






点击蓝字 关注我们
土壤养分周转与作物地上养分代谢之间的联系 — 土壤微生物的作用

iMetaOmics主页:http://www.imeta.science/imetaomics/
研究论文
● 原文链接DOI: https://doi.org/10.1002/imo2.55
● 2025年2月3日,中国农业科学院植物保护研究所王秋霞、方文生等在iMetaOmics在线发表了题为“Linkages between soil nutrient turnover and above-ground crop nutrient metabolism: the role of soil microbes”的文章。
● 本研究探讨了微生物群落在土壤氮养分周转及作物地上部氮代谢中的关键纽带作用,揭示土壤微生物群落的变化破坏土壤氮循环的平衡后,产生的级联效应引发硝酸盐及铵转运蛋白的特异性表达,导致作物地上部氮素空间分布发生改变。研究结果为管理土壤微生物群落提高作物养分效率提供了支撑。
● 第一作者:方文生
● 通讯作者:王秋霞(wqxcasy@163.com)
● 合作作者:田文凤、颜冬冬、李园、曹坳程
● 主要单位:植物病虫害综合治理全国重点实验室、中国农业科学院植物保护研究所,湖南农业大学园艺学院
亮 点
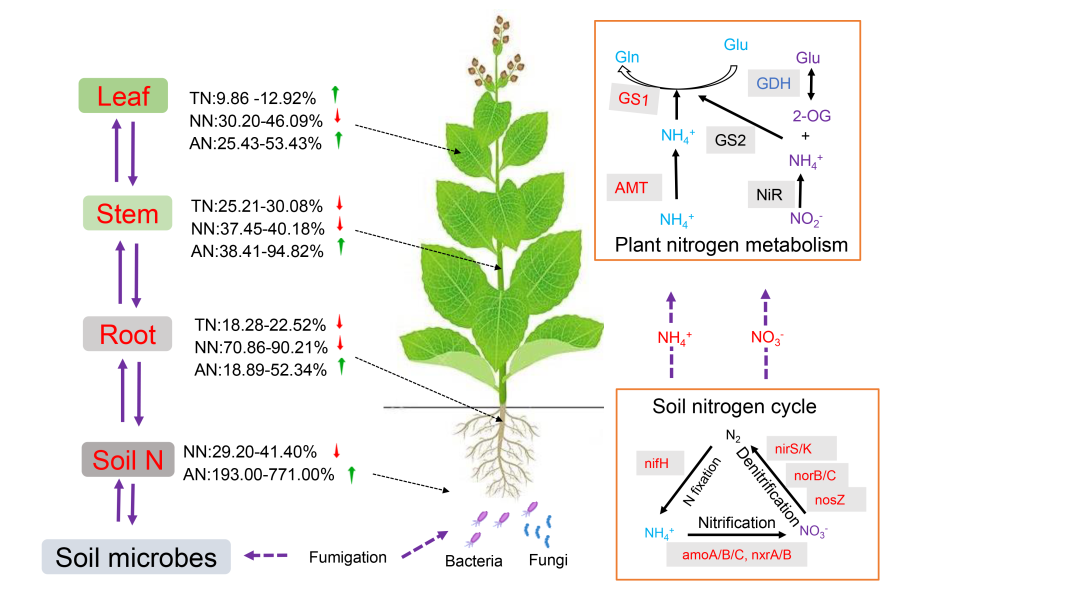
● 土壤熏蒸显著改变了植物体内氮的空间分布;
● 熏蒸后土壤微生物群落发生改变促进反硝化微生物丰度;
● 微生物群落的变化导致土壤氮平衡被打破;
● 氮代谢酶活性增加且氮转运基因的表达被刺激;
● 土壤微生物通过改变养分周转过程影响植物地上部养分代谢。
摘 要
破解土壤养分周转和作物养分吸收之间的复杂关系对于提高作物生产力至关重要。本研究聚焦棉隆土壤熏蒸后的氮转化过程,重点探讨了土壤微生物如何影响土壤氮循环和作物地上部氮代谢。结果发现,土壤熏蒸导致烟草氮素空间分布发生显著改变,烟草根、茎、叶中铵态氮增加了18.89% ~ 94.82%,硝态氮减少了30.20% ~ 90.21%。土壤熏蒸引起微生物群落发生明显变化,抑制了铵氧化微生物,同时刺激了反硝化微生物。这些变化不仅破坏了氮平衡—使土壤硝化强度减少72.91% ~ 86.51%,反硝化强度增加197% ~ 324%,而且还对作物地上部的氮代谢产生了连带影响。通过改变微生物群落的组成,直接影响土壤养分的周转,进而影响作物养分的供应和分布。氮平衡的破坏进一步导致关键氮代谢酶和转运体基因的表达发生变化。具体而言,熏蒸促进了硝酸盐和铵转运体相关的基因以及氨基酸代谢酶基因的表达。结构方程模型证实,微生物群落的变化是土壤养分周转和作物氮素分布变化的核心驱动力。这些发现强调了土壤微生物群落作为土壤养分循环和地上部养分代谢之间的纽带作用,突出了微生物群落在调节植物养分吸收和利用方面的重要性。因此,优化微生物管理具提高作物养分效率的潜力。
视频解读
Bilibili:https://www.bilibili.com/video/BV1uuNcesEdZ/
Youtube:https://youtu.be/9xwEFh-TaOU
中文翻译、PPT、中/英文视频解读等扩展资料下载
请访问期刊官网:http://www.imeta.science/imetaomics/
全文解读
引 言
土壤微生物与作物间的相互作用是生态系统中一个复杂而动态的过程,涉及物质循环、能量流动和信息传递。这种错综复杂的相互作用对维持生态平衡和作物生产力至关重要。尽管科学研究取得了重大进展,但仍有许多未解之谜,特别是关于土壤微生物如何调节土壤养分周转并影响作物体内养分的分布尚不清。解析这些相互作用对揭示生态系统功能的基本机制和提高农业生产力至关重要,将助力农业可持续发展。
土壤熏蒸是一种有效的土传病害管理方法,它通过消灭病原微生物(包括真菌、细菌和线虫)并重构微生物群落,显著改善作物生长的土壤环境,具有显著的“肥料效应”,可改善作物对养分和水分的吸收,促进作物生长。然而,前期研究表明土壤熏蒸破坏氮代谢,导致土壤中无机氮的存在形式和数量发生显著变化。这些干扰是否影响作物对氮的吸收及氮在植物组织的空间分布仍然不清。
微生物在土壤养分周转中起着关键作用,承担着有机物的分解和矿化,将复杂的有机化合物转化为作物易于吸收的氮、磷、钾等必需的无机养分。这些微生物同时影响作物地上部养分的供应和代谢,如固氮菌和菌根真菌可显著提高养分吸收率,从而直接影响作物的生长和产量。土壤养分与作物之间的相互作用是通过根系分泌物来实现的,根系分泌物为土壤微生物提供碳和能量来源,刺激其生长和活动。这种互惠关系促进了养分的高效循环和供应,从而支持作物的养分吸收和新陈代谢。作物和微生物之间持续的养分交换保障了必需元素的有效转换和利用。此外,通过改变养分供应和循环过程,微生物还能调节作物所需的养分供应速率。这种相互作用凸显了管理土壤微生物群落对提高养分周转和改善作物养分吸收的重要性。有效的管理方法可以提高作物养分利用效率,提高作物产量、促进农业可持续发展。
棉隆(DZ)是中国广泛使用的土壤熏蒸剂,施用到土壤可迅速分解为其活性成分异硫氰酸甲酯,发挥熏蒸作用。本研究探讨了DZ 熏蒸对氮代谢的影响,特别关注了土壤微生物群落的变化如何影响作物的氮吸收和氮素空间分布。研究目标有两个:(1)阐明DZ熏蒸后作物组织中氮的空间分布及代谢机制;(2)探索土壤微生物影响作物地下-地上养分分布的潜在机制。研究结果将为了解土壤养分-微生物-作物之间的相互作用提供重要见解,为农业可持续性提供理论基础。
结 果
DZ熏蒸条件下氮素的时空变化
