






点击蓝字 关注我们
限制时间饮食通过假长双歧杆菌-丙酸-FFAR3通路减缓阿尔兹海默相关的认知功能损伤

iMeta主页:http://www.imeta.science
研究论文
● 原文: iMeta (IF 23.8)
● 原文链接DOI: https://doi.org/10.1002/imt2.70006
● 西北农林科技大学刘志刚、刘学波以及陕西师范大学施琳、北京大学深圳医院胡俊等在iMeta在线联合发表了题为“Time-restricted feeding mitigates Alzheimer's disease-associated cognitive impairments via a B. pseudolongum-propionic acid-FFAR3 axis”的文章。
● 本研究首次开展了时间限制饮食(TRF)对阿尔兹海默病(AD)患者的临床干预,发现TRF具有显著的神经保护作用,确定了假长双歧杆菌(B. pseudolongum)和丙酸(PA)在AD发病机制中的核心作用。
● 第一作者:赵忆航、贾梦真
● 通讯作者:刘志刚(zhigangliu@nwsuaf.edu.cn)、刘学波(xueboliu@nwsuaf.edu.cn)、施琳(linshi198808@snnu.edu.cn)、胡俊(dochj@163.com)
● 合作作者:丁辰、鲍炳昆、李航棋、马嘉彬、董伟璇、高蕊、陈旭辉、陈娇、代晓霜、邹远强
● 主要单位:西北农林科技大学食品科学与工程学院、西安交通大学第一附属医院、北京大学深圳医院、Xbiome深圳未知君生物科技有限公司、华大集团、陕西师范大学、西北农林科技大学深圳研究院
亮 点
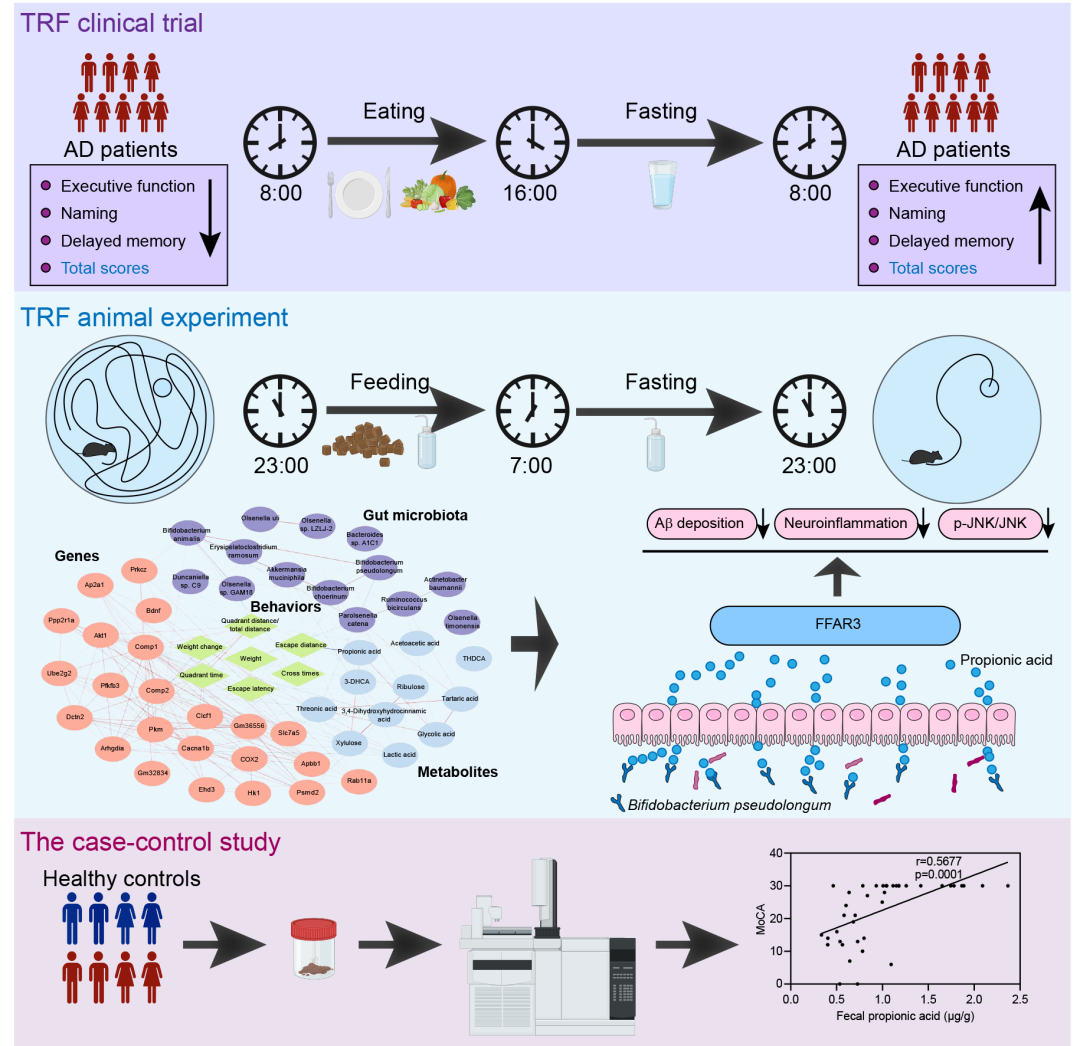
● 限制时间饮食(TRF)可改善阿尔兹海默病(AD)患者的认知功能,尤其是显著提高执行能力;
● AD小鼠的多组学整合分析发现,假长双歧杆菌(B. pseudolongum)和丙酸(PA)是TRF改善认知功能的关键靶点;
● TRF缓解AD引起的认知损伤的潜在分子机制为B. pseudolongum-PA-游离脂肪酸受体3(FFAR3)通路;
● 队列研究和TRF临床干预均表明PA是AD的潜在生物标志物。
摘 要
限制时间饮食(TRF)有望缓解衰老过程中认知能力的下降,尽管通过肠脑轴的确切机制仍不清楚。在一项临床试验中,我们首次观察到四个月的TRF改善了阿尔兹海默病(AD)患者的认知障碍。在5xFAD小鼠中证实了TRF对减轻认知功能障碍、淀粉样蛋白沉积和神经炎症的肠道微生物群依赖性作用。多组学整合分析将假长双歧杆菌(B. pseudolongum)和丙酸(PA)与 AD 发病机制中的关键基因联系起来。口服补充B. pseudolongum或PA可发挥与TRF相同的神经保护作用。正电子发射断层扫描成像证实PA可穿透血脑屏障,而游离脂肪酸受体3(FFAR3)的敲低削弱了TRF的认知益处。值得注意的是,我们观察到粪便中PA的含量与AD患者认知状态之间呈显著正相关,这进一步表明TRF促进了PA的产生。这些发现强调了微生物群-代谢物-脑轴是TRF发挥认知益处的关键,表明B. pseudolongum或PA是潜在的AD治疗的关键靶点。
视频解读
Bilibili:https://www.bilibili.com/video/BV1GEPxehE6X/
Youtube:https://youtu.be/JeVHiATyc4Q
中文翻译、PPT、中/英文视频解读等扩展资料下载
请访问期刊官网:http://www.imeta.science/
全文解读
引 言
阿尔兹海默病(AD)病情持续蔓延,影响全球超过4亿人。尽管AD的具体发病机制尚未完全阐明,但β淀粉样蛋白(Aβ)沉积和tau蛋白纤维缠结被公认为是AD的经典病理特征,并最终导致认知功能障碍、神经炎症和神经元损伤。越来越多的实验研究和临床数据揭示了肠道菌群-脑轴在AD进程中的关键作用。深入理解肠-脑轴的调控机制,对开发基于微生物的干预措施和治疗策略,以预防和控制AD及其他神经精神疾病具有重大潜力。AD的发生与肠道菌群的失调密切相关,其特征包括促炎菌群的增加和抗炎菌群的减少,同时伴随微生物代谢产物的显著变化,如短链脂肪酸(SCFAs)、支链氨基酸和胆汁酸。这些复杂的肠道菌群与脑功能的相互作用凸显了针对微生物生态系统的潜在治疗途径,为AD及其他神经退行性疾病的研究和管理提供了创新策略。
限制时间饮食(TRF)是一种安全有效的间歇性禁食策略,限制每日食物摄入时间至8-10小时,但不限制总能量及营养素的摄入。相较于传统的卡路里限制和隔日禁食,TRF更温和、更易于坚持,已在有限的临床试验中显示出对肥胖和糖尿病的治疗潜力。近期研究发现了TRF通过调节氧化应激、葡萄糖和脂质代谢、自噬和小胶质细胞分化等途径,在神经退行性疾病(如皮层下血管性痴呆和亨廷顿氏病)中发挥神经保护作用。最新研究还强调了恢复TRF中的昼夜节律失调有助于增强AD模型小鼠的认知功能。然而,尽管已有研究报道了TRF的多方面益处,但大多数证据主要来自小鼠模型,其作用机制,特别是肠道微生物群在其中的作用,仍未被完全揭示。我们之前的研究表明,禁食可通过调节肠道微生物群有效缓解糖尿病引起的认知障碍,而TRF对肠道微生物组及微生物代谢产物的深远影响主要在动物实验和有限的人体研究中得到证实。因此,尽管已有关于肠道微生物群-代谢物-脑轴的相关研究,但对特定益生菌及其代谢产物在认知功能改善中的作用仍缺乏深入理解。此外,目前的研究大多基于动物模型,限制了其在临床应用中的直接可行性,因此,迫切需要开展系统性研究,以建立改善肠道和脑健康的饮食策略的理论基础。
本研究通过系统性调查,深入探讨了TRF在改善AD相关认知障碍方面的治疗潜力。首次发现,4个月的TRF干预显著改善了AD患者的认知状态;3个月的TRF可有效延缓转基因AD模型小鼠(5xFAD小鼠)的病理进展,该模型能够快速重现AD的淀粉样蛋白病理特征。多组学整合分析及随后的抗生素干预实验进一步证明了肠道菌群-代谢物-脑轴在TRF对AD小鼠认知功能改善中的关键作用。此外,研究发现,TRF可显著促进益生菌假长双歧杆菌(B. pseudolongum)在AD小鼠肠道的定植,而该菌种能够通过产生丙酸(PA)特异性激活游离脂肪酸受体3(free fatty acid receptor 3, FFAR3),从而模拟TRF对AD相关认知障碍的改善作用。横断面人群研究及TRF临床干预试验均进一步证实,认知功能与粪便PA水平之间存在显著关联。本研究明确且有力地证明了肠道微生物群及其代谢产物在TRF与AD生理病理过程中的关键联系,并进一步揭示了B. pseudolongum或PA作为AD营养干预治疗的潜在应用价值。
结 果
TRF缓解AD患者以及AD小鼠的认知损伤
在本研究中,我们首先进行了一项为期4个月的小规模TRF临床干预试验,以评估其对AD患者(n = 9)认知功能的影响(图1A)。认知功能测试表明,TRF显著提高了AD患者的蒙特利尔认知评估(MoCA)分数,特别是在执行功能领域(p < 0.05),强调了TRF干预对AD患者认知障碍的缓解作用(图1B)。
为研究TRF对AD引起的认知障碍和病理损伤的保护作用的潜在生物学机制,我们使用6月龄的5xFAD小鼠进行实验,让其自由进食或每天进食8小时,持续3个月(图1C)。Morris水迷宫测试显示,与野生型(WT)小鼠相比,AD小鼠在5天的初始空间训练期间的逃避潜伏期时间显著延长,而TRF干预可逆转这一现象(图1D)。此外,TRF显著提高了AD小鼠在目标象限的停留时间(图1E)和在目标象限中路程比(图S1F),恢复至WT小鼠的正常水平,表明TRF可改善AD小鼠空间记忆能力。免疫荧光染色结果进一步证实,AD小鼠皮层和海马中Aβ和离子化钙结合衔接分子1(Iba-1)大量聚集,表明AD诱导了显著的淀粉样蛋白沉积和小胶质细胞活化(图1F和图S1H)。TRF干预显著减少了AD小鼠脑内Aβ斑块的阳性面积,并增加了小胶质细胞的聚集(图1G、H和图S1I、J),同时降低了促炎细胞因子肿瘤坏死因子-α(TNF-α)和白细胞介素-1β(IL-1β)的mRNA表达水平(图1I、J)。此外,TRF干预显著增加了AD小鼠皮层中脑源性神经营养因子(BDNF)的mRNA表达(图S1K),这一基因对神经元的存活、维持及再生至关重要。我们的研究结果共同表明,TRF增强了AD患者和AD小鼠的认知记忆功能,并通过促进Aβ斑块周围小胶质细胞的聚集,同时减少Aβ沉积和神经炎症,显著缓解AD的病理表现。
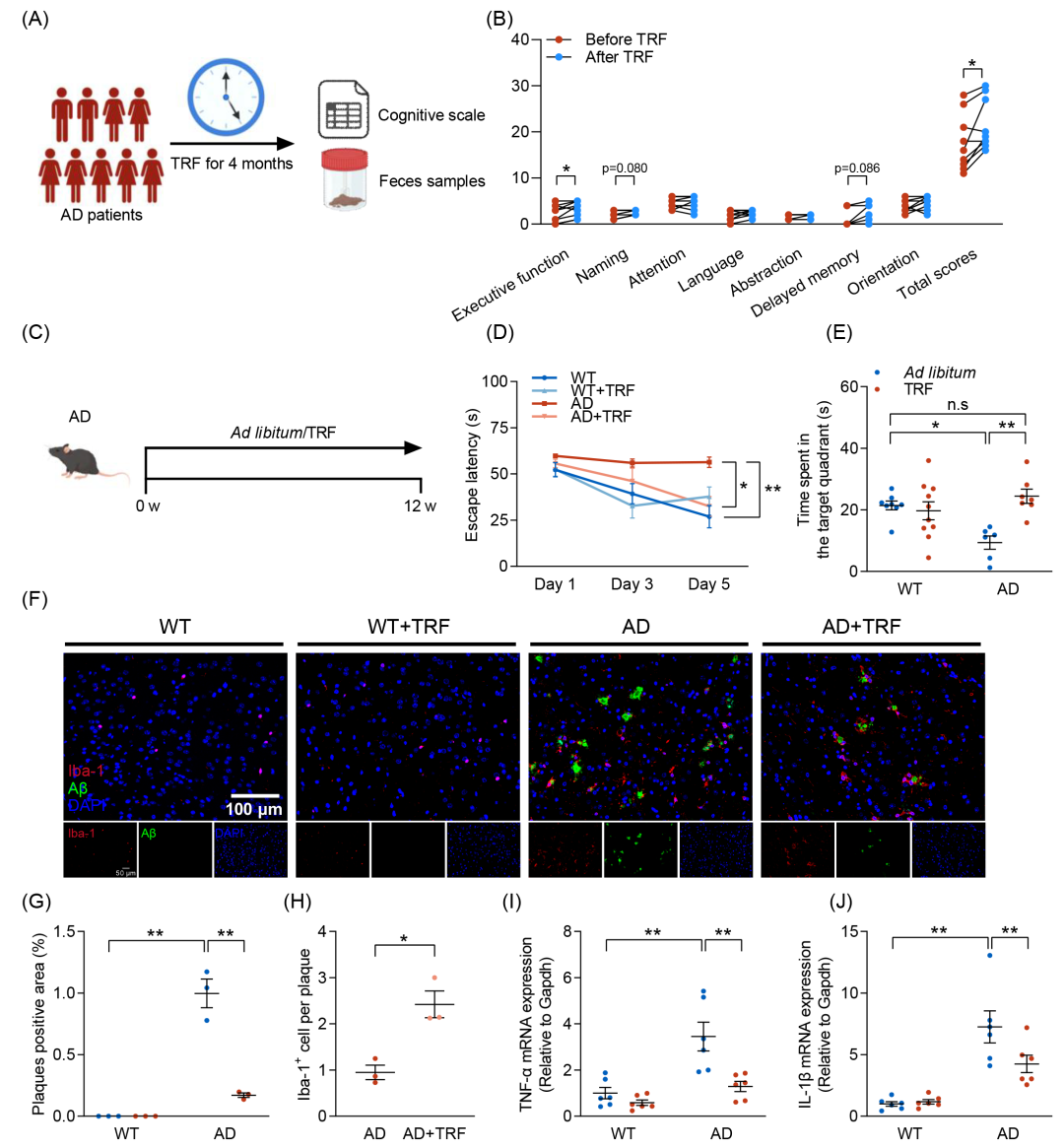
图1. 限制时间饮食(TRF)缓解AD相关的认知损伤
