肪积累。然而,在脂肪组织中,LCA并未显著影响Fxr和Tgr5表达,而是下调了Pparγ的表达。这表明Fxr、Tgr5和Pparγ可能是LCA的潜在靶点,但这一假设需要在基因敲除模型中进一步验证。尽管本研究已证明LCA可减少脂肪沉积,但当前的机制研究仍然有限。未来的研究应进一步探讨这一效应的机制。
研究的局限性:首先,宁乡猪和杜长大猪具有不同的遗传背景,肠道微生物群的遗传背景效应尚未完全理解;其次,本研究忽略了性别对肠道微生物群和胆汁酸谱的影响;第三,未来应进行假小链双歧杆菌的BSH过表达和无菌小鼠研究。此外,每次胆汁酸的肠肝循环中,约95%的胆汁酸被肠道回收,只有5%的胆汁酸进入结肠并转化为二级胆汁酸。因此,在后续实验中,更全面地考虑回肠胆汁酸的变化可能有助于我们更好地理解这一问题。
结 论
总的来说,我们的研究表明,结肠微生物群对脂肪沉积的作用在脂肪型猪和瘦肉型猪之间存在差异。通过宏基因组学和代谢组学分析,我们发现假小链双歧杆菌和LCA类胆酸在杜长大猪的结肠中显著富集。此外,灌胃活的假小链双歧杆菌可以减少肝脏和脂肪组织中的脂肪沉积,同时增加结肠中的LCA浓度。通过药理学抑制假小链双歧杆菌的BSH会导致HFD饲喂小鼠脂肪沉积增加。进一步,LCA的膳食补充也能减少HFD饲喂大鼠和宁乡猪的脂肪沉积。这些结果证明了假小链双歧杆菌通过增强次级胆汁酸的合成来减少脂肪沉积。
方 法
实验动物与处理方法
动物实验 1:杜长大和宁乡猪样本收集
本研究使用了40头杜长大猪和40头宁乡猪(楚沩香集团有限公司,长沙,中国),体重约为100公斤。宁乡猪和杜长大猪的饲料特别根据它们不同的营养需求配制,所有猪都有自由进食和饮水的权利。取材后,迅速收集组织样本。在第6和第7胸椎交界处,用游标卡尺测量背膘厚度。
动物实验 2:结肠微生物群移植实验(常规饲料喂养)
根据先前描述的方法制备结肠微生物群悬液。简而言之,采集健康杜长大和宁乡猪的结肠内容物,使用无菌生理盐水混匀,经过无菌不锈钢筛网过滤,并离心去除不溶性物质。将悬液的OD600调整为1.0,并加入10%无菌甘油(最终浓度),然后存放在-80°C。
实验设计如图2A所示。7-8周龄的小鼠(C57BL/6J,SLAC,上海,中国)在饮水中加入抗生素混合物(氯霉素0.5 g/L、庆大霉素1 g/L、甲硝唑1 g/L、氨苄西林1 g/L,均来自Sigma)喂养2周。然后将小鼠随机分为两组,分别用不同品种猪的结肠细菌悬液处理:(1)DLY-CD组:接受200 µL来自杜长大猪的结肠细菌悬液(n = 8);(2) NX-CD组:接受200 µL来自宁乡猪的结肠细菌悬液(n = 8)。所有小鼠都喂标准常规饲料(脂肪能量占10%;3.25 kcal/g;SLAC)。实验8周后,每组8只小鼠被安乐死,收集肝脏、脂肪组织和血清样本以供进一步分析。
动物实验 3:结肠微生物群移植实验(高脂饲料喂养)
实验设计如图2A所示。7-8周龄的小鼠(C57BL/6J,SLAC)用抗生素混合物处理两周后,分别用不同品种猪的结肠细菌悬液处理:(1) DLY-HFD组:接受来自杜长大猪的结肠细菌悬液(n = 8);(2) NX-HFD组:接受来自宁乡猪的结肠细菌悬液(n = 8)。所有小鼠都喂高脂饲料(HFD,脂肪占60%,D12492)。实验8周后,每组8只小鼠被安乐死,收集肝脏、脂肪组织和血清样本以供进一步分析。
动物实验 4:假小链双歧杆菌单菌定殖实验
实验设计如图4A所示。假小链双歧杆菌(CGMCC 1.2277)来自中国普通微生物菌种保藏管理中心。假小链双歧杆菌在无菌BBL培养基(MM1921,Coolaber)中,于37°C下培养48小时,在厌氧箱(85% N2:10% H2:5% CO2)中培养。
7-8周龄健康雄性小鼠随机分为5组:(1) NC组:常规饲料(n = 13);(2) HFD组:高脂饲料(n = 13);(3) BPS组:高脂饲料,并灌胃200 µL 假小链双歧杆菌培养上清液(n = 13);(4) HK组:高脂饲料,并灌胃200 µL热灭活的假小链双歧杆菌(1×108 CFU/200 µL BBL培养基)(n = 13);(5) BP组:高脂饲料,并灌胃200 µL活的假小链双歧杆菌(1×108 CFU/200 µL BBL培养基)(n = 13)。实验9周后,每组8只小鼠被安乐死,收集肝脏、脂肪组织、结肠和血清样本以供进一步分析。
动物实验 5:伪无菌小鼠中的假小链双歧杆菌单菌定殖实验
实验设计如图S6A所示。7-8周龄健康雄性小鼠(C57BL/6J,SLAC)用抗生素混合物处理2周。然后将小鼠随机分为两组:(1) ABX组:喂高脂饲料,并灌胃500 µL BBL培养基(n = 6);(2) ABP组:喂高脂饲料,并灌胃500 µL活的假小链双歧杆菌(5×108 CFU/500 µL BBL培养基)(n = 6)。实验5周后,每组6只小鼠被安乐死,收集肝脏、脂肪组织、结肠和血清样本以供进一步分析。
动物实验 6:药理学抑制假小链双歧杆菌的胆盐水解酶
实验设计如图5A所示。8周龄雄性C57BL/6J小鼠喂高脂饲料8周后,使用抗生素混合物处理2周。通过有氧和厌氧平板涂布来验证微生物群的去除情况。然后将小鼠随机分为两组:(1) ABP组:喂高脂饲料,并灌胃500 µL活的假小链双歧杆菌(5×108 CFU/500 µL BBL培养基)(n = 8);(2) GR7组:喂高脂饲料,灌胃500 µL活的假小链双歧杆菌(5×108 CFU/500 µL BBL培养基)并加入10 mg/kg肠道特异性GR7(HY-135747,MedChemExpress)(n = 7)。实验3周后,15只小鼠被安乐死,收集肝脏、脂肪组织、结肠和血清样本以供进一步分析。