在北京土壤中,熏蒸第 30 天时,根、茎和叶中的总氮 (TN) 含量没有发生变化(图 1A-1C)。到第 50 天,与未熏蒸组相比,根(减少 21.89%,p < 0.05)、茎(减少 45.81%,p < 0.05)和叶(减少 20.95%,p < 0.05)的总氮含量明显减少(图 1A-1C)。相反,在云南土壤中,熏蒸组根部和茎部的氮含量在第 30 天(减少 32.11%-33.29%,p < 0.05)和第 50 天(减少 18.44%-25.35%,p < 0.05)出现持续下降(图 1A-1C)。熏蒸组与未熏蒸组相比,叶片在第 30 天最初没有差异,但在第 50 天时,熏蒸组的氮含量明显高于未熏蒸组(增加 12.92%,p < 0.05)。上述结果表明,随着时间的推移,DZ 熏蒸导致北京土壤中的氮逐渐减少。相比之下,在云南土壤中,熏蒸的影响更为直接,早期达到峰值,随后氮含量下降。在北京和云南土壤中,与对照组相比,根、茎和叶中的铵态氮(AN)含量在 DZ 熏蒸后明显升高(分别增加 37.49%-52.34%、31.91%-94.82% 和 25.44%-53.96%;p < 0.05,图 1D-1F)。值得注意的是,在云南土壤中,与第 30 天相比,第 50 天茎中 AN 的含量显著增加(94.82% vs. 31.91%,p < 0.05)(图 1E)。同样,在北京土壤中,根中 AN 的增加量(50.68% vs. 37.49%,p < 0.05)也明显高于第 30 天(图 1E)。同样,在北京土壤中,根(50.68% vs. 37.49%,p < 0.05)、茎(48.89% vs. 38.14%,p < 0.05)和叶(53.96% vs. 25.44%,p < 0.05)在第 50 天的增加量显著高于第 30 天。这些结果表明,土壤类型显著影响DZ熏蒸后作物氮素的时空分布。
在云南土壤中,DZ熏蒸显著刺激了根、茎和叶中硝态氮(NN)的代谢,使其分别减少了70.85%-88.64%、47.45%-40.18%和4.5%-30.20%(除第50天的叶片外,其余均p < 0.05,图1G-1I)。这种影响在第 30 天更明显,尤其是根部,其次是茎和叶。相反,在北京土壤中,促进 NN 代谢主要是在第 30 天(根部减少 66.39%,p < 0.05;茎部变化可忽略不计,叶片减少 40.28%,p < 0.05)。到了第 50 天,熏蒸效果过渡到抑制作用,导致根(200.43%)、茎(108.74%)和叶(74.09%)的显著增加(p < 0.05)。空间分布分析发现,无论是否熏蒸,烟草根、茎和叶中的 AN 含量都会急剧下降,而 TN 和 NN 则会逐渐增加,并主要在叶片中积累(图 1J)。
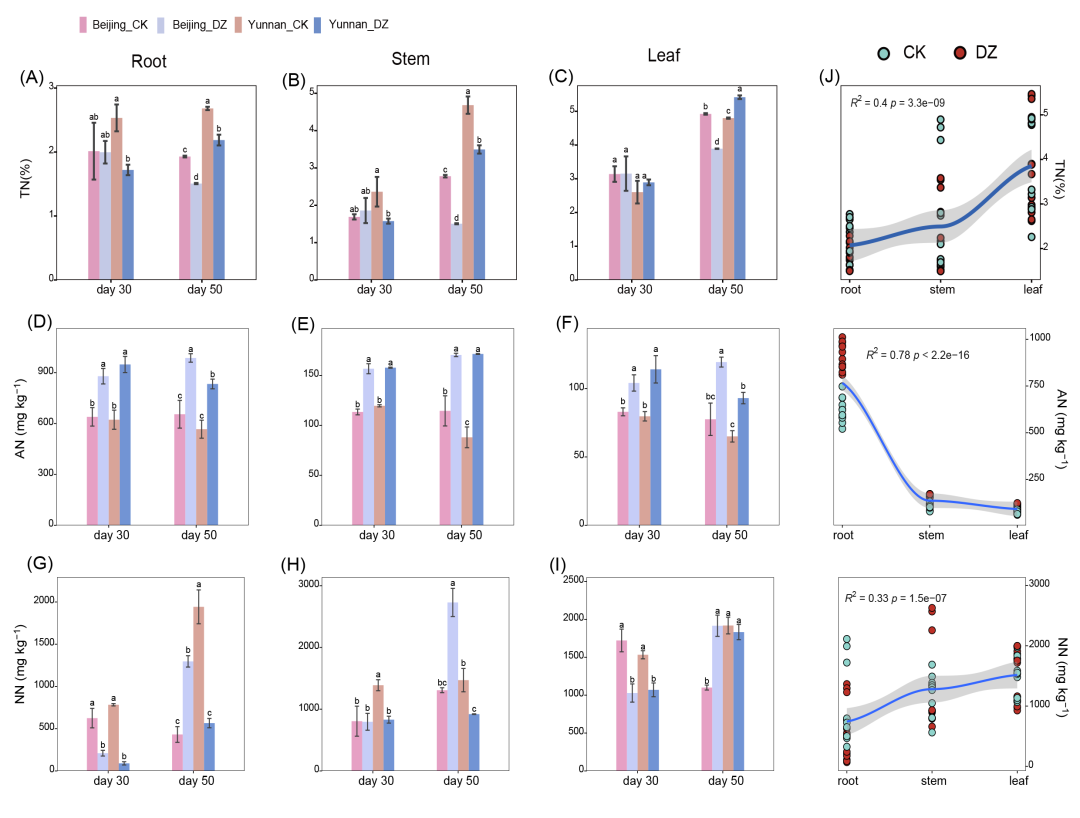
图1. 总氮 (TN)、铵态氮 (AN) 和硝态氮 (NN) 的时空分布
根(A)、茎(B)或叶(C)中 TN 的变化。根(D)、茎(E)或叶(F)中 AN 的变化。根(G)、茎(H)或叶(I)中 NN 的变化。各组的显著性用小写字母表示,不同字母代表显著差异。(J) 三种氮形态与空间分布的 Spearman 相关性。
DZ 熏蒸后不同分室中氮代谢酶动态变化
在北京和云南土壤中,总氮(TN)的减少与根、茎和叶中亚硝酸盐还原酶(NiR)、谷氨酰胺合成酶(GS)和谷氨酸合成酶(GOGAT)活性的升高一致(图 2),表明土壤熏蒸诱导的这些酶活性刺激了 TN 的减少。此外,两种土壤中铵态氮(AN)水平的升高与 NiR、硝酸还原酶(NR)、GS 和 GOGAT 活性的增强有关(图 2)。然而,硝态氮(NN)的反应在两种土壤之间存在显著差异:在北京土壤中,硝态氮与干重和 GOGAT 活性呈正相关,而在云南土壤中,硝态氮与某些酶(GS、NR、GOGAT)呈负相关(图 2)。总之,土壤熏蒸显著影响烟草氮酶活性的动态变化。酶活性的增加始终与 TN 的减少和 AN 的增加相关。然而,土壤类型不同,氮代谢酶的响应也不同,这凸显了土壤特性、氮素形态和氮代谢酶活性之间复杂的相互作用。
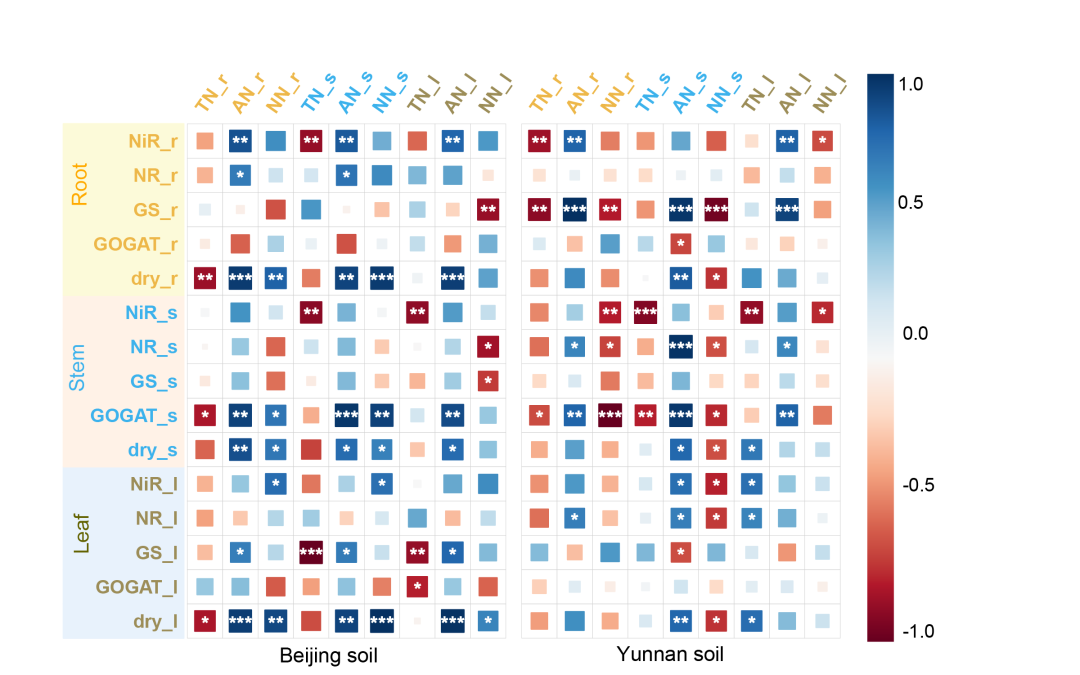
图2. 北京和云南土壤中烟草根、茎和叶的总氮(TN)、铵态氮(AN)和硝态氮(NN)与氮代谢酶之间的相关性热图
蓝色表示正相关,红色表示负相关。方框内的星号表示显著性水平:p < 0.05,p < 0.01,*p < 0.001。
DZ 熏蒸后土壤氮动态变化及微生物响应
DZ熏蒸对北京和云南土壤的氮素产生显著影响,增加AN而减少NN,影响时间超过50天(图3和S2)。在第 50 天,熏蒸土壤中的 AN 含量增加了 193%-771% (p < 0.05)(图 3A),而 NN 含量减少了 29.20%~41.40% (p < 0.05)(图 3B),这种变化在云南土壤中更为明显。土壤氮素形态和含量的变化进一步导致根际氮供应速率发生了显著变化。具体来说,北京和云南的 AN 供应率分别增加了 250% 和 807%(p < 0.05),而 NN 供应率则分别显著下降了 82.47% 和 45.16%(p < 0.05)(图 3C 和 3D)。因此,土壤硝化潜力下降了 72.91~86.51%(p < 0.05)(图 3E),反硝化潜力增加了 197~324%(p < 0.05)(图 3F)。这些发现与先前的研究结果一致,即熏蒸强烈抑制硝化作用,同时提高反硝化作用。
熏蒸后(第 0 天),微生物群落多样性和基于 KEGG 的功能多样性均显著增加(图 S1A),然而在熏蒸后期(第 50 天)熏蒸组和对照组没有显著差异(图 S1B)。PCA 结果显示物种分类和功能基因组成存在明显差异(图 S1C 和 S1D),表明熏蒸选择性地富集了特定物种和功能通路。
在DZ熏蒸的云南土壤中,重点关注了硝化、反硝化、硝酸盐还原、氨氧化、固氮及有机氮代谢等氮转换过程(图3G和S2C)。最初,只有反硝化作用受到熏蒸的显著刺激,增加了 nirS 和 norB 基因的丰度(图 S2C 和表 S3)。第 50 天时,大多数参与反硝化的基因如 nar 家族、nirK/S、norB 和 nosZ 均显著增加,而氨氧化基因如 amoC 则有所减少(图 3G)。基因丰度的变化与土壤硝化、反硝化率一致(图 3E 和 3F),这表明熏蒸促进了反硝化微生物的富集。
在熏蒸 50 天的土壤中,变形菌门显著富集(图 S2D 和 3H),反硝化类群如鞘氨单胞菌科、拟杆菌科显著增加(图 3I)。例如,拟杆菌科增加了 127% (p < 0.05),棒状杆菌科增加了 150% (p < 0.05),鞘氨单胞菌科增加了 113% (p < 0.05)(图 S2G)。此外,鞘氨单胞菌在熏蒸后第 0 天增加了 43.81% (p < 0.05),第 50 天增加了 147% (p < 0.05),表明熏蒸对该属有持续的刺激作用(图 S2F 和 S2G)。