(A)AD患者接受为期4个月的TRF干预的示意图(n = 9);(B)TRF干预前后AD患者蒙特利尔认知评估(MoCA)总分及各子项目(执行功能、命名、注意力、语言、抽象功能、延迟记忆、定向能力)的量化分析;(C)每组接受TRF或自由进食(ad libitum)干预的实验流程示意图(n = 6-10);(D)逃避潜伏期;(E)目标象限的停留时间;(F)小鼠皮层中淀粉样β(Aβ)沉积(绿色)及离子化钙结合适配分子1(Iba-1+)阳性小胶质细胞(红色)的免疫荧光染色图像(n = 3)(比例尺:100 μm);(G)Aβ斑块阳性区域的定量分析;(H)Aβ斑块相关小胶质细胞的定量分析;(I)肿瘤坏死因子-α(TNF-α)的mRNA水平测定(n = 6);(J)白细胞介素-1β(IL-1β)的mRNA水平测定(n = 6)。数据以均值 ± SEM表示。*p < 0.05,**p < 0.01;双因素方差分析(two-way ANOVA)结合Tukey多重比较检验。
整合多组学分析揭示肠道微生物群-代谢物-脑轴在TRF依赖性改善AD小鼠认知功能障碍中的作用机制
尽管目前关于禁食对神经生物学健康影响的研究仍较为有限,但已有研究表明其具有积极作用,这与本研究结果一致。然而,TRF改善AD的具体机制仍未完全阐明。肠道菌群通过复杂的双向交流系统——肠-脑轴,与脑进行信息传递。该轴通过神经-免疫-内分泌介质,将外周肠道功能和微生物代谢与脑的情绪及认知中枢相互整合。鉴于肠道菌群及其代谢产物与AD发病机制之间日益密切的联系,深入理解TRF对AD的益处以及肠道微生物群-代谢物-脑轴在这一过程中的作用,对于AD及其他神经退行性疾病的预防和管理至关重要。
本研究综合运用海马转录组、粪便代谢组和肠道微生物组数据,从多层面挖掘证据,解析TRF依赖性改善AD认知障碍的微生物群-代谢物-脑轴机制。具体而言,我们对小鼠海马组织进行RNA测序,并将所有小鼠的183 Gb高质量RNA测序数据比对至小鼠(Mus musculus)基因组,共检测到10,272个基因,其中包括517个尚无注释的新预测基因。基因集富集分析(GSEA)揭示,在AD+TRF组中高表达的基因富集于多个生物学过程,包括IL-6信号通路、磷脂酸生物合成、线粒体调控及行为的正向调节(Benjamini-Hochberg校正后p < 0.01),而涉及神经系统发育、细胞过程的负调控及神经发生的基因则在TRF处理后呈现下调(p < 0.05)。进一步分析AD病理生理过程中重要的KEGG信号通路,如神经营养因子信号通路、mTOR信号通路、凋亡、AMPK信号通路及线粒体代谢(图2A),共筛选出381个基因用于偏最小二乘判别分析(PLSDA)。结果显示,AD组与AD+TRF组在PLSDA分析中可被明显区分(图2B),其中,电压门控钙通道亚基α1B(Cacna1b)、蛋白激酶C zeta(Prkcz)及环氧合酶-2(COX2)被鉴定为核心贡献因子,并在单变量分析中表现出显著的差异表达(图2C)。
随后,我们对粪便样本进行靶向代谢组学分析,以探究TRF处理对AD小鼠粪便代谢的影响,从而评估其对微生物活性的调节作用。共检测174种代谢物,包括氨基酸(n = 36)、苯甲酸类(n = 10)、胆汁酸(n = 32)、碳水化合物(n = 11)、肉碱类(n = 3)、脂肪酸(n = 32)、咪唑及吲哚类(n = 7)、有机酸(n = 24)、酚类及苯丙酸类(n = 10),以及短链脂肪酸(SCFAs,n = 9)(图2D及表S2)。结果显示,TRF显著增加了AD小鼠粪便中的胆汁酸(上升9.1%)及SCFAs(上升7.1%)总浓度,同时降低了脂肪酸(下降8.2%)及有机酸(下降6.7%)(图2D)。其中,TRF显著上调了咪唑丙酸(AD+TRF vs. AD,Fold change = 2.1, p = 0.04)及丙酸(PA)(AD+TRF vs. AD,Fold change = 1.5, p = 0.05)(图2E)。此外,TRF显著降低了牛磺去氧胆酸、3,4-二羟基氢桂皮酸、丙酰肉碱、异阿洛石胆酸、乳酸、木酮糖、核酮糖及硫酸化石胆酸的水平(图2E)。
鉴于肠道菌群在改善认知障碍中的关键作用,我们采用16S rRNA基因测序及全基因组鸟枪法宏基因组测序分析小鼠粪便样本,结果一致。TRF未能显著改变AD小鼠的肠道微生物群α-多样性、宏基因组功能通路丰度及功能变异性(图2F及图S3)。然而,TRF显著改变了肠道微生物组成,共检测到84种微生物分类单元在AD与AD+TRF组间呈现差异,其中36种在TRF干预后显著上调,48种显著下调(图S4A)。
进一步地,我们利用模糊C均值(Fuzzy C-means)聚类算法进行趋势聚类分析,以识别AD小鼠不同干预条件下的生物功能参数、AD病理相关差异表达基因、肠道菌群及粪便代谢物(变量数 = 6,372)的动态变化模式。最终确定16种不同的趋势聚类模式(图S4B)。其中,TRF在AD小鼠中上调的变量主要集中于Cluster 1、Cluster 4、Cluster 5、Cluster 14及Cluster 16,而下调的变量则聚集于Cluster 3、Cluster 8及Cluster 10。值得注意的是,大量与认知功能障碍及AD病理生理相关的TRF调控生物功能参数,与SCFAs(如PA)、特定胆汁酸及肠道菌群(Cluster 4及Cluster 16)呈现一致变化模式(图S4B)。其中,Akkermansia muciniphila与B. pseudolongum在AD小鼠中被TRF显著富集,且为丰度最高的菌种(图2G)。此外,TRF还增加了Bifidobacterium animalis及Bifidobacterium choerinum的相对丰度,尽管其水平低于B. pseudolongum(图2G,H)。进一步分析显示,TRF引起的生物功能参数变化、AD病理相关差异表达基因、肠道菌群及粪便代谢物之间存在密切的关联(图2I)。
综上,整合多组学分析表明,TRF通过调控肠道微生物群、微生物代谢物(如PA)及脑基因表达,构建了微生物群-代谢物-脑轴,从而介导TRF对AD小鼠认知功能障碍的改善作用。
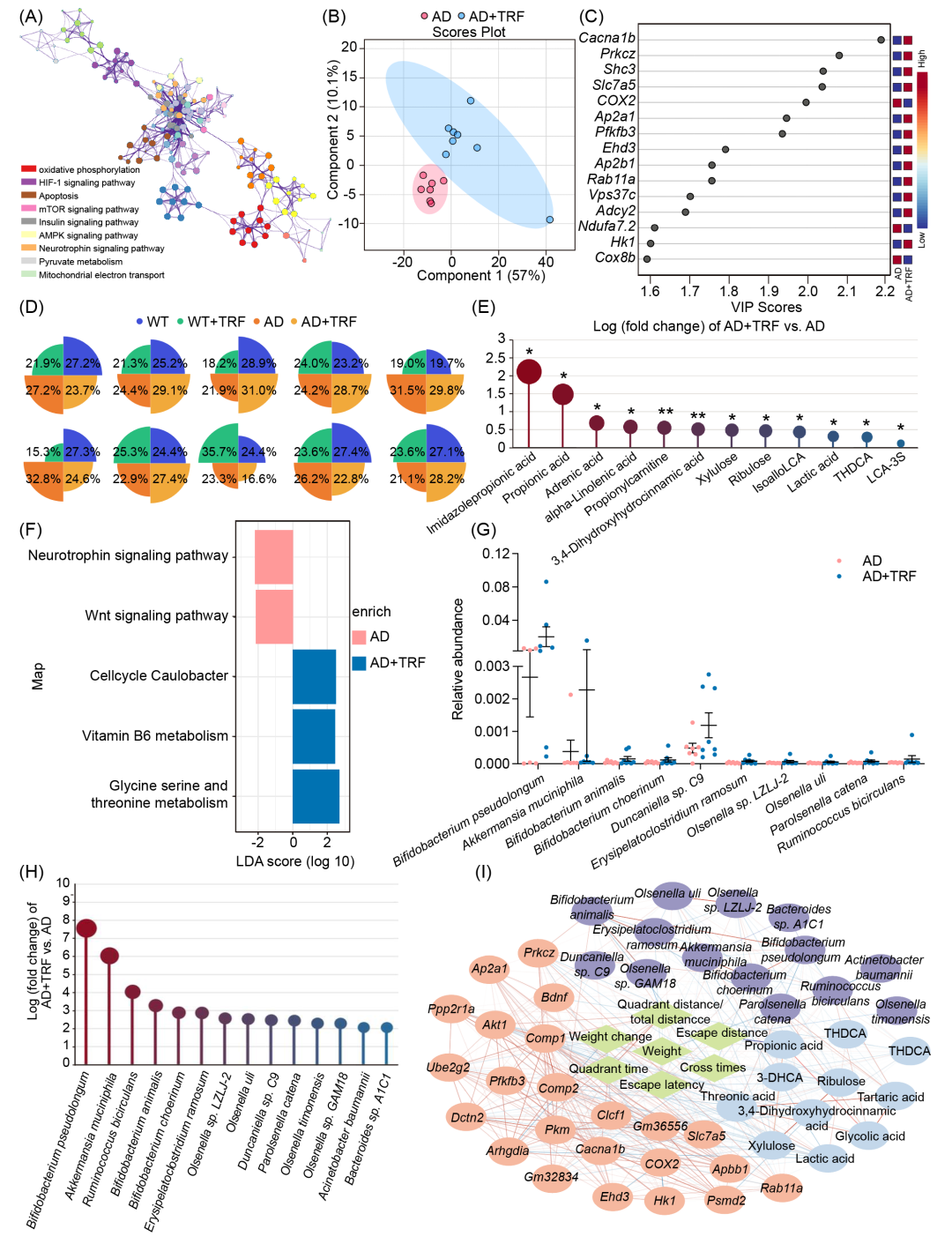
图2. 综合多组学分析揭示了肠道微生物-代谢物-脑轴在TRF改善AD小鼠认知功能障碍中的作用机制
(A)基因本体(GO)分析显示通过基因集富集分析(GSEA)鉴定的关键基因,这些基因涉及AD的病理生理过程(n = 7-10);(B)采用偏最小二乘判别分析(PLSDA)对GSEA筛选出的关键AD病理生理相关基因进行分析,实现了AD+TRF组与AD组的显著区分;(C)PLSDA分析识别出的关键基因贡献者,可用于区分AD+TRF组和AD组;(D)归纳总结四组小鼠中10类主要代谢物的相对百分比;(E)AD+TRF组相较于AD组代谢物变化最显著的代谢物倍数变化;(F)功能富集分析显示AD+TRF组与AD组之间存在差异的肠道微生物功能通路;(G)AD+TRF组与AD组间显著变化的关键菌的相对丰度;(H)AD+TRF组相较于AD组变化最显著的肠道菌群倍数变化;(I)TRF调控的基因、代谢物、肠道微生物、生化参数与认知参数之间的相关性分析。
肠道菌群在TRF改善AD小鼠认知功能障碍中的关键作用
为了进一步验证肠道菌群在TRF改善AD小鼠认知功能障碍中的介导作用,本研究采用广谱抗生素(ABX)对小鼠进行2周的肠道菌群清除处理,随后进行4周的TRF干预。实验全程每日给予ABX,以确保肠道菌群的持续消除(图3A)。结果显示,肠道菌群的耗竭消除了TRF对AD小鼠认知障碍的改善作用,这一现象可通过巴恩斯迷宫的逃避潜伏期、到达目标孔的潜伏期以及接近目标孔的次数等指标加以验证(图3B、C和图S5D)。此外,相较于AD+TRF组,仅接受抗生素处理的AD小鼠未表现出认知能力的显著改善(图3B)。免疫荧光染色结果显示,尽管ABX干预可减少Aβ沉积,其作用相较于TRF干预仍较为有限(图3D-F和图S5F,G)。肠道菌群的耗竭消除了TRF对AD病理损伤的改善作用(图3D-F和图S5F-H),并阻断了TRF诱导的小胶质细胞向Aβ斑块的募集(图3D-F和图S5F-H)。抗生素处理对脑内神经炎症因子的表达产生显著影响,表现为TNF-α水平的上升及IL-1β水平的下降(图3G,H)。然而,在TRF干预下,TNF-α和IL-1β均显著降低,而这一效应在去除肠道菌群后消失(图3G,H)。结果表明,肠道菌群在TRF对AD小鼠认知功能障碍的保护作用中发挥着关键调控作用。
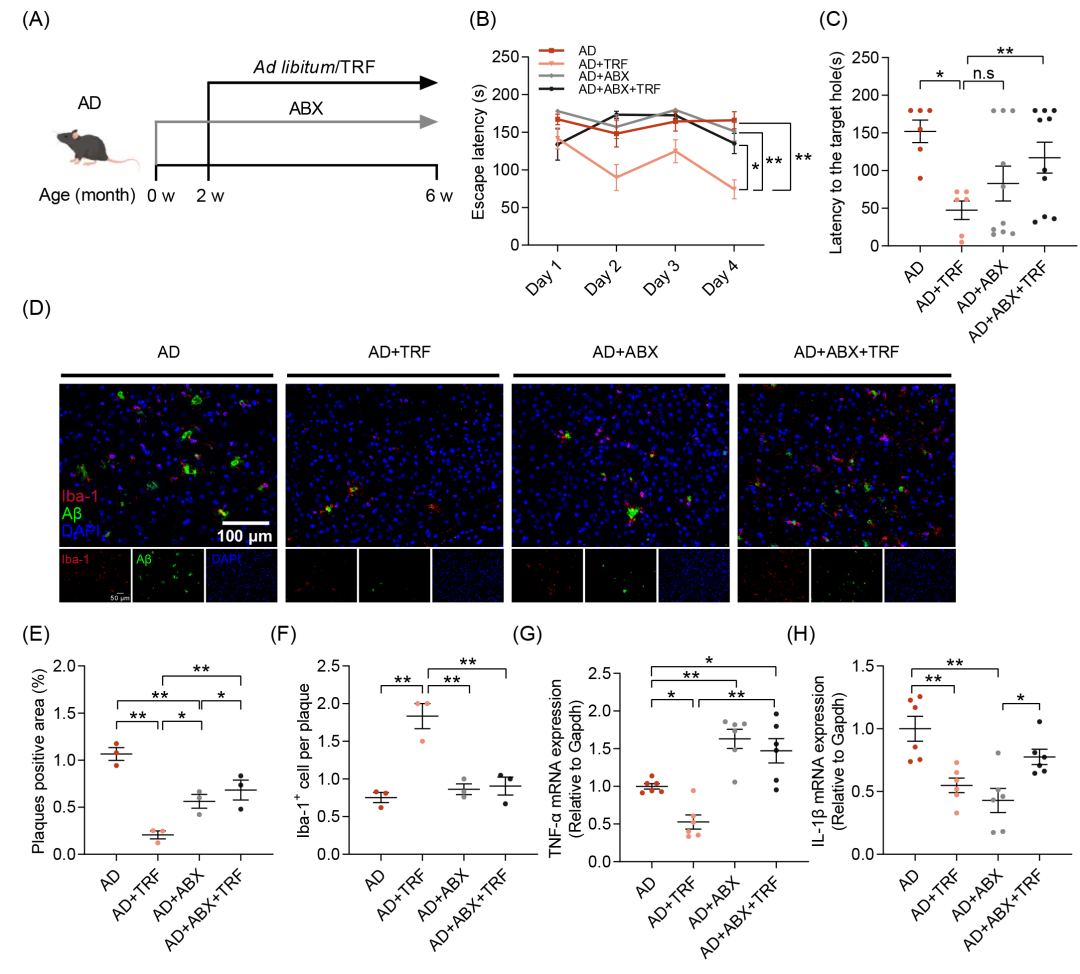
图3. 肠道菌群移除会消除TRF对AD小鼠的改善作用
(A)每组接受广谱抗生素(ABX)和TRF干预的实验流程示意图(n = 6-10);(B)逃避潜伏期;(C)到达目标孔的潜伏期;(D)小鼠皮层中Aβ沉积(绿色)及Iba-1+阳性小胶质细胞(红色)的免疫荧光染色图像(n = 3)(比例尺:100 μm);(E)Aβ斑块阳性区域的定量分析;(F)Aβ斑块相关小胶质细胞的定量分析;(G)TNF-α的mRNA水平测定(n = 6);(H)IL-1β的mRNA水平测定(n = 6)。数据以均值 ± SEM表示。*p < 0.05,**p < 0.01;单因素方差分析(one-way ANOVA)结合Tukey多重比较检验。