动物实验 7:LCA处理实验(大鼠)
8周龄健康雄性大鼠(SLAC,中国)根据体重随机分为2组:(1) HFD组:喂高脂饲料(n = 8);(2) LCA组:0.3% LCA 添加的高脂饲料(L106779,阿拉丁)(n = 8)。LCA的剂量参考先前的研究。大鼠在12小时光暗周期下饲养。实验持续7周,实验结束后,收集肝脏、脂肪组织、结肠和血清样本以供进一步研究。
动物实验 8:LCA处理实验(宁乡猪模型)
共12头猪,按体重随机分为2组:(1) NC组:喂基础饲料;(2) LCA组:喂基础饲料并补充792 mg/kg LCA(银丰生物,中国)。基础饲料配方见表S3,满足宁乡猪的营养需求。所有猪有自由进食和饮水的权利。LCA的剂量基于杜长大猪结肠中LCA浓度的测定。实验10周后,每组5头猪被安乐死,收集肝脏、脂肪组织、结肠和血清样本以供进一步研究。
DNA提取与16S rRNA基因测序
结肠内容物中的细菌DNA使用Mobio PowerSoil R DNA提取试剂盒(Zymo Research, Irvine, CA)按照制造商说明提取。提取的DNA通过2%琼脂糖凝胶电泳和Nanodrop-1000(Thermo Scientific, Wilmington, MA)进行评估。条形码引物为341F(5'-CCTACGGGNGGCWGCAG-3')和805R(5'-GACTACHVGGGTATCTAATCC-3')。这些引物用于扩增16S rRNA的V3-V4超变区。纯化的PCR扩增产物用于构建测序文库。所有扩增子通过Novaseq平台(Illumina, San Diego, CA)采用双端测序方法进行测序。序列数据可以在SRA数据库中找到(PRJNA816269)。通过FLASH(版本1.2.7)获得了6113705个原始标签,每个样本平均为76421 ± 57个原始标签。通过Trimmomatic(v. 0.33)获得了5763836个清洁标签,每个样本平均为72048个清洁标签。最后,通过UCHIME(v. 4.2)除嵌合体,获得有效标签。有效标签根据QIIME v. 1.9.1管道进行处理,并通过开放参考算法以97%的相似性聚类为操作性分类单元(OTUs)。使用RDP分类器与Greengenes数据库(2013年5月发布)其进行分类。多样性分析基于OTU表和特征序列进行。α多样性通过观察的OTUs、Shannon、Simpson和Pielou指数计算;β多样性基于Bray-Curtis距离和加权UniFrac距离计算。序列数据可在国家生物技术信息中心(NCBI)中找到(GSA: PRJNA816269)。
宏基因组测序分析
DNA测序文库使用TruSeq Nano DNA LT文库制备试剂盒–A套件(FC-121-4001,Illumina)和无核酸酶水(AM9932,Ambion,Austin,Texas)进行制备。在验证测序文库后,通过NovaSeq 6000平台(PE150,Illumina)进行高通量测序(LC-Bio)。
原始测序数据经过处理以生成用于深入分析的清洁数据。接头修剪使用Trim Galore(v. 0.6.5)进行,参数如下:去除小于75 bp的读取、修剪质量分数低于20的碱基,去除接头序列。使用BMTagger去除宿主基因组序列,参考Sus scrofa 11.1基因组(NCBI登录号GCA_000003025.6)。使用FastQC (v. 0.11.9)对处理后的数据进行质量控制检查。通过Kaiju对数据进行分类,结果与NCBI RefSeq数据库对齐。使用Prodigal(v. 2.6.3)预测开放阅读框(ORFs)。功能注释使用Diamond(v. 2.0.15)进行,E值阈值设定为 ≤ 10−10,并与KEGG数据库(https://www.kegg.jp/)进行对齐。多样性分析基于OTU表和特征序列进行。序列数据可以在中国国家生物信息中心找到(登录号PRJCA022867)。
结肠代谢组学分析
结肠代谢组学分析使用UPLC-MS/MS(ACQUITY UPLC-Xevo TQ-S,Waters)进行。所有代谢物标准品均由Sigma-Aldrich,Steraloids Inc和TRC Chemicals提供。由UPLC-MS/MS生成的原始数据文件使用Mass Lynx软件(v. 4.1,Waters)进行处理。统计分析使用自主开发的平台iMAP(Metabo-Profile),包括主成分分析、单变量分析和通路分析。
假小链双歧杆菌定量
从结肠内容物中提取总DNA,使用DNA提取试剂盒(Vazyme, 南京,中国)。使用特异性引物(BiCAT-F: 5'-CGGATGCTCCGACTCCT-3',BiCAT-R: 5'-CGAAGGCTTGCTCCCGAT-3')对假小链双歧杆菌进行定量。采用实时荧光定量试剂盒(SYBR Green,艾科瑞)检测其拷贝数。
血清生化指标的测定
血清ALT、AST、ALP、总胆汁酸、葡萄糖、甘油三酯、胆固醇、LDLC、HDLC和NEFA浓度使用自动生化分析仪和试剂盒(罗氏)进行测定。肝脏甘油三酯水平使用甘油三酯测定试剂盒(NJJCBIO)按照制造商说明进行测定。