图3. 第 50 天时土壤氮转化特征和微生物响应
(A) 氨态氮(AN),(B) 硝态氮(NN),(C) 根际氨态氮供应速率,(D) 根际硝态氮供应速率,(E) 根际土壤硝化潜力,(F) 根际土壤反硝化潜力。(G)与氮循环途径相关的功能基因变化概览。彩色线条代表不同途径,沿线标注了负责各途径的基因。与对照相比,基因上调明显的用红色标出,下调明显的用蓝色标出,无明显变化的用黑色标出。*表示显著性水平。(H) LEfSe 多级物种层次图。不同颜色的节点表示在相应处理组中显著富集的微生物群。浅黄色节点表示组间无明显差异或影响的微生物群。(I) LDA 判别直方图。LDA 分数越高,表明物种丰度对差异效应的影响越大。
DZ 熏蒸刺激作物氮代谢基因表达
与茎相比,根和叶富含更多具有特定功能的代谢途径。例如,根部富集了细胞壁生物合成、纤维素代谢、木聚糖生物合成、木质素分解、苯丙素分解、次生细胞壁生物发生及原生细胞壁生物合成等途径(图 S3A)。而叶片则主要富集了与光系统 II 氧发生复合体、光合作用调节、含卟啉化合物生物合成有关的途径(图 S3C)。结果显示,熏蒸导致根部 1497 个基因和叶片 614 个基因显著上调,而茎部只有 6 个基因显著上调(图 4A-4C)。这些结果表明,根部是作物的功能热点,熏蒸对根部基因表达的影响最大,其次是叶片和茎。我们从转录组数据中筛选了参与硝酸盐转运、铵转运、氨基酸转运、硝酸盐还原、铵代谢和氮调节的基因表达情况,同时分析了这些基因的表达水平与根、茎和叶中氮含量的关系(图 4D)。在根中,较低的总氮(TN)和硝酸盐氮(NN)水平以及较高的铵态氮(AN)水平与特定的硝酸盐转运体(NRT1.1、NRT1.2、NRT1.6 和 NRT2. 7)、铵转运体(AMT1.1、AMT3.1)、氨基酸转运体(AAP3、AAP6 和 LHT1)、谷氨酸合成酶(GS-N1)、谷氨酰胺合成酶(GOGAT1、Fe-GOGAT2)和氮调节基因(NAC48)有关。在茎中, TN 和 NN 水平与 NRT1.2、NRT1.5、NRT2.6、NRT3.1、NRT3.2、AMT3.1、AAP6 和 LHT1 呈负相关,而这些基因与 AN 呈正相关。在叶片中,NN 的减少或 AN 的增加与 NRT1.1、NRT1.6、NRT1.8、NRT2.5、NRT3.2、AMT1.1、AMT1.2、AMT3.1、GS-N1、Fe-GOGAT、NLP7、NLP14、NLP41 和 NLP48 有关。熏蒸诱导的基因表达和氮分布变化在根部最为显著,根部的许多氮转运体和代谢基因被上调,导致氮含量发生明显变化。这表明根部在调节氮素代谢中起着关键作用,其次是叶片,因此根部是提高作物氮利用率的重要目标。
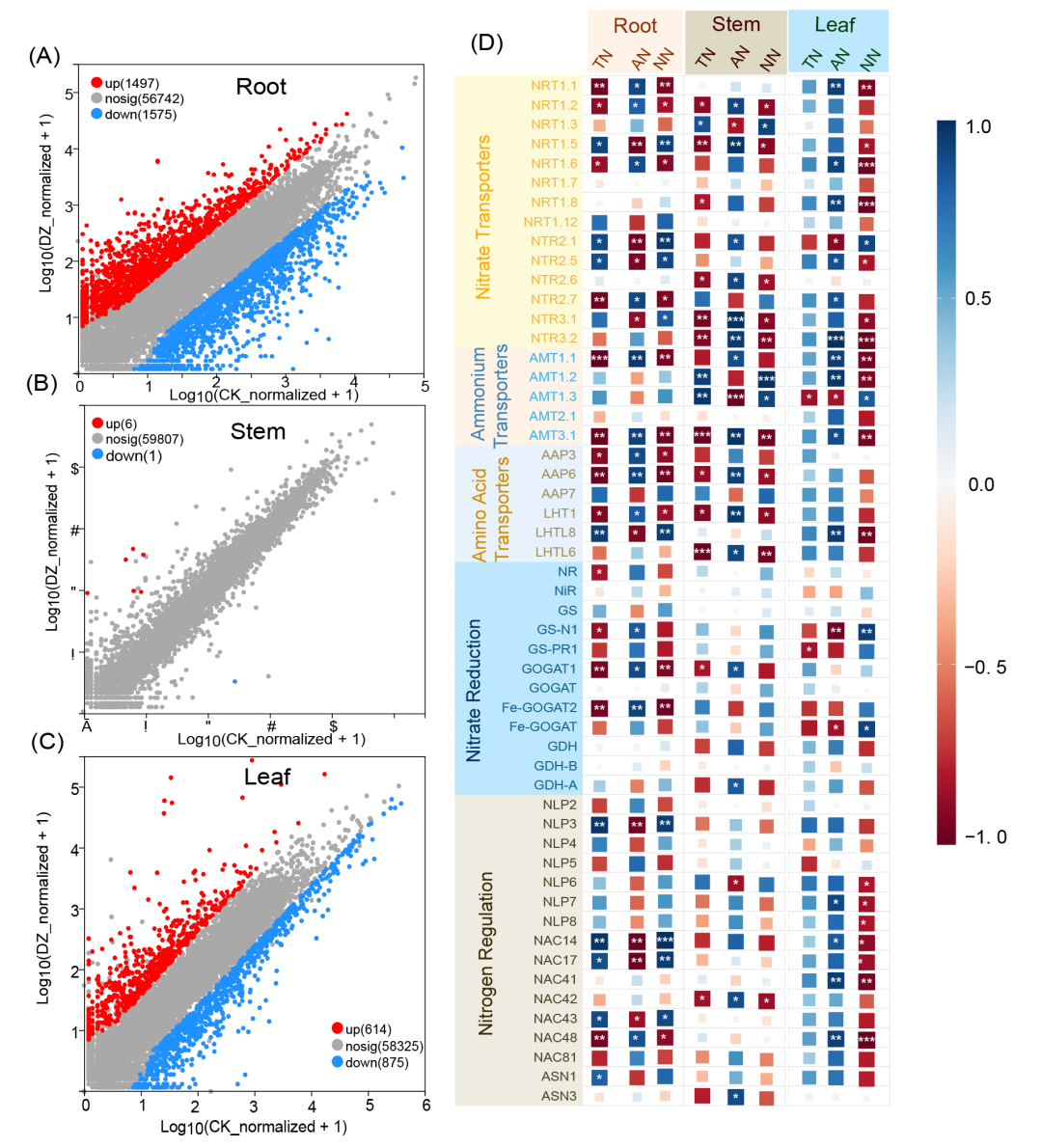
图4. 基因表达特征
散点图显示了根(A)、茎(B)和叶(C)的基因表达差异,横轴和纵轴分别为对照组和处理组的对数转换表达水平。红点表示明显上调的基因,绿点表示明显下调的基因,灰点表示差异不明显的基因。离原点较近的点表示表达水平较低,而离对角线较远的点表示两个样本之间基因表达差异较大。(D) 热图显示云南土壤中根、茎或叶的全氮(TN)、铵态氮(AN)和硝态氮(NN)与氮代谢基因表达矩阵之间的相关性。蓝色表示正相关,红色表示负相关。方框内的星号表示显著性水平:p < 0.05,p < 0.01,*p < 0.001。
驱动氮素空间分布的关键因素
结构方程模型结果表明,氮相关微生物群落是调节土壤铵态氮和硝态氮水平的主要因素,这种影响在根部尤为显著。具体来说,烟草体内铵态氮的积累及硝态氮水平的降低,是由微生物参与的硝化和反硝化过程间接实现的(图 5G-5I)。微生物群落直接对土壤铵态氮具负调控及对土壤硝态氮具正调控,改变了根、茎和叶的总氮和硝态氮水平,但对叶的影响不明显。此外,氮基因表达也直接受到氮相关微生物群落的调控,但不受土壤中铵态氮和硝态氮水平的调控(图 5B、5F、5H)。总体效应分析表明,氮相关微生物群落对根、茎、叶的总氮水平有负向调节作用,但对硝态氮有正向作用,尤其是根部更显著(图 S4)。这些结果表明,DZ熏蒸显著改变了氮循环微生物群落的结构,进而影响了土壤中铵态氮和硝态氮的供应,这种改变导致氮转运和代谢基因表达发生变化,最终影响氮在根茎、叶中的分布。
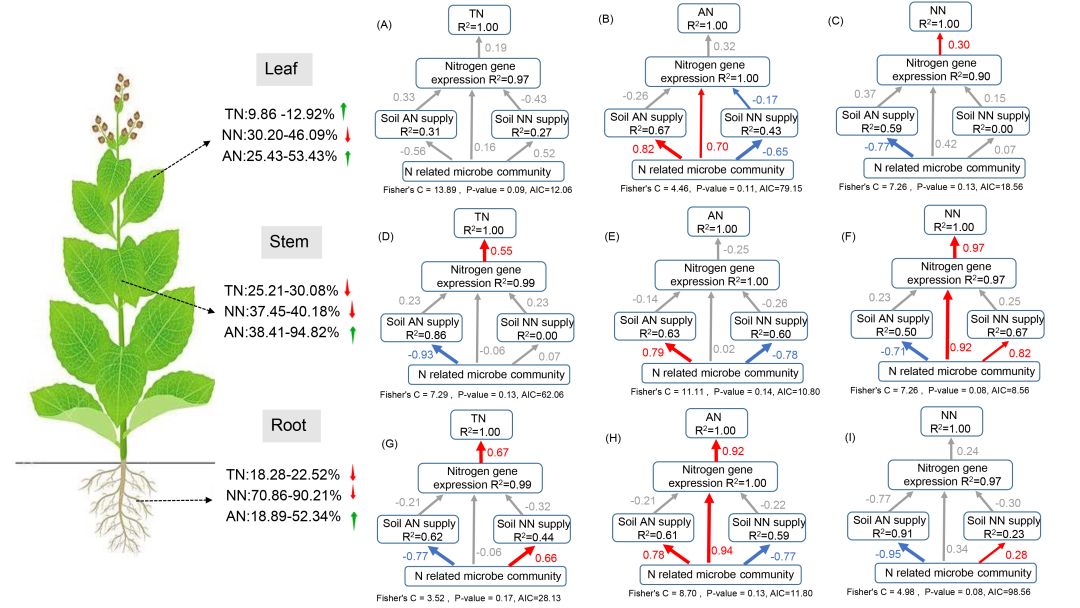