定植B. pseudolongum同时产生PA可改善AD小鼠的认知功能障碍
鉴于肠道菌群在TRF介导的AD认知功能障碍缓解过程中发挥着不可替代的作用,以及TRF诱导的B. pseudolongum丰度显著增加与AD病理生理学相关生物功能参数的改善密切相关,本研究进一步探讨了4周口服B. pseudolongum干预对AD小鼠认知障碍的影响(图4A)。正如预期的那样,B. pseudolongum干预显著改善AD小鼠的认知功能,表现为逃避潜伏期(图4B)、到达目标孔的潜伏期(图4C)和接近目标孔的次数(图S6D)的显著降低。此外,B. pseudolongum干预减少了AD小鼠脑内Aβ积累,并增加了Aβ斑块周围的Iba-1+小胶质细胞的聚集(图4D-F和图S6D-F)。此外,B. pseudolongum干预还显著降低了TNF-α水平,但对IL-1β的表达没有产生影响(图 4G、H)。与先前的研究一致,B. pseudolongum可促进SCFAs的产生。本研究观察到B. pseudolongum的相对丰度与粪便PA水平之间存在相关性,而TRF进一步增强这种相关性。值得注意的是,B. pseudolongum干预显著恢复了AD小鼠粪便中乙酸、PA和丁酸的水平,而这些代谢产物在AD小鼠中均明显下降(图4I)。这些结果表明,TRF诱导B. pseudolongum丰度增加,该菌与认知功能改善密切相关,并能够通过促进SCFAs生成,在缓解AD认知功能障碍及病理损伤方面发挥与TRF相似的作用。
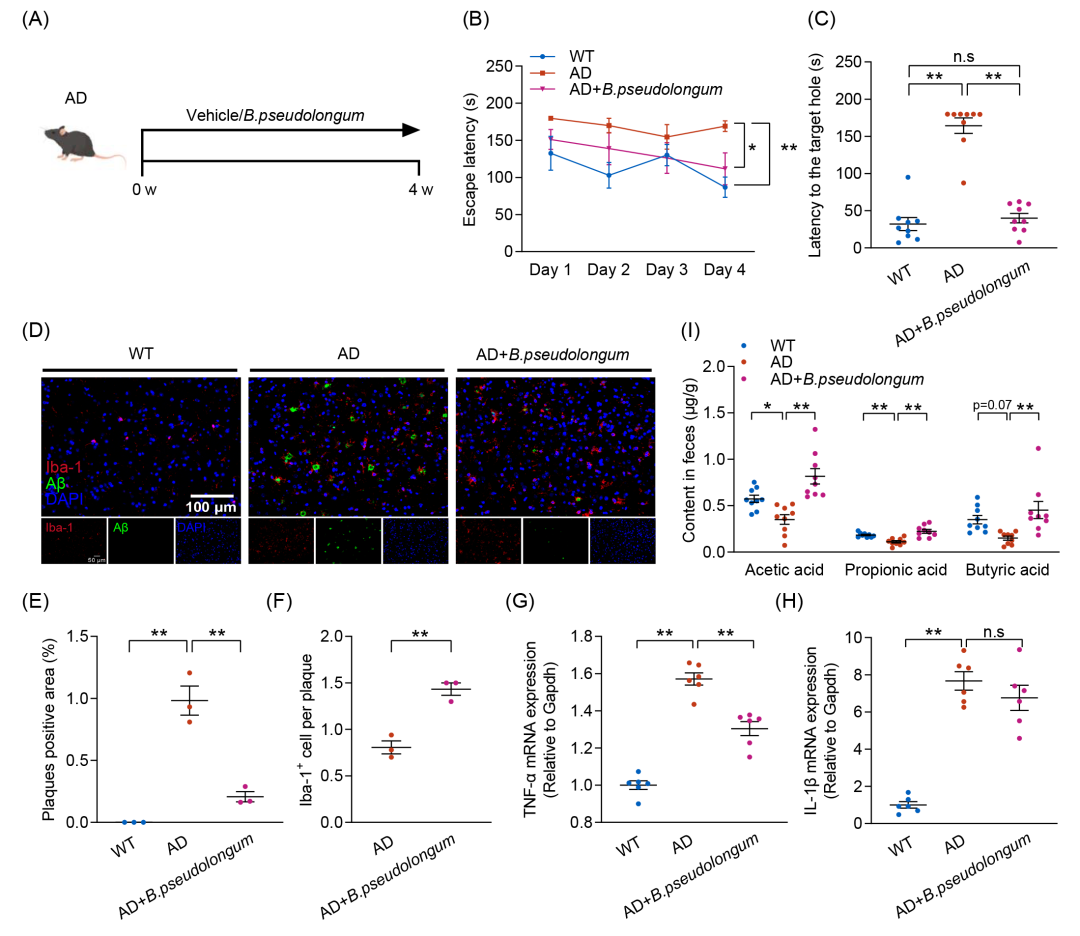
图4. B.pseudolongum干预缓解AD小鼠的认知损伤
(A)每组接受B.pseudolongum或生理盐水干预的实验流程示意图(n = 9);(B)逃避潜伏期;(C)到达目标孔的潜伏期;(D)小鼠皮层中Aβ沉积(绿色)及Iba-1+阳性小胶质细胞(红色)的免疫荧光染色图像(n = 3)(比例尺:100 μm);(E)Aβ斑块阳性区域的定量分析;(F)Aβ斑块相关小胶质细胞的定量分析;(G)TNF-α的mRNA水平测定(n = 6);(H)IL-1β的mRNA水平测定(n = 6);(I)粪便短链脂肪酸水平(n = 9)。数据以均值 ± SEM表示。*p < 0.05,**p < 0.01;one-way ANOVA结合Tukey多重比较检验。
PA 穿透血脑屏障并改善认知能力
为了进一步证实PA在AD发病机制中的重要性,并研究PA对认知缺陷的治疗作用,使用丙酸钠对AD小鼠进行干预(图5A)。腹腔注射PA显著增强了AD小鼠的认知功能(图5B、C和图S7C),显著减少了脑内Aβ聚集,促进了小胶质细胞向斑块的募集,并增加了BDNF表达(图5D-F 和图S7E-I)。此外,PA干预降低了TNF-α和IL-1β水平(图5G、H)。为评估PA在AD小鼠脑中的穿透性及代谢情况,本研究利用正电子发射断层扫描(PET)成像进行追踪分析。PET成像显示PA可通过血脑屏障(图5I),其代谢主要集中在下丘、中脑、纹状体和海马等关键脑区(图5J、K)。PET定量分析进一步揭示,AD小鼠的PA代谢信号显著降低,尤以海马、皮质及纹状体区域最为明显,然而PA治疗可有效恢复这些信号(图5K)。已有研究表明,SCFAs通过上调FFAR3和抑制核因子κB(NF-κB)和c-Jun N末端激酶(JNK)的磷酸化发挥抗神经炎症作用。为进一步探讨PA增强的脑内代谢信号对神经炎症及Aβ沉积的影响,测定了NF-κB和JNK的磷酸化水平。结果发现,PA干预显著抑制AD诱导的JNK磷酸化,但对NF-κB磷酸化无显著影响(图5L和图S7L),最终下调β-分泌酶(BACE1)的表达(图S7K)。因此,PA可穿透血脑屏障,恢复AD小鼠脑内PA代谢紊乱,并通过抑制JNK磷酸化和上调BDNF表达,改善认知损伤和减缓AD病理进展。
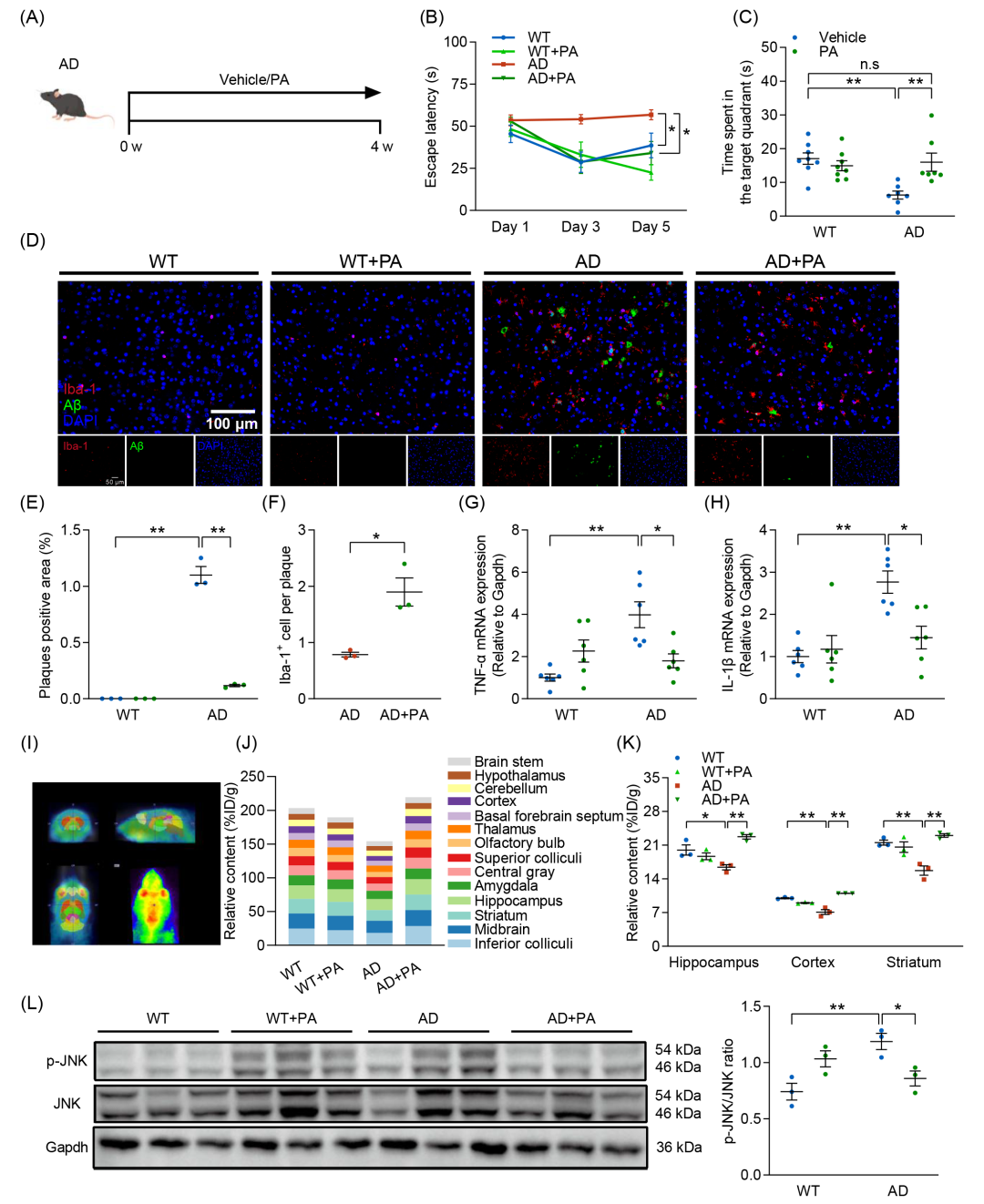
图5. 丙酸(PA)补充改善AD小鼠的认知功能损伤
(A)每组接受PA或生理盐水干预的实验流程示意图;(B)逃避潜伏期(n = 7-8);(C)目标象限的停留时间;(D)小鼠皮层中Aβ沉积(绿色)及Iba-1+阳性小胶质细胞(红色)的免疫荧光染色图像(n = 3)(比例尺:100 μm);(E)Aβ斑块阳性区域的定量分析;(F)Aβ斑块相关小胶质细胞的定量分析;(G)TNF-α的mRNA水平测定(n = 6);(H)IL-1β的mRNA水平测定(n = 6);(I)轴位、矢状位和冠状位正电子发射断层扫描(PET)图像,显示PA在脑中的摄取情况;(J)脑中[18F]-FPA的摄取水平(%ID/g)(n = 3)(K)海马、皮层和纹状体[18F]-FPA的PET定量分析。(L)Western blot分析p-JNK和JNK蛋白表达水平(n = 3)。数据以均值 ± SEM表示。*p < 0.05,**p < 0.01;two-way ANOVA结合Tukey多重比较检验。
FFAR3的敲低消除TRF的神经保护作用
已有研究表明,FFAR3可与PA结合并调节心肌损伤。为探讨FFAR3在TRF干预AD过程中的作用,本研究采用小发夹RNA(shRNA)特异性敲低FFAR3的mRNA水平和蛋白质表达,并通过双侧侧脑室注射shRNA-FFAR3进行干预,同时以shRNA-con作为病毒载体对照(图6A及图S8A,B)。认知行为测试结果表明,与AD+TRF组相比,FFAR3的敲低消除了TRF对AD小鼠认知功能的改善作用(图6B,C和图S8E)。此外,与FFAR3的敲低消除TRF对认知功能障碍的保护作用一致,FFAR3的剥夺显著逆转了TRF干预后所导致的Aβ过度聚集的减少,并减少了小胶质细胞向Aβ斑块的募集(图6D-F和图S8G-I)。同样地,敲低FFAR3也消除了TRF对神经炎症因子表达的抑制作用(图6G,H)。结果表明,FFAR3在TRF清除Aβ、缓解神经炎症以及改善认知功能的过程中发挥着不可或缺的作用。
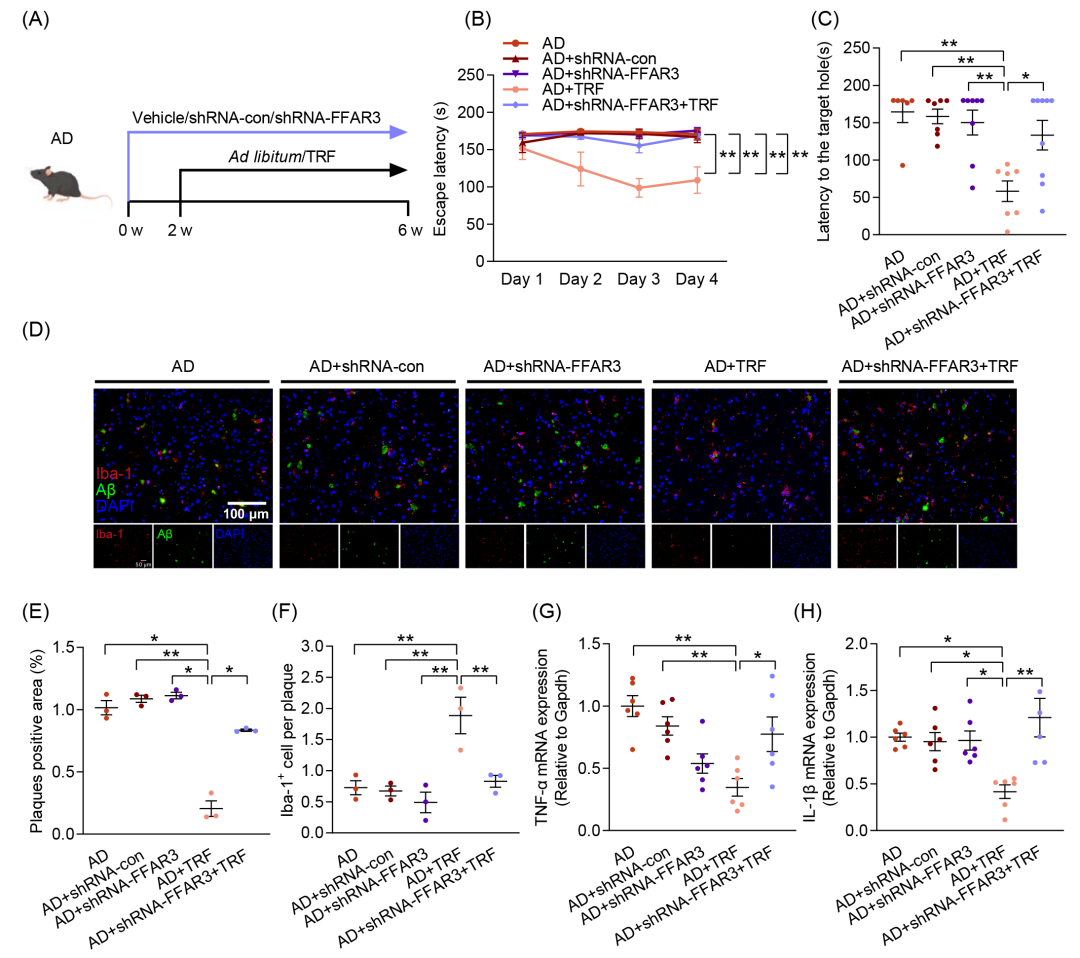
图6. 游离脂肪酸受体3(FFAR3)敲低消除TRF对AD小鼠的益处
(A)每组接受shRNA-FFAR3、shRNA-con和TRF干预的实验流程示意图(n = 6-9);(B)逃避潜伏期;(C)到达目标孔的潜伏期;(D)小鼠皮层中Aβ沉积(绿色)及Iba-1+阳性小胶质细胞(红色)的免疫荧光染色图像(n = 3)(比例尺:100 μm);(E)Aβ斑块阳性区域的定量分析;(F)Aβ斑块相关小胶质细胞的定量分析;(G)TNF-α的mRNA水平测定(n = 6);(H)IL-1β的mRNA水平测定(n = 6)。数据以均值 ± SEM表示。*p < 0.05,**p < 0.01;one-way ANOVA结合Tukey多重比较检验。
PA作为AD生物标志物在缓解认知障碍中的关键作用
为了研究SCFAs是否可作为AD的潜在生物标志物,本研究开展了一项病例对照研究,分析了AD患者(n = 21)和健康受试者(n = 20)粪便中PA和丁酸的浓度(表 S4)。认知功能评估结果表明,与健康个体相比,AD患者的认知能力显著下降,其蒙特利尔认知评估量表(MoCA)及简易精神状态检查(MMSE)评分均明显降低(表S4)。结果显示AD患者粪便中PA水平显著低于健康受试者(图7A)。相关性分析进一步揭示,粪便PA浓度与认知功能呈显著正相关(图7B 和图S9A、B)。此外,粪便丁酸水平同样与认知功能呈显著正相关,尽管其相关系数低于PA(图 S9C、D)。
为进一步明确TRF对PA水平的影响以及其在缓解AD认知障碍中的作用,本研究检测图1中提到的4个月TRF临床干预前后的AD患者粪便PA水平(图S9E)。结果发现,粪便PA水平的提升与MoCA总分及多个特定认知领域(包括执行功能、命名、注意力和语言)呈显著正相关(图7C,D)。通过病例对照研究和小规模的TRF临床干预试验,本研究证实了粪便PA水平与认知状态呈显著正相关,提示PA可以作为AD的潜在生物标志物。
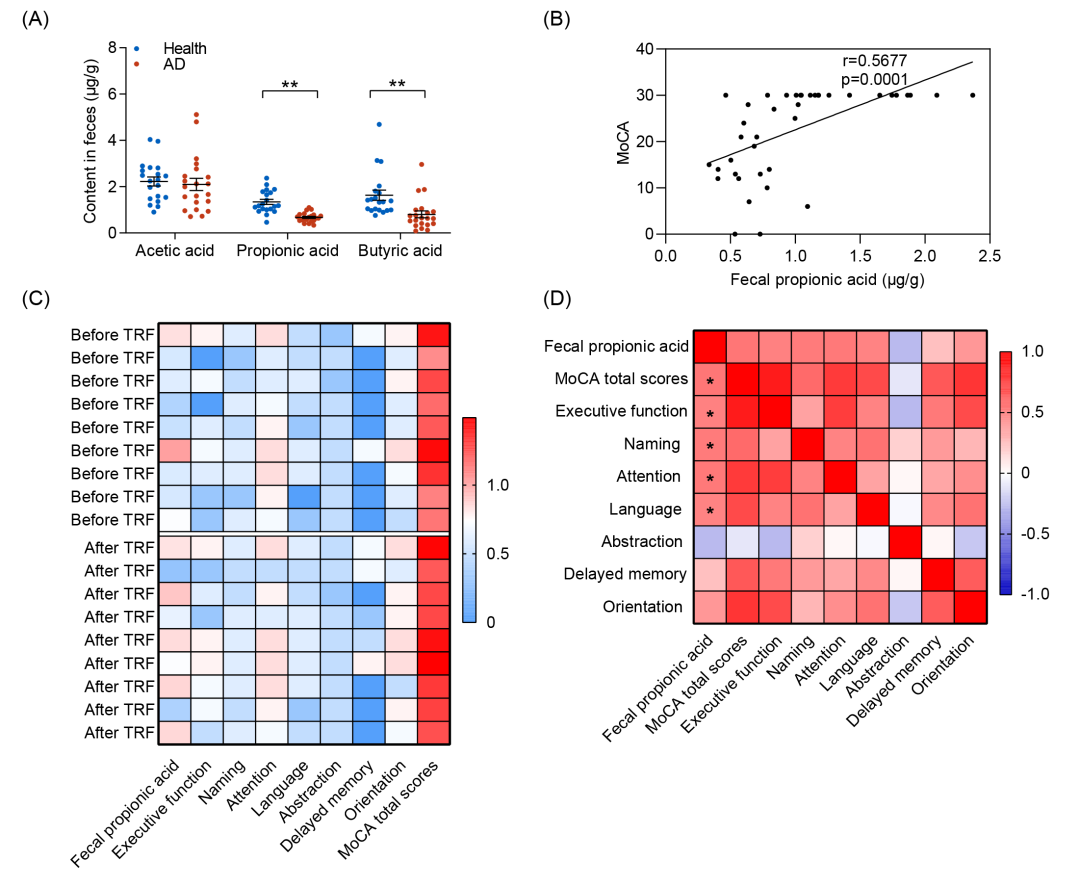
图7. PA作为AD患者的生物标志物在缓解认知障碍中发挥关键作用
(A)健康受试者与AD患者粪便中SCFAs水平的比较(n = 20-21);(B)散点图显示MoCA评分与粪便PA水平之间存在显著正相关(Pearson相关系数 r = 0.5677, p = 0.0001);(C)热图显示接受TRF治疗前后的AD患者粪便PA浓度和MoCA评分;(D)热图展示粪便PA水平变化与MoCA评分之间的Spearman秩相关分析结果。数据采用双尾Student’s t检验分析,*p < 0.05,**p < 0.01。