肝脏和白色脂肪组织的组织学分析
组织样本在安乐死后收集并固定在4%多聚甲醛溶液中。固定后的样本通过乙醇脱水,石蜡包埋,H&E染色后用中性树脂封片。肝脏脂肪变性评估使用数字显微镜(BA210Digital,Motic)进行。脂肪细胞大小通过Motic Images Advanced软件测量。脂肪细胞面积通过ImageJ 6.0进行测量。
LPL活性测定
组织样本被匀浆,蛋白质浓度使用BCA蛋白定量试剂盒(碧云天)进行测定。LPL活性使用试剂盒(CUSABIO)进行测定,结果根据蛋白质浓度进行标准化。
细菌基因组功能注释
假小链双歧杆菌的基因组序列(GCA_001025215.1)从NCBI下载。细菌基因组的功能注释使用Proksee进行。
次级胆汁酸的测定
将20 mg结肠内容物与1 mL甲醇-水溶液(v/v = 5:5)混合,溶液中加入2%内标物(D4-CA,D4-LCA,60 µg/mL每种)。混合物进行超声提取,频率为60 Hz,时间为10分钟,之后在4°C下以13,000 g离心10分钟。收集上清液,并重复提取三次。然后,将700 µL汇集的上清液通过固相萃取处理,最后通过0.22 µm滤膜过滤。最终的上清液转移至1 mL小瓶中,采用UPLC-MS进行分析。本研究中检测的次级胆汁酸包括:胆酸(CA)、牛磺胆酸(TCA)、脱氧胆酸(DCA)、甘氨胆酸(GCA)、熊脱氧胆酸(UDCA)、牛磺脱氧胆酸(TDCA)、甘氨脱氧胆酸(GDCA)、3β-熊脱氧胆酸(3β-UDCA)、豚鼠脱氧胆酸(HDCA)、异脱氧胆酸(iso-DCA)、石胆酸(LCA)、表石胆酸(epi-LCA)、异石胆酸(iso-LCA)、3α-羟基-5α-胆酸(3α-ol-5α-CA)、7-酮石胆酸(7-KLCA)、12-酮石胆酸(12-KLCA)、5α-胆酸-3α-醇(5α-CA-3α-ol)、甘氨胆氨酸(GCDCA)、甘氨熊脱氧胆酸(GUDCA)、牛磺胆氨酸(TCDCA)、牛磺熊脱氧胆酸(TUDCA)、胆脱氧胆酸(CDCA)、甘氨醇脱氧胆酸(GHDCA)、牛磺醇脱氧胆酸(THDCA)、异胆酸(ACA)、牛磺石胆酸(TLCA)。
实时定量PCR
样本被研磨成粉末,使用TRIZOL(15596-026,Invitrogen)方法提取总RNA。随后,使用Nanodrop ND-1000(Thermo Scientific)测量总RNA的浓度和质量。总RNA通过逆转录试剂盒(Takara)反转录为cDNA。引物见表S4。使用实时荧光定量试剂盒(SYBR Green,艾科瑞)检测目标基因的mRNA表达,使用甘油三磷酸脱氢酶(GAPDH)作为内参基因进行基因表达的归一化。
统计分析
数据使用Excel 2016整理。两因素重复测量方差分析(ANOVA)用于比较多个时间点之间两条曲线的趋势。弗里德曼检验用于五组比较的重复测量。两组之间的比较使用双尾非配对学生t检验。对于多组比较,正态分布数据使用单因素ANOVA,非正态分布数据使用Kruskal-Wallis检验。统计分析使用IBM SPSS Statistics 22.0软件(IBM Corp. 2013年发布,IBM SPSS Statistics for Windows,版本22.0,纽约,IBM Corp.)进行,显著性设定为p < 0.05。数据可视化使用R(v. 4.1.2)和Graphpad Prism 8.0软件(IBM)进行。其他数据分析,包括主坐标分析、Mantel检验、线性判别分析效应大小和Spearman相关分析,均在RStudio(v 2024.04.2)中进行。随机森林分析使用R的“randomForest”包(v. 4.1.2)进行。
代码和数据可用性
本文报告的宏基因组原始序列数据已存入基因组序列档案(Genomics, Proteomics & Bioinformatics 2021)于中国国家基因组数据中心(Nucleic Acids Res 2022),中国科学院北京基因组研究所(GSA: CRA014539),并可公开访问:https://ngdc.cncb.ac.cn/gsa。本文报告的16S rRNA测序原始数据已存入NCBI(GSA: PRJNA816269,https://www.ncbi.nlm.nih.gov/bioproject/PRJNA816269/)。数据和脚本可在GitHub上获得:https://github.com/ZAD-Vitamin/ZhaiMeta。补充材料(图表、表格、图形摘要、幻灯片、视频、中文翻译版和更新材料)可通过在线DOI或iMeta Science(http://www.imeta.science/)获取。
引文格式:
Andong Zha, Ming Qi, Yuankun Deng, Hao Li, Nan Wang, Chengming Wang, Simeng Liao, et al. 2024. “Gut Bifidobacterium pseudocatenulatum protects against fat deposition by enhancing secondary bile acid biosynthesis.” iMeta 3. e261. https://doi.org/10.1002/imt2.261.
作者简介