图5. 土壤微生物群落与氮在不同植物区系中分布关系的路径图
图(A)-(C)表示叶片中的总氮(TN)、铵态氮(AN)和硝态氮(NN);图(D)-(F)表示茎中的总氮、铵态氮和硝态氮;图(G)-(I)表示根中的总氮、铵态氮和硝态氮。蓝色箭头表示负相关,红色箭头表示正相关。箭头旁边显示的是标准化路径系数,箭头大小与系数强弱成正比。蓝色或红色系数表示关系显著(p < 0.05),而灰色箭头和系数表示关系不显著(p > 0.05)。R² 值表示每个变量解释的变异比例。与对照组相比,熏蒸处理下根、茎和叶中的 TN、AN 和 NN 百分比变化情况,向上和向下箭头分别表示增加和减少。
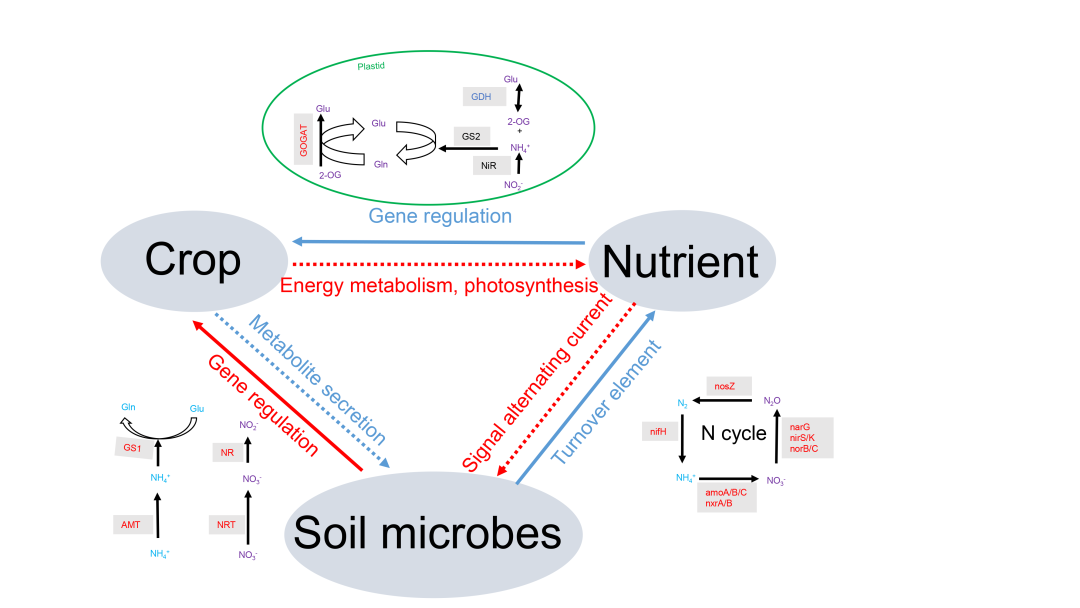
图6. 作物-微生物-养分相互作用的概念模型
熏蒸通过改变微生物群落结构,从而影响土壤养分周转,改变养分类型和水平。这些变化会影响作物基因表达和养分吸收。作为回应,作物通过光合作用调节养分分布,从而间接影响微生物的活动。此外,作物还能通过根部渗出物直接改变微生物群落,进一步影响土壤养分动态。
讨 论
DZ 熏蒸刺激土壤氮循环微生物群落生长
DZ熏蒸显著提高了特定微生物的定殖,在氮循环微生物中,反硝化微生物如鞘氨醇单胞菌和与反硝化相关的功能基因包括 nirS、nosZ、nirK、nar 和 norB,在熏蒸后均明显增加(表 S3)。同时反硝化强度也大幅上升(图 3F),表明熏蒸显著刺激了土壤中的反硝化过程。其他熏蒸剂,如氯化苦、1,3-二氯丙烯和二甲基二硫,也不同程度刺激反硝化微生物。在早先的一项研究中,我们发现 DZ 熏蒸暂时降低反硝化功能基因(napA、narG、nirK、nirS、cnorB、qnorB 和 nosZ)的丰度,尤其是在酸性红壤中更明显。在本研究中,在熏蒸后种植作物的土壤中这些基因的丰度明显高于对照土壤(图 3G),表明作物的存在促进了熏蒸后的微生物演替。例如,氯化苦熏蒸后,种植番茄土壤中的细菌群落恢复速率比未种植番茄土壤恢复得更快。这表明,熏蒸诱导的微生物群落变化可以促进作物生长。
DZ 熏蒸促进反硝化微生物繁殖可能的机制包括:(1)有机物供应增加:对熏蒸敏感的微生物死亡后释放出大量可溶性蛋白质,促进微生物快速生长。熏蒸后,土壤中的可溶性氨基酸增加了 1.99~2.32 倍。此外,熏蒸剂本身也是一种碳源。例如,异硫氰酸甲酯的降解半衰期从几小时到几天不等,生物降解占总降解量的 80%~90% 。(2) 减少竞争压力:熏蒸可杀死或抑制许多病原微生物和竞争微生物,通过产生抗生素或其他抑制物质来减少病原体数量,这反过来又有利于反硝化细菌的生长。此外,DZ 熏蒸减少了硝化细菌丰度(如氨氧化细菌AOB和氨氧化古细菌AOA) 的数量,它们能将氨(NH₃)或铵(NH₄⁺)转化为亚硝酸盐(NO₂-),而亚硝酸盐浓度较高时会抑制反硝化细菌的活性。(3) 更强的微生物群落恢复能力:熏蒸对微生物群落具有重构作用,后期微生物开始逐步恢复,反硝化细菌等适应性强的微生物迅速重新占据生态位。研究表明,与硝化细菌相比,反硝化细菌对熏蒸引起的变化表现出更强的抵抗力和复原力。
土壤微生物驱动的氮分布机制
熏蒸扰乱土壤微生物群落后,通过抑制氨氧化和促进反硝化来改变土壤生态功能,导致铵态氮的积累和硝态氮的减少,破坏了土壤氮循环的平衡(图 3E 和 3F)。熏蒸引起的土壤氮平衡破坏极大地改变了植物组织(包括根、茎和叶)对氮的吸收和分布(图 5)。这种氮的重新分配与关键氮代谢和转运基因的特异性表达密切相关。与氮转运和同化有关的多个基因的表达明显升高,这些基因包括与氮吸收、转运和代谢相关的基因,它们表达量的上调表明作物体内氮的利用率提高了(图 4D)。这反映了氮相关过程在基因水平发生了重大变化,是观察到的氮素分布变化的分子基础,凸显了植物氮素平衡响应土壤熏蒸的复杂调控机制。
熏蒸后氮在植物地上组织的重新分配主要归因于根、茎、叶中氮代谢基因表达水平的变化。值得注意的是,作物易吸收的硝态氮也是一种信号分子,可刺激根部氮转运基因的表达,从而调节氮的吸收和转运。结构方程模型表明,虽然没有观察到土壤氮对基因表达的直接调控,但土壤微生物群落的变化对氮代谢基因在各组织中的表达有显著影响(图 5)。这凸显了土壤微生物在介导作物养分吸收和运输方面的关键作用。微生物群落对土壤养分循环至关重要,土壤养分含量和形态的改变会直接影响作物基因的表达,进而影响养分的吸收和代谢(图 6)。