讨 论
越来越多的证据证明了TRF具有神经保护作用,主要是通过动物研究。然而,TRF对AD相关认知障碍的神经保护作用的机制仍然不清楚。本研究报告发现,4个月的TRF干预极大地改善了AD患者的认知功能,并提供了令人信服的证据,表明TRF干预对改善AD引起的认知缺陷、过量Aβ积累和神经炎症的肠道菌群和SCFAs依赖性作用。TRF特别提高了B. pseudolongum和PA的粪便丰度。补充B. pseudolongum或其微生物代谢物PA可有效复制TRF在AD小鼠中的神经保护作用。
从机制上讲,PA表现出穿过血脑屏障和调节脑病理相关的PA代谢的能力,从而通过激活FFAR3增强AD小鼠的认知功能,FFAR3调节下游信号通路,包括JNK磷酸化和BDNF表达。值得注意的是,粪便中PA浓度与AD患者的认知功能障碍之间存在显著的负相关,进一步强调了PA是TRF减轻认知障碍的关键因素。我们的研究结果明确证明了微生物群(B. pseudolongum)-代谢物(PA)-FFAR3轴是TRF认知益处的基础,并强调了非药物干预策略(包括TRF和其他肠道微生物衍生的营养干预)在阻止AD进展方面的转化潜力。
先前的研究表明,维持TRF干预6个月可改善老年人的认知状态。虽然统计能力受到样本量的限制,但我们的结果证明4个月的TRF增强了AD患者的认知能力(图1)。最近的研究表明,TRF通过上调BDNF水平、降低炎症因子(IL-6和TNF-α)的表达和促进神经发生发挥神经保护作用。研究表明,间歇性禁食和卡路里限制对AD样病理有抑制作用,表现为降低脂蛋白脂肪酶表达、降低Aβ40和Aβ42水平、降低磷酸化tau水平。最新研究从昼夜节律的角度解释了TRF对AD认知和Aβ沉积的改善作用,重点强调TRF干预通过恢复脑转录过程的紊乱,从而改善AD引起的睡眠障碍。尽管TRF对认知障碍和神经损伤的作用机制和影响尚有争议,但本研究证实TRF增强了AD小鼠的空间记忆能力、减少了Aβ的沉积、促进了小胶质细胞向斑块的聚集、提高了BDNF水平(图1)。海马转录组的PLSDA有效地区分了AD+TRF组和AD组之间的基因贡献者,包括Cacna1b、Prkcz和COX2。TRF显著增加Cacna1b和Prkcz表达,同时降低COX2表达(图S2)。据报道,Cacna1b参与维持突触信号传导,其失调与AD的发病机制有关。由Prkcz编码的蛋白激酶C的激活可调节BACE1的活性并控制Aβ前体APP的裂解。此外,COX2被发现介导IL-1β和Aβ之间的相互调节,促进AD的发病机制。这些发现表明TRF对神经退行性疾病的发展具有有益作用。具体而言,TRF诱导的差异表达基因可作为治疗AD的潜在靶点。
肠道菌群-脑轴因其在代谢和神经退行性疾病中的重要作用而得到了全世界的认可。值得注意的是,将AD患者和AD模型小鼠的粪便定植在无菌小鼠体内会降低成年海马神经发生率、加剧记忆障碍并加剧AD的发展。针对肠道菌群的干预措施,如抗生素疗法、粪便菌群移植和益生菌给药,已证明具有缓解AD进展的潜力。研究表明,使用抗生素消除肠道菌群可减少AD小鼠脑中的Aβ沉积,同时增加可溶性Aβ水平。我们的研究结果表明,去除肠道菌群可减轻Aβ沉积。然而,ABX干预消除了TRF对AD引起的认知功能障碍的有益作用(图3),这可以通过ABX干预消除与AD相关的有害肠道微生物群和TRF诱导的有益肠道微生物群来解释。
间歇性禁食对肠道微生物群的影响已被广泛研究和记录。我们之前的研究表明,隔日禁食会增加乳酸杆菌的丰度,从而提高认知能力、抑制神经胶质细胞过度激活和增强线粒体的生物合成。隔日禁食通过提高肠道细菌丰富度和促进乳酸杆菌科、拟杆菌科和普氏菌科的丰度,在多发性硬化症模型小鼠中发挥神经保护作用,从而影响与抗氧化反应相关的微生物代谢途径。在AD中,肠道微生物群失调的特点是促炎门增加和抗炎门减少的不平衡。既往研究报道健康个体与AD患者粪便菌群组成和多样性存在显著差异,主要集中于瘤胃球菌科、梭菌科、阿克曼菌和酪酸球菌的交替变化。但目前对于AD患者粪便中双歧杆菌丰度的变化存在争议,一些研究认为双歧杆菌的增多与AD的病理过程有关,而另一些研究则强调双歧杆菌的减少可能是由于健康个体与AD患者肠道菌群组成的差异所致,这些差异可能受到AD患者的入组条件和饮食环境的影响。但一些临床干预试验已经证明双歧杆菌BGN4和长双歧杆菌BORI对患有认知功能障碍和抑郁症的老年个体具有认知益处。与我们的发现一致,TRF极大地抑制了AD小鼠体内双歧杆菌(尤其是B. pseudolongum)的衰退,而B. pseudolongum干预可改善认知功能并减轻AD的病理症状(图4)。这些争议凸显了进一步研究双歧杆菌及其衍生代谢物对AD模型和患者的直接影响的必要性。
我们的研究还强调了B. pseudolongum与粪便SCFAs之间的相关性,这种相关性在AD小鼠中被TRF显著增强(图2)。已证明补充SCFAs可促进神经发育、增强认知功能和改善行为障碍。SCFAs和FFAR3的组合可抑制ERK/JNK/NF-κB信号传导,促进认知缺陷和AD型病理的恢复。我们的研究进一步表明,PA干预可有效抑制AD诱导的JNK磷酸化、降低BACE1表达、减轻Aβ聚集并减弱神经炎症反应(图5)。这强调了PA在预防神经退行性疾病发展方面的直接影响。具体而言,调节JNK通路可降低Aβ沉积以及Aβ40和Aβ42的水平,已知Aβ40和Aβ42可介导与AD相关的病理性细胞死亡。尽管PA对认知功能有益作用的潜在机制仍不清楚,但最近的一项临床研究表明,多发性硬化症患者脑脊液中PA浓度的增加可能会增强Treg细胞线粒体的功能和形态。此外,自闭症谱系障碍患者的淋巴母细胞已表现出使用PA作为能量产生燃料的能力增强,尽管对线粒体功能的浓度依赖性影响可能导致过量活性氧产生和能量产生受损。根据我们的研究结果,AD小鼠脑中的PA代谢受损,表明PA不足以消耗能量。PA补充有效恢复了脑对PA的利用率(图5)。总的来说,TRF的神经保护作用可能部分归因于恢复涉及PA的能量代谢途径和抑制JNK过度磷酸化,这可能会影响下游信号通路。然而,有必要进一步研究PA对脑线粒体质量控制系统和呼吸酶的影响。
PA通过被动扩散或通过载体蛋白(例如SCFAs受体FFAR2和FFAR3)进入细胞。研究表明,FFAR3被认为是肠神经系统中PA的唯一受体,并在人脑内皮细胞中表达,而FFAR2尚未在中枢神经系统或周围神经系统中检测到。据报道,PA与FFAR3相互作用,通过降低氧化应激来防止心肌缺血-再灌注损伤。然而,PA与脑中的FFAR3结合对认知功能的影响仍未得到充分探索。在本研究中,我们使用FFAR3-AAV验证了TRF通过PA-FFAR3通路改善了AD诱导的认知障碍(图6)。与我们的研究一致,在炎症性神经病大鼠模型中,PA被发现通过与FFAR3结合在周围神经系统中发挥神经再生作用。此外,我们观察到PA干预显著抑制了FFAR3下游信号分子JNK的磷酸化(图5),表明PA通过与FFAR3结合抑制了JNK磷酸化。TRF干预降低了NF-κB调控的下游神经炎症因子(TNF-α和IL-1β)的水平,敲低FFAR3后这种作用消失(图6)。FFAR3在不同神经胶质细胞中的表达和功能尚不清楚,需要更多的研究来探索FFAR3对不同神经胶质细胞神经炎症反应的抑制作用。
最近一项针对轻度认知障碍和帕金森病患者的队列研究发现,粪便SCFAs显著减少,并且粪便SCFAs的减少与认知相关脑区Aβ沉积呈负相关,这表明SCFAs可以作为区分健康个体和神经退行性疾病患者的早期诊断生物标志物。最近的研究支持了类似的发现,表明轻度认知障碍患者的粪便样本中PA和丁酸的浓度与Aβ42表达呈负相关。我们还观察到,接受TRF干预的AD患者其粪便中PA水平较对照组更低,并且PA水平与认知状态呈负相关(图7)。这一结果支持PA可能作为TRF缓解认知功能障碍的关键调节因子,并且其水平与MoCA评分呈显著相关性(图7)。
我们研究的优势在于对小鼠模型进行了广泛的生理分析和多组学研究,并得到了人体试验的证实,为TRF改善认知的机制提供了新的见解,并确定了关键的肠道贡献者,即假长双歧杆菌和PA。然而,我们的研究有几个局限性。PA对神经系统的影响是有争议的。过量或急性PA干预,例如剂量超过200 mg/kg,与自闭症和丙酸血症有关。研究结果的不一致可能归因于不同干预措施中使用的剂量和给药方法的差异,这至关重要地强调了剂量和给药方法在确定PA疗效方面的重要性。在此,小鼠腹膜内注射0.1 g/kg体重的丙酸钠,导致AD小鼠的认知能力改善,并延缓了小鼠AD病理的进展(图5)。此外,据报道,TRF可通过多种机制改善认知能力。例如,长时间禁食会诱导酮体的产生。酮体补充已证明对轻度认知障碍患者的执行功能、记忆力和语言能力有益。此外,除了PA,其他SCFAs(如乙酸和丁酸)也显示出认知益处。乙酸已被证明可以减轻1型糖尿病小鼠的认知障碍,丁酸通过与其受体FFAR2和FFAR3结合改善AD小鼠模型中的认知缺陷。然而,在我们的研究中,乙酸和丁酸在TRF后均未发生显著变化,并且未观察到乙酸与AD患者的认知参数之间的显着关联。最后,我们进行的小规模TRF人群临床干预试验样本量有限,并且未考虑性别和遗传等因素。尽管如此,它提供了有价值的见解,并为未来设计更大规模的TRF临床干预研究提供了有用的参考。此外,收集不同神经退行性疾病患者的样本以确定粪便PA水平降低是AD所特有的还是神经退行性疾病的共同特征,也十分有意义。脑脊液的生化分析也是必要的。未来有必要对PA、丁酸及其相关肠道微生物群对AD患者的影响进行大样本研究。