查安东(第一作者)
● 中南大学基础医学院青年教师。
● 研究方向为肠道菌群和能量代谢,以第一作者在iMeta、Animal nutrition、Ecotoxicology and Environmental Safety等期刊发表SCI论文11篇。

王婧(通讯作者)
● 湖南农业大学动物科技学院教授,硕士生导师。
● 研究方向为仔猪发育与营养调控、母仔营养传递与精准饲喂技术。入选2018年博士后创新人才计划、湖南农业大学神农学者ш层次人才计划,在Science Bulletin 等高影响力期刊发表论文20余篇。

印遇龙(通讯作者)
● 中国科学院亚热带农业生态研究所研究员,院士,博士生导师。
● 畜禽养殖污染控制与资源化技术国家工程实验室主任,中国农学会微量元素与食物链分会理事长,世界中医药学会联合会中医与农业产业分会、芳香产业分会名誉理事长,中国饲料工业协会副会长,Animal Nutrition杂志主编,中国科学 生命科学中英文版编委,Journal of Animal Science编委。长期从事畜禽健康养殖与环境控制研究,先后主持完成院、省、国家、国际合作科研项目30多项。在PNAS、CNS子刊等刊物发表高质量论文300多篇,英文论文总被引50,000多次,H-index in Google Scholar 102,以第一完成人获国家科技进步奖二等奖二项和国家自然科学奖二等奖一项,曾获湖南省杰岀贡献奖,何梁何利科技进步奖,2018年在澳大利亚布里斯班举行的第14届国际猪消化生理学大会上获Asia-Pacific Nutrition Award (杰出成就奖)。