土壤熏蒸改变作物与微生物相互作用的潜在机制包括:(1) 加强了有机物的分解和土壤微生物介导的矿化过程,从而增加了植物对可溶性氨基酸、磷和氮等关键养分的吸收。(2) 熏蒸改善土壤结构,优化根系环境提高了水分和养分的吸收效率。(3) 熏蒸引起的微生物群落结构变化可能刺激作物生长素、细胞分裂素和赤霉素等激素的产生,进而调节根系发育、幼芽生长和养分运输。这些机制共同作用使作物能够更有效地利用土壤资源,优化地上组织的养分分配,并提高植物的整体生长和产量。然而,要充分了解土壤微生物、植物内生菌和养分代谢之间复杂的相互作用,还需要进一步借助同位素标记等技术进行深入研究。
结 论
棉隆土壤熏蒸导致作物根、茎和叶中的铵态氮增加,硝态氮减少,从而显著提高了氮代谢。熏蒸引起的微生物群落变化,特别是氨氧化微生物的减少和反硝化微生物的增加,加剧了土壤氮循环过程。此外,微生物群落的变化影响作物氮代谢和转运蛋白基因的表达,从而促进氮从土壤向作物地上部有效的转运和利用。本研究阐明了土壤氮动态与作物氮代谢之间的联系,为优化作物养分管理提供了新的视角。
方 法
土壤样品制备和熏蒸
土壤样品采集自北京房山(北纬 39.7077°,东经 115.9605°)和云南文山(北纬 23.3681°,东经 104.2493°),前者代表碱性潮土,后者代表酸性红壤。土壤的理化性质详见表 S1。采集的土壤过 2 mm筛,并调整至绝对含水量为 13%。实验前,土壤中添加了硝酸钠、磷酸钾和氯化钾,最终浓度分别为 200 mg/Kg N、150 mg/Kg P和 150 mg/Kg K。
熏蒸时,将棉隆(98%颗粒剂,江苏南通施壮化工有限公司)与土壤混合,剂量为 150 mg/Kg。然后将土壤放入用凡士林密封的干燥器中进行熏蒸,以确保密闭性,并在 28°C 的黑暗环境中培养 7 天。此外,还设立了不使用棉隆的对照组。熏蒸结束后,在通风橱中通风散气。收集土壤样本,并将其保存在 4°C 和 -80°C(指定为熏蒸后第 0 天),以测定土壤化学参数和微生物指标
烟草移植和样本采集
将烟草种子(由中国农业科学院烟草研究所提供)播种在塑料培养箱中。当幼苗长到三叶期时,选择生长均匀的幼苗。在移栽到熏蒸土和非熏蒸土中之前,先用无菌水清洗根部。烟草被移植到单个花盆中,每个花盆装 1 Kg土。将花盆放置温室中:温度为 28°C,相对湿度为 60%,光照时间为 12 h光照/12 h黑暗,熏蒸后第 30 天和第 50 天进行采样。收集烟草样本时,将整株作物连根拔起,用蒸馏水清洗根、茎和叶,然后晾干。将根、茎和叶分开;部分样本在 55°C 下烘干至恒重,用于测定干重和总氮。其他样本放入预冷离心管中,在液氮中速冻 5 分钟,然后保存在 -80°C,用于测定无机氮浓度、氮代谢酶活性和转录组分析。在根际土壤取样时,松散附着的土壤被抖落,紧密附着的土壤被刷落并收集;部分样本用于土壤氮转化测定,其他样本用于宏基因组测序。
土壤氮相关参数测定
土壤理化性质按照标准方法测定。有效磷用 NaHCO3 提取,用流动分析仪分析;速效钾用 CH3COONH4 提取,用火焰光度计分析;pH 值用电位计按 2.5:1 的水土比测定;电导率按 2.5:1 的水土比测量;有机质用重铬酸钾滴定法测定。硝态氮和铵态氮用氯化钾提取,并用流动分析仪测量。
通过将 10 g新鲜土壤放入装有去离子水的 20 ml顶空瓶中,密封后在 28°C 黑暗中培养,用于测量反硝化的潜力。采用氯化钾提取法测定 0 h和 24 h的硝态氮浓度,并根据单位时间内的硝态氮消耗量计算反硝化强度,计算公式如下:
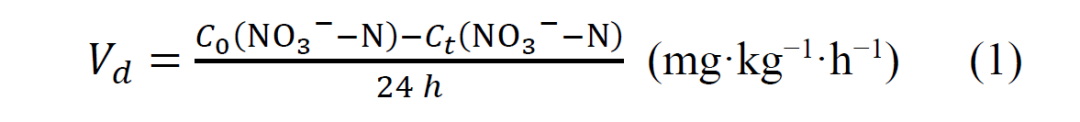
测定硝化潜力的方法是将 10 g新鲜土壤放入 20 ml顶空瓶中,用无菌棉花密封,在 28°C 黑暗环境中培养,并在 0 h和 24 h后用氯化钾提取法测定铵态氮浓度。硝化强度按单位时间内的铵态氮消耗量计算,公式如下:
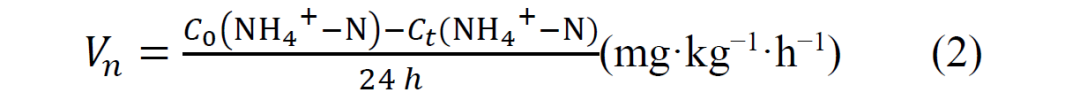
根系中铵态氮和硝态氮的供应率是通过作物根际有效氮供应模型(3)和(4)计算得出的:
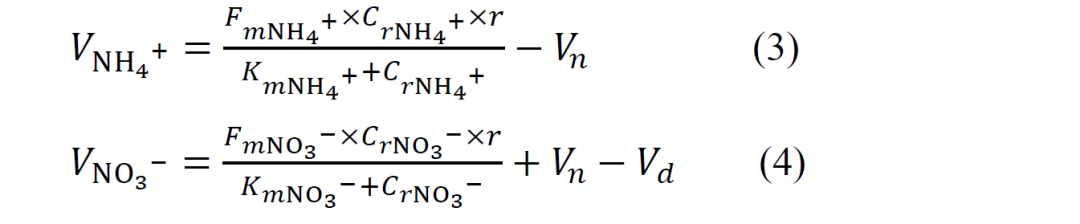
VNH4+别VNO3-分别代表根瘤土壤中铵态氮和硝态氮的供给率(mg·kg−1·h−1)。Cr 表示根际土壤中铵态氮和硝态氮的浓度(mg·kg−1·h−1)。Vn和Vd分别指硝化率和反硝化率(mg·kg−1·h−1)。参数 r 设为 0.1 mm,代表根系吸收氮的半径。Fm 和 Km 分别表示铵态氮和硝态氮进入根系的最大速率,以及铵态氮和硝态氮在土壤中的传导消耗常数。