结 论
总而言之,我们的研究结果支持TRF通过调节B. pseudolongum-PA-FFAR3 轴改善AD小鼠的认知缺陷。为期4个月的TRF临床干预(每日禁食16小时)可缓解AD患者的认知障碍。TRF对AD认知衰退的有益作用可能源于B. pseudolongum的相对丰度增加及PA的增强,从而在肠道菌群依赖的机制下抑制神经炎症和Aβ沉积。SCFAs受体FFAR3的敲低消除了TRF对认知功能障碍的改善作用。此外,在病例-对照研究中,我们发现PA与认知状态之间存在显著相关性。总体而言,我们的研究为靶向肠道微生物及其代谢产物(特别是双歧杆菌和PA)的非药物干预策略在神经退行性疾病中的应用提供了新的研究方向。
方 法
动物实验
携带5xFAD突变的雄性转基因小鼠(库存号:006554),以C57BL/6J为遗传背景,从Jackson Laboratory(Bar Harbor,ME,USA)购入。5xFAD小鼠在人淀粉样前体蛋白(APP)和人早老素1(PSEN1)基因上携带五个与阿尔兹海默病(AD)相关的突变,其中包括APP基因中的瑞典突变(K670N/M671L)、佛罗里达突变(I716V)和伦敦突变(V717I),以及PSEN1基因中的M146L和L286V突变。5xFAD小鼠的构建方法已在先前研究中描述。实验中,小鼠定期与Jackson实验室提供的雌性C57BL/6J小鼠回交。为了鉴定后代基因型,使用聚合酶链式反应(PCR)对小鼠的APP和PS1转基因进行检测,基因组DNA样本来自耳部组织(DNA提取试剂盒,北京倍特克生物科技有限公司,北京,中国)。
本研究共进行五项动物实验。实验严格遵循西北农林科技大学实验动物管理条例,并获得伦理批准(批准号:XN2023-0610)。所有实验均严格符合动物研究的伦理标准和相关法规。
实验小鼠均喂养标准饲料,并单独饲养于恒温(22 ± 2°C)、相对湿度(55 ± 5%)的环境中,采用12/12 h的昼夜光照循环,光照时间:上午8:00(ZT0)开始。动物实验包括行为学测试和病理学分析。实验结束后,海马组织被解剖并存储于-80°C以备进一步分析。具体实验方案如下:
(1)限制时间饮食(TRF)对阿尔兹海默病(AD)小鼠认知障碍的影响
进行12周的TRF干预,以研究其对AD小鼠认知障碍的影响。6月龄的雄性5xFAD(AD)小鼠及其年龄匹配的野生型(WT)小鼠(相同遗传背景)被随机分为自由进食(ad libitum)组和TRF组(n = 7-10)。TRF干预的小鼠每天在晚11点至次日早7点(ZT15-ZT23),每日进食8小时,水摄入不限。干预结束后,收集粪便样本用于代谢组学和微生物组分析。
(2)肠道微生物群在TRF缓解AD认知障碍中的介导作用
为了验证肠道微生物群在TRF缓解AD认知障碍中的不可替代作用,研究比较了在有无肠道微生物的情况下,AD小鼠在4周TRF干预后的行为和病理学表现。TRF干预方案同前述。为了耗竭肠道微生物群,对7月龄雄性5xFAD小鼠给予口服抗生素混合物(ABX),包括新霉素(1 g/L)、氨苄西林(1 g/L)、甲硝唑(1 g/L)和万古霉素(0.5 g/L)(Sigma-Aldrich,上海,中国),如先前研究所述。ABX从TRF干预前两周开始添加于饮用水中,并持续整个实验以维持肠道菌群的去除。AD小鼠被随机分为AD、AD+TRF、AD+ABX、AD+ABX+TRF四组(n = 6-10)。采用厌氧和需氧培养技术以及定量qRT-PCR检测粪便样本,以验证肠道菌群的去除情况。qRT-PCR引物为:8F: AGAGTTTGATCCTGGCTCAG;338R: CTGCTGCCTCCCGTAGGAGT。
(3)假长双歧杆菌(B. pseudolongum)对AD小鼠认知障碍的影响
为了研究B. pseudolongum干预是否可缓解AD小鼠的认知障碍,实验首先在37°C厌氧培养B. pseudolongum(ATCC 25526; zlzt, 武汉,中国)24小时,然后5000 rpm、5 min、4°C离心收集菌体,并重悬于PBS,浓度为1×10⁹ CFU/mL。7月龄雄性AD小鼠被随机分为3组(n = 9):每日灌胃100 μL B. pseudolongum悬液或PBS对照,持续4周。实验结束后,提取粪便DNA并进行qRT-PCR以检测B. pseudolongum的定植情况(16S引物:Forward: 5'-CGCCGATGATGGGATGCTTTACA-3';Reverse: 5'-AGATCATCTAAACACCACCCACAC-3')。同时,收集粪便样本以检测短链脂肪酸(SCFAs)。
(4)丙酸(PA)处理对AD小鼠认知障碍的影响
7月龄雄性AD小鼠被随机分为4组(n = 7-8),每日腹腔注射0.1 g/kg体重的丙酸钠(Sigma-Aldrich,上海,中国),溶于生理盐水,或注射等量生理盐水作为对照,持续4周。丙酸钠的剂量参考了先前文献。
(5)游离脂肪酸受体3(FFAR3)在TRF改善AD小鼠认知障碍中的作用
为了研究短链脂肪酸受体FFAR3在TRF对AD认知功能影响中的作用,我们进一步在FFAR3基因敲低的AD小鼠中探讨TRF的神经保护作用。构建AAV2/9载体(携带CMV启动子和GFP报告基因)以敲低FFAR3(FFAR3-shRNA序列:5'-TTTGCTAAACCTGACCATTTC-3')(合源生物,上海,中国)。AD小鼠被随机分为5组(7月龄,n = 6-9):AD组(PBS注射);AD+shRNA-con组(AAV2/9-GFP注射);AD+shRNA-FFAR3组(AAV2/9-GFP-FFAR3注射);AD+TRF组(PBS注射);AD+shRNA-FFAR3+TRF组(AAV2/9-GFP-FFAR3注射)。实验采用2%异氟烷麻醉小鼠,使用68025(RWD,深圳,中国)立体定位仪向双侧侧脑室(坐标:AP -0.3 mm, ML ±1.0 mm, DV -2.5 mm)随机注射1×10¹² AAV(1 μL,200 nL/min),采用LEGATO 130(KD Scientific Inc)微量注射泵。注射后针头保持10 min防止回流。小鼠恢复1周后,进行4周TRF干预。
病例对照研究
为了评估粪便SCFAs水平与AD症状之间的关联,我们于2023年10月至2024年6月期间,在北京大学深圳医院(中国深圳)招募了30名健康受试者和30名AD患者。依据《DSM-IV》标准筛选出因AD导致痴呆的患者,排除因手术或其他原因引起的痴呆个体。本研究的受试者排除标准包括:(1)具有炎症性肠病、碳水化合物吸收不良、激素失衡、已知食物添加剂过敏或其他严重疾病史者;(2)既往接受胆囊切除术、胃肠道或颅脑手术者;(3)存在寄生虫感染者;(4)有自杀倾向、尝试或攻击行为者;(5)使用已知影响胃肠功能、血压、血脂的药物,或服用激素补充剂、抗过敏/哮喘药物、质子泵抑制剂者;(6)在测试前8周内服用益生菌、抗生素或益生元(膳食纤维、低聚糖)者;(7)存在酒精或药物滥用者;(8)认知功能障碍或有严重创伤史者;(9)具有脑血管疾病史或因脑肿瘤等原发性脑病变导致认知功能障碍者;(10)存在严重烟草依赖者。
所有受试者接受以下评估:(1)通过蒙特利尔认知评估量表(MoCA)和简易精神状态检查量表(MMSE)评估综合认知功能;(2)进行医学访谈以确认病史和当前用药情况;(3)采集脑脊液样本分析Aβ40、Aβ42、总Tau(total-Tau)及磷酸化Tau(p-Tau)水平(自愿参与),为AD的诊断提供依据;(4)进行正电子发射计算机断层扫描(PET-CT),包括Aβ-PET和Tau-PET(自愿参与),以辅助AD诊断;(5)采集血样分析AD相关生物标志物(包括Aβ40、Aβ42、total-Tau及p-Tau等,自愿参与),进一步辅助AD的临床诊断。
根据筛选标准,并结合NINCDS-ADRDA的诊断标准,最终纳入20名健康受试者(MoCA ≥ 27且MMSE ≥ 27)和21名AD患者(MoCA < 27且MMSE < 27,同时在脑脊液分析、PET-CT扫描或血液生物标志物检测中存在异常)进入研究队列(表S4)。所有受试者均获得详细的研究方案说明,并签署书面知情同意书。本研究经北京大学深圳医院伦理委员会批准,伦理审批编号为No. 2023-108。