谭碧娥(通讯作者)
● 湖南农业大学二级教授,博士生导师。
● 主要从事猪肠道发育和营养调控。先后主持国家自然科学基金5项、国家重点研发计划课题、国家科技支撑计划专题、973项目子课题等。近年来以第一/通讯作者发表SCI收录论文100多篇,SCI论文合计引用11000多次,单篇最高被引用1900多次,H指数48;主编著作2部,参编著作6部,授权发明专利、软件著作权和制订标准规程等10多项;获国家自然科学奖二等奖(排名第2)、中华农业科技奖一等奖2项(排名第3和5)和创新团队奖(排名第5)、湖南省自然科学一等奖(排名第2)和二等奖(排名第3)以及中科院青年创新促进会优秀会员、中科院广州分院优秀青年科学家奖、中国科学院院长优秀奖。现为湖南省畜牧兽医学会副理事长、畜禽产品品质调控湖南省重点实验室主任、中国畜牧兽医学会动物营养学分会理事等。入选教育部长江学者奖励计划青年学者、国家“万人计划”科技创新领军人才、科技部创新人才推进计划、湖南省科技创新领军人才。
更多推荐
(▼ 点击跳转)
iMeta | 引用15000+,海普洛斯陈实富发布新版fastp,更快更好地处理FASTQ数据
iMeta | 兰大张东组:使用PhyloSuite进行分子系统发育及系统发育树的统计分析
iMeta | 唐海宝/张兴坦-用于比较基因组学分析的多功能分析套件JCVI

1卷1期

1卷2期

1卷3期

1卷4期

2卷1期

2卷2期

2卷3期

2卷4期

3卷1期

3卷2期

3卷3期

3卷4期

3卷5期

1卷1期

1卷2期
期刊简介
“iMeta” 是由威立、宏科学和本领域数千名华人科学家合作出版的开放获取期刊,主编由中科院微生物所刘双江研究员和荷兰格罗宁根大学傅静远教授担任。目的是发表所有领域高影响力的研究、方法和综述,重点关注微生物组、生物信息、大数据和多组学等前沿交叉学科。目标是发表前10%(IF > 20)的高影响力论文。期刊特色包括中英双语图文、双语视频、可重复分析、图片打磨、60万用户的社交媒体宣传等。2022年2月正式创刊!相继被Google Scholar、PubMed、SCIE、ESI、DOAJ、Scopus等数据库收录!2024年6月获得首个影响因子23.8,位列全球SCI期刊前千分之五(107/21848),微生物学科2/161,仅低于Nature Reviews,学科研究类期刊全球第一,中国大陆11/514!
“iMetaOmics” 是“iMeta” 子刊,主编由中国科学院北京生命科学研究院赵方庆研究员和香港中文大学于君教授担任,是定位IF>10的高水平综合期刊,欢迎投稿!
iMeta主页:
http://www.imeta.science
姊妹刊iMetaOmics主页:
http://www.imeta.science/imetaomics/
出版社iMeta主页:
https://onlinelibrary.wiley.com/journal/2770596x
出版社iMetaOmics主页:
https://onlinelibrary.wiley.com/journal/29969514
iMeta投稿:
https://wiley.atyponrex.com/journal/IMT2
iMetaOmics投稿:
https://wiley.atyponrex.com/journal/IMO2
邮箱:
office@imeta.science

















 3139
3139

 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