植物氮和酶活性测定
根、茎和叶样品于液氮中研磨后,用氯化钾提取,然后用流动分析仪测量其无机氮含量。总氮的测定方法是将样品烘干并粉碎,然后用元素分析仪在 CN 模式下分析。
使用 BCA蛋白质测定试剂盒(Thermo Scientific)测定硝酸还原酶(NR)、亚硝酸还原酶(NiR)、谷氨酰胺合成酶(GS)和谷氨酸合成酶(GOGAT)的活性,并在 450 nm波长下读取吸光度。
转录组测序和分析
使用 RNAprep Pure Plant Kit 提取根、茎和叶的总 RNA,并使用琼脂糖凝胶电泳和 Nanodrop 2000 检测其质量和纯度。高质量 RNA 用于合成 cDNA 文库,通过末端修复、A-尾链接、适配器连接和 PCR 扩增构建 cDNA 文库。测序在 Illumina NovaSeq 6000 平台上进行 (上海美吉生物医药科技有限公司)。使用 fastp (v0.20.0)对原始数据进行过滤,去除接头、含有超过 10% 未知碱基(N)的读数和低质量读数(Q 值≤ 20)。经 FastQC(v0.11.9)质检后,用 Trinity(v2.11.0)进行等位基因组装以获得单基因。基因注释使用 BLASTX 将单基因与蛋白质数据库(包括 NR、Swiss-Prot 和 KEGG)进行比对。使用 RSEM(v1.3.3)计算基因表达水平,并使用 DESeq2(v1.28.1)识别差异表达基因,设置阈值为 |log2 fold change| ≥ 1 且调整后的 p 值小于 0.05。
宏基因组测序和分析
使用 E.Z.N.A.® Soil DNA Kit 提取土壤总 DNA,并使用琼脂糖凝胶电泳和分光光度计 (Nanodrop 2000)检测其质量。对DNA 进行片段化、末端修复、连接和 PCR 扩增,构建测序文库,在 Illumina NovaSeq 平台 (上海美吉生物医药科技有限公司)上进行测序。使用 fastp (v0.20.0)对原始数据进行质检后,使用MEGAHIT (v1.1.2)对其进行组装。采用 MetaGeneMark (v3.38)进行基因预测,并使用 CD-HIT (v4.6)对相似度为 95% 的基因进行聚类,从而构建非冗余基因集。物种注释基于NR 数据库的比对结果,功能注释来自 KEGG、eggNOG 和 CAZy 数据库。
数据统计分析
采用 Tukey ANOVA分析熏蒸后根、茎、叶不同部位总氮、铵态氮和硝态氮变化的显著性。采用spearman相关系数计算氮形态与空间位置之间的相关性。利用 R语言 (4.4.1)中 Hmisc包绘制氮与酶活性相关性热图。使用 Goatools进行 GO 富集分析,并通过 Fisher 检验确定显著变化的 GO 功能。采用线性判别分析 (LDA)估计各物种丰度对差异效应值,LDA阈值设定为大于 3.5。结构方程模型 (SEM)借助piecewiseSEM包来实现,用于比较土壤微生物群落对不同区系氮分布的直接和间接影响。
代码和数据可用性
本研究的转录组(CRA021448, https://bigd.big.ac.cn/gsa/browse/CRA021448)和宏基因组(CRA021339, https://bigd.big.ac.cn/gsa/browse/CRA021339)原始数据已保存在中国科学院北京基因组研究所国家生物信息中心国家基因组数据中心补充材料(图、表、图表摘要、幻灯片、视频、中文翻译版本和更新资料)也可从在线DOI或iMeta Science(http://www.imeta.science/imetaomics/.)获取。
引文格式:
Wensheng Fang, Wenfeng Tian, Dongdong Yan, Yuan Li, Aocheng Cao, Qiuxia Wang. 2025. "Linkages between soil nutrient turnover and above-ground crop nutrient metabolism: The role of soil microbes. iMetaOmics e55. https://doi.org/10.1002/imo2.55
作者简介