TRF临床试验
本研究于2023年10月至2024年6月期间招募受试者,并于2024年3月完成数据收集。AD患者的纳入标准与上述病例对照研究一致。最终,共有10名AD患者(2名男性,8名女性,年龄范围58至81岁)经脑脊液分析、PET-CT扫描或血液生物标志物检测确诊,并纳入TRF干预研究。在TRF干预前一周,所有受试者接受了饮食和生活方式指导,该指导内容由专家讲座和印刷宣传材料提供,并基于《中国居民膳食指南(2016)》及《中国全民健康生活方式行动(2017-2025)》制定。
受试者被要求每天8小时内自由进食,并可自行选择进食时间窗口(8:00-16:00、9:00-17:00或10:00-18:00),持续4个月。在禁食期间,受试者不得摄入含热量的食物或饮料(黑咖啡或无糖茶除外),并被鼓励在整个干预期间保持足够的饮水量。在研究过程中,研究人员分别在基线和干预结束时收集认知评估量表(MoCA和MMSE)评分、血清样本及粪便样本,并在每次随访时记录不良事件。
不良事件的初步评估由临床营养师进行,并根据情况建议是否继续干预。研究过程中,受试者报告的不良事件由研究人员实时记录并上报研究团队。在干预期间,研究团队要求对不良事件进行3日内的随访,以确认是否有所缓解;若症状未改善,则由临床专家组决定是否终止受试者的干预过程。
在TRF干预结束后,共9名受试者完成研究(2名男性,7名女性,年龄范围58至81岁),1名受试者因无法坚持TRF而退出。所有受试者均获得详细的研究方案说明,并签署书面知情同意书。本研究经北京大学深圳医院伦理委员会批准,伦理审批编号为No. 2023-108,并已在中国临床试验注册中心(ChiCTR)注册,注册号为ChiCTR2400092653。
Morris水迷宫实验
Morris水迷宫用于研究与空间学习和记忆相关的脑区功能评价,实验方法基于先前研究并进行了少许修改。实验装置为直径150 cm、高度35 cm的圆形水池,池内填充不透明水(23–25°C)。在水池东北象限中心放置一个直径4.5 cm、高14.5 cm的平台。水池被划分为四个等面积象限,每个象限均设有显著视觉标记。该实验包括两个阶段:初始空间训练和探测实验。实验第一天,所有小鼠被训练在平台上停留60 s,以识别隐藏平台的存在并熟悉实验环境。接下来的3天进行初始空间训练,以评估小鼠的空间学习能力。训练过程中,实验水池中加入食品级二氧化钛使水呈白色,平台隐藏在水面以下0.5-1.0 cm。每天小鼠从四个象限之一进入水池,并允许其自由游泳直至找到并爬上平台,若60 s内未能找到平台,则由实验人员引导至平台并停留30 s。每只小鼠每天进行四次训练,每次训练的起始位置不同。视频追踪系统记录逃避潜伏期(s)。在最后一次训练后24 h,进行探测实验以评估空间记忆保持能力。实验第四天,移除平台,每只小鼠在水池中自由游泳60 s。利用光电传感器记录小鼠在目标象限的停留时间及穿越平台位置的次数,并使用计算机化视频追踪系统(Super Maze软件,上海欣软信息科技有限公司,中国)进行数据分析。
Barnes迷宫实验
采用Barnes迷宫实验对空间学习和记忆能力进行检测,实验方法参考先前研究。该迷宫由一个直径92 cm的圆形平台组成,平台周围均匀分布20个孔,其中一个孔底部放置逃生箱,其余孔为空。实验为期5天,第1-4天进行训练,每天训练1次,使小鼠学习定位逃生箱。实验开始时,小鼠被放置于中心的黑色起始箱中5 s,随后起始箱移除,小鼠可自由探索迷宫。若小鼠在3 min内找到并进入逃生箱,则实验结束,否则3 min后实验自动终止。记录逃避潜伏期及初始潜伏期。第5天进行探测实验,移除逃生箱,将小鼠置于迷宫中央自由探索3 min。每次实验结束后,迷宫及逃生箱使用70%乙醇擦拭,并旋转迷宫以避免位置依赖效应。使用Smart 3.0追踪软件(Super Maze软件,上海欣软信息科技有限公司,中国)分析小鼠在目标象限的停留时间百分比。
实时荧光定量PCR(Real time-qPCR)
为了检测神经营养因子及炎症因子相关调控基因的mRNA表达水平,采用Trizol试剂(Invitrogen, Carlsbad, CA)从脑组织中提取总RNA,并按照制造商说明进行操作。从组织样本中提取的总RNA(5 μg)使用Prime Script TM RT Master Mix逆转录试剂盒(TaKaRa Prime Script RT Master Mix,大连,中国)在CFX96T实时荧光定量PCR系统(Bio-Rad, Hercules, CA)上进行逆转录反应。基因特异性引物详见表S3。Ct值归一化至内参基因GAPDH,并采用2-△△Ct方法计算相对基因表达水平。
Western blot检测
Western blot实验按照先前描述的方法进行。从4只小鼠/组的肝组织(30 mg)提取蛋白,使用冰冷的RIPA裂解液(P0013C,碧云天生物技术研究所,上海,中国),其中含1 mM PMSF及蛋白酶抑制剂(P1005,碧云天生物技术研究所,上海,中国)。蛋白样品经变性处理后在10% SDS-PAGE凝胶上进行电泳分离,并转移至0.45 μm PVDF膜。随后,使用5%脱脂奶粉在37°C封闭2 h,并在4°C孵育一抗过夜,包括FFAR3(1:1000, ER64606; HuaBio, 杭州,中国)、GAPDH(1:1000, Lot#5174S; Cell Signaling Technology, 波士顿,美国)、JNK(1:1000, sc-7345; Santa Cruz, 加利福尼亚,美国)、p-JNK(1:500, sc-6254; Santa Cruz, 加利福尼亚,美国)、NFκB(1:1000, sc-8008; Santa Cruz, 加利福尼亚,美国)、p-NFκB(1:1000, sc-136548; Santa Cruz, 加利福尼亚,美国)。次日,用TBST洗涤膜,并在室温下孵育相应二抗2 h。采用化学发光检测蛋白条带(Bio-Rad Laboratories, Hercules, CA),并使用Image J软件对Western blot结果进行定量分析。
免疫荧光染色
免疫荧光染色的步骤依据先前的研究进行。固定后的脑组织切片依次置于二甲苯和梯度乙醇中进行脱蜡和水化处理。随后,石蜡切片用3%过氧化氢孵育10 min,以阻断内源性过氧化物酶活性。接着,在室温下用山羊血清封闭组织切片20 min,然后于4°C下过夜孵育一抗,包括Aβ(1:1000,SIG-39320;Biolegend,Boston)、Iba-1(1:1000,Cat# ab178847;Abcam,Cambridge,UK)、BDNF(1:500,Cat# ab108319;Abcam,Cambridge,UK)。
第二天,PBS洗涤后,组织切片孵育荧光二抗(Cy3 标记的山羊抗兔 IgG (H+L),Servicebio,武汉,中国)20 min。随后,PBS清洗8次,并用含DAPI的封片剂封片。Aβ和Iba-1的共染色步骤为:第二天洗去一抗后,孵育相应的二抗,洗涤后,再孵育另一种一抗过夜。第三天,组织切片洗涤一抗后,再孵育相应的二抗,随后进行洗涤和封片。切片在荧光显微镜(Olympus,Tokyo,Japan)下观察并拍照,阳性区域的面积使用 Image J 进行测量。
短链脂肪酸(SCFAs)水平分析
粪便中的 SCFAs 水平采用气相色谱法(GC)测定,方法参考先前研究。简要而言,称取180-200 mg粪便样本,加入1 mL MilliQ纯水,在室温下振荡10 min。随后,加入150 μL 50% H₂SO₄(w/w),充分振荡混合。接着,加入1.6 mL乙醚,并在冰上孵育20 min。随后,样本以8000 rpm离心5 min(4°C),收集上清液,并使用0.2 µm滤膜(天津市亿肺实验设备有限公司,天津,中国)过滤。最终,上清液转移至干净的GC进样瓶,并以乙醚作为内标。
SCFAs的浓度通过GC-2014C气相色谱仪(岛津公司,京都,日本)进行分析,配备DB-FFAP毛细管柱(30 m × 0.25 µm × 0.25 mm)(Agilent Technologies,Wilmington,DE)。使用不同浓度的标准混合物(乙酸A116173、丙酸P110445、丁酸B11se0438)(阿拉丁生化科技有限公司,上海,中国)绘制标准曲线。峰面积由岛津GC solution软件进行积分计算。所有SCFAs的浓度均以µg/g(粪便)表示。
正电子发射断层扫描(PET)成像
[¹⁸F]-氟代丙酸([¹⁸F]-FPA)由前体物质乙基-2-溴丙酸酯合成,具体的[¹⁸F]放射标记和水解反应参照先前研究进行。总衰变校正后的放射化学产率超过40%,比活度约50 GBq/μM,放射化学纯度超过 98%。PET成像采用[¹⁸F]-FPA,并使用MadicLab Box071(山东麦迪克科技有限公司)设备进行。小鼠经静脉注射11.7 ± 5.2 MBq的[¹⁸F]-FPA,并在30 min后进行扫描。在PET成像前,动物先吸入2.5%异氟烷进行麻醉,并放置于加热的扫描床上,随后在整个扫描过程中保持1.5%-2%异氟烷麻醉。扫描时间为90 min。
PET数据采用二维子集期望最大化(2D-OSEM)算法进行重建,并校正时间及放射性衰变。所有后续PET图像的处理以及不同脑区的放射性分布分析均使用PMOD 4.3版本(PMOD Technologies LLC,瑞士)软件进行。
RNA测序分析
总RNA提取采用TriZol试剂(Invitrogen, Carlsbad, USA)按照生产商说明进行操作,并使用NanoDrop及Agilent 2100生物分析仪(Thermo Fisher Scientific, MA)对RNA质量进行检测和定量。RNA测序文库构建采用BGISEQ-500平台(华大基因,深圳,中国)。测序数据使用Trimmomatic v0.38(Bolger, Lohse, & Usadel, 2014)进行过滤和修剪,以获得高质量的清洁读段数据。清洁读段随后通过Hisat2 v2-2.1.0比对到小鼠(Mus musculus)基因组序列(ftp://ftp.ncbi.nlm.nih.gov/genomes/all/GCF/000/001/635/GCF_000001635.26_GRCm38.p6)。然后使用StringTie v1.3.4d对每个样本的读段进行转录组拼接,并与参考基因模型进行比对。使用StringTie-Merge程序合并35个转录本以获得共识转录本。对于未在小鼠编码序列(CDS)数据库中发现的转录本,进一步提取用于新基因预测。基因表达FPKM(Fragments Per Kilobase of transcript per Million mapped reads)值通过StringTie Merge程序计算。KEGG(京都基因与基因组百科全书)及Gene Ontology(基因本体论)富集分析使用WebGestalt 2019(WEB-based Gene Set Analysis Toolkit)进行,并利用Metascape(http://metascape.org/)进行可视化分析。
代谢组学分析
粪便样本采用超高效液相色谱-串联质谱(UPLC-MS/MS)系统(ACQUITY UPLC-Xevo TQ-S,Waters Corp.,Milford,MA)进行靶向定量代谢组学分析,涵盖肠道菌群来源及相关代谢物。简要而言,冻干粪便样本使用含内标的冰甲醇充分振荡,收集上清液。随后,加入冰冷50%甲醇溶液稀释样品,并于-20°C存放20 min后,以4000 g离心。收集上清液并与内标混合后密封,用于UPLC-MS/MS分析。
仪器参数设定如下:C18分析柱(2.1 × 100 mm,1.7 μM);柱温40°C;流动相A(含 0.1% 甲酸的水),流动相B(乙腈:异丙醇,70:30)。质量控制样本由混合样本组成,每间隔14个样本进行一次运行。UPLC-MS/MS 产生的原始数据使用MassLynx 软件(v4.1,Waters,Milford,MA)进行峰积分、校准和定量。共检测到174种代谢物,包括氨基酸、脂肪酸和胆汁酸(详见表 S2)。样本制备和代谢组学分析由上海麦特博生物科技有限公司(上海,中国)完成。
16S rRNA基因测序及宏基因组测序
采用16S rRNA基因测序及全基因组鸟枪法宏基因组测序分析粪便样本中的肠道菌群组成,该分析由华大基因(深圳国家基因库,深圳,中国)完成。简要而言,在无污染环境下收集粪便样本,并使用E.Z.N.A.粪便DNA提取试剂盒(Omega, Norcross, GA)按照生产商说明提取总DNA。选取细菌16S rRNA超可变V3-V4区域,采用MiSeq(Illumina, CA)平台进行双端300 bp扩增子测序,使用引物341_F(5'-CCTACGGGNGGCWGCAG-3')和802_R(5'-TACNVGGGTATCTAATCC-3')。文库构建参照已发表的方法9。原始读段进行合并、修剪,并去除嵌合序列,利用UNOISE在Search(v2.6.0)中实现零半径操作分类单元(zOTUs)分群。分类注释采用绿色基因(Greengenes)16S rRNA基因数据库(13.8 版本)。稀释OTU数据后,使用 PICRUSt(v1.1.3)预测功能基因,并按照KEGG不同层级进行功能注释,采用edgeR进行统计分析(FDR-p < 0.1 )。
宏基因组的分类学分析方法参照先前研究。清洁读段使用Kraken2 v2.1.2(k-mer 分析工具)比对至Kraken数据库(k2_pluspfp_20220908),生成分类学概况。Kraken2结果随后由Bracken v2.5处理,计算每个样本中相对丰度,即分配的读段数量相对于总清洁读段的比率。使用R语言包“vegan”计算α和β多样性,分别采用Shannon指数及Bray-Curtis不相似性进行评估,具体计算使用“diversity”及“vegdist”函数。功能注释使用HUMAnN3 v3.5进行每个样本的宏基因组功能分析。KEGG Orthology(KO)丰度通过HUMAnN3工具基于201901版官方数据库进行计算。16S rRNA基因测序及全基因组鸟枪法宏基因组测序均由华大基因有限公司(深圳,中国)完成。
数据分析
数据以均值 ± 标准误(SEM)表示,基于至少三次独立实验。组间差异的显著性采用双侧非配对Student’s t检验(两组间)、配对Student’s t检验(TRF临床干预前后比较)、单因素方差分析(ANOVA,三组或五组比较)或双因素方差分析(ANOVA,四组比较),并使用GraphPad Prism 8.0进行Tukey事后检验。显著性设定为p < 0.05,图中用*p < 0.05和**p < 0.01表示。数据分析采用盲法进行。
OMICs数据的统计分析使用 R 语言(4.1 版本)。转录组学数据采用基因集富集分析(GSEA)评估AD组和AD+TRF组之间的生物学功能差异(R语言包 ClusterProfile),显著性设定为p < 0.05。特别关注与AD病理生理密切相关的KEGG信号通路,包括神经营养因子信号通路、mTOR信号通路、凋亡、AMPK 及线粒体代谢(表S1)。最终选取381个基因,并进行偏最小二乘判别分析(PLSDA,R 语言包 mixOmics)。
采用Wilcoxon检验评估TRF对AD小鼠肠道微生物种类的影响。Chao1和ACE指数反映微生物群落丰富度,Shannon和Simpson指数反映群落多样性。FDR-p < 0.05视为显著性差异。最后,采用Fuzzy C-means聚类技术(R语言包 ClusterGVis)整合TRF诱导的生物功能参数变化、与AD病理生理相关的差异基因、微生物分类群及粪便代谢物。该方法是数据挖掘中的典型聚类算法,广泛用于高通量及高维数据的模式识别。
代码和数据可用性
数据存储:16S rRNA基因序列数据可在NCBI BioProject登录号 PRJNA1014591(https://www.ncbi.nlm.nih.gov/bioproject/PRJNA1014591)上获取。转录组和宏基因组序列数据已存放在基因组序列档案中,登录号为CRA022584 和CRA022537(https://bigd.big.ac.cn/gsa/browse/CRA022584和 https://bigd.big.ac.cn/gsa/browse/CRA022537。原始数据已存放在figshare数据集中,如下所示:https://doi.org/10.6084/m9.figshare.26129467.v1。更详细的数据信息可联系通讯作者获取。补充材料(文本、图、表、中文翻译版本或视频)也可从线上(http://www.imeta.science/)获取。
引文格式:
Yihang Zhao, Mengzhen Jia, Chen Ding, Bingkun Bao, Hangqi Li, Jiabin Ma, Weixuan Dong, et al. 2024. Time-restricted feeding mitigates Alzheimer's disease-associated cognitive impairments via a B. pseudolongum-propionic acid-FFAR3 axis. iMeta 4: e70006. https://doi.org/10.1002/imt2.70006.
作者简介