方文生(第一作者)
● 中国农业科学院植物保护研究所 副研究员。
● 研究方向为土壤消毒防控土传病害作用机理研究,以第一作者/通讯作者在Journal of Hazardous Materials,Soil Biology and Biochemistry等发表论文18篇,累积被引1547次(谷歌学术)。

王秋霞(通讯作者)
● 中国农业科学院植物保护研究所研究员,博士生导师,中国农业科学院“土壤有害生物防控”创新团队首席科学家。
● 研究方向为土壤消毒技术与应用原理研究。近五年承担重点研发计划项目等7项,参与创制了系列土壤消毒装备,大幅度提升了土壤消毒效率、效果与稳定性。作为主要完成人之一构建了以土壤消毒技术为核心的土传病害防控技术体系,可有效破解作物连作障碍,并成功入选农业农村部主推技术,产业扶贫效果显著。获中华全国妇女联合会巾帼文明岗称号(第1)及中国农科院杰出科技创新奖(第2)、北京市科技进步二等奖(第2),发表科技论文76篇,其中以第一或通讯作者发表论文34篇,主编著作2部、参与制定标准4项。
iMetaOmics
更多资讯
● iMeta姊妹刊iMetaOmics(定位IF>10)欢迎投稿!(2024.2.27)
● iMeta姊妹刊iMetaOmics编委招募 (定位IF>10) (2024.3.2)
● iMeta姊妹刊iMetaOmics电子版和印刷版ISSN申请获批(2024.4.1)
● iMeta姊妹刊iMetaOmics投稿系统正式上线(2024.4.17)
● iMeta姊妹刊iMetaOmics主编正式官宣(2024.4.22)
● 出版社iMetaOmics主页正式上线!(2024.4.28)
● iMetaOmics | 浙江大学宗鑫组揭示两猪种宿主-肠道菌群互作差异
● iMetaOmics | 罗鹏/袁硕峰/苗凯/程全发表STAGER: 生成式人工智能可靠性的标准化测试和评估推荐
● iMetaOmics | 徐州医科大杨欢组揭秘沙门氏菌-宿主-微生物群在免疫与代谢中的相互作
● iMetaOmics | 中科院动物所金坚石组综述16S rRNA基因扩增子测序技术的“前世今生”
● iMetaOmics | 浙大张天真组完成二倍体棉种泛基因组构建
● iMetaOmics | 张勇/李福平-先进糖蛋白组学在男性生殖研究中的潜在应用
● iMetaOmics | 暨南大学潘永勤/杨华组-炎症蛋白联合检测利于诊断甲状腺乳头状癌和结节性甲状腺肿
● iMetaOmics | 张开春组利用多组学方法揭示甜樱桃加倍后果色变化的候选基因
● iMetaOmics | 杜娟/林婷婷-慢性泪囊炎患者眼部菌群类型和纵向菌群变化
● iMetaOmics | 陈汉清/陈俊综述有关肝细胞癌治疗的新兴纳米医学策略
● iMetaOmics | 基因组所刘永鑫/卢洪评述微生物在提高杂种优势中的作用
● iMetaOmics | 上科大刘雪松组开发基于通路的肿瘤细胞鉴别工具TCfinder
● iMetaOmics | 中山大学刘鹏/邹宇田-整合人工智能实现HER2阳性乳腺癌精准管理
● iMetaOmics | 安徽农大李晓玉组-丛枝菌根真菌对玉米内生菌群的影响
● iMetaOmics | 徐涛/黄蓉/苏国海-急性冠脉综合征纵向多组学队列建设
● iMetaOmics | 通过整合宏组学促进人类与环境健康发展
● iMetaOmics | 苏州大学林俊组-揭示活性微生物及益生元/益生菌与关节炎联系
● iMetaOmics | 中国药科大学徐文波开发叶绿体基因组数据分析软件
● iMetaOmics | 清华刘晓组和复旦王久存组揭示特定细菌在皮肤老化中的作用
●iMetaOmics | 中南大学夏晓波团队揭示青光眼和SLE发病机制新关联
●iMetaOmics | 庐山植物园刘芬组揭示了自噬在植物-根微生物互作机制中的调控作用
●iMetaOmics | 杨瑞馥/袁静综述微生物组与“同一健康”的联系
●iMetaOmics | 同济/上海交大-开发支持群体分组分析的宏基因组测序综合分析软件
●iMetaOmics | 陈绍鸣-关于靶向NF-κB的潜伏逆转剂及其在HIV潜伏期的表观遗传和突变影响的评论
● iMetaOmics | 甘肃农大刘自刚组-强抗寒甘蓝型冬油菜的基因组组装和基因组特征解析
● iMetaOmics | 南京农大朱伟云组-外周血清素在结肠稳态中的作用
● iMetaOmics | 魏来/贾慧珏/何明光-多组学助力揭示塑造转录组的基因型-微生物组相互作用
● iMetaOmics | 徐州医科大学朱作斌组-微生物对寿命的调节:机制和治疗策略
● iMetaOmics | 白立景/邢凯组-解析脊椎动物肠道微生物多样性的影响因素
● iMetaOmics | 刘永鑫/陈同-用于食物微生物组成和时间序列研究的微生物组数据库FoodMicroDB
● iMetaOmics | 重庆大学王贵学组-肠道微生物细胞外囊泡在神经退行性疾病中的新作用及其治疗策略
● iMetaOmics | 四川大学王红宁组-解析产气荚膜梭菌的基因组宿主适应性
● iMetaOmics | 北京协和医院杨启文组-ramR基因突变增强免疫激活和依拉环素耐药性
● iMetaOmics | 香港中文苏奇组-抗菌多肽开发中的见解: 一个多学科视角的观察
● iMetaOmics | 上科大刘雪松组开发CD4 TCR特异性预测工具Pep2TCR
● iMetaOmics | 江苏省农科院植物细菌团队-解析中国梨火疫菌特征及溯源分析
● iMetaOmics | 基因组所刘永鑫组-易扩增子(EasyAmplicon):用户友好的扩增子测序数据分析指南
● iMetaOmics | 东京科学大学奥村学组-Hyena架构蛋白质语言建模
● iMetaOmics | 兰大南志标/段廷玉组-丛枝菌根网络影响邻近植物对病原菌的响应
● iMetaOmics | 汝之蜜糖彼之砒霜:源自益生菌LGG的研究证据
● iMetaOmics | 中国农大汪杰组-解析微塑料胁迫下玉米的分子响应
● iMetaOmics | 陈嘉莉/唐少军-奶源动物双歧杆菌乳亚种的膳食健康
● iMetaOmics | 西北农林曹阳春组综述艰难梭菌感染病理机制及饮食模式对其影响策略
● iMetaOmics | 宁波大学叶央芳组-解析驱动蟹肠道菌群稳定性的关键物种
● iMetaOmics | 中国农科院毕研亮组-精确消化道微生物群调节策略促进宿主健康
● iMetaOmics | 深圳大学李猛组-沉积物中古菌的多样性和代谢潜能
● iMetaOmics | 华东理工叶邦策组-多组学分析分枝杆菌侵染宿主过程
● iMetaOmics | 中国农业大学曹志军组-MicroRNA-微生物群互作调控宿主健康
更多推荐
(▼ 点击跳转)
iMeta | 引用16000+,海普洛斯陈实富发布新版fastp,更快更好地处理FASTQ数据
iMeta | 兰大张东组:使用PhyloSuite进行分子系统发育及系统发育树的统计分析
iMeta | 唐海宝/张兴坦-用于比较基因组学分析的多功能分析套件JCVI