赵忆航(第一作者)
● 西北农林科技大学食品科学与工程学院在读博士研究生。
● 研究方向为膳食结构调控阿尔兹海默病的机制探索,以第一作者在iMeta、Free Radical Biology and Medicine、Journal of Nutritional Biochemistry期刊发表SCI论文3篇。

贾梦真(第一作者)
● 西北农林科技大学食品科学与工程学院在读博士研究生。
● 研究方向为食品营养与健康,以共同第一作者在iMeta、Journal of Neuroinflammation发表SCI论文2篇。

刘志刚(通讯作者)
● 西北农林科技大学食品科学与工程学院教授,博士生导师。
● 主要研究内容包括:重点围绕阿尔兹海默症、孤独症、焦虑抑郁等特殊人群脑健康问题开展肠脑互作机制阐释、特色膳食模式改善脑健康作用及机理研究、食品营养功能组分发掘与功效评价等。成果在Cell Metabolism、Nature Communications、Redox Biology等领域内的高水平刊物上发表论文110余篇,总引用超过5900余次,H-index 49。主、参编著作5部,申请或授权专利8项。主、参编专著4部。担任Food & Humanity杂志主编。

刘学波(通讯作者)
● 西北农林科技大学食品科学与工程学院教授,博士生导师,校长助理。
● 研究方向为膳食因子调节糖脂代谢及脑认知功能的营养功能性评价、膳食模式的营养学干预策略和新型健康食品研发。入选国家级人才,教育部“新世纪优秀人才”支持计划,科技部“中青年科技创新领军人才”计划,荣获2015年度中国食品科学技术学会科技创新奖-杰出青年奖,享受国务院政府特殊津贴。成果在Cell Metabolism、Nature Communications、Gut Microbes等专业期刊上发表论文200余篇。任《中国食品学报》、《中国农业科学》、《食品科学》、Food Science and Human Wellness编委。

施琳(通讯作者)
● 陕西师范大学食品工程与营养科学学院副研究员,硕士生导师。
● 主要研究方向为膳食营养与肠道菌群互作调控及其健康改善作用机制研究。在Nature Communications、Microbiome、Journal of Internal Medicine、Journal of Agricultural and Food Chemistry、Redox biology 和American Journal of Clinical Nutrition等具有国际影响力的食品营养学、生物学和医学领域期刊发表论文50余篇。主持参与国家级、省部级科学研究项目10余项,联合主持瑞典FORMAS基金一项。

胡俊(通讯作者)
● 北京大学深圳医院神经内科主任、教研室主任,眩晕诊疗中心主任。
● 主要研究方向为阿尔兹海默病、脑血管病、中枢神经系统感染、脱髓鞘疾病、癫痫等疾病的诊治。在Journal of Neuroinflammation、Annals of Clinical and Translational Neurology、Frontiers in Neurology等权威学术期刊发表SCI论文46篇,累计影响因子150分。
更多推荐
(▼ 点击跳转)
iMeta | 引用16000+,海普洛斯陈实富发布新版fastp,更快更好地处理FASTQ数据
iMeta | 兰大张东组:使用PhyloSuite进行分子系统发育及系统发育树的统计分析
iMeta | 唐海宝/张兴坦-用于比较基因组学分析的多功能分析套件JCVI

1卷1期

1卷2期

1卷3期

1卷4期

2卷1期

2卷2期

2卷3期

2卷4期

3卷1期

3卷2期

3卷3期

3卷4期

3卷5期

3卷6期

1卷1期

1卷2期
期刊简介
“iMeta” 是由威立、宏科学和本领域数千名华人科学家合作出版的开放获取期刊,主编由中科院微生物所刘双江研究员和荷兰格罗宁根大学傅静远教授担任。目的是发表所有领域高影响力的研究、方法和综述,重点关注微生物组、生物信息、大数据和多组学等前沿交叉学科。目标是发表前10%(IF > 20)的高影响力论文。期刊特色包括中英双语图文、双语视频、可重复分析、图片打磨、60万用户的社交媒体宣传等。2022年2月正式创刊!相继被Google Scholar、PubMed、SCIE、ESI、DOAJ、Scopus等数据库收录!2024年6月获得首个影响因子23.8,位列全球SCI期刊前千分之五(107/21848),微生物学科2/161,仅低于Nature Reviews,学科研究类期刊全球第一,中国大陆11/514!
“iMetaOmics” 是“iMeta” 子刊,主编由中国科学院北京生命科学研究院赵方庆研究员和香港中文大学于君教授担任,是定位IF>10的高水平综合期刊,欢迎投稿!
iMeta主页:
http://
www.imeta.science
姊妹刊iMetaOmics主页:
http://www.imeta.science/imetaomics/
出版社iMeta主页:
https://onlinelibrary.wiley.com/journal/2770596x
出版社iMetaOmics主页:
https://onlinelibrary.wiley.com/journal/29969514
iMeta投稿:
https://wiley.atyponrex.com/journal/IMT2
iMetaOmics投稿:
https://wiley.atyponrex.com/journal/IMO2
邮箱:
office@imeta.science


















 被折叠的 条评论
为什么被折叠?
被折叠的 条评论
为什么被折叠?
 到【灌水乐园】发言
到【灌水乐园】发言