1卷1期

1卷2期

1卷3期

1卷4期

2卷1期

2卷2期

2卷3期

2卷4期

3卷1期

3卷2期

3卷3期

3卷4期

3卷5期

3卷6期

1卷1期

1卷2期
期刊简介
“iMeta” 是由威立、宏科学和本领域数千名华人科学家合作出版的开放获取期刊,主编由中科院微生物所刘双江研究员和荷兰格罗宁根大学傅静远教授担任。目的是发表所有领域高影响力的研究、方法和综述,重点关注微生物组、生物信息、大数据和多组学等前沿交叉学科。目标是发表前10%(IF > 20)的高影响力论文。期刊特色包括中英双语图文、双语视频、可重复分析、图片打磨、60万用户的社交媒体宣传等。2022年2月正式创刊!相继被Google Scholar、PubMed、SCIE、ESI、DOAJ、Scopus等数据库收录!2024年6月获得首个影响因子23.8,位列全球SCI期刊前千分之五(107/21848),微生物学科2/161,仅低于Nature Reviews,学科研究类期刊全球第一,中国大陆11/514!
“iMetaOmics” 是“iMeta” 子刊,主编由中国科学院北京生命科学研究院赵方庆研究员和香港中文大学于君教授担任,是定位IF>10的高水平综合期刊,欢迎投稿!
iMeta主页:
http://www.imeta.science
姊妹刊iMetaOmics主页:
http://www.imeta.science/imetaomics/
出版社iMeta主页:
https://onlinelibrary.wiley.com/journal/2770596x
出版社iMetaOmics主页:
https://onlinelibrary.wiley.com/journal/29969514
iMeta投稿:
https://wiley.atyponrex.com/journal/IMT2
iMetaOmics投稿:
https://wiley.atyponrex.com/journal/IMO2
邮箱:
office@imeta.science


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









